
Abstract
Background:
c-Myc is overexpressed in different types of cancer, including thyroid cancer, and has been considered undruggable. There is evidence showing that MLN8237, a type of aurora A kinase (AURKA) inhibitor, destabilizes c-Myc proteins in liver cancer cells through disruption of the c-Myc/AURKA complex. However, the role of MLN8237 in thyroid cancer remains largely unclear. The aims of this study were to test the therapeutic potential of MLN8237 in thyroid cancer, and to analyze determinant factors affecting the response of thyroid cancer cells to MLN8237 and clarify the corresponding mechanism.
Methods:
The phenotypic effects of MLN8237 in thyroid cancer cells were evaluated through a series of in vitro and in vivo experiments, and the mechanism of c-Myc affecting MLN8237 response were explored using Western blot, ubiquitination, and cycloheximide chase assays.
Results:
The data show that the levels of c-Myc protein were strongly associated with MLN8237 cellular response in thyroid cancer cells. Only the cells with high c-Myc expression exhibited growth inhibition upon MLN8237 treatment. However, MLN8237 barely affected the growth of those with low c-Myc expression. Mechanistically, MLN8237 dramatically promoted proteasomal degradation of c-Myc proteins through disruption of the c-Myc/AURKA complex in the cells with high c-Myc expression. A similar antitumor activity of MLN8237 was also found in xenograft tumor models.
Conclusions:
The data demonstrate that c-Myc is a major determinant for MLN8237 responsiveness in thyroid cancer cells. Thus, indirectly targeting c-Myc by MLN8237 may be an effective strategy for thyroid cancer overexpressing c-Myc.
Introduction
T
The basic helix-loop-zipper (bHLHZ) protein c-Myc belongs to the Myc family, which includes c-Myc, N-Myc, and L-Myc (8,9), and has been demonstrated to be frequently elevated in a variety of human cancers (10). Similarly, increased expression of c-Myc has also been found in thyroid cancer, and it is associated with dedifferentiation and poor prognosis (11,12). Given that c-Myc is involved in regulating cell cycle, cell growth, protein synthesis, differentiation, and transformation (13), great efforts have focused on the development of new agents designed to target c-Myc for cancer therapy. Unfortunately, c-Myc does not harbor any cavities into which small molecules can easily bind (14,15). Therefore, it is a huge challenge to target c-Myc directly.
Aurora A kinase (thereafter AURKA), a member of the aurora kinase family, is a serine threonine kinase that is a key mitotic regulator (16). It is highly expressed in different types of cancer and plays a critical role in tumorigenesis (17,18). A growing number of AURKA inhibitors have been developed in recent years. Among them, MLN8237, a type of conformation-changing AURKA inhibitor, shows remarkable anticancer activities through selectively inhibiting AURKA in preclinical studies and clinical trials (19). A previous study reported that AURKA can form a complex with and stabilizes N-Myc through protecting it from proteasomal degradation in neuroblastoma, and MLN8237 disrupts the complex to promote its degradation (14). Like N-Myc, c-Myc also forms a stable complex with AURKA in NRAS-driven TP53-altered liver cancer cells, and MLN8237 has been shown to promote proteasomal degradation of c-Myc, thereby inhibiting cancer cell growth (20).
Based on the above observations, an effective strategy is suggested to treat c-Myc overexpressing thyroid cancers by indirectly targeting c-Myc. Indeed, a previous study has demonstrated that an indirect method aimed at suppressing the transcriptional activity of c-Myc with JQ1 is efficient in ATC cells (21). The present study attempted to test the therapeutic potential of MLN8237 in thyroid cancer cells and to explore whether c-Myc is a major determinant for the response of thyroid cancer cells to MLN8237 through a series of in vitro and in vivo experiments.
Methods
Human thyroid cancer cell lines and drug treatments
Human thyroid cancer cell lines K1, FTC133, 8305C, 8505C, BCPAP, and TPC-1 were obtained from Dr. Haixia Guan (The First Affiliated Hospital of China Medical University, Shenyang, P.R. China). All cell lines used in this study were authenticated by short tandem repeat analysis at Genesky Co. Ltd, (Shanghai, P.R. China), and the results were consistent with a previous study (22) and the COSMIC database (
Cell proliferation assay
Cells (2 × 103/well) were seeded in 96-well plates and cultured with various concentrations of MLN8237 for 72 h. The MTT assay was then performed to evaluate cell proliferation, as previously described (23). Three triplicates were done to determine each data point.
Colony formation assay
Cells were seeded in 24-well plates at a density of 6000–8000 cells/mL, and treated with the indicated concentration of MLN8237 or DMSO for five days. Cells were then fixed with methanol and stained with crystal violet. Each experiment was performed in triplicate.
Apoptosis and cell cycle assays
For the apoptosis assay, cells were treated with 250 nM MLN8237 or DMSO for 24 h, harvested, washed with phosphate-buffered saline (PBS), and stained with Annexin V-FLUOS Staining Kit (Roche Applied Science, Penzberg, Germany) according to the manufacturer's protocol. The apoptotic cells were analyzed by a Flow Cytometer (BD Biosciences, Franklin Lakes, NJ). Each experiment was performed in triplicate. For cell cycle assay, 8505C and K1 cells in the logarithmic growth phase were serum starved for 12 h, and treated with 250 nM MLN8237 or DMSO for the indicated time. Next, cells were harvested and fixed in ice-cold 70% ethanol for at least 30 min. After washing twice with PBS, propidium iodide (PI) solution (50 μg/mL PI, 50 μg/mL RNase A, 0.1% Triton-X, 0.1 mM EDTA) was added to stain the cells. Cells were then subjected to flow cytometry analysis to determine cell cycle distributions.
Short interfering RNAs transfection
Oligonucleotides of short interfering RNAs (siRNAs) targeting AURKA (si-AURKA #1 and si-AURKA #2) and control siRNA (si-NC) were obtained from Ruibobio (Guangzhou, China), and the sequences are presented in Supplementary Table S2. One day prior to transfection, 2 × 105 cells were plated on a six-well plate to achieve 50% confluence overnight. Using X-tremeGENE siRNA Transfection Reagent (Roche Diagnostics, Mannheim, Germany), cells were transfected with the above siRNAs at a final concentration of 1 μg/mL.
