
Abstract
Background:
The high constitutive, or ligand-independent, activity of the thyrotropin receptor (TSHR) is of clinical importance in some thyroid conditions, particularly well-differentiated thyroid carcinoma remnants following incomplete ablative therapy (surgery and radioiodine). Under these conditions, even total suppression of TSH by thyroid hormone administration does not fully reduce TSHR activity, a driver of thyrocyte growth.
Methods:
CS-17 is a murine monoclonal antibody that has inverse agonist activity in that it suppresses TSHR constitutive activity. This study crystallized the CS-17 Fab and determined its atomic structure at a resolution of 3.4 Å.
Results:
In silico docking of this structure to that of the TSHR extracellular domain was accomplished by targeting to TSHR residue tyrosine 195 (Y195) known to contribute to the CS-17 epitope. High affinity interaction between these two molecules, primarily by the CS-17 immunoglobulin heavy chain, was validated by energetic analysis (KD of 8.7 × 10–11 M), as well as by previously obtained data on a number of individual TSHR amino acids in three regions whose mutagenesis reduced CS-17 binding as detected by flow cytometry.
Conclusions:
Structural insight at atomic resolution of a TSHR antibody with inverse agonist activity opens the way for the development of a molecule with therapeutic potential, particularly in thyroid carcinoma. For this purpose, CS-17 will require “humanization” by substitution of its constant region (Fc component). In addition, with its epitope defined, the CS-17 affinity can be increased further by mutagenesis of selected amino acids in its heavy- and light-chain complementarity determining regions.
Introduction
T
High TSHR constitutive activity is of clinical importance in thyroid carcinoma, the great majority of which are well differentiated (7). It has been recognized for >60 years that thyroid hormone suppression of TSH secretion can reduce the post-ablative (surgery or radioiodine) recurrence of thyroid carcinoma both locally and distantly (8,9). Because moderate or complete suppression of TSH secretion requires supraphysiological doses of thyroxine (T4), awareness in recent years of deleterious consequences has led to guidelines restricting this approach to incomplete removal of thyroid carcinoma, a common outcome (10). However, even complete TSH suppression does not reduce TSHR constitutive activity—hence the current interest in the use of TSHR inverse agonists to suppress this activity in concert with physiological T4 replacement following thyroid ablation.
Previously, a mouse monoclonal antibody, CS-17, was generated with inverse agonist activity (11). The present study reports the crystal structure of the CS-17 Fab molecule. In silico docking of this structure with that of the TSHR ECD provides insight into potential mechanisms by which CS-17 constrains the high level of TSHR constitutive activity.
Methods
CS-17 Fab purification
Monoclonal antibody CS-17, an IgG2a, is one of a panel of TSHR monoclonal antibodies (mAb) generated in the authors' laboratory by immunizing BALB/c mice by intramuscular injection of an adenovirus expressing the human TSHR A-subunit, as reported previously (11). Murine SP-2/0 hybridoma cells were transferred to QED Biosciences (San Diego, CA) for ascite generation in SCID mice lacking endogenous mouse immunoglobulin G (IgG). CS-17 IgG was extracted from the ascites using Protein G Hi-Trap columns (GE Healthcare, Piscataway NJ) following which the IgG was digested and Fab purified using the ImmunoPure Fab Preparation kit (Pierce, Rockford, IL). The nucleotide and amino acid sequences of the heavy and light chains of CS-17 Fab have been submitted to GeneBank (accession numbers MH036357 for H chain and MH036358 for the light chain). CS-17 Fab amino acid residues are shown in Figure 1.

Primary amino acid sequence of the CS-17 heavy (H) chain and light (L) chain variable regions. The complementarity determining regions (CDRs) are in bold. FR, framework region.
