
Abstract
Background:
Serum thyroid state in older adults correlates with extended longevity. We hypothesized that age impacts not only systemic but also organ-specific thyroid state and response to thyroxine (T4).
Methods:
Young (3 months) and old (23 months) male mice were analyzed at baseline and after acute T4 challenge. Age effects on circulating thyrotropin (TSH) and thyroid hormone (TH) concentrations, transcript expression in the pituitary and thyroid were compared with organ-specific responses characterized by hepatic and cardiac content of TH and TH metabolites and expression of TH-target genes, as well as hepatic deiodinase 1 activity.
Results:
Circulating TH concentrations and hepatic and cardiac TH content were lower in old versus young mice. After injection with T4, conversion of T4 to triiodothyronine was decreased in old mice while TH transport in liver and heart was not affected. Organ-specific TH response was augmented in old mice in liver but not heart, indicating age- and tissue-specific sensitivity to TH. A compensatory increase of thyroid stimulating hormone subunit beta expression in the pituitary and increased serum TSH concentrations, but reduced expression of thyroid differentiation markers were found in old mice.
Conclusions:
We suggest that a reduced activity of the aged thyroid is responsible for the systemic low TH state in old mice. Further, divergent TH metabolism and tissue response in liver and heart occur after T4 treatment in an aged organism. These rodent data are in agreement with a much narrower window for T4 substitution in the older adults to avoid overtreatment.
Introduction
Aging modulates thyroid hormone (TH) action and metabolism. In aged patients, the prevalence of thyroid dysfunction increases, but it is associated with an attenuated and oligosymptomatic clinical presentation of hyper- and hypothyroidism (1,2). Thyroid function, defined by serum thyrotropin (TSH), thyroxine (T4), and triiodothyronine (T3), has been found to change with aging toward higher TSH and lower free T3 (fT3) and free T4 (fT4) concentrations in humans (3 –5). Interestingly, decreased thyroid function in older adults is suggested to contribute to an increased lifespan (6,7). Thus, it remains under debate whether subclinical hypothyroidism, defined by TSH concentrations above a laboratory reference range, in older adults is a pathological state or whether its treatment is beneficial. Importantly, in a recent study, levothyroxine treatment over 1 year did not improve symptoms in patients with biochemical subclinical hypothyroidism older than 65 years of age (8).
The underlying cause of the age-dependent changes in systemic thyroid function may be caused by changes in the hypothalamic-pituitary-thyroid-axis (HPT-axis) (7), but the precise molecular mechanisms are not known. Further, it is still unknown whether systemic changes are also reflected in organ-specific adaptation of TH action with age.
In this study, we aimed at characterizing the age-dependent TH state in a systemic and organ-specific manner. Systemic effects represented by circulating TSH, TH concentrations, and transcript expression in the pituitary and thyroid were compared with organ-specific responses characterized by hepatic and cardiac content of TH and TH metabolites, the expression of TH-target genes, as well as hepatic deiodinase 1 activity. Further, by challenging mice with a single T4 injection, the age-dependent response was assessed in young (3 months) and old (23 months) male mice.
Materials and Methods
Animals
Male C57Bl/6J mice were purchased from Janvier Labs, St. Berthevin, France. Two age cohorts were investigated: twelve 3-month-old (young cohort) and twenty-two 23-month-old mice (aged cohort). Animals were single-housed in temperature- (23 ± 1°C) and light-controlled (inverse 12:12 hour light-dark cycle) conditions. Food and water were provided ad libitum. All animal experiments were performed in accordance with the German regulations for Laboratory Animal Science (GVSOLAS) and the European Health Law of the Federation of Laboratory Animal Science Associations (FELASA). The protocols for animal studies were approved by the Landesamt für Natur, Umwelt und Verbraucherschutz, Nordrhein-Westfalen (LANUV-NRW), Germany.
