
Abstract
Background:
Cyclic peptides derived from some cylindrical loops of the leucine-rich repeat domain (LRD) of the thyrotropin receptor (TSHR) have been shown to treat disease manifestations in a mouse model of Graves' disease during a long-term protocol of four-weekly immunizations with adenovirus coding for the TSHR A-subunit (Ad-TSHR289).
Methods:
In a follow-up study, two additional cyclic peptides were tested, which were shortened in order to obtain additional information on the minimally involved epitopes and to enable easier production conditions. In addition, a linear peptide was tested, which mimics parts of three loops of the native TSHR LRD structure, and is potentially able to block the discontinuous epitopes of anti-TSHR antibodies.
Results:
The novel peptides markedly reduced thyroid size, serum thyroxine levels, retro-orbital fibrosis, and tachycardia in Ad-TSHR289-immunized mice. In immunologically naïve mice, administration of the peptides did not induce any immune response.
Conclusions:
In summary, novel cyclic peptides mitigate many clinical findings in a mouse model of established Graves' disease and orbitopathy, and may therefore provide an additional therapeutic option compared to existing drugs or interventions.
Introduction
An autoimmune attack targeting the thyrotropin receptor (TSHR) in the thyroid gland results in Graves' disease (GD) and hyperthyroidism (1), with an annual incidence of 15–80/100,000 people throughout the world. The condition is generally mediated by autoantibodies, but also involves subsets of immune cells (1), although neonatal GD occurs following placental transfer of maternal immunoglobulin G (IgG) without immune cells, and many GD mouse models do not clearly show lymphocyte infiltrates in the thyroid. The existing treatment options are characterized by relatively high relapse rates and significant side-effect profiles (2). Still, treatment is necessary, since untreated GD incurs significantly increases morbidity and mortality (3).
Treatment of orbitopathy is a relevant issue. Ophthalmopathy occurs in almost 50% of all GD patients—up to 16/100,000 women per year (4)—and must frequently be treated by giving intravenous (i.v.) corticoids over many weeks with a high risk of relevant side effects (5).
An alternative therapeutic approach may be offered by specific immune therapies, which have been used for the treatment of allergic autoimmune conditions for >100 years (reviewed, e.g., in Larché and Wraith and Soyka et al.) (6,7). Since treatment with broad-range immunosuppressive drugs may cause serious side effects, allergen-specific therapies have been conceived to induce tolerance in a variety of related conditions. As a novel option, immunogen-mimicking cyclic peptides have been developed for the treatment of anti-TSHR antibody-mediated GD (8). In contrast to some existing specific immune therapies, a primary target of these cyclic peptides is to reduce antibody-secreting immune cells.
A long-term disease model for human GD was successfully established using nine immunizations of recombinant adenovirus expressing the extracellular A subunit of the TSHR (9,10) to boost antibody production permanently in mice. Monthly i.v. administration of cyclic peptides, which were derived from the tertiary structure of the TSHR A-subunit, resulted in strong potency to treat hyperthyroidism and its symptoms in TSHR-immunized diseased mice and a clear decrease of retro-orbital fibrosis (8).
In this follow-up study, further peptides were investigated, which were designed upon close analysis of available published structural data of the interaction of TSHR with the GD patient-derived antibody M22 (11). The peptides are derived from the major TSHR epitopes, which include the exclusive M22 antibody-binding amino acids (aa) (11), and are outlined in Table 1. First, the study investigated the cyclic peptide 19, which was derived from the first TSHR loop, and includes aa residues EED, which are considered to be crucial for the interaction of the TSHR and M22 at positions 34–36 (11). A multi-epitope peptide 12 was also designed as a discontinuous epitope reflecting the binding of anti-TSHR antibodies to the TSHR leucine-rich domain (LRD) consecutive loops 5, 6, and 7, and includes the crucial aa K129 (11). Lastly, a shortened 13-meric cyclic version of the previously described peptide 836 (8) was included, which includes the interaction aa N208 (11).
TSHR-Derived Peptides
TSHR amino acid sequences from which peptides used in the study were derived. After synthesis, peptide 836 13-mer was cyclized head-to-tail, and peptide 19 was cyclized by spontaneous formation of disulfide bonds. Peptide 12 lines up several epitopes, which are considered to be relevant for the interaction of anti-TSHR antibodies with the TSHR, which were fused by PG linkers. The first and third loops were derived from the forward peptide sequence; the second was derived from the respective reverse peptide sequence.
TSHR, thyrotropin receptor.
The study shows that repeated i.v. administrations of these novel cyclic peptides, which were derived from the eighth or the first cylindrical loop of the LRD of the TSHR, successfully treated thyroid disease in Ad-TSHR-immunized animals, resulting in reduced thyroid size and normalized thyroxine (T4) levels, starting only eight weeks after initiation of peptide therapy. It also shows that retro-orbital fibrosis was mitigated, suggesting a positive effect on Graves' orbitopathy. Tachycardia and cardiac hypertrophy were consistently reduced by the novel therapy. Several of these effects were also exerted by the novel multi-epitope peptide 12.
Methods
Recombinant adenovirus
The DNA sequence coding for the first 289 aa of the human TSHR (12,13) was cloned into the Microbix Admax™ adenovirus expression system, as described before (9).
HEK293A cells were used to propagate until first viral plaques became visible. This system results in recombinant replication-deficient E1 and E3-deficient adenovirus type 5. A control adenovirus containing only the reporter gene green fluorescent protein (Ad-GFP) was amplified and purified in the same manner.
Synthesis of cyclic peptides
Peptides were designed, as outlined in Table 1. The peptides are derived from the major TSHR LRD epitopes, which include the exclusive M22 antibody-binding aa (11). Specifically, the sequence of the 13-meric peptide 836 is a shortened version of the previously published 24-meric 836 (8), which replicates the eighth TSHR LRD cylindrical loop. This 13-meric variant, as well as the previously described peptide 836, includes the interaction aa N208 (11). Peptide 19 is a shortened version of peptide 829, which was designed in analogy to the first loop of the TSHR LRD, and includes aa residues EED, which are considered crucial for the interaction of the TSHR and M22 at positions 34–36 (11). A multi-epitope peptide 12 was also designed, which includes the crucial aa K129 (11) and other sequences in close proximity. As one example of a control peptide, results for peptide 14 are shown, which was derived from the first and second TSHR loops: aa 37–43 oriented in forward sense, followed by aa 55–62 in reverse order (FRVTSKDPGTEILKLTQPG), but which did not include crucial interaction aa (11). This peptide is shown as one example for a control, since inclusion of further control groups into the figures would compromise their legibility. Further controls were included in the studies but did not yield significantly different results from this control when analyzable. All peptides were fully soluble in 0.9% NaCl.