RNA extraction and quantitative reverse transcription polymerase chain reaction
Total RNA was isolated from cell lines using TRIZOL reagent (Takara, Inc., Dalian, P.R. China), and was converted to cDNA using PrimeScript RT reagent Kit (Takara, Inc.) according to the manufacturer's protocol. Quantitative reverse transcription polymerase chain reaction (qRT-PCR) was carried out on a CFX96 Thermal Cycler Dice™ real-time PCR system (Bio-Rad Laboratories, Hercules, CA) using SYBR Premix Ex Taq™ (Takara, Inc.). 18S rRNA was used as a reference gene to normalize mRNA levels of c-Myc. Three triplicates were done for each sample. The sequences of primers are listed in Supplementary Table S3.
Western blot analysis
Cells were harvested and washed with PBS, and then lysed in prechilled RIPA buffer containing protease inhibitors. Xenograft tumor tissues were homogenized in TissueLyser LT (Qiagen, Hilden, Germany) with RIPA buffer containing protease inhibitors. Equal amounts of protein lysates were separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis and transferred to polyvinylidene fluoride membranes (Roche Diagnostics). The membranes were then incubated overnight with the following primary antibodies: anti-c-Myc (Santa Cruz Biotechnology, Inc., Santa Cruz, CA), anti-AURKA (Bio-Rad Laboratories), anti-phospho-AURKA (Thr288; Cell Signaling Technology, Danvers, MA), and anti-GAPDH (Bioworld, Inc., Minneapolis, MN). This was followed by incubation with their respective horseradish peroxidase–conjugated secondary antibodies from ZSGB-BIO, and immunoblotting signals were visualized using the Western Bright ECL detection system (Advansta, Inc., Menlo Park, CA).
Co-immunoprecipitation
Cells were lysed in prechilled RIPA buffer containing protease inhibitors. The amount of proteins was adjusted to equal incorporation before the immunoprecipitaion assays. Protein extracts were first incubated with antibodies against c-Myc (Santa Cruz Biotechnology, Inc.), AURKA (Santa Cruz Biotechnology, Inc.), or immunoglobulin G at 4°C for 3 h, and then incubated with protein A/G-agarose beads (Santa Cruz Biotechnology, Inc.) at 4°C overnight. Immunoprecipitates were washed with RIPA buffer, and further analyzed by Western blot analysis.
In vitro protein ubiquitination assay
Cells were lysed with ice-cold RIPA buffer. After centrifuging, the supernatant was incubated with 10 μL anti-c-Myc antibody (Santa Cruz Biotechnology, Inc.) and Protein A/G PLUS-Agarose (Santa Cruz Biotechnology, Inc.). Eluted proteins were immunoblotted with an anti-ubiquitin antibody (Abcam, Cambridge, MA).
CHX chase assay
To assess protein stability, cells were treated with 200 μg/mL CHX (MP Biomedicals) to stop de novo protein synthesis following MLN8237 treatment for 24 h. At the indicated time points, cell lysates were harvested and then subjected to immunoblotting.
Animal studies
Four- to five-week-old female athymic nude mice were purchased from SLAC Laboratory Animal Co., Ltd. (Shanghai, P.R. China), and housed in a specific pathogen-free environment. Cells (8505C [3 × 106] and K1 [5 × 106]) cells were injected subcutaneously into the flanks of nude mice. When tumors grew to around 5 mm in diameter, mice were grouped into two groups (five mice per group), and MLN8237 (30 mg/kg) or vehicle was administered by oral gavage at a dose volume of 0.2 mL/20 g body weight in 10% 2-hydroxypropyl-β-cyclodextrin/1% sodium bicarbonate for 14 consecutive days. Tumor volumes were measured by a Vernier caliper every other day and calculated by the following formula: tumor volume (mm3) = l × w 2 × 0.5, where l is the length and w is the width. All mice were killed 2 h after the last treatment. Tumors were then harvested and weighted. All animal experiments were conducted and approved by the Laboratory Animal Center of Xi'an Jiaotong University.
Xenograft tumor tissues were fixed in 15% formalin for 48 h, and then embedded in paraffin, sectioned at 5 μm, deparaffinized, and rehydrated. Immunohistochemistry (IHC) was then performed to evaluate the levels of Ki-67 in these tumors. Next, the TUNEL assay was used to assess cell apoptosis in tumor tissues by using the TUNEL Andy Fluor™ 488 Apoptosis Detection Kit (GeneCopoeia, Inc., Rockville, MD) according to the manufacturer's protocol.
Statistical analysis
Data were compared using the t-test (SPSS Statistics for Windows v16.0, Chicago, IL). All values are expressed as the mean ± standard deviation. A p-value of <0.05 was considered to be statistically significant. Unless indicated, the data shown in the figures are representative examples.
Results
Response of thyroid cancer cells to MLN8237 correlates with c-Myc levels
First, the levels of c-Myc, phosphorylated AURKA (p-AURKA at Thr288), and AURKA were analyzed in six human thyroid cancer cell lines by Western blot analysis. The results showed that c-Myc levels were positively associated with the levels of p-AURKA (Thr288) and AURKA in these cell lines (Fig. 1A). Next, to determine in vitro growth response of these cell lines to the AURKA inhibitor MLN8237, these cells were treated with increasing concentrations of MLN8237 from 6.25 to 800 nM for 72 h to calculate IC50 values. Surprisingly, it was found that MLN8237 caused distinct growth responses among these cell lines (Fig. 1B). MLN8237 cellular responsiveness was strongly correlated with c-Myc levels. The c-Myc overexpressing cell lines 8505C, 8305C, and TPC-1 were more sensitive to MLN8237 (IC50 = 410–668 nM), while MLN8237 had little impact on the growth of K1, FTC133, and BCPAP cells with low c-Myc expression (IC50 = 2109–6108 nM; Fig. 1B). There was an average 7.3-fold change in the sensitivity between the cells with high and low c-Myc expression. This was also supported by colony formation assay (Fig. 1C). MLN8237 significantly inhibited colony formation in 8505C, 8305C, and TPC-1 cells in a dose-dependent manner, while it barely had an effect in K1, FTC133, and BCPAP cells (Fig. 1C). These findings indicate that c-Myc may be a major determinant for the response of thyroid cancer cells to MLN8237.