Treatment and organ collection
Mice were randomly divided into control (n = 6 for 3 months, n = 8 for 23 months) or T4 treatment (n = 6 for 3 months, n = 11 for 23 months) groups, 12 hours before sacrifice. Treated mice were injected intraperitoneally (i.p.) once with 1 μg/g body weight L-thyroxine (T4, T2376; Sigma-Aldrich, St. Louis, MO); stock-solution: 2 mg/mL T4 dissolved in 0.01 M NaOH, 0.1% bovine serum albumin (BSA, A7906; Sigma-Aldrich); injection solution; and stock-solution diluted 1:10 with phosphate-buffered saline (PBS, 18912014; Thermo Fisher Scientific, Waltham, MA). Control mice received an i.p. injection of 150 μL PBS. Twelve hours after treatment, mice were deeply anesthetized by an i.p. injection of 200 μL Ketamine/Xylazine mixture (150 μL of 100 mg/mL Ketamine [Beta-pharm, Vechta, Germany] and 50 μL of 20 mg/mL Xylazine [Ceva, Düsseldorf, Germany]), and final blood was obtained by heart puncture. For tissue collection, mice were perfused with heparinized saline through a needle placed in the left heart ventricle. Tissues were shock-frozen in liquid nitrogen, and they were stored at −80°C until further processing.
Serum TH and TSH measurements
fT4 and fT3 concentrations in serum of mice were measured by using commercial ELISA kits according to the manufacturer's instructions (fT3: EIA-2385; fT4: EIA-2386; DRG Instruments GmbH, Marburg, Germany). Serum samples with known TH concentrations were used as standards. According to the manufacturer's instructions, the minimum detectable TH concentration is 0.05 ng/dL for fT4 and 0.05 pg/mL for fT3, and the inter-assay variation is <9% for fT4 and <11% for fT3. TSH serum concentration was determined by using the Milliplex Map mouse pituitary magnetic bead panel (MPTMAG-49K; EMD Millipore Corporation, Billerica, MA) on the Luminex 200 system (Thermo Fisher, Waltham, MA). The minimal detectable concentration is 1.9 pg/mL, and the inter-assay variation is <20% in this assay.
TH measurements in liver and heart
TH content in liver and heart tissue of mice was analyzed as previously described (9,10), and all solvents were purchased from Promochem (Wesel, Germany). Between 10 and 50 mg of tissue was homogenized in 300 μL of pure methanol (MeOH) and sonicated by using an ultrasound probe. After homogenization, 60 μL of internal standard and 600 μL of chloroform (CHCl3) were added. The mixture was centrifuged at 2348 × g for 10 min, and the supernatant was then collected. This extraction procedure was repeated two more times. The CHCl3-MeOH extracts were transferred into a 12-mL tube for back-extraction of the iodothyronine with 500 μL of 0.05% calcium chloride (CaCl2) in water at 4°C. The extraction procedure was repeated two more times by adding 500 μL of MeOH and 500 μL of 0.05% CaCl2 in water each time. The pooled aqueous phase was concentrated under nitrogen stream at 40°C (to eliminate the excess of MeOH). Phosphoric acid was added into the combined extracts to reach a final concentration of 2%, followed by the addition of an antioxidant solution (0.2 mL; 10 mg ascorbic acid +10 mg citricacid +10 mg dithiothreitol in 1.5 mL MeOH). After vortexing, the mixture was loaded onto a Bond Elut Plexa PCX cartridge, which was preconditioned sequentially with, 1.5 mL of pure MeOH and 1.5 mL of water. The cartridge was first washed with 2 mL of 2% formic acid in water and then with 2 mL of MeOH: acetonitrile (1:1, v/v). Analytes were eluted into a vial with 1 mL of 5% ammonium hydroxide (28–30%) in MeOH: acetonitrile (1:1, v/v). The solvent was evaporated, and compounds were re-dissolved in 60 μL of a mixture of 20% acetonitrile in water containing 0.1% formic acid for instrumental analysis. The quantification limits were 2.5 pg injected into column for 3,5-diiodothyronine (T2), reverse T2 (rT2) and T3; 3.8 pg injected into column for reverse T3 (rT3) and T4; and 5 pg injected into column for monoiodothyronine (T1). The inter-assay variation was determined by measuring mouse liver samples. For this experiment, three different mouse livers were chosen: (i) mouse livers in which the concentration of T3 and T4 was in the lower range of the experimental window, (ii) mouse livers in which the concentration of T3 and T4 was in the medium range of the experimental window, and finally, (iii) mouse livers in which the concentration of T3 and T4 was in the higher range of the experimental window. Per each mouse liver, four different batches were prepared and measured at four different days within two weeks. The inter-assay variation was <15% for T3 (low, medium, and high concentration) and <20% for T4 (low, medium, and high concentration).