The peptides were synthesized by Biosyntan Berlin according to described protocols of fluorenylmethoxycarbonyl (FMOC) resin-based aa chain elongation, and subsequent head-to-tail cyclization. FMOC aa or FMOC dipeptide was attached to the 2-chlorotrityl chloride resin (RAPP Polymere GmbH), yielding a loading of 0.30 mmol/g resin. Peptide synthesis was done by a standard cycle of deblocking with 30% piperidine/N,N-dimethylformamide (DMF; 5 + 12 min) and coupling with 3 eq. FMOC aa/O-(7-Azabenzotriazol-1-yl)-N,N,N′,N′-tetramethyluronium-hexafluorophosphate/6 eq. N-methylmorpholine in DMF (double coupling, 2 × 30 min). After cleavage from the resin by 20% hexafluoroisopropanol/DCM (2 × 20 min), the isolated crude peptide was cyclized by 1.5 eq. 7-azabenzotriazol-1-yloxy)tripyrrolidinophosphonium hexafluorophosphate/3 eq. diisopropylethylamine in DMF overnight, the solvent evaporated, and the crude peptide deblocked by trifluoroacetic acid/water/thioanisol (95:5:3) for two hours. Then, the peptides were purified up to 95% by means of high-performance liquid chromatography and analyzed by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry. This quality control reconfirmed that aa had been correctly included into the peptides, and their cyclization was evident from the experimental determination versus theoretical prediction of molecular weights: As examples, values of observed versus predicted molecular weight were 1446.2 versus 1445.7 for peptide 13-mer 836, and 1364.6 versus 1363.5 for peptide 19.
Studies in immunized mice
Female BALB/c mice were delivered from Charles River, and were adapted for at least one week before starting the experiments at the age of six weeks. Animals were kept under standard housing conditions (23 ± 2°C, 55 ± 10% relative humidity) in groups of 10 mice in GR1800DD cages (Tecniplast®). All animal experiments were approved by the local animal welfare authority and ethics committee at the Regierung von Oberbayern (Government of Upper Bavaria; no. 55.2-1-54-2531-25-12), and were carried out in accordance with the World Medical Association (Declaration of Helsinki) and the European Commission guidelines (Directive 2010/63/EU. All guidelines for the care of animals were respected.
Mice received 1010 pfu of adenovirus carrying the A-subunit of the TSHR gene. In addition, age-matched immunologically naïve mice that were not immunized and were not treated (n = 10) were studied for comparison. For immunization, mice were anesthetized with isoflurane (introduction 5%, maintenance 1.5–2%) and placed on a heating pad. The adenovirus was injected into the left and right femoral muscles at a volume of 25 μL each.
For blood withdrawal, mice were moved to a restrainer. Blood (100 μL) was drawn from the left or right tail vein with a 27G needle and centrifuged at 2400 g for 15 minutes at room temperature (RT) before being stored at −20°C. At the end of the study (before euthanasia), blood was drawn intracardially in deep anesthesia (170 mg/kg ketamine +17 mg/kg xylazine) with a 1 mL syringe and a 24G needle, and treated as mentioned above.
The study protocol used 3 × three-weekly immunizations (“initiation”), followed by a “maintenance” phase with further regular four-weekly boosts until the ninth immunization, as described before (9). Strict inclusion and exclusion criteria were defined before starting the study. If T4 values in Ad-TSHR289-immunized mice were at least 25% higher at week 11 compared to baseline measurement at week 0, mice were included for further study. Consequently, 25% of immunized mice were excluded before randomization because they did not meet this threshold. Also, mean T4 values and baseline heart rates of all groups had to be consistently altered. Mice were randomly assigned to therapeutic groups, allocations were concealed, and veterinarians and all other staff who handled the animals or who analyzed the results were strictly blinded to the treatment groups. Therapy (either peptides, all dissolved in 0.9% NaCl without further additives, or 0.9% NaCl vehicle control) was given one week after the fourth immunization by i.v. injection into the tail vein, and then continued at four-weekly intervals, as described before (8). A detailed immunization schedule is shown in Figure 1A. All animals that were excluded after randomization are reported in the results.
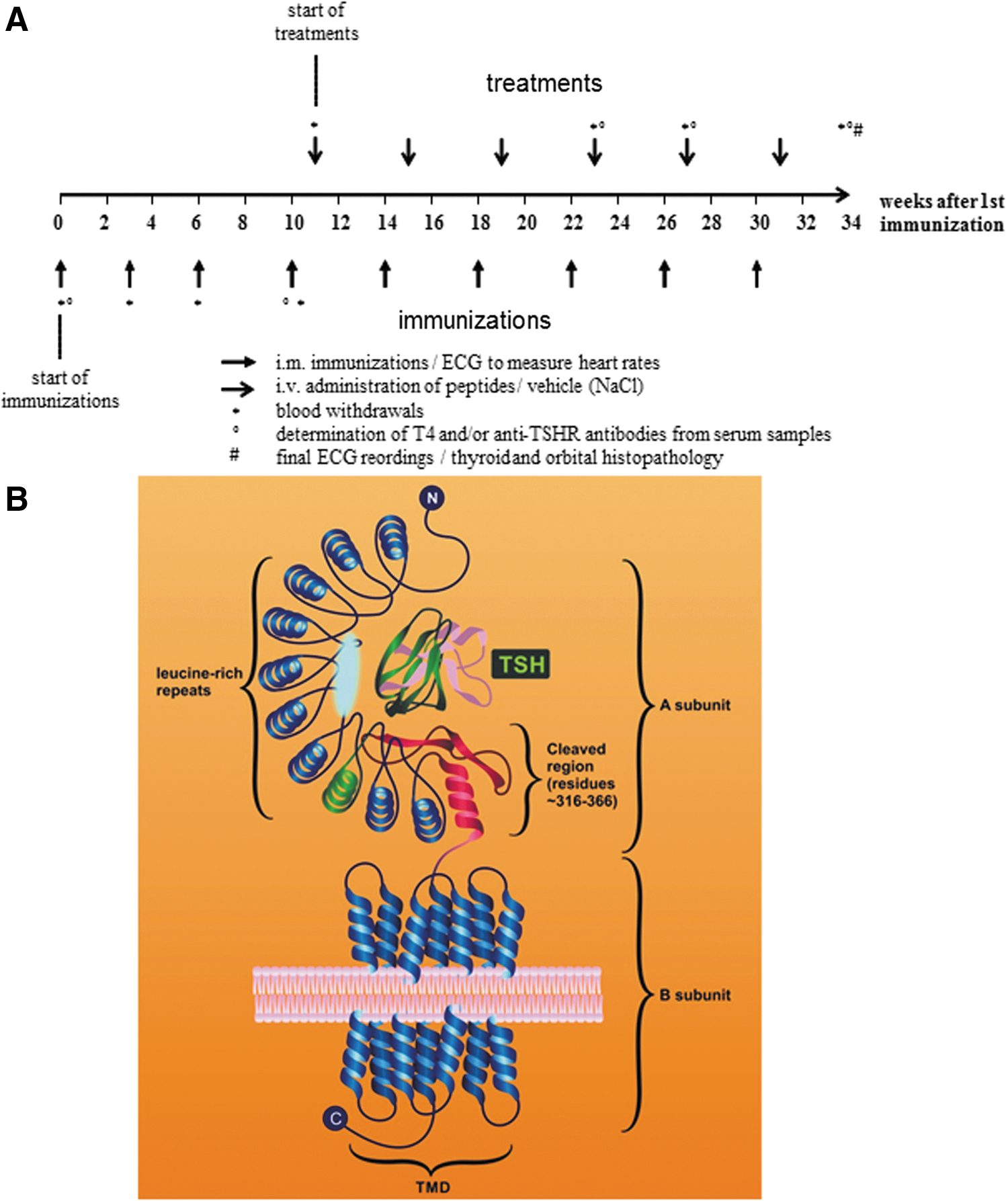
When mice were subjected to anesthesia for immunization, their heart rate was monitored by an electrocardiogram (ECG; ECG amplifier module, Harvard Apparatus, Hugo Sachs Electronics) and recorded with a special software, which allows the heart rate to be determined from the ECG reading (Haemodyn, Hugo Sachs Electronics). An ECG was also performed in anesthesia before the animals were euthanized for histological exam at the end of the study.
