
Abstract
Background:
Thyrotoxicosis increases bone turnover, resulting in net bone loss. Sympathetic nervous system (SNS) activation, via β2-adrenoceptor (β2-AR) signaling, also has osteopenic effects. Because thyroid hormones (TH) interact with the SNS to regulate several physiological processes, we hypothesized that this interaction also occurs to regulate bone mass. Previous studies support this hypothesis, as α2-AR knockout (KO) mice are less susceptible to thyrotoxicosis-induced osteopenia. Here, we evaluated whether TH-SNS interactions in bone involve β2-AR signaling.
Methods:
Thyrotoxicosis was induced in 120-day-old female and male mice with β2-AR gene inactivation (β2-AR−/− ) by daily treatment with supraphysiological doses of triiodothyronine (T3) for 12 weeks. The impact of thyrotoxicosis on femoral bone microarchitecture, remodeling, fracture risk, and gene expression of the receptor activator of nuclear factor-kappa-B (RANK)-RANK ligand (RANKL)-osteoprotegerin (OPG) pathway was evaluated. In addition, the effect of the β2-AR-specific agonist clenbuterol (CL) on cAMP accumulation was determined in osteoblastic (MC3T3-E1) cells treated with T3 and/or 17β-estradiol (E2).
Results:
Thyrotoxicosis negatively affected trabecular bone microarchitecture in wild-type (WT) females, but this effect was milder or nonexistent in β2-AR−/− animals, whereas the opposite was seen in males. T3 treatment increased the femoral RANKL/OPG mRNA ratio and the endosteal perimeter and medullary area of the diaphysis in WT females and males, but not in β2-AR−/− mice, suggesting that T3 promotes endosteal resorption in cortical bone, in a mechanism that involves β2-AR signaling. T3 treatment increased endocortical mineral apposition rate only in WT females but not in β2-AR −/− mice, suggesting that TH also induce bone formation in a β2-AR signaling-dependent mechanism. T3 treatment decreased femoral resistance to fracture only in WT females, but not in KO mice. E2 and CL similarly increased cAMP accumulation in MC3T3-E1 cells; whereas T3 alone had no effect, but it completely blocked E2-stimulated cAMP accumulation, suggesting that some T3 effects on bone may involve E2/cAMP signaling in osteoblasts.
Conclusions:
These findings sustain the hypothesis that T3 interacts with the SNS to regulate bone morphophysiology in a β2-AR signaling-dependent mechanism. The data also reveal sex as an important modifier of skeletal manifestations of thyrotoxicosis, as well as a modifier of the TH-SNS interactions to control bone microarchitecture, remodeling, and resistance to fracture.
Introduction
Thyroid hormones (TH) are key regulators of bone metabolism, controlling both bone formation and resorption. In conditions of TH excess, both osteoblastic and osteoclastic activities are increased, but the latter predominates, leading to hypercalcemia and bone loss (1,2). The osteopenic effects of TH were first described more than a century ago (3) and overt thyrotoxicosis is recognized as a well-established cause of secondary osteoporosis (4,5). In the past two decades, the sympathetic nervous system (SNS) has also been identified as a pivotal regulator of bone remodeling and bone mass (6 –8). Evidence shows that SNS activation decreases bone formation and increases bone resorption (9), leading to bone loss, through a mechanism that involves β2-adrenoceptors (β2-AR), expressed in osteoblasts (6).
The role of the SNS in the control of bone remodeling is supported by a high bone mass (HBM) phenotype in mice deficient in dopamine β-hydroxylase (DbH−/−), the limiting enzyme responsible for the synthesis of catecholamines (10), and in mice with global (11) and osteoblast-specific (12) gene inactivation of β2-AR (β2-AR−/− and β2osb-AR−/− mice, respectively). These β2-AR knockout (KO) mice present normal body weight (BW) and a HBM phenotype by six months of age, which occurs due to increased bone formation and decreased bone resorption (11,12). In addition, the administration of β-adrenergic agonists and antagonists to rodents has been shown to decrease and increase bone mass, respectively (10,13,14).
On the other hand, studies by our group have demonstrated that α2-adrenergic receptors (α2A-ARs) also mediate SNS actions in the skeleton (15,16), and that mice with a double KO for genes that encode α2A-AR and α2C-AR subtypes (α2A/2C-AR−/− mice) also present an HBM phenotype (16). In addition, we demonstrated that all α2-AR isoforms (α2A-AR, α2B-AR, and α2C-AR) are expressed in the major bone cells (osteoblasts, osteocytes, and osteoclasts) and in chondrocytes (16). α2-ARs are presynaptic autoreceptors that regulate the release of catecholamines. Thus, α2A/2C-AR−/− mice present a chronic elevation of the sympathetic tone (17). The fact that α2A/2C-AR−/− animals present HBM phenotype, regardless of showing increased sympathetic tone and normal expression of β2-AR, demonstrates that the latter is not the only adrenoceptor involved in the control of bone metabolism, and that α2A-AR and/or α2C-AR play an important role in mediating the SNS actions on the regulation of bone mass (16).
Interestingly, an important feature of TH, but still poorly understood, is their interaction with the SNS. It is well known that interactions between TH and catecholamine actions are necessary for maximum thermogenesis, lipolysis, glycogenolysis, and gluconeogenesis (18). Remarkably, patients with hyperthyroidism usually present numerous clinical features of increased adrenergic activity, which include tachycardia, increased cardiac output, increased glycogen and lipid mobilization, enhanced thermogenesis, tremor, hyperkinetic behavior, and sweating (19). The well-known effectiveness of β-blockers in ameliorating the signs and symptoms of hyperthyroidism suggests important roles of β-adrenoceptor signaling in the TH-SNS interactions to control many physiological processes.
Interestingly, treatment of hyperthyroid patients with propranolol (a β-adrenergic antagonist) was shown to correct the hypercalcemia secondary to thyrotoxicosis (20 –22); to decrease the urinary excretion of hydroxyproline (23), a biochemical marker of bone resorption (24); and to lower serum alkaline phosphatase (22), a biochemical marker of bone formation (24). These findings suggest that TH also interact with the SNS to control bone mass and remodeling, in a process that involves β-adrenergic receptor-dependent mechanisms. It is noteworthy that a TH-SNS interaction to control bone mass and function has been supported by the findings that mice with double KO for α2A- and 2C-AR (α2A/2C-AR−/− ) and mice with single KO for α2C-AR (α2C-AR−/− ) are resistant to the thyrotoxicosis-induced osteopenia (15,25), which indicates an important role of α2-AR signaling in this interaction.
To investigate the hypothesis that β2-AR signaling is also relevant to TH-SNS interactions in bone physiology, in this study, we evaluated the effect of supraphysiological doses of triiodothyronine (T3) on the microarchitecture and biomechanical parameters of the femur of female and male mice with β2-AR gene inactivation (β2-AR−/− mice). These new data show that lack of β2-AR signaling impairs or blocks the deleterious effects of thyrotoxicosis on the microarchitecture of trabecular and cortical bone and on the biomechanical properties of the femur in female mice; whereas, in males, similar effects were observed only in cortical bone. These findings sustain the hypothesis that TH interact with the SNS, in a β2-AR-dependent mechanism, to regulate bone microarchitecture, remodeling, and resistance to fracture.
Materials and Methods
Animals and treatments
β2-AR−/−
mice were generated as previously described (26). Briefly, disruption of the β2-AR gene was achieved by using a positive
A cohort of female and male congenic β2-AR−/− mice and the WT FVB/N controls were studied. All animals were 120°days old at baseline. The animals were kept under light- and temperature-controlled conditions (alternating cycles of light/dark for 12 hours at a temperature of ∼25°C), with ad libitum access to food and water. Thyrotoxicosis was induced by daily and intraperitoneal administration of T3 (Sigma, St Louis, MO), at a dose of 3.5 or 7 μg T3/100 g BW/day, which are equivalent, respectively, to 10 and 20 times the physiological dose of T3 per day (10 × T3 and 20 × T3, respectively). Both T3 treatments and saline (T3-untreated) were administered for 12 weeks, at the same time each day. Animals were weighed once a week to monitor BW changes over the experimental period and for adjusting the amount of T3 to be administered, to maintain the 10 × T3 and 20 × T3 doses during the 12-week treatment period.