CS-17 Fab crystals and X-ray diffraction
CS-17 Fab, in 20 mM of Tris, pH 7.4, 150 mM of NaCl, and at a concentration of 14 mg/mL, was crystallized using the hanging drop vapor diffusion method. An equal volume of well solution (0.1 M of HEPES, pH 7.5, 0.3 M of ammonium citrate, and 20% PEG 3350) was added to CS-17 Fab and equilibrated over well solution at 18°C. Crystals were cryoprotected by soaking in well solution containing 35% PEG 3350 followed by flash-freezing in liquid nitrogen. Initially, low-resolution diffraction data were collected in-house using a Rigaku Micromax 007HF X-ray diffractometer with an R-Axis IV++ detector and processed/scaled with XDS/XSCALE (12). Then, crystals were also sent to the Stanford Synchrotron Radiation Lightsource (SSRL) facility to obtain high-resolution data. The crystals diffracted to a maximum resolution of 3.4 Å. The structure was solved by molecular replacement with Phaser (13) using the heavy and light chains of the Fab for monoclonal OKT3 (PDB ID: 1SY6) (14) as a search model. The crystallographic model was built using Coot (15) and refined using Phenix (16) and BUSTER (17).
Docking of the CS-17 atomic structure to the TSHR ECD
ZDOCK (18) was used for in silico docking of CS-17 to the TSHR ECD. The structure of the latter was based on the known structure of TSHR residues 22–260 (largely the LRD; structure database 3G04 and 2XWT) (19,20), together with TSHR residues 261–410 modeled on the crystal structure of the FSHR hinge region (structure data base 4AY9) (21), as described by Kreuchwig et al. (22,23) (
CS-17 binding energetics
The energetics of the CS-17/TSHR ECD complex were analyzed by PRODIGY (24,25).
Results
Crystal structure of the CS-17 Fab
Crystals of purified CS-17 Fab (Fig. 2) were subjected to X-ray diffraction analysis. Despite analysis of multiple sets of CS-17 crystals generated over a period of nearly two years, and repeated efforts to collect high-resolution data at the SSRL facility, only moderate resolution of its atomic structure (3.4 Å) was obtained. Nonetheless, the overall structure for CS-17 Fab was determined using molecular replacement. The backbone conformation of all six CDR loops was determined unambiguously. Due to resolution limit, neither solvents nor side chains could be determined with greater accuracy. Side chains were built using COOT (26) and optimized by molecular simulations using Desmond (Schrodinger, Inc., San Diego, CA). Data collection and refinement statistics are shown in Table 1. The final structure of Fab CS-17 is shown in Figure 3.