Total T4 was measured by using an immunoassay kit (#T4044T-100; Calbiotech, Inc.) in duplicate.
Histological analysis
After euthanasia, dissection of the thyroid glands was performed under a stereomicroscope, as described before (8). The orbital sections were viewed with a 4 × objective lens (Axioscope; Zeiss), captured with an Axiovision digital cam system and recorded with 2560 × 1920 pixel resolution. Focus was adjusted for each new field, but light conditions were kept identical. All sections were evaluated in a blinded fashion.
Fibrotic areas in the extra-orbital adipose tissue and extra-orbital muscle regions were identified by their green color. Digitized image analysis of green color pixels was carried out using the luminescence tool of Adobe Photoshop vCS5 extended on the basis of pixel areas. In order to correct for possible bias between stainings, the green staining intensity of the respective orbital bone in each section was taken as an internal standard for each measurement. Accordingly, all fibrotic tissue throughout a whole orbital section was quantified, and the results of all sections were added in the end to yield a total fibrosis volume (mm3) of each investigated orbit (taking into account the 0.63 mm interval thicknesses between sections).
Measurements in mouse sera
Anti-TSHR autoantibody titers and the potency of antibodies to stimulate TSHR-dependent cyclic adenosine monophosphate (cAMP) levels in test cells were determined before the start of immunization (baseline value), 56, 133, and 189 days after the first immunization, and at the end of the experiment. To this end, two different assays were used:
(i) A third-generation assay: Antibodies against TSHR were detected by a commercially available third-generation enzyme immunoassay provided by RSR Ltd., in which the human Graves' patient-derived M22 monoclonal antibody and serum antibodies compete for binding sites on immobilized TSHR. The assay is also used in Roche's Cobas® assay (04388790) for electrochemiluminescence assay with minor modifications. The assay was performed using 30 μL 1:10 (phosphate-buffered saline [PBS]) diluted serum in at least double determination according to the manufacturer's instructions.
(ii) Thyroid stimulating antibodies in the serum of hyperthyroid mice were analyzed by measuring cAMP generation in JP2626 Chinese hamster ovary (CHO) cells expressing the human TSHR (kindly provided by Dr. Gilbert Vassart, Brussels, Belgium). CHO cells were seeded onto 96-well plates (30,000 cells per well) and incubated for 24 hours in Dulbecco's modified Eagle's medium (DMEM; Invitrogen) containing 2% fetal calf serum. Then, DMEM was removed, and mice serum was diluted 1:8 in 40 μL: Hank's Balanced Salt Solution buffer (20 mM Hepes, 1.26 mM CaCl2, 5.33 mM KCl, 0.44 mM KH2PO4, 0.5 mM MgCl2, 0.4 mM MgSO4, 4.2 mM NaHCO3, 5.6 mM glucose, and 222 mM sucrose, pH 7.2) supplemented with 1.5% bovine serum albumin and 0.5 mM isobutyl-1-methylxanthine (Sigma–Aldrich) and added to each well. After incubation for 2.5 hours at 37°C, the cAMP release in the medium was measured in duplicate by a competitive immunoassay enzyme-linked immunosorbent assay (ELISA; #EMSCAMPL; Thermo Fisher Scientific).
In vitro studies (third anti-TSHR antibody assay)
In order to investigate the direct binding of peptides to anti-TSHR antibodies in vitro, the fusion protein TSHR-Fc was used, which consists of the first 289 aa of the human TSHR (extracellular TSHR A-subunit) followed by a GGR linker and the Fc portion of human IgG2. The cDNA sequence adapted for hamster codon usage was produced synthetically and cloned into the plasmid vector pcDNA5/FRT. This expression vector was transfected into CHO Flp-In cells together with the plasmid pOG44, providing site-directed recombination. After selection of a stably expressing clone, the fusion protein was purified from suspension culture supernatant by Protein G chromatography and dialyzed against PBS. It represents the conformationally intact A domain of the TSHR.
ELISA plates were coated with 0.5 μg/mL TSHR-Fc in coating solution for one hour. All subsequent procedures were performed at RT, and incubations were on a microtiter plate shaker. The coated plates were washed three times with PBST, blocked with 100 μL/well of blocking solution (PBS with 0.1% Tween, 3% milk powder) for one hour, and washed again. In another non-coated plate, polyclonal anti-TSHR antibody-containing mouse sera was diluted 1:400, and incubated (30 minutes) with different concentrations of peptide 19 diluted in PBST.
The blocked ELISA plates were then incubated for one hour with 100 μL of this mix (final volume: 0.25 μL polyclonal mouse anti-TSHR antibodies per well). After washing with PBST, the ELISA plates were incubated with 100 μL/well of anti-mouse IgG detection antibody, and labeled with horseradish peroxidase (POD) for one hour. After washing, POD was detected by incubation with 100 μL/well of TMB substrate until a maximal optical density (OD) of about 1–2 was reached. Finally, the colorimetric reaction was stopped with 100 μL/well stopping solution, and the OD was determined at a wavelength of 450 nm with a reference wavelength of 595 nm with the Tecan Infinite F200 plate reader.
The peptides were also investigated in third-generation assays with minor modifications: 30 μL of the diluted monoclonal M22-Bio antibody was mixed with 30 μL diluted peptide (final concentration of 100 μg/mL in PBS) and added to the TSHR-pre-coated microtiter plate. After an incubation step for two hours at RT, the protocol was continued according to the manufacturer's instructions.
Studies in native mice
BALB/c mice were adapted for at least one week before the start of the experiments at the age of 12 weeks. Animals were kept as described before. This study was approved by the local animal welfare authority and ethics committee at the Regierung von Oberbayern (Government of Upper Bavaria; no. 55.2-1-54-2532.0-32-15) and carried out in accordance with the European Commission guidelines.
Peptides (1 mg/kg body weight) or NaCl vehicle control were given by i.v. injection into the tail vein of these naïve mice (n = 6 per group), and then continued at four-weekly intervals for six months, as described before for the therapeutic study in immunized mice. Serum samples were taken at identical intervals.
Statistics
Differences between the groups were analyzed by analysis of variance (ANOVA) for comparison between groups using IBM SPSS Statistics for Windows v19 (IBM Corp.), followed by least significant difference post hoc testing or Student's t-test where appropriate. For comparison of values at various times within one group, repeated-measures ANOVA was used where appropriate.