Female and male mice were grouped as follows: WT and β2-AR−/− (T3-untreated animals, receiving saline), WT +10 × T3 and β2-AR−/−+10 × T3, and WT +20 × T3 and β2-AR−/− +20 × T3 (8–11 animals per group). At the end of the treatment period, the animals were euthanized by exposure to CO2. The experimental procedures were carried out in accordance with the ethical principles and guidelines for animal research set by the Brazilian Society of Animal Experimentation, and they were approved by the Ethics Committee on Animal Use (ECAU) of the Institute of Biomedical Sciences and the Ethics Committee on the Use of Animals of the School of Medicine of the University of São Paulo (Protocol No. 121/2015 and 158/15, respectively).
Serum levels of total T3 and T4
At the end of the T3 treatment period, after 1 hour of the last T3 administration, the animals were euthanized; the blood was collected; and the serum was separated by centrifugation and immediately frozen at −20°C. Serum levels of total thyroxine (T4) and total T3 (TT4 and TT3, respectively) were determined by means of an electro-chemiluminescence immunoassay, using the commercial kits Elecsys® T4 and Elecsys® T3 Cobas® (Roche Diagnostics GmbH, Mannheim, Germany).
Body and femoral length and heart mass
Body length was measured immediately after euthanasia, from the tip of the snout to the base of the tail, by using a ruler. The length of the right femur was measured between the proximal and distal ends, using a digital caliper (Digimess) in dissected bones. Immediately after the animals were euthanized, the heart was dissected out and weighed for wet-weight determination. The heart samples were dehydrated at 60°C for 48 hours and weighed again for dry-mass determination. All masses were expressed in milligrams per gram of BW.
Micro-computed tomography
The right femur of each animal was scanned by using the micro-computed tomography (μCT) unit Skyscan 1174 (Bruker microCT, Kontich, Belgium). The X-ray settings were standardized to 100 kV for the baseline specimens, with an exposure time of 590 ms. A 0.05-mm-thick aluminum filter and a beam-hardening algorithm were used to minimize beam-hardening artifacts. The bone parameters were obtained with the CtAn Software, Version 1.5 (BrukermicroCT). The region of interest analyzed was the proximal half of the distal third of the femur. This region was delimited by dividing the total femoral length by 3, for the selection of the distal third, whose length was divided by 2, for the selection of its proximal half.
The trabecular parameters analyzed were bone volume/total volume (BV/TV; %), which is a measure of bone volume; trabecular thickness (
The cortical parameters were total tissue area, which is the total area delimited by the periosteum (
Bone histomorphometry
The skeletons of all animals were double labeled with tetracycline (Amresco, Solon, OH) and calcein (Sigma) administered subcutaneously. Tetracycline was administered in a dose of 2 mg/100 g of BW 8 and 9 days before euthanasia, and calcein was administered in a dose of 1.5 mg/100 g of BW 3 and 4 days before euthanasia. Just after euthanasia, the tibias were carefully dissected and processed as previously described (27). Briefly, undecalcified tibias were fixed in 70% ethanol, dehydrated, and embedded in methyl methacrylate. Ten-micrometer-thick sections of the mid-diaphysis and of the proximal metaphysis of the tibia were cut by using a Polycut S equipped with a tungsten carbide knife (Leica, Heidelberg, Germany).
Endocortical mineral apposition rate (
Biomechanical test of the femur
Biomechanical parameters of the femurs were obtained by the three-point bending test, using the Instron testing system Model 3344 (Instron Corporation, MA). A load cell with a maximum capacity of 100 kgf was used. During the test, the anterior cortex of the femur was placed in compression and the posterior cortex in tension. The femur was supported on two anvils, spaced by a distance that equals the half of the femoral length (femoral length/2). A force was applied perpendicular to the longitudinal axis of the femur, on the diaphysis midpoint (between the two anvils), and in the antero-posterior direction of the bone by a crosshead, at a constant velocity of 5 mm/min, until the bone completely ruptured. To stabilize the specimen, a small preload (5% of the average maximal load) was applied before actual testing.
During the bending test, the load displacement data were collected, at a sampling rate of 80 Hz, by a computerized data-acquisition system, the Bluehill 3 (Instron Corporation), which constructed force-strain curves, from which the following biomechanical parameters were obtained: (i) Maximum load (N), which corresponds to the greatest force applied during the test (refers to the maximum force at which the bone is able to withstand); (ii) stiffness (N/mm), a measure of the extrinsic rigidity of the bone tissue; (iii) resilience (J), a measure of the ability of a bone to suffer elastic deformity (corresponds to the energy absorbed by a material during the elastic deformation); and (iv) tenacity (J), which represents the bone's ability to withstand a fracture (corresponds to the energy absorbed until the complete fracture).
RNA isolation and real-time polymerase chain reaction
Gene expression of receptor activator of nuclear factor kappa-B (RANK), RANK ligand (RANKL), and osteoprotegerin (OPG) was determined by real-time polymerase chain reaction (PCR) in the whole femur. The femurs were dissected and then crushed in a steel mortar and pestle set (Fisher Scientific International, Inc., Hampton, NH) as previously described (28). Total RNA was isolated from the crushed bones by using Trizol (Invitrogen, Carlsbad, CA), following the manufacturer's instructions. Total RNA was reverse transcribed by using RevertAid-H-Minus M-MuLV Reverse Transcriptase (Fermentas, Hanover, MD) to synthesize the first-strand cDNA. SYBR Green Super Mix (Applied Biosystems, Warrington, United Kingdom) was used for the real-time PCR by using the ABI Prism 7500 sequence detector (Applied Biosystems, Foster City, CA).
Real-time PCR analyses were done in duplicate. Threshold cylce (CT) values were normalized by using 18S expression as the internal control, which was validated for this study, showing it to be stable (its expression did not vary due to mice lineage or T3 treatment). The primers used in this study [RANK_F: TCT GCA GCT CTT CCA TGA CAC T and R: CGA TGA GAC TGG GCA GGT AAG (NM_009399), RANKL_F: GGC CAC AGC GCT TCT CAG and R: GAG TGA CTT TAT GGG AAC CCG AT (NM_011613.2), OPG_F: AGT CCG TGA AGC AGG AGT G and R: CCA TCT GGA CAT TTT TTG CAA A (NM_U94331), 18S_F: GTA ACC CGT TGA ACC CCA TT and R: CCA TCC AAT CGG TAG TAG CG (NM_11188)] were designed by using the Primer Express software (Applied Biosystems™) and were synthesized (Integrated DNA Technologies, Coralville, IA) specifically for real-time PCR. Relative gene expression quantification was assessed by the CT method, as previously described by Livak and Schmittgen (29).
Cell culture and cAMP content quantification
Mouse calvaria-derived MC3T3-E1 cells were cultured in Minimum Essential Medium Eagle (Sigma) supplemented with 10% Fetal Bovine Serum (Gibco-BRL, Grand Island, NY), and they were maintained in a CO2 automatic incubator (at 37°C, 5% CO2 in 95% air). For the cAMP content estimation experiments, cells were grown to confluence and differentiated into mature osteoblasts for 11 days. The cells were then left untreated (controls) or treated with 10−8 M T3 (Sigma) and/or with 10−8 M 17β-Estradiol 3-benzoate (E2; Sigma) for 72 hours. Finally, T3, E2, and untreated cells were treated with 10−4 M clenbuterol (CL; Sigma) during the last 30 minutes of culture.