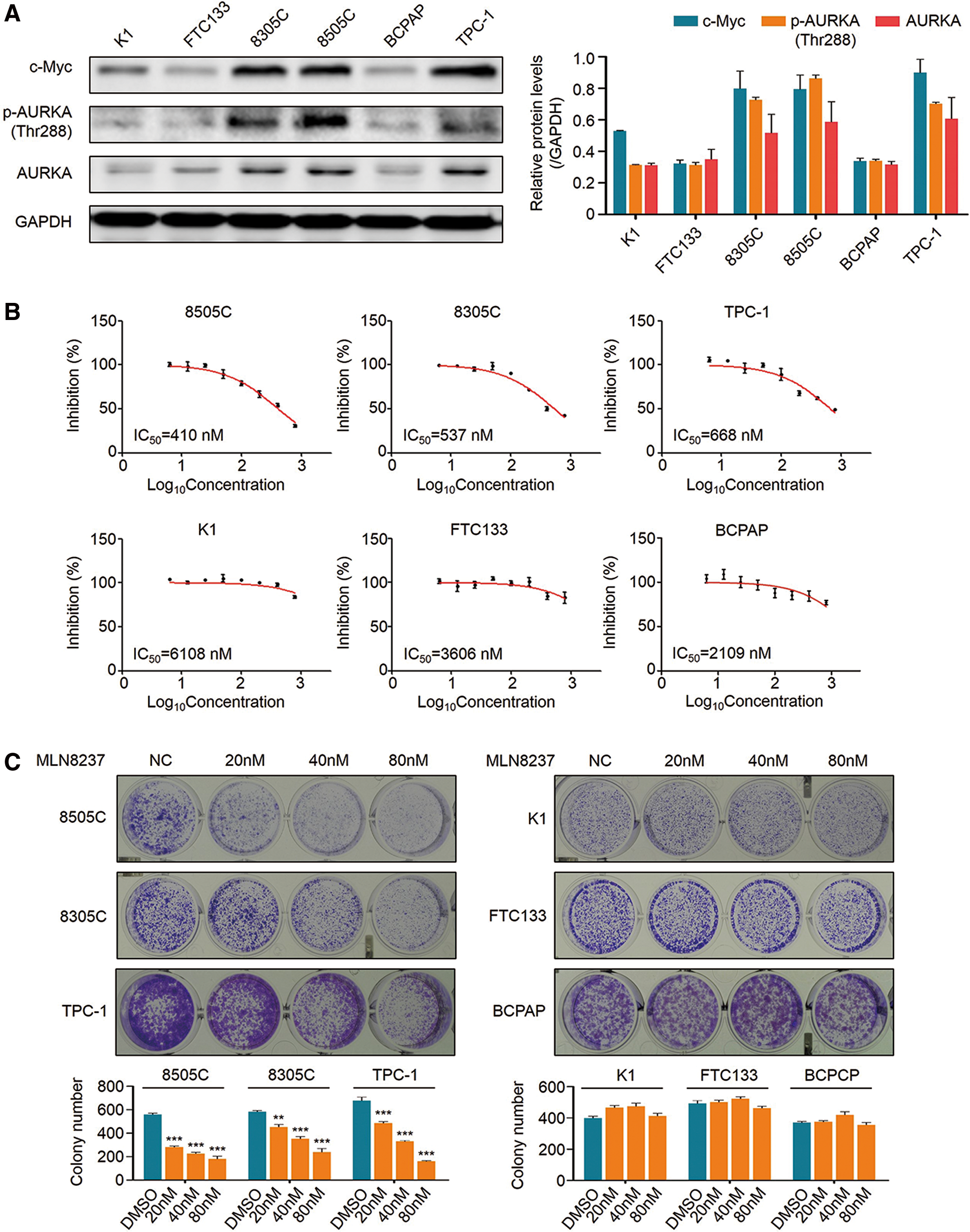
Cellular effects of MLN8237 on thyroid cancer cell lines. (
MLN8237 induces apoptosis in the cells with high c-Myc expression
To test the effect of MLN8237 on cell apoptosis, 8505C, 8305C, K1, and FTC133 cells were treated with 250 nM MLN8237 for 24 h. As shown in Figure 2A, 8505C and 8305C cells exhibited a remarkable increase in both early and late apoptosis upon MLN8237 treatment relative to the control. The percentage of apoptotic cells increased from 8.1 ± 0.2% to 14.5 ± 0.1% in 8505C cells, and from 2.3 ± 0.1% to 9.4 ± 0.3% in 8305C cells. However, a significant effect of MLN8237 was not found on apoptosis in K1 and FTC133 cells compared to the control (Fig. 2B). These data further support that cells with high c-Myc expression are more sensitive to MLN8237 relative to those with low c-Myc expression.