Results
Selection of cyclic peptides and of a linear multi-epitope peptide
Cyclic peptides were synthesized whose aa sequences and tertiary structures were derived from the TSHR LRD (Table 1). The short peptide 836 is a 13-meric cyclic peptide derived from the eighth TSHR LRD loop, and peptide 19 (a 11-meric peptide) from the first loop of the TSHR LRD.
In several pilot studies, optimal dosing of these peptides was evaluated by studying their effects on major outcome parameters. Hence, it was determined that 0.3 mg/kg body weight of the 13-mer peptide 836 yielded the best results, as well as 0.3 mg/kg of peptide 12 and 0.1 mg/kg of peptide 19. Therefore, these doses were chosen to carry out a randomized, blinded main study, and it is these results that are shown in the following. The control peptide 14 was used at a standard dose of 1 mg/kg body weight in order not to miss out on any effects that may occur at somewhat higher doses.
No animals were prematurely taken out of the study after randomization, and the prespecified protocol was completed in all animals. Results reflect the respective means of all values of all randomized animals.
Thyroid sizes, as determined from serial sections
Thyroid volumes (mm3) were determined from the sum of the areas of each section over the whole cutting region (between 5 and 10 slides, depending on the respective size of the thyroid gland) multiplied by the slice thickness of 0.5 mm. This macroscopic investigation showed clearly increased thyroid sizes in mice that had received nine immunizations of Ad-TSHR289 (see Fig. 2) compared to the healthy mouse group. In contrast, mice treated with peptide 19 showed decreased thyroid sizes, whereas treatment with peptide 12 or the 836 13-mer failed to reach statistical significance.
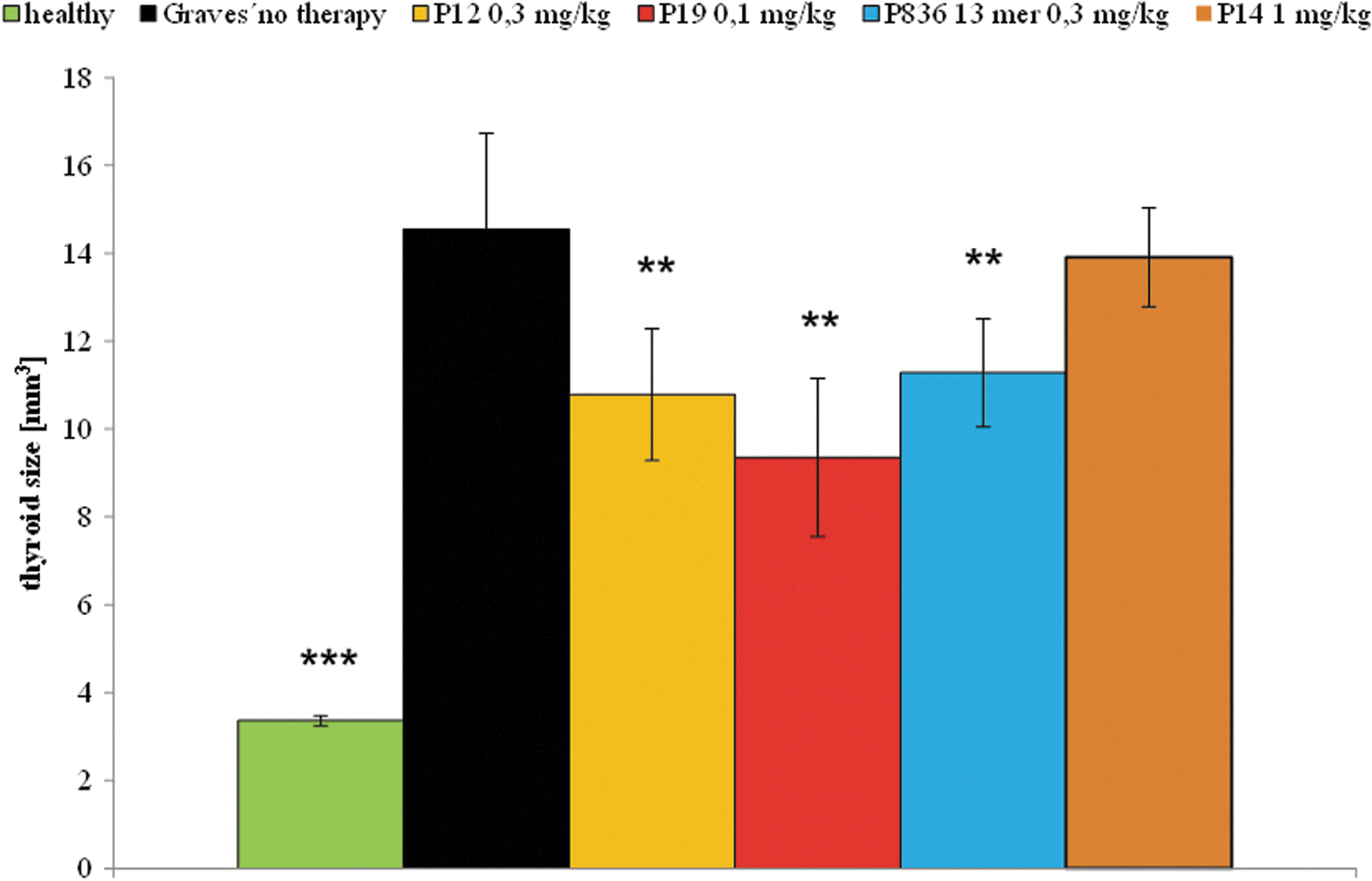
Effects on macroscopically measured thyroid size. The effects of peptide therapy on thyroid size were investigated at the end of the experiment. The measurements were carried out in Ad-TSHR-immunized mice treated by either four-weekly injections with vehicle (0.9% NaCl, “Graves' no therapy,” 10 mice), or administrations of 0.3 mg/kg body weight of peptide 836 13-mer (11 mice), 0.3 mg/kg of peptide 12 (10 mice), 0.1 mg/kg of peptide 19 (10 mice), or 1 mg/kg control peptide 14 (11 mice). In addition, age-matched immunologically naïve unimmunized mice (“healthy,” 10 mice) were investigated. The mean thyroid sizes in mm3 are shown with standard error of the mean (SEM). Differences between groups were tested by analysis of variance (ANOVA) followed by post hoc least significant difference (LSD) testing. **p < 0.05 and ***p < 0.001 compared to the TSHR-immunized group treated with only NaCl (“Graves' no therapy”).
Determination of T4 serum levels
T4 levels did not differ between groups at the start of the study, and mean T4 levels in eligible animals of the Ad-TSHR289-immunized groups were significantly higher than controls at week 11 (start of therapy; Fig. 3) and exceeded the normal T4 value range, which was defined as 5.97–6.81 μg/dL in the authors' laboratory (means ±3 standard errors of control mice). After the start of therapy, mice treated with peptide 19 and peptide 12 showed progressively decreasing T4 levels. These values even reverted to normal values in the group treated with peptide 19. In contrast, values in mice treated with peptide 836 failed to reach statistical significance.