After the indicated times of treatment, cells were scraped off and dissociated in 0.1 M HCl, by pipetting up and down, to a homogenous suspension. After top-speed centrifugation for 10 minutes, the supernatant was used to measure the intracellular cAMP content by using a cAMP immunoassay kit (ab65355; Abcam), according to the manufacturer's instructions. All samples and standards were run in triplicate.
Statistical analysis
Two-way ANOVA (analysis of variance) was used to compare more than two groups and was always followed by the Tukey's test to detect differences between groups. For all tests, p < 0.05 was considered statistically significant. The unpaired Student's t-test was used for pairwise comparisons of the groups. All results are expressed as the mean ± standard error of the mean. For statistical analysis, we used the GraphPad Instat Software (GraphPad Software, Inc., San Diego, CA).
Results
The effect of T3 treatment on serum levels of total T3 and T4
Serum levels of TT3 were not different between saline-treated WT and β2-AR−/− female and male mice (Fig. 1A, C). In females, TT4 levels were also quite similar between WT and KO mice (Fig. 1B), but in males, they were 32% lower in β2-AR−/− versus WT mice (Fig. 1C). As shown in Figure 1A, 10 × T3 similarly increased serum levels of TT3 in WT and KO female mice (6.6- and 7.1-fold vs. saline, respectively), and 20 × T3 further increased serum levels of TT3 in these animals (15- and 26-fold, respectively). In males, 10 × T3 and 20 × T3 similarly increased serum levels of TT3, in a range of 12- to 20-fold versus saline (Fig. 1C), which certainly suppressed the hypothalamic-pituitary-thyroid axis (30), leading to decreased serum levels of TT4 in WT and KO mice (Fig. 1B, D).
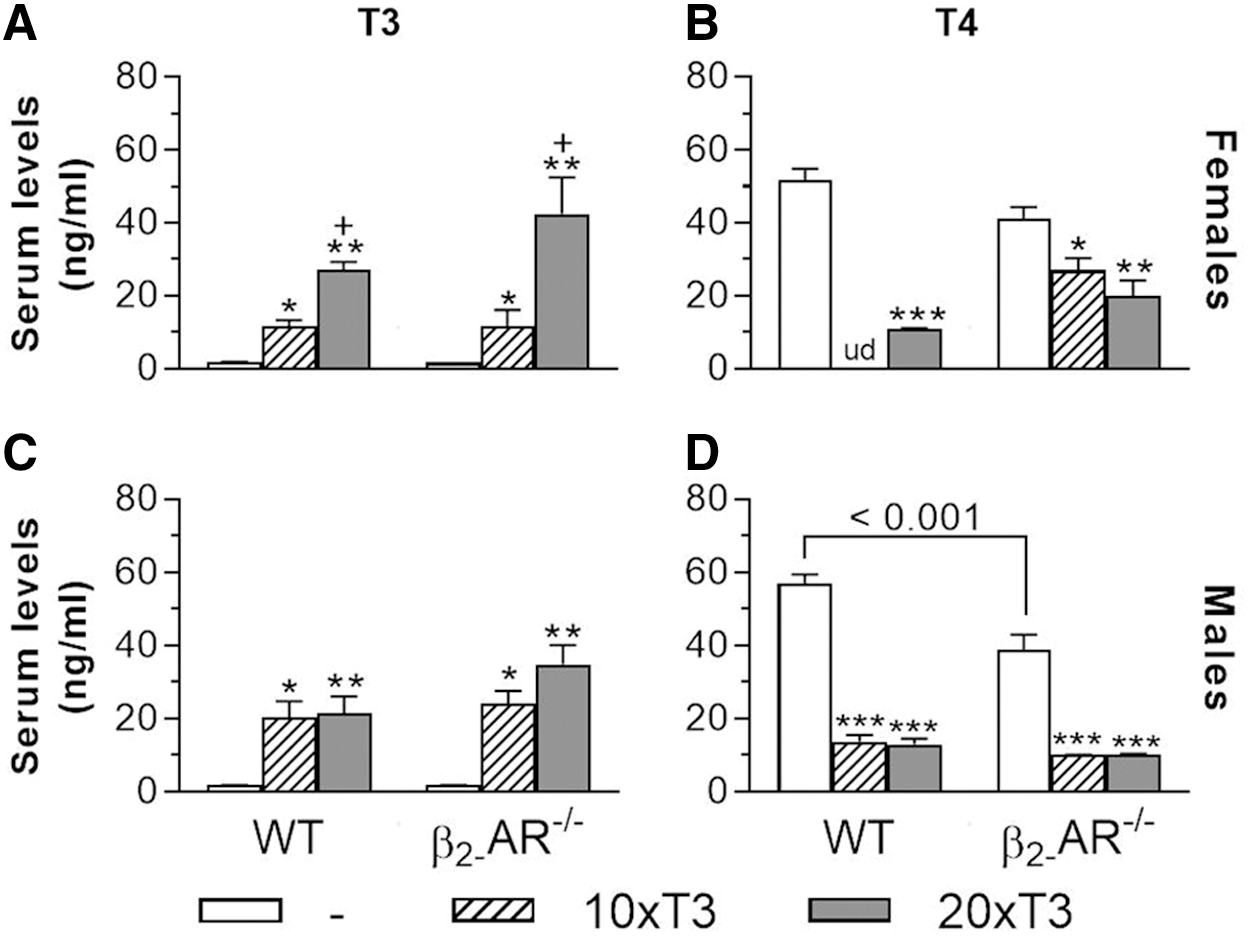
Serum levels of T3 and T4 in WT and β2-AR
−/−
mice. Animals were treated with 10 times (10 × T3) or 20 times (20 × T3) the physiological dose of T3 (3.5 and 7 μg/100 g BW/day, respectively) or with saline for 12 weeks, by daily IP injections. Serum levels of T3 and T4 were determined after one hour of the last T3 injection. (
Effect of thyrotoxicosis on BW, body length, and heart mass
Corroborating previous studies (11,26), there were no BW differences between saline-treated WT and β2-AR−/− animals, for both females and males, during the whole experimental period (Supplementary Fig. S1A, D). Treatment with 10 × T3 and 20 × T3 did not promote significant BW changes in female and male WT and KO mice (Supplementary Fig. S1B, C, F). However, treatment with 20 × T3 promoted significant increases in BW in WT males, mainly between weeks 9 and 12 (Supplementary Fig. S1E), which is probably due to the thyrotoxicosis-induced increase in water and food intake. It is noteworthy that increases in BW, as a consequence of hypothyroidism, have also been identified in C57BL/6NTac mice, but mainly in females (31,32), suggesting that sex and the genetic background influence the effects of TH on BW.
Despite the presence of β2-AR in the epiphyseal growth plate (16), body length and femoral length (data not shown) were not different between WT and β2-AR−/− mice (females and males), suggesting that β2-AR has no relevant role in bone elongation. Treatment with T3 did not promote changes in this bone parameter (not shown), which is expected for mice at the age range selected for this study. Corroborating previous studies reporting cardiac hypertrophy in conditions of thyrotoxicosis (33), both WT and KO male mice treated with 20 × T3 for 12 weeks presented increased heart mass (an indirect measure of cardiac hypertrophy) versus their saline-treated controls (29%, p < 0.01 and 19%, p < 0.05, respectively), which confirms a thyrotoxic state in our animals. Heart mass tended to be higher in T3-treated WT and KO female mice, but it was not statically significant (not shown).