Selection of CS-17 crystals, from which some were subjected to Synchrotron X-ray diffraction analysis. Crystal birefringence under polarizing light is shown in the right panel.
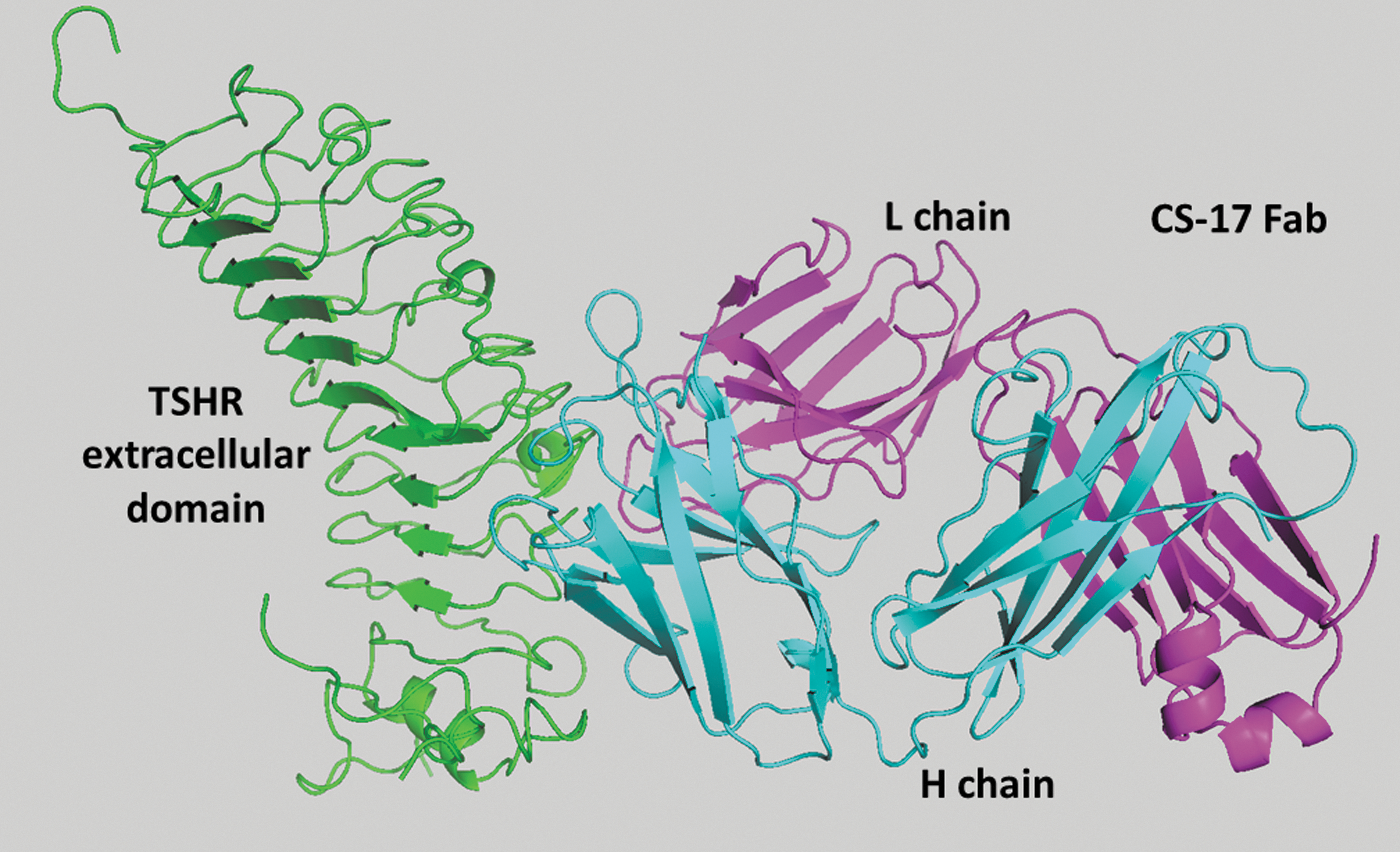
Molecular structure of the CS-17 Fab in complex with the thyrotropin receptor (TSHR) extracellular domain (ECD). The TSHR is shown in green; the L chain of CS-17 Fab is in magenta and the H chain in blue. The ECD structure was based on the crystal structure of TSHR residues 22–260 (19,20), together with TSHR residues 261–410 modeled on the crystal structure of the FSHR hinge region (21), as described by Kreuchwig et al. (22,23) (see Methods).
Values in parentheses are for highest-resolution shell.
TSHR, thyrotropin receptor; rms, root mean square.
Docking of the CS-17 Fab to the TSHR ECD
To facilitate in silico docking, extensive studies had previously provided information regarding the CS-17 epitope. First, the immunogen for generating CS-17 was the recombinant human TSHR A-subunit (amino acids 22–289; residues 1–21 being the signal peptide) (27,28). Second, flow cytometric studies using chimeric receptors involving substitution of human TSHR segments with the corresponding region of the closely related rat LHR localized a major portion of the CS-17 epitope to be downstream of TSHR residue 260 (11). Taken together, these observations localized a significant portion of the CS-17 epitope to lie between residues 261 and 289. Third, based on the observation that CS-17 recognized the human but not the porcine TSH holoreceptor, individual substitution of six non-homologous human TSHR amino acid residues with its porcine counterpart within this region identified human TSHR Y195 as having a significant influence on CS-17 binding, while that of a control TSHR antibody to a distant location was unaffected (29) (Fig. 4). Consequently, in silico docking of the atomic structure of the CS-17 Fab with the TSHR ECD was performed, selecting TSHR Y195 as a target.