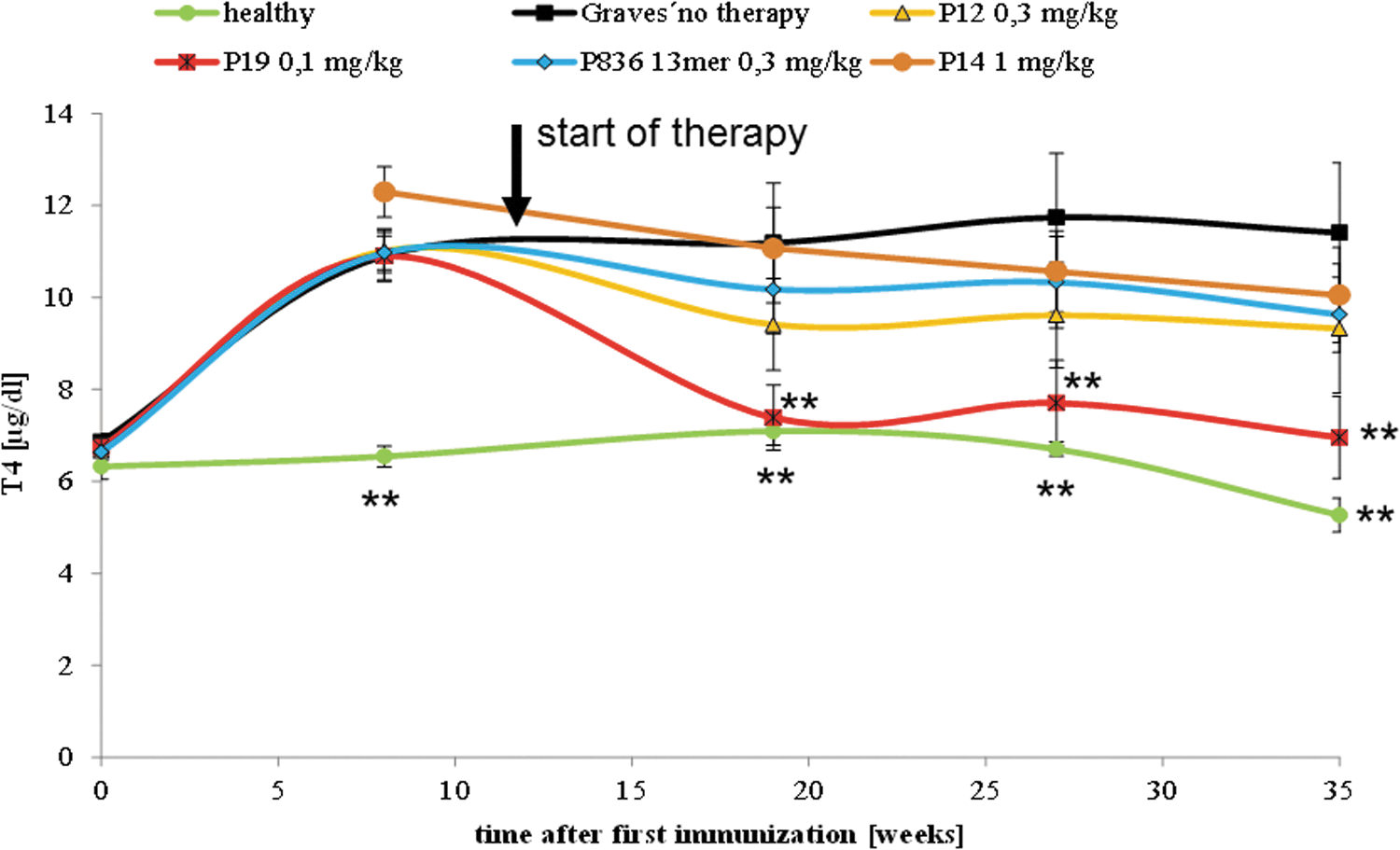
Effect on serum thyroxine (T4) levels. The effects of peptide therapy on serum T4 levels were evaluated. The measurements were carried out in Ad-TSHR-immunized mice treated by either four-weekly injections with vehicle (0.9% NaCl, “Graves' no therapy,” 10 mice), or administrations of 0.3 mg/kg body weight of peptide 836 13-mer (11 mice), 0.3 mg/kg of peptide 12 (10 mice), 0.1 mg/kg of peptide 19 (10 mice), or 1 mg/kg control peptide 14 (11 mice). In addition, age-matched immunologically naïve unimmunized mice (“healthy,” 10 mice) were investigated. Data are represented as means ± SEM. Significance over time was tested by ANOVA of groups at given time points, and controlled by ANOVA for repeated measurements within one group, followed by LSD post hoc testing. **p < 0.01 compared to the TSHR-immunized group treated with only NaCl (“Graves' no therapy”).
Histopathological changes of the orbits
Histological investigation of retro-orbital fibrosis was carried out after serial sectioning of the orbits. In nine AdTSHR-treated mice, a significant increase of retro-orbital fibrosis was observed upon digitized image analysis, which was markedly reduced in mice treated with peptide 12 and 836 13-mer, whereas a trend was observed in the group treated with peptide 19, which did not reach statistical significance (Fig. 4). Unfortunately, no data for the peptide 14 control group were obtained because the respective tissue samples were not available.

Effects on digitized analysis of retro-orbital fibrosis volumes after histological preparation of orbital sections. The effects of peptide therapy on the severity of retro-orbital fibrosis were evaluated in histological sections of all available animals. The measurements were carried out in Ad-TSHR-immunized mice treated by either four-weekly injections with vehicle (0.9% NaCl, “Graves' no therapy,” 10 mice), or administrations of 0.3 mg/kg body weight of peptide 836 13-mer (11 mice), 0.3 mg/kg of peptide 12 (10 mice), or 0.1 mg/kg of peptide 19 (10 mice). In addition, age-matched immunologically naïve unimmunized mice (“healthy,” 10 mice) were investigated. The mean total fibrosis volumes of all orbits (left and right), as assessed by digitized image analysis of all sections, and consecutive integrations are shown with SEM. Differences between groups were tested by ANOVA. **p < 0.01 and ***p < 0.001 compared to the TSHR-immunized group treated with only NaCl (“Graves' no therapy”).
ECG to determine heart rates
Starting from the third immunization, a significant increase in heart rate in the hyperthyroid Ad-TSHR289 immunized group was observed (Fig. 5). In contrast, the heart rate in the native, healthy group was only mildly increased at older age, but did not change significantly.