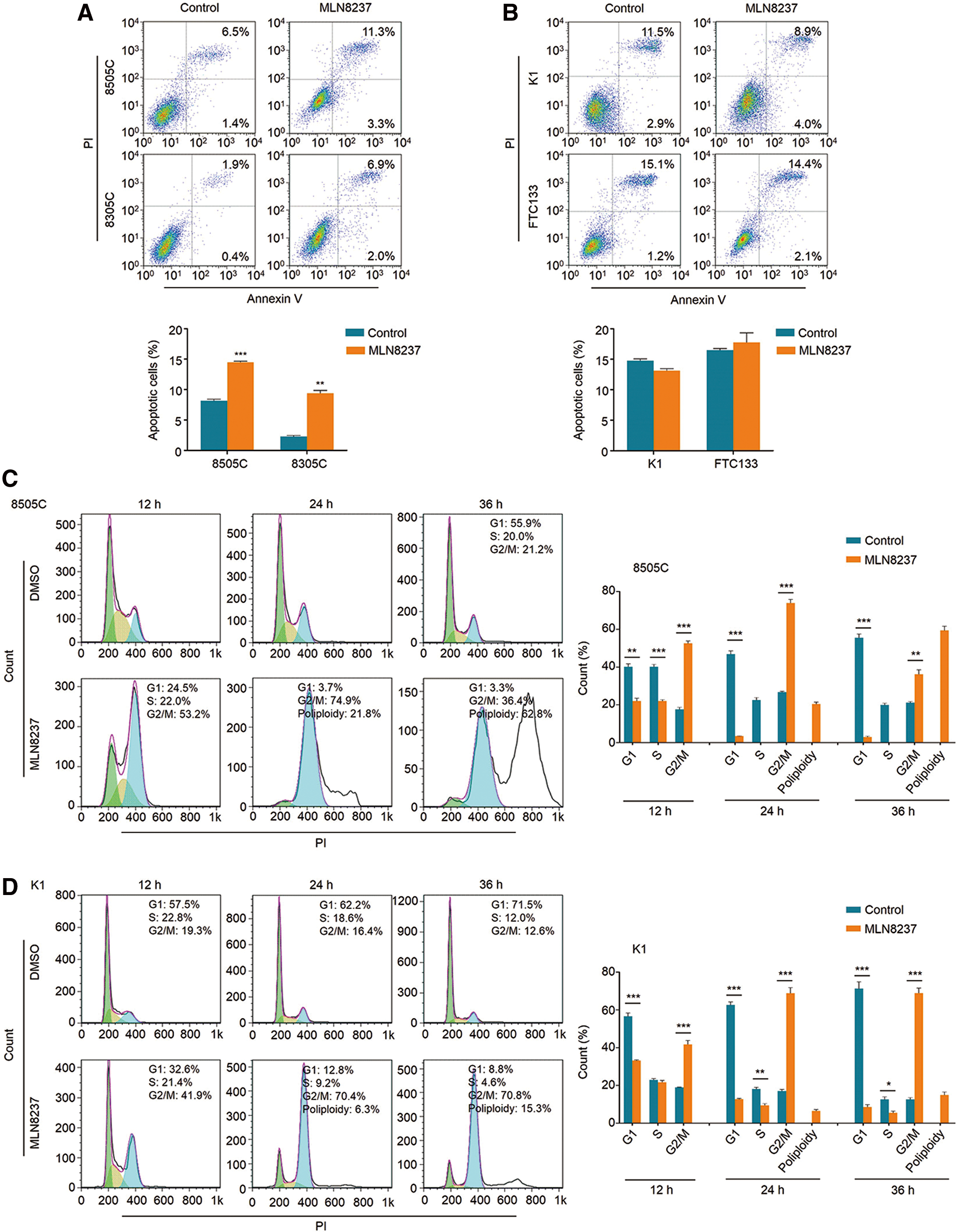
Induction of apoptosis and cell cycle arrest by MLN8237 in thyroid cancer cells. (
MLN8237 induces G2/M phase arrest and polyploidy in thyroid cancer cells
AURKA is a serine/threonine kinase, which plays a pivotal role in mitosis, and inhibition of AURKA can lead to cell cycle arrest and polyploidy in the cell cycle (24,25). This study treated 8505C and K1 cells with 250 nM MLN8237 for the indicated times. Cell cycle analysis showed a statistically significant increase in G2/M phase cells in both 8505C and K1 cells at 12, 24, and 36 h (Fig. 2C and D). The percentage of G2/M phase cells increased from 17.6 ± 1.1% to 52.6 ± 1.3% at 12 h, 26.7 ± 0.5% to 73.9 ± 1.9% at 24 h, and 21.1 ± 0.6% to 36.2 ± 2.3% at 36 h in 8505C cells, and increased from 19.0 ± 0.2% to 41.7 ± 2.0% at 12 h, 17.1 ± 0.8% to 68.9 ± 2.9% at 24 h, and 12.6 ± 1.0% to 68.9 ± 2.6% at 36 h in K1 cells. Like AURKA, c-Myc also plays an essential role in the regulation of the cell cycle (26). Inhibition of c-Myc will block entry into the S phase, while inhibition of AURKA will cause a G2/M arrest (24). In this study, MLN8237 treatment induced a G2/M arrest in thyroid cancer cells, indicating that the effect of MLN8237 on cell cycle distributions of thyroid cancer cells is mainly caused by inhibition of AURKA phosphorylation. In addition, the data show that MLN8237 treatment at 24 and 36 h results in a prominent increase in the number of polyploidy cells in 8505C cells compared to that in K1 cells (Fig. 2C and D), further supporting the above conclusion.
MLN8237 inhibits xenograft tumor growth derived from thyroid cancer cells with high c-Myc expression
To determine whether the observed effects of MLN8237 in vitro can be seen in vivo, 8505C and K1 tumor xenografts were established in nude mice, and these mice were treated with MLN8237 (30 mg/kg) for the indicated times. As shown in Figure 3A (left panel), MLN8237 significantly inhibited the growth of 8505C cell-derived xenograft tumors in nude mice compared to the control. At the end of the experiments, the tumors were isolated and weighted. The mean tumor weight was significantly lower in MLN8237-treated mice relative to control group mice (Fig. 3A, right panel). As expected, MLN8237 did not affect the growth and weight of K1 cell–derived xenograft tumors in nude mice relative to the control (Fig. 3B).