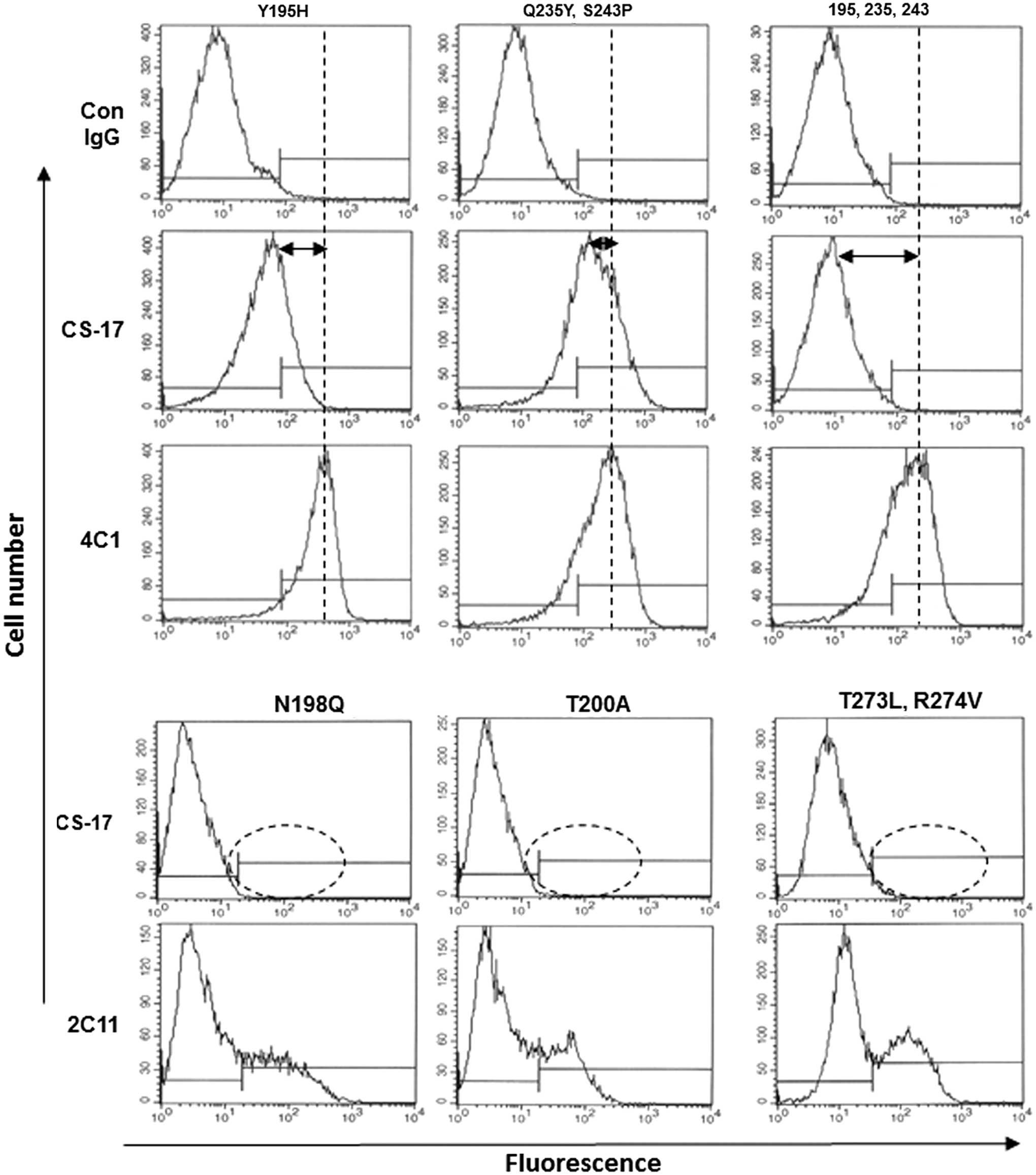
TSHR ECD mutations previously observed to reduce CS-17 binding as detected by flow cytometry. TSH holoreceptors with Y195H and Q235/S243P (dual mutation) and Y195H/Q235/S243P (triple mutation) were stably (and clonally) expressed on the surface of intact Chinese hamster ovary (CHO) cells (29). Included as negative control for CS-17 recognition is normal mouse immunoglobulin G (IgG; Con) and mouse monoclonal antibody (mAb) 4C1, whose epitope is distant from CS-17, as a positive control. The horizontal arrows depict the degree of reduction in CS-17 binding compared to binding of 4C1; the triple mutation totally abrogates CS-17 binding whereas 4C1 recognition is unaffected. For the N198Q, T200A, and T273/R274V (dual) mutations, TSHR holoreceptors were transiently (non-clonally) expressed, and the TSHR null cells in the pool serve as negative controls (30). The dotted ovals depict the loss of CS-17 binding relative to that for another control TSHR antibody, 2C11. The panels in this figure were selected from an extensive series of mutations, the majority of which had no effect on CS-17 binding and are reproduced from the following manuscripts with permission from the Oxford University Press: Chen CR et al., Identification of key amino acid residues in a thyrotropin receptor monoclonal antibody epitope provides insight into its inverse agonist and antagonist properties. Endocrinology 2008;149(7):3427–3434; and Chen CR et al. A monoclonal antibody with thyrotropin (TSH) receptor inverse agonist and TSH antagonist activities binds to the receptor hinge region as well as to the leucine-rich domain. Endocrinology 2009;150(7):3401–3408.