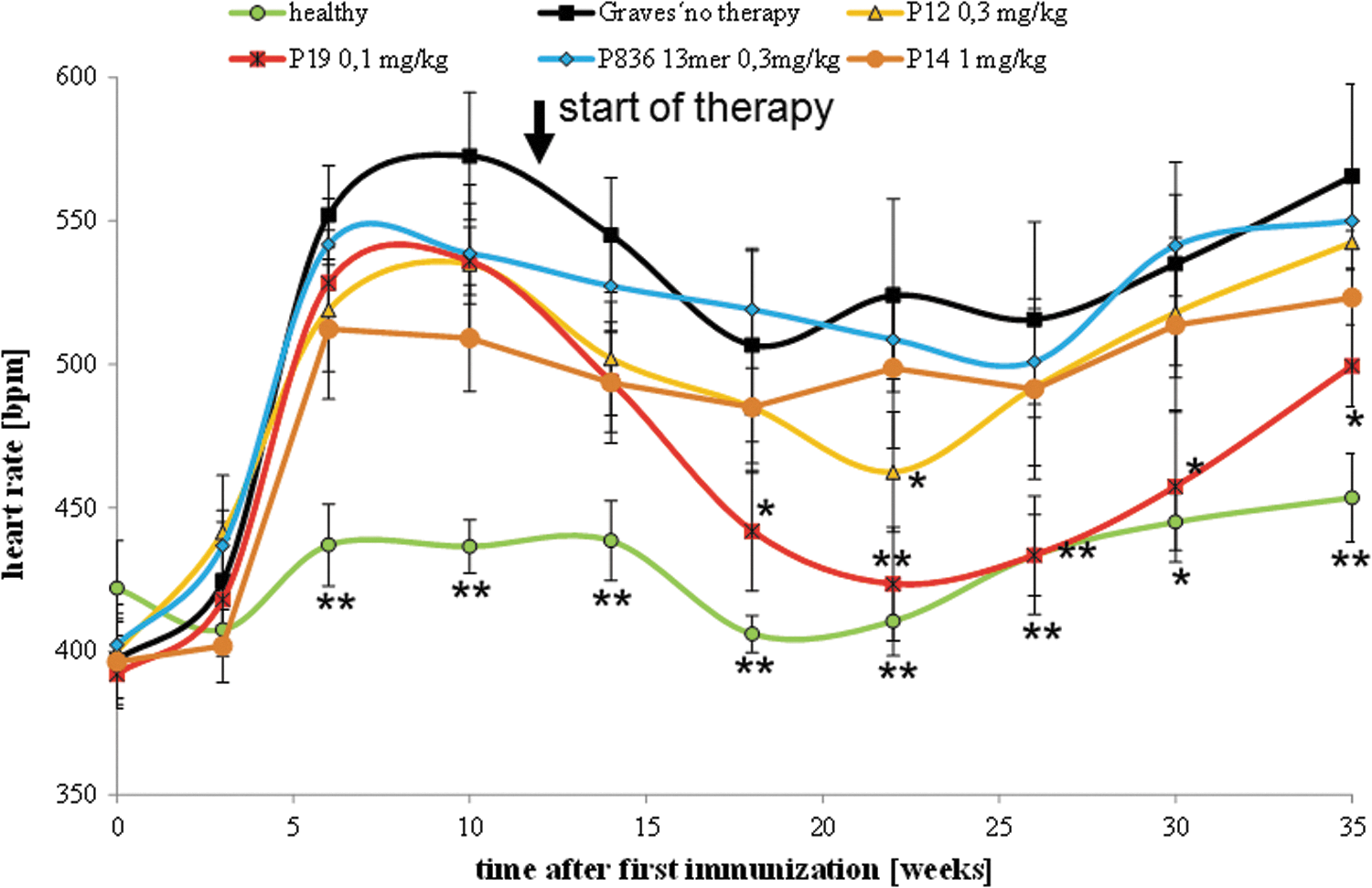
Effect on heart rates. The effects of peptide therapy on heart rates at various times were evaluated in all animals. The measurements were carried out in Ad-TSHR-immunized mice treated by either four-weekly injections with vehicle (0.9% NaCl, “Graves' no therapy,” 10 mice), or administrations of 0.3 mg/kg body weight of peptide 836 13-mer (11 mice), 0.3 mg/kg of peptide 12 (10 mice), 0.1 mg/kg body weight of 19 (10 mice), or 1 mg/kg control peptide 14 (11 mice). In addition, age-matched immunologically naïve unimmunized mice (“healthy,” 10 mice) were investigated. Data are represented as means ± SEM. Significance over time was tested by ANOVA of groups at a given time points, and controlled by ANOVA for repeated measurements within one group, followed by LSD post hoc testing. *p < 0.05 and **p < 0.005 compared to the TSHR-immunized group treated with only NaCl (“Graves' no therapy”).
Upon consecutive Ad-TSHR289 immunizations, a further strong increase in heart rate in the hyperthyroid vehicle-treated group was observed. In contrast, animals treated with peptide 19 showed a larger variation but significantly decreased resting heart rates at later time points. In contrast, mice treated with peptide 836 13-mer and peptide 12 showed trends toward reduced heart rates, which did not reach statistical significance.
Anti-TSHR antibody titers and capacity to stimulate cAMP in test cells
Anti-TSHR antibodies were determined from serum samples by investigating the ability of the respective mouse sera to inhibit the binding of the monoclonal Graves' patient antibody M22 to the TSHR (third-generation ELISA). Highly significant titers were detected in all Ad-TSHR289-immunized animals (Supplementary Fig. S1).
As in previous studies, there were no statistically significant variations of anti-TSHR titers in vivo between groups.
In addition, the stimulatory activity of these antibodies was determined as the capacity of mouse serum samples to induce an increase in TSHR-dependent cAMP levels in test cells (Fig. 6). Anti-TSHR antibodies from almost all TSHR-immunized mice showed potency to stimulate cAMP in TSHR-expressing test cells. The maximum inducible cAMP levels showed considerable variation. Peptide therapy did not impact on these mean TSHR-triggered intracellular cAMP levels.