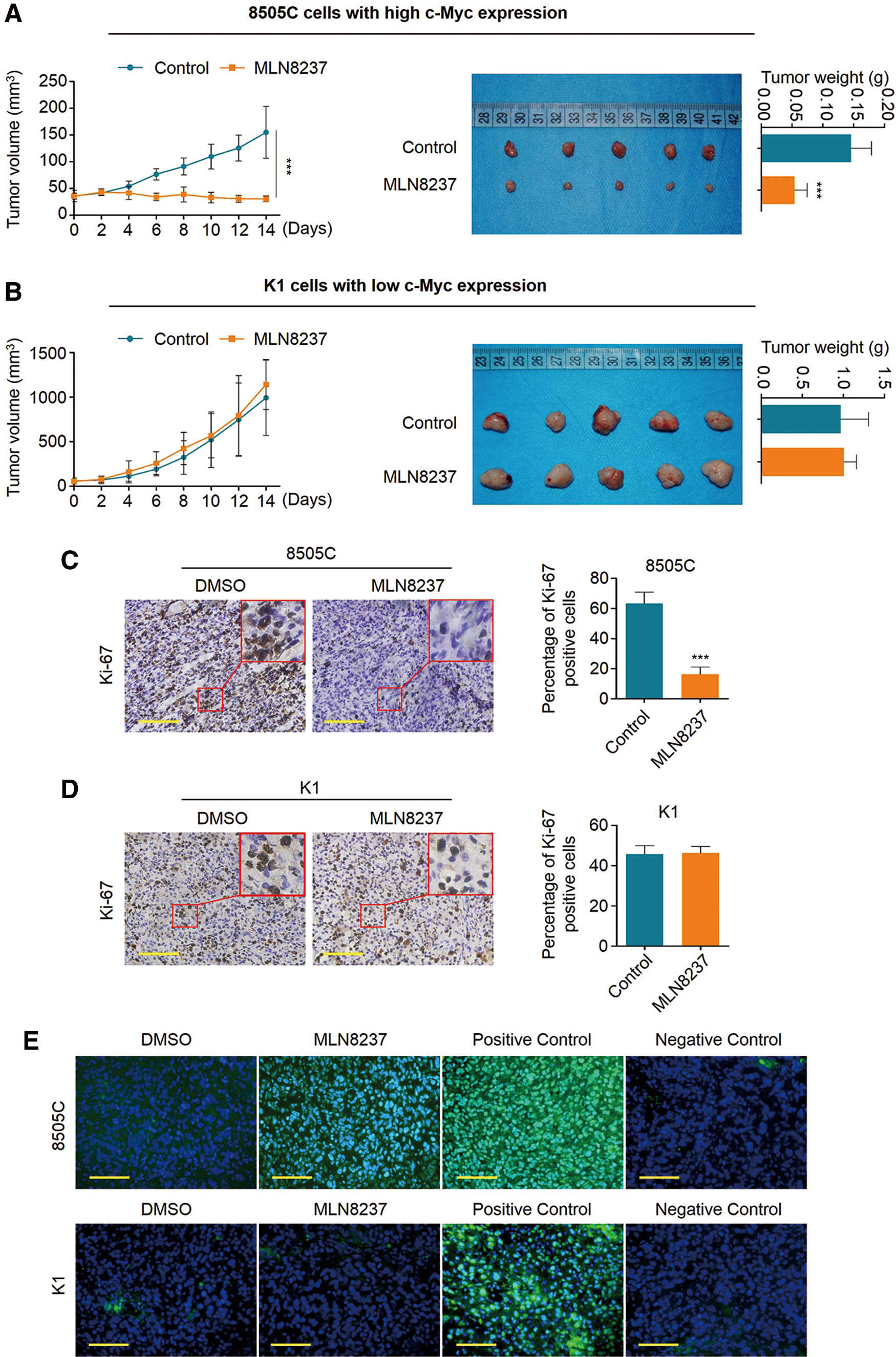
Antitumor effect of MLN8237 in xenograft tumors. (
Ki-67 is present in all proliferating cells, and serves as a proliferation marker (27). Thus, the levels of Ki-67 were evaluated using an IHC assay in this study. As shown in Figure 3C, the percentage of Ki-67-positive cells dramatically decreased in 8505C cell-derived xenograft tumors upon MLN8237 treatment compared to the control. However, MLN8237 treatment did not affect the percentage of Ki-67-positive cells in K1 cell–derived xenograft tumors compared to the control (Fig. 3D). Next, a TUNEL assay was also performed to evaluate apoptosis in xenograft tumors. The results show that the number of apoptotic cells was significantly increased upon MLN8237 treatment in xenograft tumors compared to the control (Fig. 3E, upper panels), while MLN8237 barely affected apoptosis in K1 cell–derived xenograft tumors (Fig. 3E, lower panels). Altogether, the data suggest that MLN8237 exhibits remarkable antitumor activity in thyroid cancer cells with high c-Myc expression both in vitro and in vivo.
MLN8237 downregulates c-Myc expression in thyroid cancer cells at a posttranscriptional level
To determine the regulatory role of MLN8237 in c-Myc expression, the thyroid cancer cell lines 8505C, 8305C, K1, and FTC133 were treated with 250 nM MLN8237 for one, two, and three days, respectively, and the effect of MLN8237 on c-Myc expression was evaluated by Western blot analysis. Expectedly, with increasing treatment time, MLN8237 significantly inhibited AURKA phosphorylation in 8505C and 8305C cells. Meanwhile, the levels of c-Myc proteins were also significantly downregulated upon MLN8237 treatment in these two cell lines (Fig. 4A). To exclude an off-target effect of MLN8237 on c-Myc expression, the expression of AURKA was inhibited by using a si-RNA approach, and it was found that c-Myc was also downregulated upon AURKA knockdown (Supplementary Fig. S1). However, in contrast to similar inhibitory effect of MLN8237 on AURKA phosphorylation in K1 and FTC133 cells, MLN8237 had little impact on the levels of c-Myc proteins in these two cell lines (Fig. 4B). This was further supported by an immunoblotting analysis of c-Myc and phosphorylated AURKA in 8505C/K1 cell-derived xenograft tumors (Fig. 4C). In addition, the effect of MLN8237 on c-Myc expression was also tested at the transcriptional level in 8505C, 8305C, K1, and FTC133 cells by qRT-PCR assay. The results show that mRNA expression of c-Myc was even upregulated upon MLN8237 treatment compared to the control, particularly in 8505C and 8305C cells (Supplementary Fig. S2). Taken together, these data indicate that MLN8237 downregulates c-Myc expression through a posttranscriptional mechanism in thyroid cancer cells with high c-Myc expression. In addition, to determine further whether the change in c-Myc protein levels mediated by MLN8237 can affect its transcriptional activity, the effect of MLN8237 on mRNA levels of three downstream targets of c-Myc (CAD, CDK4 and PTMA) was tested in 8505C, 8305C, K1, and FTC133 cells (13). The results show that MLN8237 decreased the levels of these three genes in 8505C and 8305C cells, as expected, but it had no impact in K1 and FTC133 cells (Supplementary Fig. S3).