Of the top 10 predicted models, cluster #5 of the CS-17 Fab TSHR ECD complex (Fig. 3) was validated on the basis of energetic analysis taken together with the experimental data described above. The interface of this complex was predicted by PRODIGY (18,19) to involve a very high affinity interaction, with a DeltaG (kcal/mol) of −13.7 and a KD of 8.7 × 10–11 M. Individual TSHR amino acids whose experimental mutagenesis reduced CS-17 binding were located in three regions: (1) Besides a Y195H mutation, mutation of closely related residues, N198Q and T200A, also had a major effect on CS-17 binding to the TSH holoreceptor expressed on the surface of Chinese hamster ovary (CHO) cells (30) (Fig. 4). These functional effects are supported by the in silico docking analysis indicating contact of TSHR Y195 with CS-17 L chain H91, TSHR N198 with CS-17 H chain V105, and TSHR K201 with CS-17 H chain residues Y101, E99, and S31 (Fig. 5A and Table 2). Of note, all CS-17 contact residues are within the Fab complementarity determining regions. (2) Docking analysis predicted interaction between TSHR H271 and CS-17 H chain Y60 (Fig. 5B and Table 2). This information is consistent with the dual mutation of adjacent TSHR amino acid residues T273L and R274V completely abrogating CS-17 binding (30) (Fig. 4). (3) A third region in the docking analysis involved TSHR K244 and G245 interacting with L chain S92 and H chain N59, respectively (Fig. 5C and Table 2). These data are consistent with the TSHR dual Q235Y, S243P mutations on CS-17 binding (29). Though small on their own, these mutations together with the Y195H mutation synergistically abrogated CS-17 binding (Fig. 4). The segment of the TSHR between Q235 and S243 is a single strand (shown in yellow). Proline residues introduce a fixed bend in the secondary structure of a protein. Therefore, mutation of S243 to a proline immediately adjacent to K244 and G245, as well as the Q235Y mutation, will alter the position of K244 and G245 in this strand.