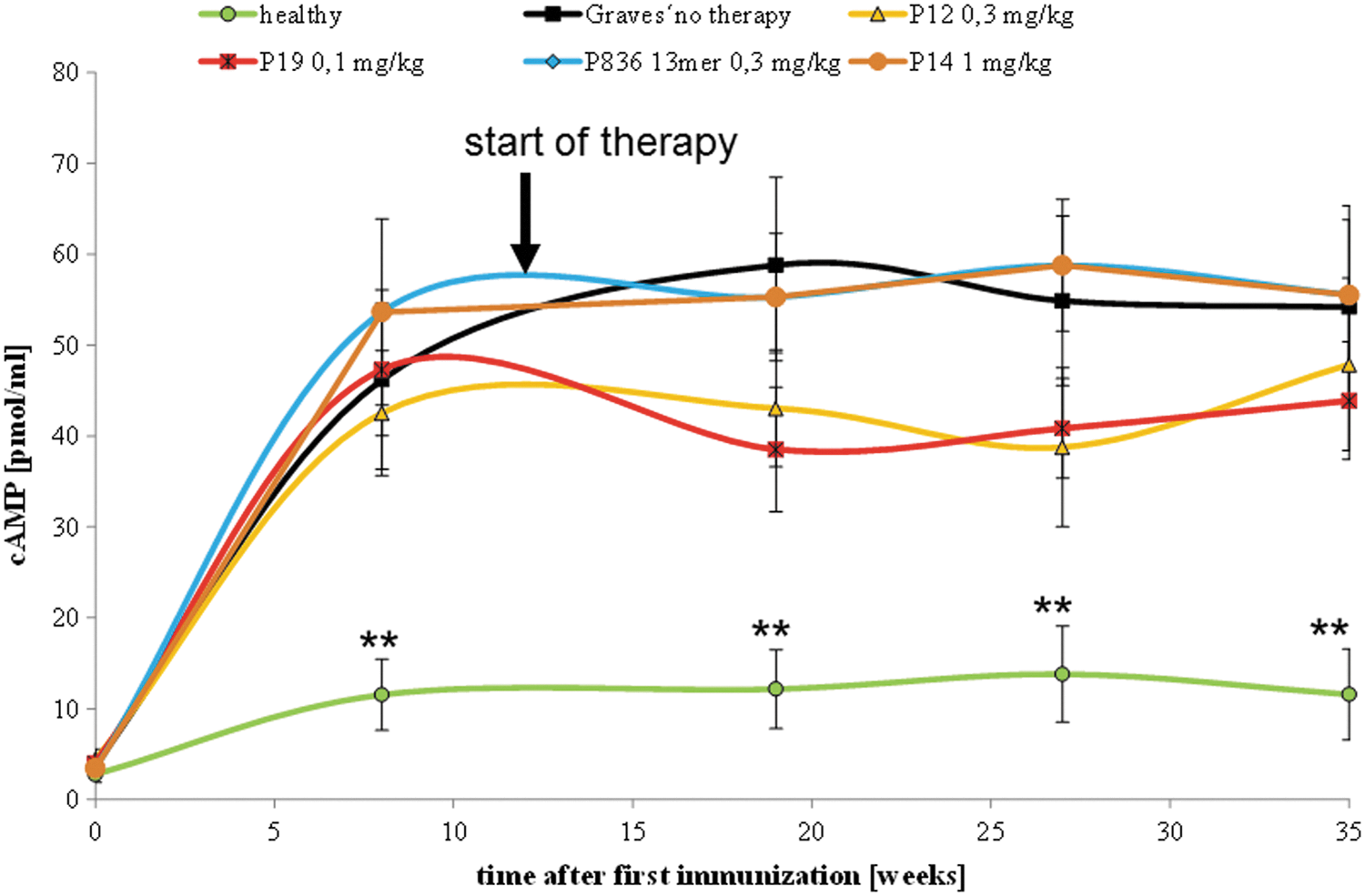
Effect on cyclic adenosine monophosphate (cAMP) stimulation in TSHR-expressing test cells, as determined in sera taken from the mice during the experiment. The effects of peptide therapy on the capacity of anti-TSHR antibodies to stimulate cAMP generation were evaluated in Chinese hamster ovary cells expressing the human TSHR. The measurements were carried out in Ad-TSHR-immunized mice treated by either four-weekly injections with vehicle (0.9% NaCl, “Graves' no therapy,” 10 mice), or administrations of 0.3 mg/kg body weight of peptide 836 13-mer (11 mice), 0.3 mg/kg of peptide 12 (10 mice), 0.1 mg/kg of peptide 19 (10 mice), or 1 mg/kg control peptide 14 (11 mice). In addition, age-matched immunologically naïve unimmunized mice (“healthy,” 10 mice) were investigated. Data are represented as means ± SEM. Differences between groups were tested by ANOVA followed by post hoc LSD testing. **p < 0.01 compared to the TSHR-immunized group treated with only NaCl (“Graves' no therapy”).
In vitro studies: ELISA
To characterize the effects of the cyclic peptides ex vivo/in vitro, two ELISA assays were carried out. In contrast to the observed lacking inhibitory effects of all peptides in the canonical third-generation assay in vivo (Supplementary Fig. S1), a clear inhibition of peptide 19 was found on an in vitro assay. When added to sera from Ad-TSHR289-immunized mice, peptide 19 inhibited the binding of the polyclonal anti-TSHR antibodies, which were contained therein, to coated TSHR-Fc effectively at a low IC50 value of 65 ng/mL, corresponding to 47 nmol/L (Fig. 7). However, these ex vivo effects were not observed with peptide 836 13-mer or peptide 12.
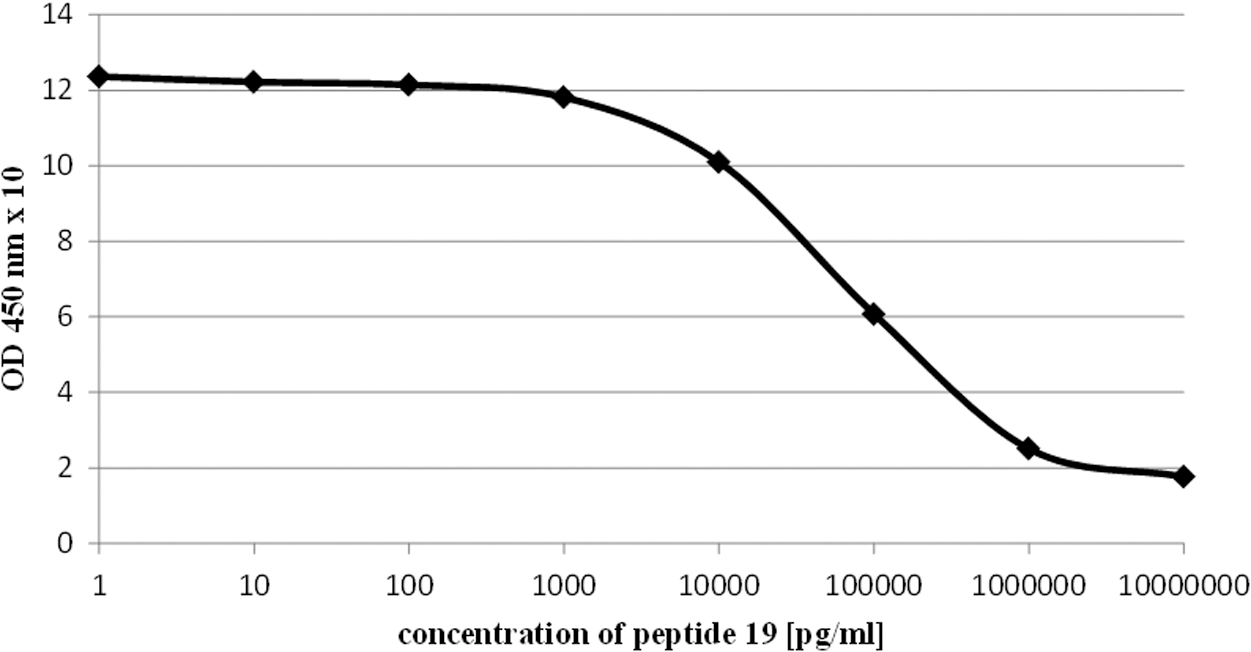
Effect of peptide 19 on anti-TSHR antibody titers in Ad-TSHR-immunized mouse serum samples. The effects of peptides on anti-TSHR antibody titers from Ad-TSHR-immunized mice were studied ex vivo. Each measurement was carried out in four samples. Results are shown as optical density (OD) 450 values with SEM.