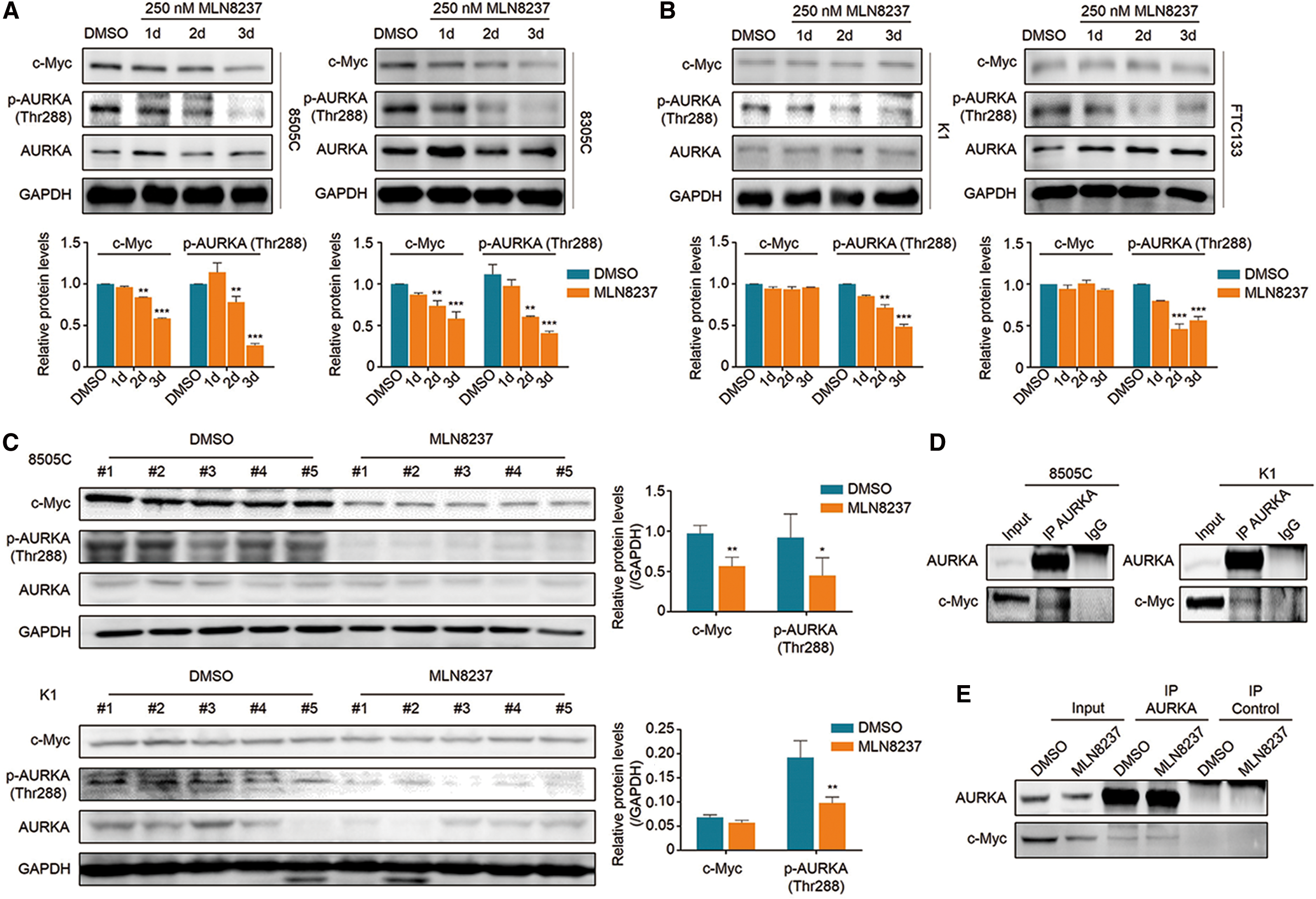
Effect of MLN8237 on c-Myc expression in thyroid cancer cell lines. (
There is evidence showing that AURKA forms a complex with c-Myc to stabilize c-Myc proteins in TP53-altered liver carcinomas (20). Thus, it was hypothesized that the c-Myc/AURKA complex could also contribute to the stabilization of c-Myc in thyroid cancer cells. First, the interaction between AURKA and c-Myc in 8505C and K1 cells was demonstrated using a reciprocal co-immunoprecipitation (Co-IP) assay, as shown in Figure 4D and Supplementary Figure S4. Given that AURKA inhibitors such as MLN8237 can induce conformational changes of AURKA proteins, thereby preventing the de novo formation of the complex and resulting in the degradation of c-Myc proteins (20), these cells were treated with MLN8237 at the indicated concentrations and times. The results indicate that MLN8237 treatment had almost no influence on the amount of preformed complex (Fig. 4E and Supplementary Fig. S5), as supported by a previous study that demonstrated that the preformed c-Myc/AURKA complex is very stable and will prevent AURKA inhibitors to access their binding site (20).
MLN8237 promotes proteasomal degradation of c-Myc proteins in thyroid cancer cells with high c-Myc expression
There is evidence showing that inhibition of AURKA can lead to an increased turnover of N-Myc proteins in MYCN-amplified cells (28). Thus, it was speculated that MLN8237 downregulates c-Myc expression through inducing destability of c-Myc proteins in thyroid cancer cells with high c-Myc expression. To assess this, first CHX was used to block new protein synthesis, and then the effect of MLN8237 on the stability of c-Myc proteins was tested by Western blot analysis in 8505C and K1 cells. The results showed that MLN8237 significantly accelerates the turnover of c-Myc proteins in 8505C cell (Fig. 5A), whereas it had little impact on c-Myc stability in K1 cells (Fig. 5B). Next, 8505C and K1 cells were treated with the proteasome inhibitor MG132 (25 μM) for 4 h to block the ubiquitin–proteasome pathway. As shown in Figure 5C, MG132 expectedly reversed the inhibitory effect of MLN8237 on c-Myc expression in 8505C cells (Fig. 5C), indicating that MLN8237 promotes c-Myc proteolysis by the ubiquitin–proteasome pathway. In addition, it was also found that the levels of AURKA proteins were increased upon MG132 treatment in both 8505C and K1 cells (Fig. 5C). This is consisitent with a previous study demonstrating that the stability of AURKA proteins are also regulated by the ubiquitin–proteasome pathway (29).