Quantitative real-time polymerase chain reaction
Total RNA from pituitaries, thyroid gland, liver, and heart was isolated by using the RNeasy Kit according to the manufacturer's instruction (Qiagen, Hilden, Germany) and stored at −80°C as previously described (11). RNA was reverse transcribed into cDNA with SuperScriptIII (Life Technologies, Darmstadt, Germany) and hexamer primers. Quantitative real-time polmerase chain reaction (qRT-PCR) was performed by using Roche SYBR Green I master mix (Roche, Mannheim, Germany). Primers were designed to be intron-spanning to exclude genomic DNA signals (sequences are provided in Supplementary Table S1). In compliance with the MIQE guidelines for RT-PCR (12), we used a set of two to three reference genes per tissue to assure accurate normalization and calculation (pituitary: 18S ribosomal RNA [18S], peptidylprolyl isomerase A, cyclophilin A [Ppia]; thyroid gland: 18S, hypoxanthine guanine phosphoribosyl transferase [Hprt]; liver: 18S, Ppia, ribosomal protein L13a [Rpl13a]; heart: 18S, polymerase RNA II [Polr2a], glyceraldehyde-3-phosphate dehydrogenase [Gapdh]). Analysis and calculation of the fold-change in gene expression were done on Ct-values ≤35 by using the efficiency-corrected ΔΔCt method (13).
Genes known to be involved in TH regulation (pituitary: thyroid stimulating hormone, beta subunit [Tshß], thyrotropin releasing hormone receptor [Trhr], nuclear receptor co-repressor 2 [Ncor2]), TH synthesis (thyroid gland: thyroid stimulating hormone receptor [Tshr], thyroid peroxidase [Tpo], thyroglobulin [Tg], sodium iodide symporter [Nis]), TH metabolism (deiodinases type 1, 2 and 3 [Dio1, Dio2, Dio3]), and organ-specific response (liver: thyroxine binding globulin [Tbg], thyroid hormone responsive [Spot14], malic enzyme [Me1], B cell leukemia/lymphoma 3 [Bcl3]; heart: myosin heavy chain 6 and 7 [Myh6, Myh7], hyperpolarization-activated, cyclic nucleotide-gated potassium channel 2 and 4 [Hcn2, Hcn4]) were assessed.
Hepatic deiodinase 1 activity
Liver protein samples (40 μg of total proteins; n = 6 per group) were prepared by mincing and sonification, adjusted to a defined protein concentration (5 μg/μL). Dio1 activity was assayed as previously described (14). In brief, a 50-μL reaction mixture containing 40 μg of liver microsomal proteins, 10 mM 6-n-propyl-2-thio-uracil (PTU) for controls, was mixed with 50 μL of freshly prepared substrate mix (20 μM rT3 [Sigma-Aldrich, MO, USA], 0.2 M KPO4 [pH 6.8], 2 mM ethylenediaminetetraacetic acid, and 80 mM dithiothreitol). The enzyme reaction lasted for two hours at 37°C. After centrifugation (4°C, 15,000 g, five minutes), the supernatant was used for quantification of released iodide. Dowex W50-X2 resin columns were used for separation of intact rT3 and the deiodinated breakdown products from the released iodide. The iodide content was determined by the Sandell-Kolthoff reaction, using cerium solution (25 mM (NH4)4Ce(SO4)4 and 0.5 M H2SO4) and arsenite solution (25 mM NaAsO2, 0.8 M NaCl, and 0.5 M H2SO4). The changes in absorption (optical density at 415 nm) were determined at the reaction starting point and after 20 min. All protein samples were assayed in triplicate. The tubes that contained PTU were used for background subtraction. Calculation of the enzyme activity was performed by utilizing an iodide standard curve.