Next, the study investigated whether the cyclic peptides inhibit the inhibition of M22 binding by human Graves' patient serum samples ex vivo (based on the canonical third-generation titer assay). Results are shown in Figure 8. There was no difference in anti-TSHR antibody titers between peptide groups (final concentration: 100 μg/mL) versus controls.
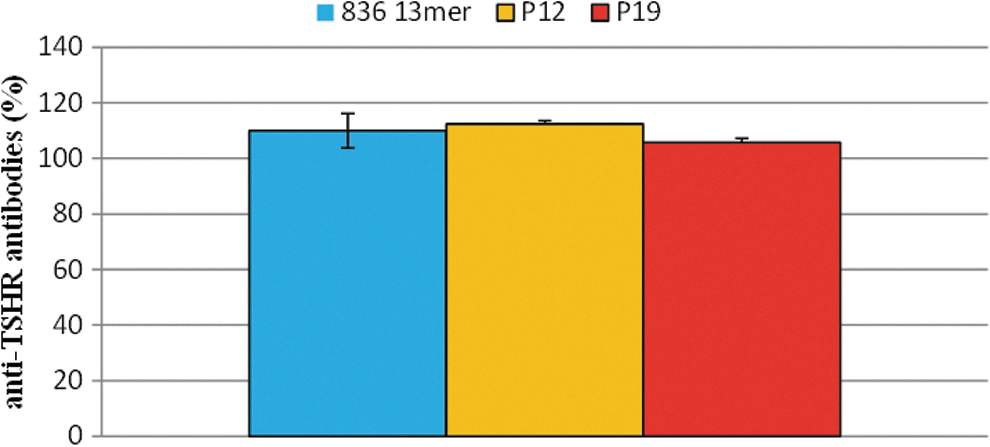
Effect of peptides ex vivo on anti-TSHR antibody titers, as determined in a double approach in a modified third-generation assay. The effects of peptides on third-generation assay anti-human TSHR antibody titers (detected by competing with the monoclonal anti-TSHR antibody M22) were studied ex vivo. Each measurement was carried out in four samples. Results are shown as percentage of untreated controls with SEM. Significance was tested by ANOVA between groups, followed by LSD post hoc testing. There were no statistically relevant differences between groups.
Studies in immunologically naïve mice
All immunologically naïve mice tolerated six-monthly administrations of either 1 mg/kg body weight cyclic peptide 836 13-mer, peptide 19, peptide 12, or vehicle (NaCl) equally well. No pathological clinical findings were observed. Generation of anti-TSHR antibodies was not observed in any of the peptide-treated animals over six months. All measured titers were below background, so that no immune response to either peptide was documented. No statistically significant difference occurred between peptide- and vehicle-treated groups (as assessed by either t-test or ANOVA).
Discussion
This study shows that repeated four-weekly i.v. administrations of 0.1 mg/kg body weight cyclic peptide 19, which is a shortened derivate from the first cylindrical loop of the LRD of TSHR, reduced thyroid hyperplasia in a long-term mouse model of GD. Elevated T4 levels and sinus tachycardia were also reduced, starting eight weeks after initiation of peptide therapy. Administration of 0.3 mg/kg body weight of the cyclic peptide 836 13-mer (shortened derivate from the eighth TSHR LRD) resulted in trends toward improvements of these parameters, which did not reach statistical significance but significantly improved retro-orbital fibrosis. A novel linear multi-epitope peptide 12 (given at a dose of 0.1 mg/kg body weight) also mitigated T4 levels and tachycardia. Administration of the peptides in immunologically naïve mice reconfirmed that neither the shorter cyclic peptides nor the linear multi-epitope peptide are immunogenic on their own. The novel peptides could offer significant advantages over existing therapies of Graves' disease and over other currently investigated therapeutic approaches, as summarized Neumann et al. (14).
Compared to a previous study (8), these data show that markedly shortened TSHR epitope-derived cyclic peptides can be designed that still result in relevant therapeutic activity in the mouse GD model. The previous study had been driven by the hypothesis that the three-dimensional confirmation of the 10 TSHR LRD loops (11) should be largely conserved by the design of cyclic peptides with potential activity, which ended up in 24- to 25-meric peptides. Since these peptides present a challenge for production and might also incur risks of immunogenicity upon long-term therapy, shorter variants were also studied. It is now shown that it is actually feasible to reduce peptide length to 11- or 13-meric variants. Surprisingly, these shorter variants retain activity at smaller doses, so that i.v. peptide load could be reduced by 10-fold for two variants.
Peptide 19 is a shortened version of the first loop-derived cyclic peptide 829, which was investigated in a previous study (8). Peptide 829 only resulted in small, nonsignificant trends to improve disease parameters. In contrast, peptide 19, whose sequence is derived from the same TSHR LRD area but presents a 11-meric variant cyclized by a disulfide bond (not via the peptide backbone), was more effective than the 24-meric peptide 829. Therefore, the cyclic structure of the peptides (e.g., C-C bonding) seems to be important, not just the aa sequence from which they derive.
Serum samples from Ad-TSHR289-immunized mice were used in an assay that determined binding of their sets of polyclonal anti-TSHR antibodies to TSHR-Fc ex vivo. Addition of cyclic peptide 19 but not of the cyclic peptide 836 13-mer or 12 ex vivo resulted in high affinity inhibition of these anti-TSHR antibody titers. In contrast, anti TSHR-antibody titers as determined in a slightly modified third-generation assay using the monoclonal anti-TSHR antibody M22 were not affected by peptide therapy in vivo. After further assay development, the study also investigated whether the peptides inhibit the inhibition of M22 binding by human Graves' patient serum samples in a third-generation-like assay ex vivo, and also did not find any significant effect of any of the investigated peptides. The failure to observe such an effect may also depend on the TSHR binding substrate in these third-generation like M22-based assays because this substrate has not been completely disclosed by the manufacturer and seems to derive from a porcine TSHR variant. The observed lack of effect on TSHR-dependent cAMP formation in test cells may depend on lacking sensitivity of this assay.
Also the effects of the novel peptides on the cardiac manifestations and complications of GD were investigated. Tachycardia is a reliable marker of disease severity in hyperthyroid patients (15,16). Regular ECG registrations served to detect the effect on heart rate. Treatment with peptide 19 significantly decreased tachycardia, which progressively developed in untreated TSHR-immunized mice over three to nine months.
There are, however, limitations in this study. The effects of the various peptides on hyperthyroidism and orbitopathy were not always consistent, and we do not currently have a full explanation for these observations, which need to be further investigated.
In summary, this study shows that treatment with a series of cyclic peptides in a mouse model of GD led to marked improvement of several disease parameters. The novel compounds, either shortened TSHR LRD loop-mimicking cyclic peptides or a multi-epitope linear combination peptide, led to differential findings. Direct induction of immune mechanisms in vivo might account for these effects. These findings are in accordance with those on cyclic peptides, which are being investigated in other autoimmune diseases (17).
Footnotes
Author Disclosure Statement
All authors are employees of a biotech company.