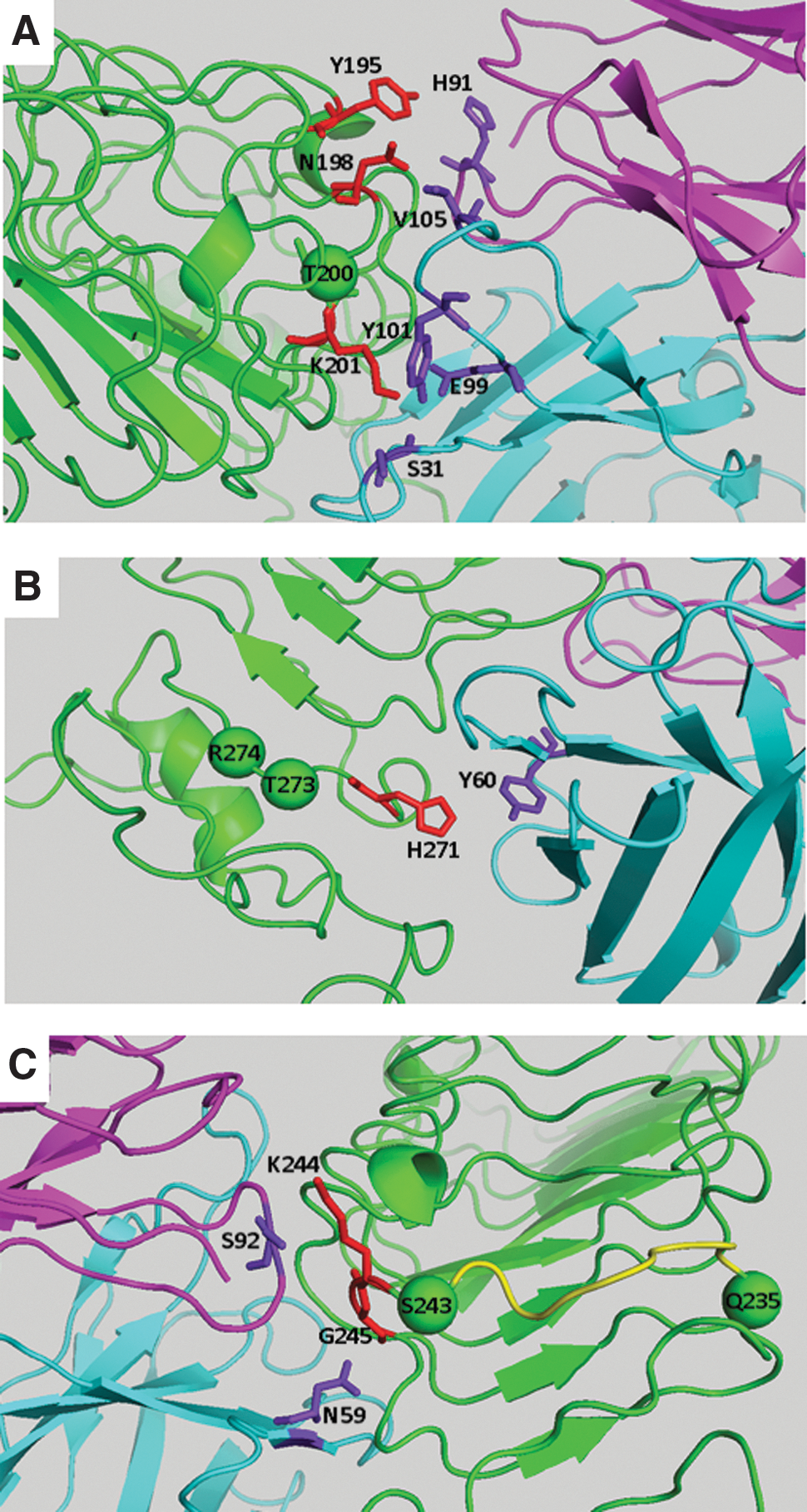
Detail of CS-17 binding to TSHR ECD in segments in which mutagenesis of the latter reduced this interaction (Fig. 4). The numerous other interfaces between these molecules are listed in Table 1. (
Asterisk indicates a salt bridge.
Putative contacts between CS-17 heavy (H) and light (L) chains and the TSHR ECD were determined using CONTACT (CCP4) (51).
Discussion
GPCR are the target of many therapeutic agents that function as antagonists or inverse agonists, some with both properties (reviewed in Bond and Ijzerman) (31). Although the great majority of inverse agonists are small molecules isolated by screening synthetic molecule libraries, mAb also provide therapeutic opportunities—reviewed in Webb et al. (32), Mujic-Delic et al. (33), and Jacobson (34). Unlike small molecules, very few mAb with inverse agonist activity have been generated, such as to the beta2-adrenergic (35), M2 muscarinic acetylcholine (36), melanocortin-4 (37), and CXCR4 chemokine (38) receptors.
An inverse agonist to the TSHR would be of value in a number of clinical conditions, particularly thyroid cancer in which TSHR constitutive activity can drive thyrocyte growth even when its natural ligand, TSH, is totally suppressed by thyroid hormone administration (see Introduction). Both small molecule (39,40) and mAb (11,41) inverse agonists for the TSHR have been generated or isolated. Pending further structural refinement, the former are either of relatively low affinity or have only partial specificity when tested against the closely related LHR and FSHR. Because small molecule allosteric modulators permeate the plasma membrane and insert into a pocket in the hepta-helical transmembrane domain, there is also the possibility of unanticipated clinical side effects, given the vast number of known and orphan receptors in the GPCR family. On the other hand, the advantages of an inverse agonist TSHR mAb are its high affinity and specificity for a very large receptor ECD absent in most GPCR.