Histological analysis
Thyroid glands were fixed in 4% formaldehyde, embedded in paraffin and 5-μm-thick sections were used for staining. For morphological evaluation, hematoxylin and eosin (H&E) staining was performed. Staining procedures followed standard protocols. All samples were viewed on the Olympus BX51 upright microscope (Olympus, Germany) with a magnification of 10 × and 20 × . Quantification of follicle size was done by using ImageJ (NIH) by determination of 78–245 follicles per animal (n = 4 mice per group).
Statistical analysis
All data are shown as mean ± standard deviation (SD) or standard error of the mean as indicated. Statistical analysis was performed by using GraphPad Prism 6 Software. One-way analysis of variance followed by Tuckey's post hoc analysis or unpaired Student's t-test were applied as indicated. Values of *p < 0.05, **p < 0.01, ***p < 0.001, and # p < 0.0001 were considered statistically significant.
Results
Aging is associated with lower T4 concentrations in serum and lower T4 content in liver and heart of male mice
Compared with 3-month-old animals, 23-month-old mice displayed slightly higher serum TSH (p = 0.0568, Fig. 1A) and significantly lower concentrations of serum fT4 (young: 2.2 ng/dL and old: 0.7 ng/dL, p < 0.001, Fig. 1B) and T4 content in the liver (young: 38.5 pg/mg and old: 19.3 pg/mg, p < 0.01, Fig. 1C) and heart (young: 7.2 pg/mg and old: 3.9 pg/mg, p < 0.01, Fig. 1D). Further, a significantly lower hepatic T3 (young: 4.5 pg/mg and old: 2.2 pg/mg, p < 0.01, Fig. 1F) content was found in old mice. Concentrations of other TH metabolites (rT3, T2, rT2, and T1) were below the detection limit in the liver and heart in all animals.
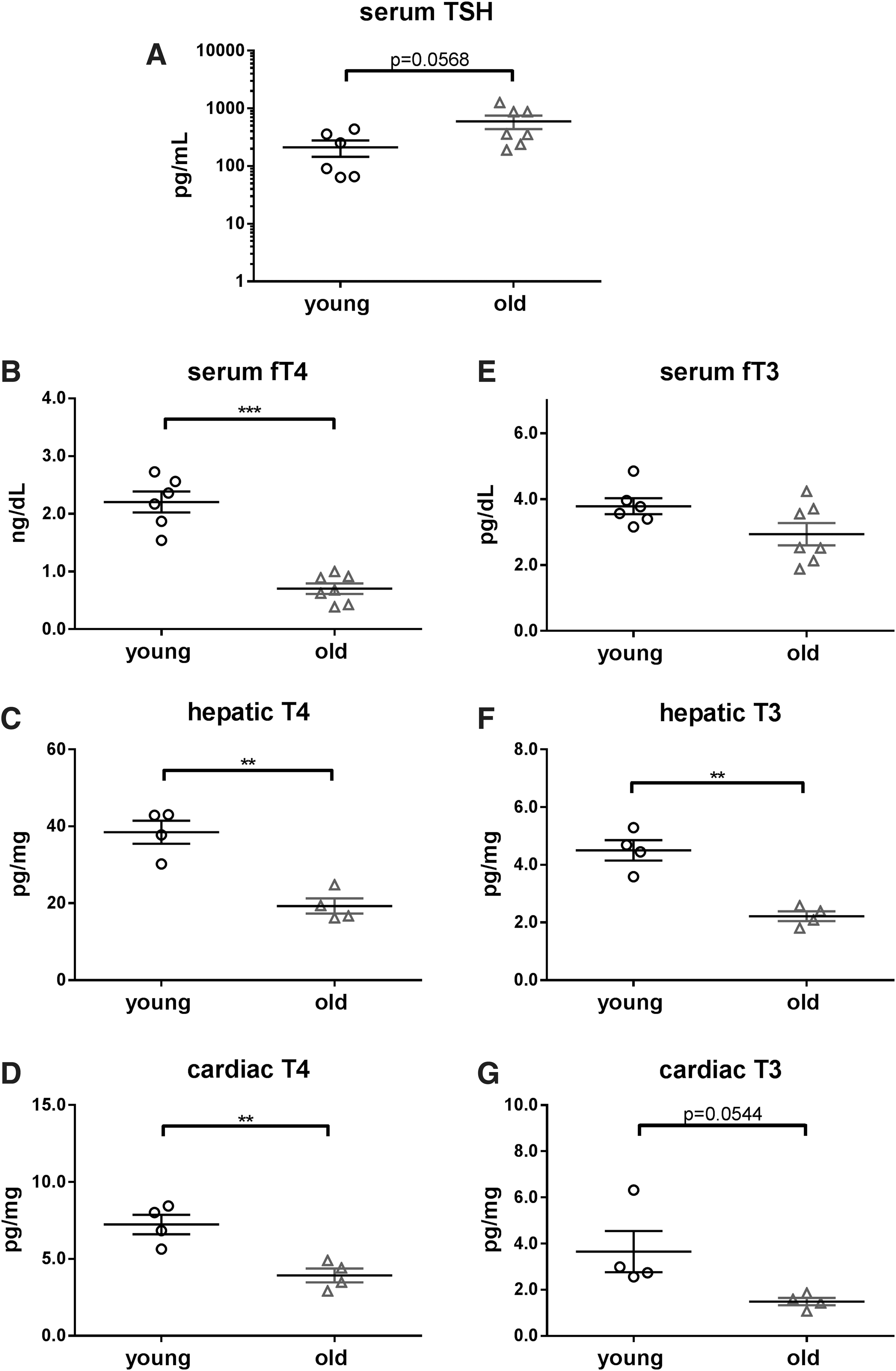
TSH and TH concentrations in sera, livers, and hearts of young and old mice. TSH (