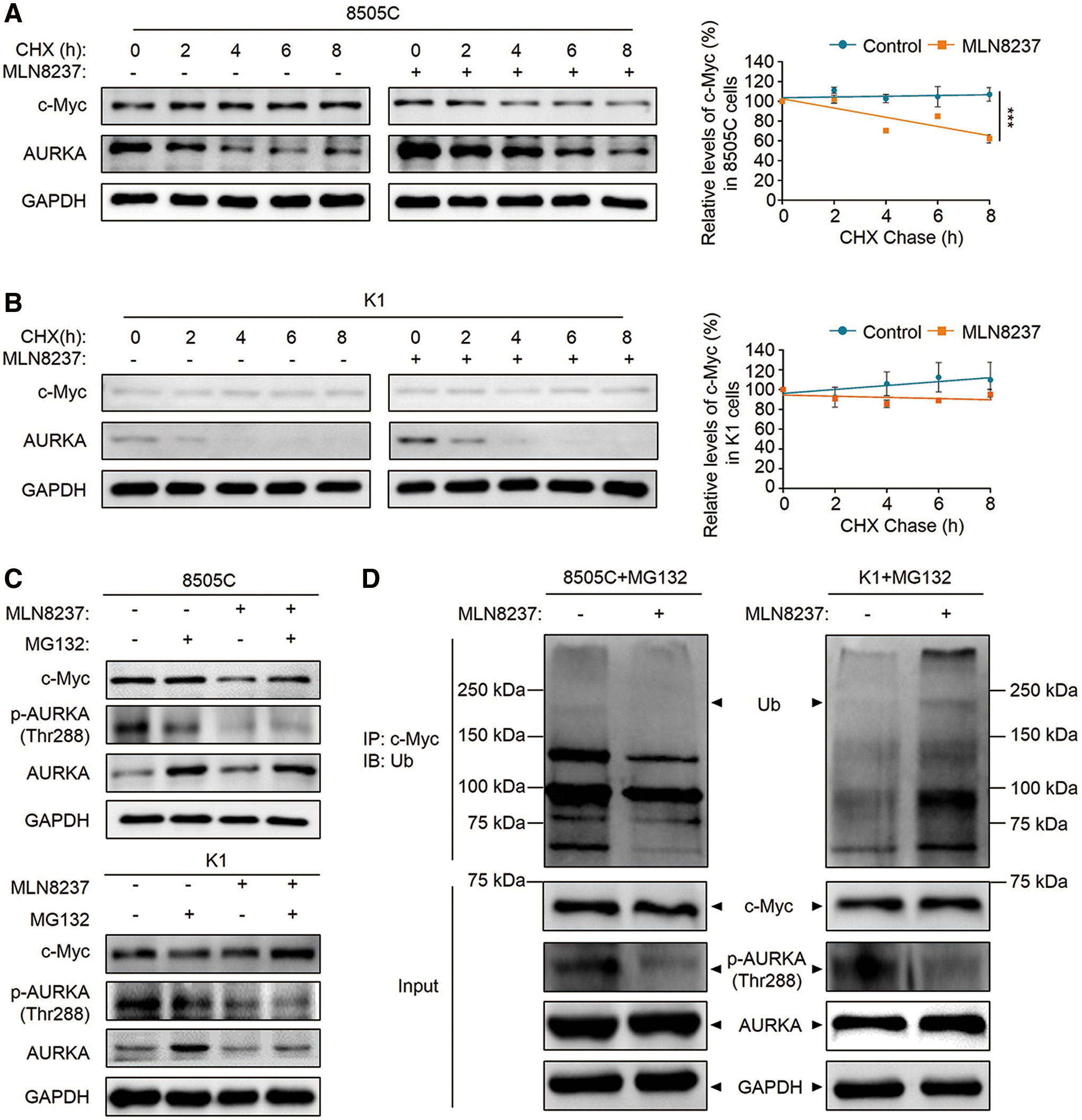
Effect of MLN8237 on c-Myc stability by proteasomal degradation. (
c-Myc is a highly unstable transcription factor, which is destroyed by ubiquitin-mediated proteolysis (30). Thus, this study tested the effect of MLN8237 on ubiquitination of c-Myc proteins in 8505C and K1 cells by Co-IP and Western blot assays. As shown in Figure 5D, surprisingly it was found that the ubiquitination of c-Myc proteins in 8505C cells was reduced upon MLN8237 treatment compared to the control, whereas it increased in K1 cells after MLN8237 treatment. This finding is supported by two previous studies demonstrating that AURKA stabilizes N-Myc by promoting the accumulation of ubiquitin chains that do not support degradation (28,31).
In brief, AURKA forms a complex with and stabilizes c-Myc proteins through protecting it from E3 ubiquitin ligase Fbw7-mediated proteasomal degradation in thyroid cancer cells with high c-Myc expression. MLN8237, a type of conformation-changing AURKA inhibitor, not only inhibits AURKA phosphorylation, but also promotes Fbw7-mediated proteolysis of c-Myc through disrupting the formation of the complex, thereby inhibiting the proliferation and inducing apoptosis and G2/M cell cycle arrest in thyroid cancer cells with high c-Myc expression.