Solving the atomic structure of the CS-17 Fab was not straightforward, with the highest resolution being 3.4°Å, despite three submissions of multiple crystals for synchrotron analysis. Nevertheless, this resolution was sufficient for in silico docking with the TSHR ECD. Based on previous mutagenesis locating TSHR Y195 as an important contributor to the CS-17 epitope, the CS-17 Fab structure to the TSHR ECD (22,23) was targeted using ZDOCK (see Methods). The validity of the interaction between CS-17 and the TSHR ECD was supported by previously obtained experimental data (see Results), in particular partial or total inhibition of CS-17 by mutation of TSHR residues (in addition to Y195) N198, T200, and the dual mutations of T273, R274, Q235, and S243. Further support was provided by the very high calculated affinity of the in silico interaction, comparable to the functional effect of CS-17 in the nanomolar range in suppressing constitutive activity of the recombinant TSHR expressed on CHO cell monolayers (11).
Characterizing the CS-17 epitope provides insight into how this antibody exerts its inverse agonist activity. The TSHR hinge region is the region involved in coupling TSH (the physiological ligand) binding to the transmembrane domain. Two critical residues in the TSHR hinge involved in receptor activation are Y385 (42,43) and S281 (44,45), reviewed in Kleinau et al. (46). Upon TSH binding, upward movement of TSHR Y385 (equivalent residue Y335 in the FSHR) into a ligand pocket is associated with flexion in the hinge region leading to signal transduction (21,47) (Fig. 6). Mutation of S281 leads to near full receptor activation, comparable to that effected by TSH and much greater than ligand independent constitutive activity. In contrast to ligand activation, the TSHR ECD (5,6), in particular the hinge region (48), partially suppresses constitutive activity. Inverse agonist CS-17 further accentuates this constraint. Based on the present data, it is proposed that CS-17 binding to the TSHR ECD flexes the hinge region in the opposite direction to the natural ligand, increasing the ectodomain suppression of constitutive activity (Fig. 6). The majority of the CS-17 contact residues are in the rigid TSHR leucine rich repeat domain, but CS-17 H chain Y60 also contacts TSHR amino acid residue H271 in the flexible hinge region, thereby clamping this hinge configuration. Another possible mechanism for CS-17 inverse agonist activity may relate to its potential impact on the association or dissociation of TSHR multimers on the cell surface. Indeed, there is evidence (though controversial) that activation by agonists (TSH and thyroid stimulating antibodies) influences the extent of TSHR multimerization, reviewed in Kleinau et al. (49). In addition, TSHR ligand independent constitutive activity is also related to negative cooperativity, the latter a function of receptor multimerization (50). Future studies will be required to determine whether CS-17 influences TSHR multimerization, such as by using bioluminescence resonance energy transfer.

Proposed mechanism of CS-17 inverse agonist activity. The TSHR ECD comprises a leucine-rich repeat domain (LRD) whose crystal structure is known (13,14) and a hinge region modeled on the closely related FSHR (21). The membrane spanning domain, modeled on rhodopsin, and the TSHR holoreceptor as a unit is from Kreuchwig et al. (22,23). Upward movement of TSHR residue Y385 following ligand binding flexes the hinge region (arrow) and transduces receptor signaling via the transmembrane domain (TMD) (21,47). Residue S281 is a key interface with the TMD and its mutation to numerous amino acids leads to receptor activation comparable to TSH (44,45). The major footprint of CS-17 is on the TSHR LRD, but it also binds to H271 in the hinge regions. It is proposed that CS-17's inverse agonist activity occurs by flexing the hinge region in the direction opposite to the ligand TSH (arrow).
In summary, solving the atomic structure of a TSHR antibody with inverse agonist activity opens the way to development of a molecule with therapeutic potential in certain thyroid conditions, particularly well-differentiated thyroid carcinoma metastases remaining after conventional ablative therapy. For this purpose, CS-17 will require “humanization” by substitution of its Fc component. In addition, with its epitope defined, the CS-17 affinity can be increased further by mutagenesis of selected amino acids in its H and L chain complementarity determining regions.
Footnotes
Acknowledgments
This work was supported by National Institutes of Health Grant DK19289 (B.R. until 2016), DK54684 (S.M.M.), and the Molecular Therapeutics Core Facility, Cedars- Sinai Research Institute. A pdb file for CS-17 docked to the TSHR ECD can be sent upon request.
Author Disclosure Statement
No competing financial interests exist.