Expression of TH-target genes correlates with tissue TH concentration
To assess how TH serum and tissue concentrations in young and aged mice are associated with altered TH tissue action, qRT-PCR analysis was performed in liver and heart. An age-dependent downregulation of the positively regulated TH target gene Dio1 (p < 0.001), and an increased expression for the negatively-regulated gene Tbg (p < 0.05, Fig. 2A) was found in livers of old versus young mice. Similarly, decreased TH action in the heart was suggested by downregulation of Hcn2 (p < 0.001) and upregulation of Myh7 (p < 0.001, Fig. 2B) in old mice versus young mice.

Age- and organ-specific expression of TH responsive genes. Transcript changes influenced by age were assessed in liver (
The function of HPT-axis is preserved while the thyroid gland is hypoactive in old mice
Next, we addressed the functional state of the HPT-axis by measuring gene expression in pituitaries and thyroids of young and old mice. An increased expression of Tshb (p < 0.01) was found in pituitaries of old compared with young mice (Fig. 2C), supporting the biochemical finding in serum of a systemic low TH state with a compensatory active response at the pituitary level. Transcript analysis of genes relevant for TH synthesis indicated a hypoactive thyroid organ with decreased Tg gene expression (p < 0.001, Fig. 2D), and a trend toward lower Nis (not significant [ns]) and Tpo (ns) expression in old versus young mice.
Age alters thyroid follicle shape and function
To verify the hypothesis of a hypofunctioning thyroid gland, H&E stained formalin-fixed thyroid gland sections of 3- and 23-month-old mice were assessed (Fig. 3A). Quantification of the follicle size displayed an age-dependent increase in size (Fig. 3B). Further, heterogeneity in follicle size with mostly flattened epithelium was noted in thyroid glands of old mice.