Lack of β2-AR modifies the response of femoral trabecular bone to thyrotoxicosis
In the femur of female mice (Fig. 2A–F), thyrotoxicosis clearly promoted more severe adverse effects on trabecular bone of WT animals than in β2-AR−/−
animals. Treatment with 10 × T3 caused a significant decrease in BV/TV (33%) and Tb.N (33%), and a significant increase in Tb.Sp (26%) only in WT animals, but not in β2-AR−/−
animals (Fig. 2A, B, D, respectively). Treatment with 20 × T3 caused a reduction in BV/TV and Tb.N and an increase in Tb.Sp in WT (56%, 49%, and 28%, respectively) and β2-AR−/−
(39%, 36%, and 27%, respectively) females, but these effects tended to be more intense in WT than in KO female mice. In addition, treatment with 20 × T3 caused a reduction in
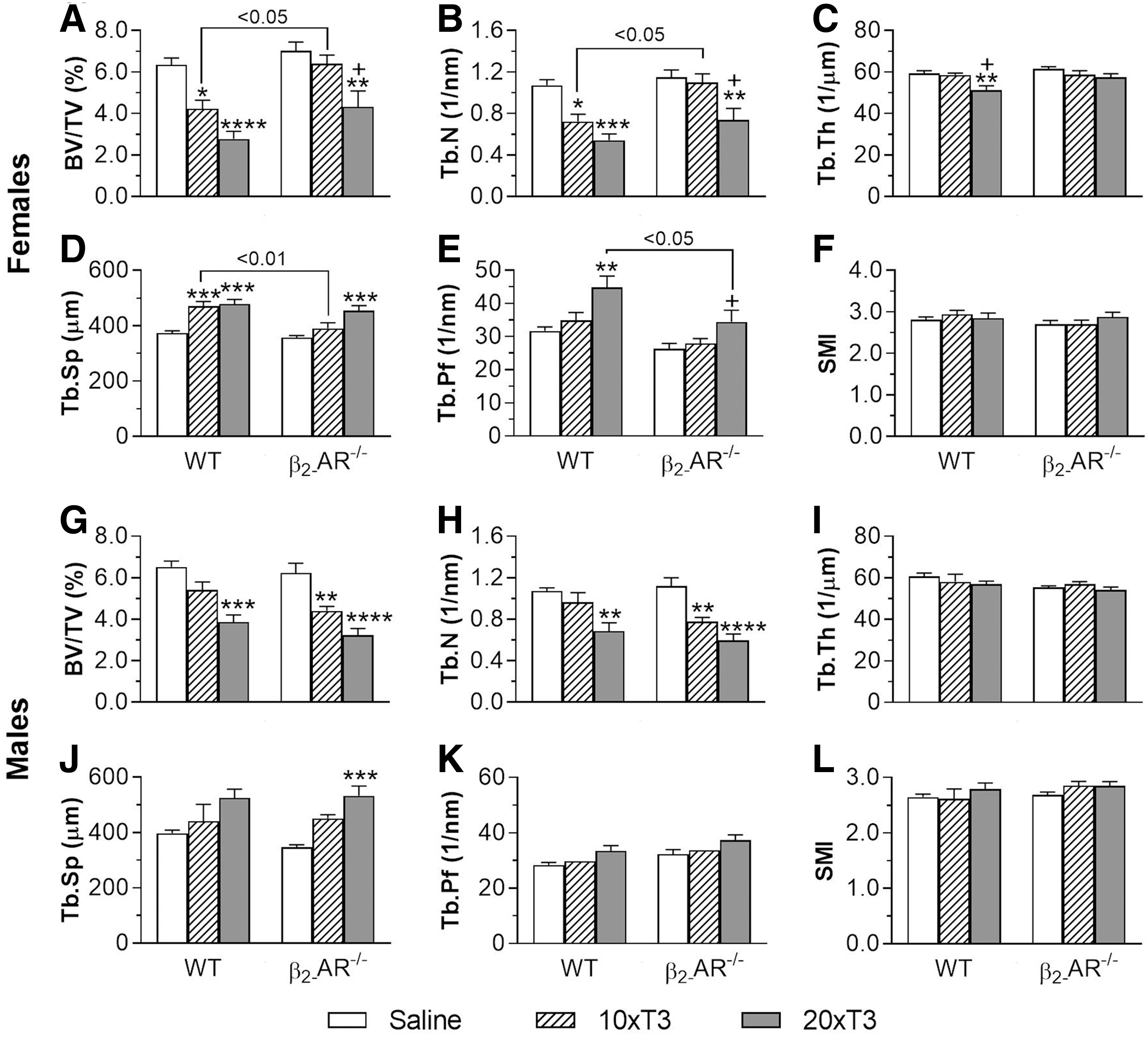
Effect of thyrotoxicosis on trabecular microarchitecture of the femur in WT and β2-AR
−/−
mice. (
There were no differences between saline-treated WT and KO animals, for any of the trabecular parameters evaluated in this study. However, among the female mice treated with 10 × T3, a higher BV/TV and Tb.N and a lower Tb.Sp (51%, 52%, and 17%, respectively) were observed in KO compared with WT animals (Fig. 2A, B, D, respectively). We also noticed a 23% lower
Interestingly, the femoral trabecular bone of WT male mice (Fig. 2G–L) was less sensitive to TH actions than that of WT females. Differently from female mice, only the highest dose of T3 (20 × T3) had negative effects on the trabecular bone of WT males, causing significant reductions only in BV/TV (41%) and Tb.N (36%) (Fig. 2G, H, respectively). In addition, and contrasting the female findings, β2-AR−/− male mice were slightly more sensitive to the detrimental effects of thyrotoxicosis on the femoral trabecular bone than WT males.
As shown in Figure 2G and H, only 20 × T3 had negative effects on BV/TV and Tb.N in WT male mice (decreases of 41% and 36% vs. saline-treated WT animals, respectively), whereas reductions in these parameters were caused by both 10 × T3 (29% and 31%, respectively) and 20 × T3 (48% and 47% respectively) in β2-AR−/− males. In addition, treatment with 20 × T3 promoted an increase in Tb.Sp (53%) only in β2-AR−/− animals (Fig. 2J). These findings show that lack of β2-AR signaling also modifies the responses of the femoral trabecular bone to thyrotoxicosis in males. In addition, they also show that sex not only modifies the responses of the trabecular bone to thyrotoxicosis but also modifies the TH-SNS interactions to control trabecular bone microarchitecture in the femur. These findings are summarized in Table 1.
Effects of Supraphysiological Doses of T3 on Bone of Wild-Type and β2-AR−/− Mice
The trabecular and cortical parameters and the biomechanical properties of long bones were determined by μCT analysis and by the three-point bending test, respectively. The
μCT parameters: BV/TV, bone volume/total volume; Ct.Ar, cortical bone area; Ct.Th, cortical thickness; Ec.Pm, endocortical perimeter; Ma.Ar, medullary area; Ps.Pm, periosteum perimeter; Tb.N; trabecular number; Tb.Pf, trabecular pattern factor; Tb.Sp, trabecular separation; Tb.Th, trabecular thickness; Tt.Ar, total tissue area.
Biomechanical parameter: ML, maximum load; Res, resilience; Stif, stiffness; Ten, Tenacity.
BW, body weight; Ec.MAR, endocortical mineral apposition rate; μCT, micro-computed tomography; n.d., no difference; n.m., not measured; RANK-RANKL-OPG, receptor activator of nuclear factor-kappa-B-RANK ligand-osteoprotegerin; WT, wild-type.
Lack of β2-AR modifies the response of cortical bone to thyrotoxicosis
The evaluation of female mice showed that there were no differences in cortical microarchitecture of the femur between saline-treated WT and β2-AR−/−
females. Only

Effect of thyrotoxicosis on cortical microarchitecture of the femur in WT and β2-AR
−/−
mice. (
These findings reveal that the femoral cortical bone of β2-AR−/− female mice is resistant to the detrimental effects of thyrotoxicosis, suggesting that β2-AR signaling has a role in the mechanisms by which T3 promotes endocortical and intracortical bone resorption in the femur. This resistance is further supported by the significant differences between WT +20 × T3 and β2-AR−/− +20 × T3 females in Ec.Pm, Ct.Th, and Ct.Po-2D of the femur (4.1-fold lower, 14% higher, and 3.7-fold lower in KO vs. WT femurs, respectively).