Discussion
The oncoprotein c-Myc is a key transcription factor with essential functions in tumorigenesis, and it has been reported to correlate with poor prognosis in thyroid cancer (11,12). However, no available cavities, found in other enzymes such as kinases, have been identified in c-Myc proteins, which limits the development of small molecule inhibitors directly targeting c-Myc (14,15). In recent years, strenuous efforts have been made to inhibit the transcriptional acitivity of c-Myc either by interrupting c-Myc/Max dimerization and subsequently inhibiting DNA binding of c-Myc/Max (32,33), or by targeting the bromodomain and extraterminal (BET) subfamily of bromodomain-containing proteins (34,35). In addition, other strategies to inhibit c-Myc indirectly have been suggested by several previous studies that have shown that proteolytic turnover of N-Myc is regulated by a kinase-independent function of AURKA in neuroblastoma (14,28,36). In brief, AURKA protects N-Myc from proteasomal degradation by forming a complex, and this process is independent of the catalytic activity of AURKA. AURKA inhibitors disrupt the N-Myc/AURKA complex and subsequently lead to the degradation of N-Myc through inducing a conformation distortion in the catalytic domain of AURKA. Similarly, AURKA also forms a complex with c-Myc in hepatocellular carcinoma cells (20). Taken together, these observations raise the need to determine whether c-Myc can be stabilized by AURKA in thyroid cancer cells, and in turn be used as a specific determinant for the efficacy of AURKA-targeted therapies.
MLN8237 is a inhibitor developed from MLN8054 (17), which preferentially inhibits AURKA versus Aurora B kinase (AURKB), with a selectivity of approximately 200-fold (24). Completed Phase II studies of MLN8237 have shown promising anticancer effects in numerous types of cancers compared to other AURKA inhibitors (19). However, its antitumor effects in thyroid cancer remain largely unclear. Data in the present study show that MLN8237 exhibits a diversity of growth responses in thyroid cancer cells in vitro and in vivo. Thyroid cancer cells with high c-Myc expression were relatively more sensitive to MLN8237 than those with low c-Myc expression. MLN8237 dramatically inhibited cell proliferation and colony formation, and induced G2/M cell cycle arrest and apoptosis in the MLN8237-sensitive cells (or cells with high c-Myc expression), whereas it barely affected the growth of MLN8237-resistant cells (or cells with low c-Myc expression). Thus, c-Myc may contribute to a distinct cellular response of MLN8237, and c-Myc levels may be a major determinant for the response of thyroid cancer cells to MLN8237.
To explore the mechanism of MLN8237 affecting the responsiveness of MLN8237 in thyroid cancer cells, the effect of MLN8237 on the expression of c-Myc proteins was tested in a panel of thyroid cancer cell lines and xenograft tumors. The results show that MLN8237 effectively decreased the abundance of c-Myc proteins in the cells with high c-Myc expression, but not in those with low c-Myc expression. In addition, MLN8237 did not decrease the transcription of c-Myc in the same cell lines, indicating that MLN8237 downregulates c-Myc expression in the MLN8237-sensitive cells at a posttranscriptional level. Next, the existence of the c-Myc/AURKA complex in thyroid cancer cells with high or low c-Myc expression was identified by using a Co-IP assay, and it was demonstrated that MLN8237 barely affected the interation between c-Myc and AURKA. This was supported by a previous study that MLN8237 only prevents the de novo formation of the c-Myc/AURKA complex. Thus, the preformed complex remains stable after treatment. These data provide a reasonable explanation for the reason why protein levels of c-Myc decrease two days after MLN8237 treatment. In addition, thyroid cancer cells were also treated with CHX to block new protein synthesis. The results showed that c-Myc proteins in the MLN8237-sensitive cells have a faster turnover upon MLN8237 treatment compared to the control, which was not the case in the MLN8237-resistant cells. Taken together, these findings further support the hypothesis. Briefly, in thyroid cancer cells with high c-Myc expression, c-Myc stabilization is mediated by the AURKA proteins via forming a c-Myc/AURKA complex, and MLN8237 destabilizes and subsequently degrades c-Myc proteins through disrupting de novo formation of this complex.
c-Myc is highly unstable protein and can be degraded by ubiquitin-mediated proteolysis (30). Thus, next, the effect of MLN8237 on ubiquitination of c-Myc proteins in thyroid cancer cells was investigated. Surprisingly, the data showed that MLN8237 decreased ubiquitination of c-Myc proteins in the MLN8237-sensitive cells, whereas it increased in the MLN8237-resistant cells. This is similarly supported by previous studies demonstrating that AURKA stabilizes N-Myc proteins through promoting the accumulation of ubiquitinated N-Myc at Lys63 (K63) and Lys11 (K11) that do not support degradation, in contrast to Lys48 (K48), which is required for proteasomal degradation (28,31). A similar mechanism may exist for c-Myc, as supported by a previous study demonstrating that ubiquitin ligase HectH9 assembles predominantly K63-linked chains on c-Myc, and ubiquitination of c-Myc by HectH9 can stimulate the transcriptional activity of c-Myc in addition to regulating turnover (37). In the MLN8237-sensitive thyroid cancer cells, through disruption of the c-Myc/AURKA complex, MLN8237 might decrease the accumulation of nondegradable ubiquitin chains on c-Myc that may be involved in regulating c-Myc stability. Meanwhile, MLN8237 promoted proteasomal degradation of c-Myc probably through increasing ubiquitination at individual lysine residues in c-Myc that are required for degradation, as supported by the data demonstrating that the proteasome inhibitor MG132 rescued the inhibitory effect of MLN8237 on c-Myc proteins. However, this effect may be missed by looking at total ubiquitination of c-Myc.
In summary, through a series of extensive in vitro and in vivo experiments, this study demonstrates that MLN8237 exhibits a promising antitumor effect in thyroid cancer cells with high c-Myc expression but not in those with low c-Myc expression. Thus, the data highlight that c-Myc may be a major determinant for MLN8237 cellular reponsiveness in thyroid cancer and suggest that targeting AURKA may be an effective therapeutic strategy for c-Myc-overexpressing thyroid cancers.
Footnotes
Acknowledgments
This work was supported by the National Natural Science Foundation of China (nos. 81572627, 81672645, and 81770787).
Author Disclosure Statement
The authors have no potential conflict of interest to disclose.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.