In males, there were no differences in cortical bone parameters between WT and KO saline-treated mice (Fig. 3I–P). We observed that treatment with 10 × T3 promoted an increase in
These findings reveal that the femoral cortical bone of β2-AR−/− male mice is also resistant to the detrimental effects of thyrotoxicosis. We could also observe that 10 × T3 treatment caused a significant increase in Ct.Po (3D) only in WT males (54 times), and, again, not in KO animals (Fig. 3P). A 4.4-fold higher Ct.Po (3D) was observed in WT +10 × T3 versus β2-AR−/− +10 × T3 mice (Fig. 3P), further revealing the resistance of β2-AR−/− males to the deleterious effects of thyrotoxicosis on the cortical bone. These findings are summarized in Table 1.
Lack of β2-AR modifies the effect of thyrotoxicosis on Ec.MAR
Considering the μCT findings suggesting that loss of β2-AR signaling impairs the response of endocortical bone surface to thyrotoxicosis, we evaluated, by bone histomorphometry, the effect of 20 × T3 treatment, for females, and 10 × T3 treatment, for males, on the
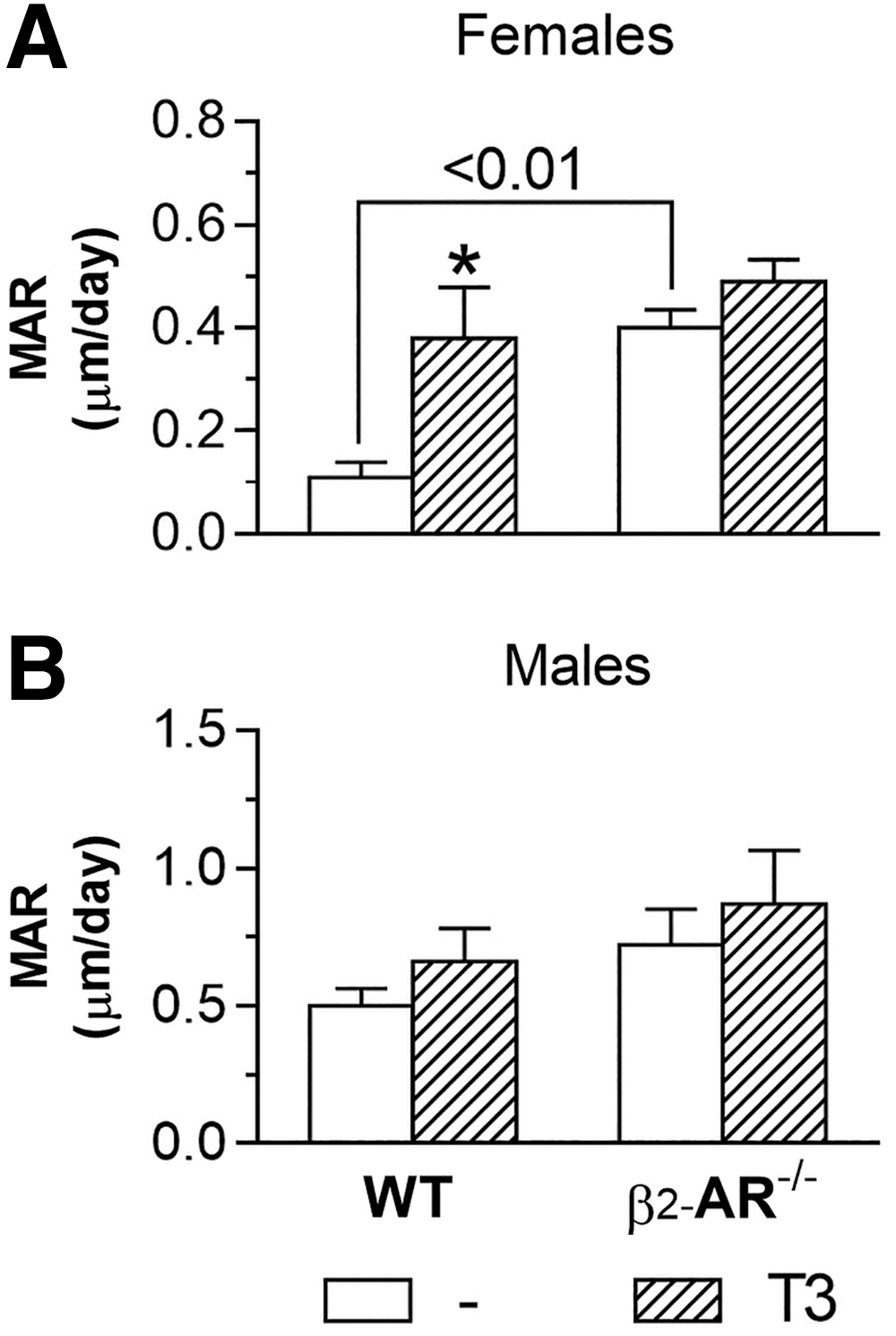
Effect of thyrotoxicosis on
In contrast to what was observed in females,
Lack of β2-AR modifies the effects of thyrotoxicosis on the biomechanical parameters of the femur
Through the three-point bending test, we observed that treatment with 20 × T3 promoted a significant reduction in all the biomechanical parameters evaluated in the femur of WT female mice (Fig. 5A–D), which reveals an increased risk of fracture. A reduction of 35%, 65%, 19%, and 82% was observed in the maximum load, tenacity, stiffness, and resilience of the femur, respectively, in WT female mice treated with 20 × T3 versus WT females treated with saline (Fig. 5A–D, respectively). Treatment with 10 × T3 only negatively affected the resilience in the femur of WT females (a reduction of 64%), in relation to the saline-treated WT females (Fig. 5D).
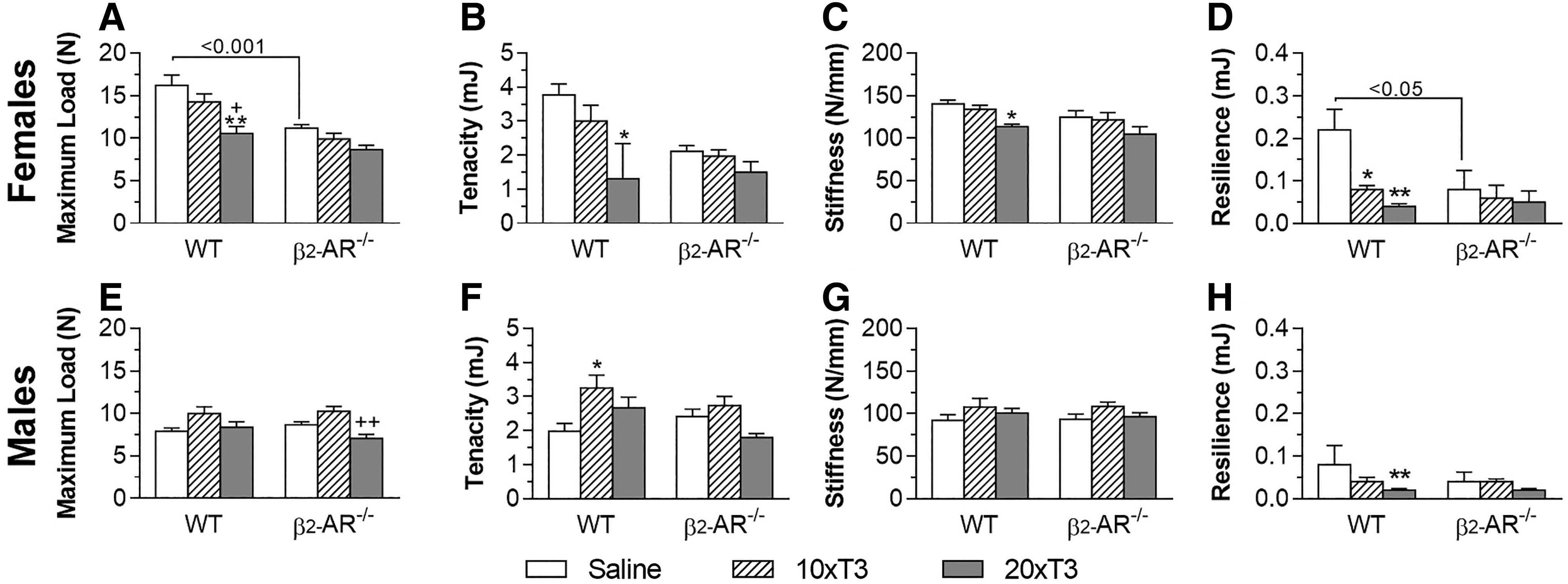
Effect of thyrotoxicosis on biomechanical parameters of the femur in WT and β2-AR
−/−
mice. (
On the other hand, treatment with 10 × T3 or 20 × T3 did not promote any effect on these parameters in KO females, revealing a lower sensibility of these animals to the deleterious effects of thyrotoxicosis on the biomechanical parameters of the femur and, ultimately, on the femoral resistance to fracture. Unexpectedly, we observed, a lower maximum load (31%) and resilience (64%) in the femur of saline-treated β2-AR−/− females versus saline-treated WT females (Fig. 5A, D), which was not observed in males.
Unlike females, WT males were insensitive to the deleterious effects of thyrotoxicosis on the biomechanical parameters of the femur (Fig. 5). Both 10 × T3 and 20 × T3 did not negatively affect any of the biomechanical parameters evaluated in WT males and KO males as well. On the other hand, 10 × T3 promoted a 64% increase in the femoral tenacity in WT animals (Fig. 5F). These results are summarized in Table 1.
Lack of β2-AR modifies the effect of thyrotoxicosis on the gene expression of the RANKL-RANK-OPG system
Considering that activation of β2-AR signaling in osteoblasts enhances bone resorption by increasing RANKL expression in these cells (11), we investigated the effect of thyrotoxicosis on gene expression of the components of the RANKL-RANK-OPG pathway in the femur of WT and β2-AR−/− animals. In the femur of female mice, RANK and OPG mRNA levels were significantly lower in saline-treated β2-AR−/− mice (48% and 60%, respectively), as compared with WT female mice (Fig. 6A, C). In contrast, in the femur of males (Fig. 6E–G), RANK, RANKL, and OPG mRNA levels were significantly higher in saline-treated β2-AR−/− mice versus WT mice (4.2-, 6.2-, and 9.3-fold, respectively).
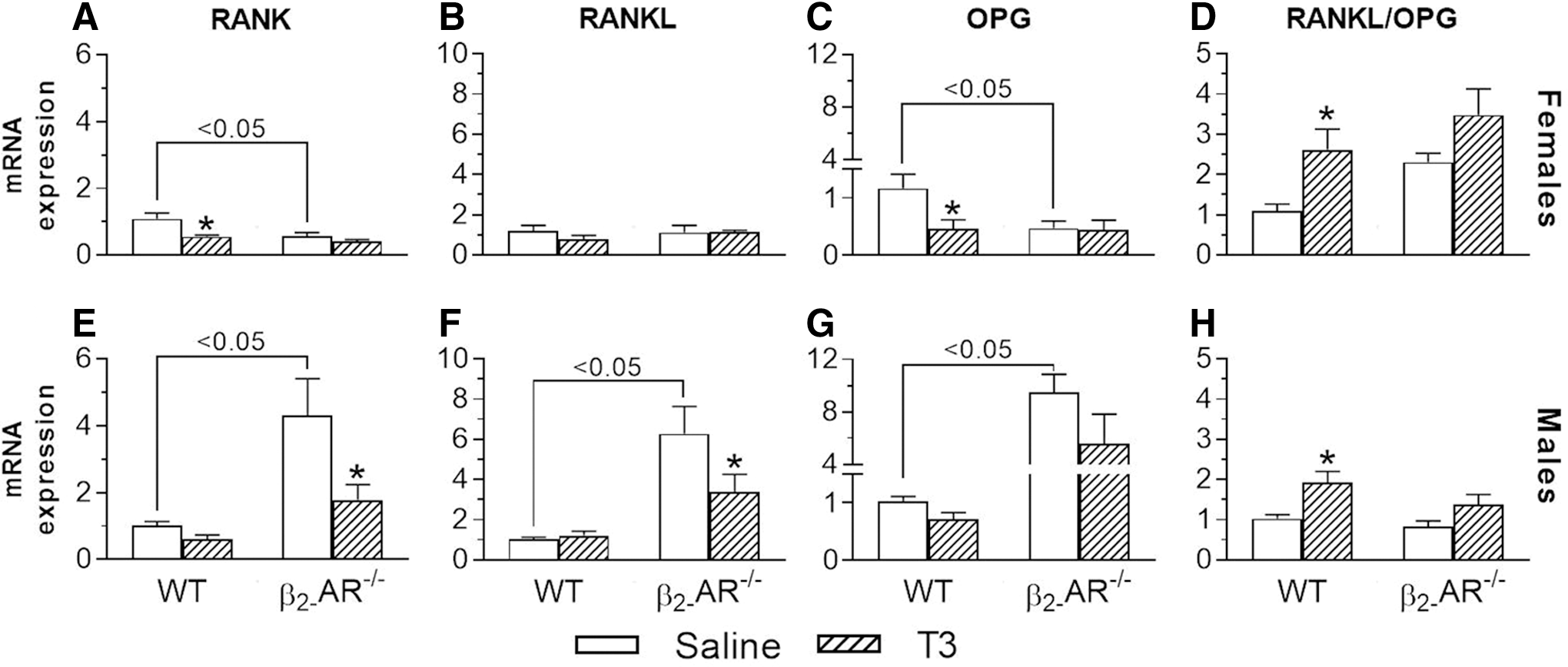
Effect of thyrotoxicosis on the mRNA expression of the RANK-RANKL-OPG system in the femur of WT and β2-AR
−/−
mice. (
In females, T3 treatment decreased RANK expression (49%) in WT but not in β2-AR−/− mice; whereas, in males, T3 treatment decreased RANK expression (59%) in β2-AR−/− but not in WT mice (Fig. 6A, E). RANKL was not affected by T3 treatment in females, but it was decreased by T3 treatment by 46% in β2-AR−/− mice (Fig. 6F). In females, T3 treatment significantly decreased OPG mRNA expression by 60% in WT but not in β2-AR−/− mice (Fig. 6C), whereas T3 tended to decrease OPG expression in both WT and β2-AR−/− male mice (30% and 40%, respectively) (Fig. 6G). Interestingly, in females and males (Fig. 6D, H), T3 treatment significantly increased the RANKL/OPG ratio in WT (2.4-fold for females and 1.9-fold for males) but this was not the case in β2-AR−/− mice. These results are summarized in Table 1.
T3 treatment prevents E2-stimulated cAMP accumulation in osteoblasts
To investigate potential underlying cellular mechanisms through which TH interacts with β2-AR signaling to control bone physiology and to investigate whether estrogen affects a possible TH/β2-AR signaling interaction in osteoblasts, we measured the effect of CL, a β2-AR specific agonist, on cAMP accumulation in osteoblastic cells (MC3T3-E1 cells) treated with T3 and/or E2 for 72 hours. T3 tended to decrease cAMP content (6.5%), whereas E2 and CL similarly increased cAMP accumulation (22% and 16%, respectively) in MC3T3-E1 cells (Fig. 7). E2 did not modify the effect of CL or T3 on cAMP content; on the other hand, T3 completely blocked E2-stimulated cAMP accumulation and had no effect on the CL-induced cAMP levels.
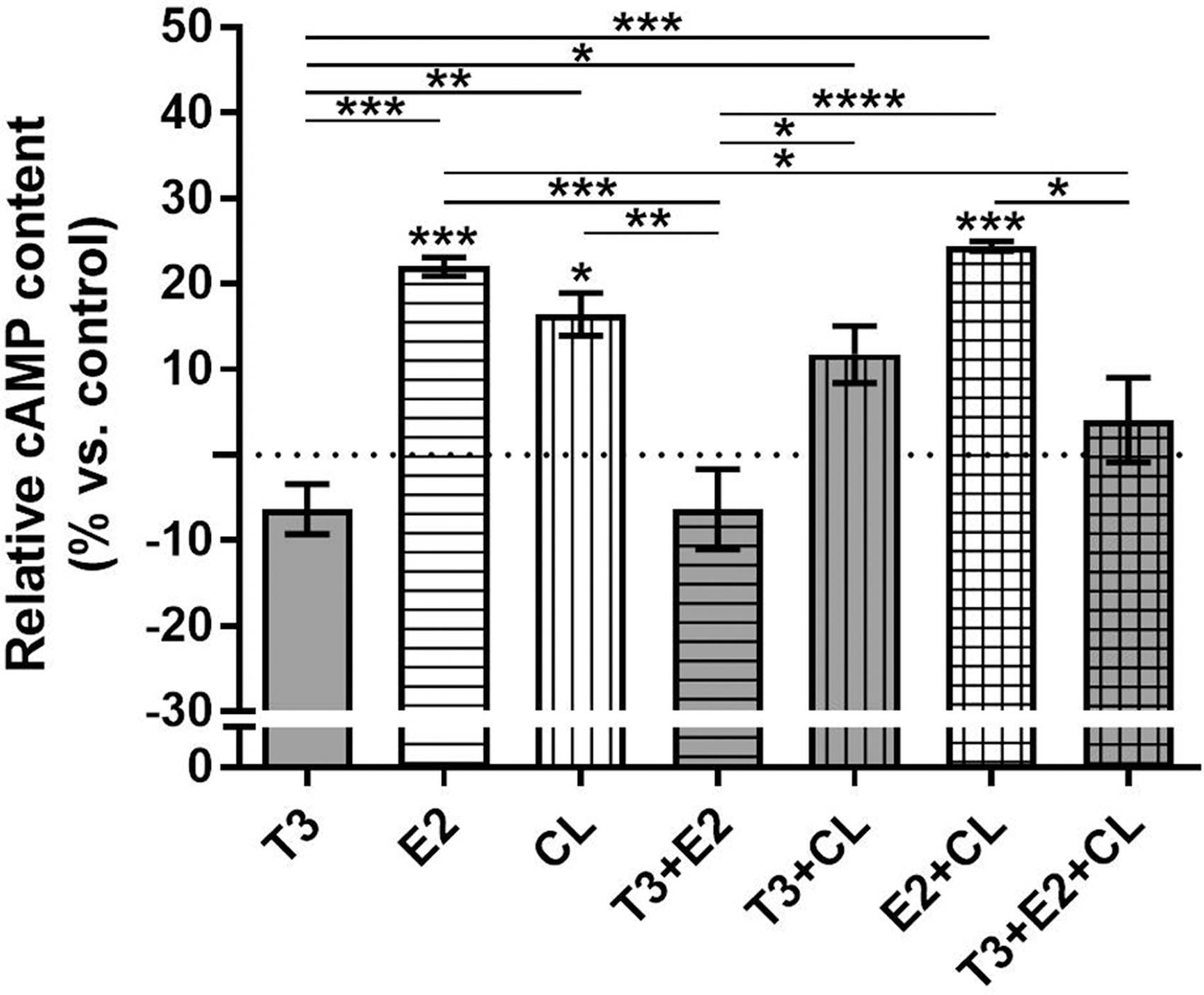
Effect of T3, 17β-Estradiol, and/or clembuterol on cAMP content in osteoblastic cells. MC3T3-E1 cells were treated with T3 (10−8 M) and/or 17β-Estradiol 3-benzoate (E2; 10−8 M) for 72 hours, or left untreated. During the last 30 minutes of T3 and/or E2 treatment, cells were also treated with 10−4 M CL, a β2-AR specific agonist. Cells were then collected, and the cAMP content was determined by immunoassay and expressed as relative content (%) versus control (untreated cells). Dotted line indicates the control mean. Bars show mean ± SEM of triplicate cultures. Statistical analysis was performed by one-way ANOVA followed by Tukey's test. Asterisks above the bars indicate p-values versus control; the other asterisks indicate p-values for comparisons indicated by the lines beneath. *p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.001. CL, clenbuterol.
Discussion
This study investigated whether a chronic state of thyrotoxicosis (12 weeks) negatively affects the femoral bone, a skeletal site that is very sensitive to TH (35,36), through a mechanism that involves the β2-AR signaling pathway. We found that TH excess caused deleterious effects on the femoral trabecular microarchitecture in WT females, while some of these effects were milder or nonexistent in the β2-AR−/− animals, demonstrating that loss of β2-AR signaling in female mice is associated with a lower sensibility of the trabecular bone to the deleterious effects of thyrotoxicosis. On the other hand, in males, the lack of β2-AR signaling made the trabecular bone slightly more sensitive to the deleterious effects of toxic doses of T3.
Interestingly, WT males were less susceptible than females to the effects of thyrotoxicosis on the femoral trabecular bone. The fact that loss of β2-AR signaling modifies the trabecular bone response to TH, in both females and males, reveals an interaction between TH and the SNS that involves β2-AR signaling to regulate trabecular bone microarchitecture. On the other hand, the contrasting sensitivity of the trabecular bone to thyrotoxicosis in female and male β2-AR−/− mice suggests that sex factors play a role in the interaction of TH with the SNS, via β2-AR signaling, to regulate bone morpho-physiology.
With regard to the femoral cortical bone, we observed that T3 excess caused an increase in endocortical perimeter (
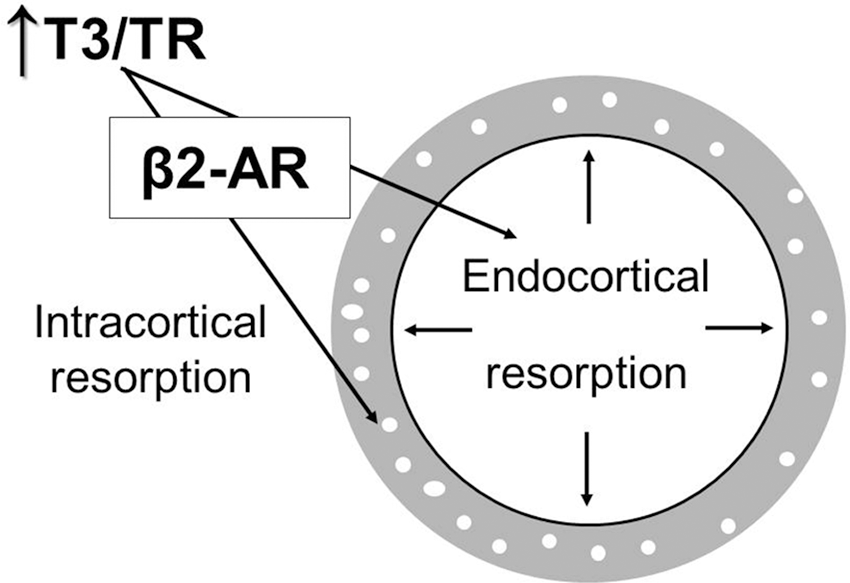
Schematic representation of the interaction between thyroid hormone and β2-AR signaling in the cortical bone. In thyrotoxicosis, T3 interacts with its receptor (TR) in the bone tissue to promote intracortical bone resorption, which results in higher cortical porosity. The higher T3/TR interaction in thyrotoxicosis also induces endocortical (endosteal) bone resorption, leading to increases in endosteal perimeter and medullary area. T3 activation of both endocortical and intracortical bone resorption occurs in a mechanism that depends on the presence of β2-AR signaling pathways, revealing that TH interacts with the SNS to promote these processes. SNS, sympathetic nervous system; TH, thyroid hormones.
The combined increases in Ec.Pm, Ma.AR, and Ec.MAR, observed in WT but not in KO females, strongly suggest that β2-signaling is necessary for the thyrotoxicosis-induced unbalance between bone formation and resorption in the endocortical bone in females, a known effect of thyrotoxicosis (1,37). T3 excess also promoted an increase in
The increased
In accordance with the μCT findings, the three-point bending test showed that thyrotoxicosis caused significant reductions in the femoral resistance to fracture in WT female mice (decreased maximum load, tenacity, stiffness, and resilience), but not in KO females, once again revealing a lower susceptibility of these β2-AR-deficient animals to the deleterious effects of thyrotoxicosis on bone tissue. These findings are substantiated by a previous study where propranolol prevented the detrimental effects of thyrotoxicosis on femoral bone mineral density and femoral resistance to fracture in B6 female mice (38). Altogether, these findings further support a TH-SNS interaction, involving β-AR signaling, to regulate bone mass and resistance to fracture. On the other hand, in this study, male mice (both WT and KO) were resistant to the deleterious effects of T3 excess on the biomechanical parameters of the femur (Table 1), further indicating the importance of sex factors in the actions of TH in the skeleton.
It is noteworthy that we have not found an HBM phenotype in the femur of β2-AR−/− mice, which contrasts the data published by Elefteriou et al. (11). A possible explanation for these discrepancies is the different genetic background of the KO animals used in these studies. While the β2-AR−/− mouse model studied here is in a FVB/N background (26), the KO animals analyzed by Elefteriou et al. were in a combined 129SvJ and FVB/N background (11,39). Interestingly, Pierroz et al. (34) showed that β2-AR KO mice in a mixed 129SvJ, FVB/N, and C57BL/6J genetic background have higher trabecular bone mass in the vertebrae and lower trabecular bone mass in the femur. These observations support the concept that the divergent skeletal phenotypes of the β2-AR KO mouse models are determined by the different genetic background of each mouse strain.
In this regard, one limitation of this study is that the WT animals are not the littermates of β2-AR heterozygous mattings, which would provide an optimal control. On the other hand, they are FVB/N inbred mice that have been maintained in the same environment for many years. Therefore, we believe that the WT animals used in this study are a reliable control for our studies.
In an attempt to get insights into the underlying mechanisms through which TH interacts with β2-AR signaling to control bone physiology, we investigated the effects of T3 treatment on the mRNA expression of the components of the RANK-RANKL-OPG system (Table 1). RANKL is an osteoblastic factor that binds its receptor RANK in osteoclast precursor cells and in mature osteoblasts, which activates osteoclastogenesis and osteoclastic activity, respectively, increasing bone resorption (40). OPG, another osteoblast-derived factor, binds RANKL, impairing the RANK-RANKL interaction, which limits bone resorption (41). Previous in vitro studies have shown that activation of β2-AR signaling in osteoblasts induces RANKL expression; and that osteoblasts derived from β2-AR−/− mice present lower RANKL mRNA expression, suggesting that SNS activation increases bone resorption via RANKL activation (11).
In contrast to these in vitro findings, we found RANKL mRNA expression to be increased in the whole femur of β2-AR−/− male mice (vs. WT) and equal to WT in β2-AR−/− female mice. Interestingly, in females, RANK and OPG mRNA levels were shown to be decreased in β2-AR−/− versus WT mice, and the opposite was seen in males, indicating that sex differences also involve the RANK-RANKL-OPG system. In view of that, T3 effects on the mRNA expression of RANK, RANKL, and OPG were different between males and females as well. It is noteworthy that, in females, T3 treatment decreased OPG expression by 60% in the femur of WT but not in β2-AR−/− mice, which may partially explain the lower susceptibility of female KO mice to the osteopenic effects of thyrotoxicosis.
Accordingly, supraphysiological doses of T3 were previously shown to decrease OPG mRNA expression in the femur of WT female mice (in B6 background), but not in the femur of α2A/2C-AR−/− and α2C-AR−/− female mice (15,25). In addition, in males and females, we found that T3 treatment significantly increased the RANKL/OPG ratio in the femur of WT mice, but not in β2-AR−/− mice. Interestingly, in a former study, T3 was also not able to increase the RANKL/OPG ratio in α2C-AR−/− female mice but it did so in the respective WT controls (B6 mice) (25). Altogether, these findings suggest that the osteopenic effects of T3 in the femur involve the RANKL-RANK-OPG system in a mechanism that involves β2-AR signaling in a sex-dependent way.
To investigate possible cellular mechanisms through which TH interacts with β2-AR signaling to control bone physiology and to investigate whether estrogen affects a possible TH/β2-AR signaling interaction in osteoblasts, we analyzed the effect of CL, a β2-AR specific agonist, on cAMP accumulation in osteoblastic (MC3T3-E1) cells treated with T3 and/or 17β estradiol (E2). We found that T3 had no significant effect on cAMP content, whereas CL and E2 similarly increased cAMP accumulation in MC3T3-E1 cells. Accordingly, positive effects of E2 on cAMP production in osteoblasts (42 –44) and decreased cAMP levels in E2-depleted osteoblasts have been previously documented (42).
It is noteworthy that high levels of intracellular cAMP have been associated with increased osteoblastic proliferation and differentiation (45 –47), trabecular thickness (48), and bone formation rate in rodents (49). In addition, there is evidence that active osteoblasts present high levels of cAMP (50 –52). Interestingly, E2 did not modify the effect of CL or T3 on cAMP content. On the other hand, T3 completely blocked the E2-stimulated cAMP accumulation and had no effect on the CL-induced cAMP contents. These findings show that there is no interaction between T3 and β2-AR signaling at the level of cAMP accumulation in osteoblasts, but that T3 critically modifies the effect of E2 on cAMP accumulation in these cells. These findings suggest that some of the osteopenic effects of T3 on the bone may involve E2/cAMP signaling in osteoblasts, which may partially explain the sex dependency of T3 effects on the bone.
In summary, this study reveals that (i) TH involves β2-AR signaling to promote its negative effects on the trabecular and cortical compartments of the femur in female mice, and on the femoral cortical bone in male mice; (ii) in general, trabecular and cortical bone compartments of the femur are less sensitive to thyrotoxicosis in male mice than in female mice; and (iii) sex is an important modifier of TH action of the TH-SNS interactions that impact bone morpho-physiology.
In conclusion, these novel findings show that β2-AR signaling has an important role in the mechanism by which T3 excess promotes its deleterious effects on bone microarchitecture, bone remodeling, and bone resistance to fracture, especially in females and the cortical bone. Therefore, the use of beta-blockers in thyrotoxicosis may not only contribute toward minimizing the adrenergic symptoms that are characteristic of this condition, such as tachycardia, tremor, and anxiety (53), but also contribute toward protecting bone tissue from the deleterious effects of thyrotoxicosis, particularly in females. Nevertheless, it is important to consider that, although β2-AR signaling likely plays a role in the actions of TH on the bone, this is most likely not the sole mechanism.
Footnotes
Acknowledgments
The authors are grateful to the Institute of Biomedical Sciences, University of São Paulo, for providing technical support to this study, and the Foundation for Research Support of the State of São Paulo (FAPESP), for providing financial support to this study (Grants No. 2015/10671-5 and 2015/12554-6).
Author Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Figure S1