Morphology of thyroid glands in young and old mice. Formalin-fixed thyroid sections of 3 and 23 months control-treated mice were H&E stained (
Conversion of T4 into T3 is less efficient in aged mice
Whether age has an impact on TH metabolism, and to investigate how it may affect systemic and organ-specific TH response, we exposed young and aged mice to a T4 challenge. A single injection of 1 μg T4/g body weight (Table 1) resulted in distinct changes in serum TH concentrations and tissue content after 12 hours in young and old mice. Suppression of TSH was comparable in young and old mice (Fig. 4A), and an age-independent increase was observed for serum fT4 concentrations and hepatic T4 content, whereas cardiac T4 content was higher in old mice (young: 139 pg/mg and old: 207 pg/mg, p < 0.05; Fig. 4B–D). Interestingly, and irrespective of the T4 increase, the conversion of T4 into T3 was less efficient in old mice (Fig. 4E–G). This resulted in smaller incremental increases in T3 serum concentrations and tissue content, further illustrated by higher T4/T3 ratios in serum, liver, and heart of old compared with young mice (Supplementary Table S2). Also, after the T4 challenge, TH metabolites (rT3, T2, rT2, and T1) were below the respective detection limit in livers and hearts of all mice.

TSH and TH concentration changes 12 h after single T4 injection in young and old mice. TSH (
Body Weight of Young and Old Male Mice at Time of Sacrifice Mean ± Standard Deviation
PBS, phosphate-buffered saline.
TH responsive gene expression is age- and organ-dependent after single T4 injection
Changes in TH responsive gene expression in organs at 12 hours after T4 injection showed an age- and organ-specific response in mice. This was observed for gene expression in the pituitary (Dio2, Trhr, and Tshb; Fig. 5A) and the thyroid gland (Tg and Nis; Fig. 5B). Surprisingly, and in contrast to the findings in the heart, TH target gene expression in the liver differed in old versus young animals after acute T4 challenge. Whereas Tbg, Spot14 and Bcl3 expression was not altered, liver Dio1 was significantly upregulated at 12 hours after T4 injection in old compared with young mice (p < 0.05; Fig. 5C). In contrast, only marginal changes in expression were noted for TH-dependent genes in the heart after T4 injection, and only Hcn2 expression increased in mouse heart, irrespective of age (Fig. 5D).

Effect of single T4 injection on gene expression in young and old mice. Gene expression changes were assessed by qRT-PCR in pituitaries (
Hepatic deiodinase type 1 activity after T4 injection reflects hepatic TH content
Since induction of Dio1 expression on T4 treatment was not in line with the higher increment in hepatic T3 content in young versus old mice, Dio1 activity was analyzed. No age differences were found in Dio1 activity under basal conditions; however, after T4 injection, Dio1 activity increased significantly in young (Fig. 6A, p < 0.01), but not old mice, and it confirmed the differences measured for hepatic T3 content in the animals.

Hepatic deiodinase 1 activity and summary of the impact of age on systemic and organ-specific TH metabolism and response. Hepatic Dio1 activity was determined in young and old PBS and T4-treated mice (
Discussion
In clinical practice, age-dependent changes of thyroid function are of high relevance, frequently leading to misinterpretation of thyroid function parameters and unindicated treatment. An association between age and a decline in serum fT4 and fT3 concentrations, accompanied by an increase of TSH in areas with iodine sufficiency, has previously been reported in humans (3,4,15), and this may be favorably associated with longevity (6). The underlying mechanisms responsible for age-dependent changes in thyroid state and their consequence for organ-specific TH supply and action are still not fully understood. Using a rodent model and the approach of a single T4 challenge, we report for the first time an impact of age on systemic and organ-specific TH metabolism and tissue response.
Aging per se is associated with mild systemic hypothyroidism caused by the thyroid gland
Old mice had lower fT4 serum concentrations than young mice, accompanied by low hepatic and cardiac T4 content in these animals. Increased expression of Tshb in the pituitaries and slightly higher TSH serum concentrations in old mice indicate an adequate function of the negative feedback loop of the HPT-axis. However, expression of genes encoding for proteins involved in TH synthesis was downregulated in the thyroid gland. Further, changes in thyroid morphology, including variable sizes and enlargement of follicles, were noted. Recently, changes in aged mouse thyroids have been attributed to a hypofunctioning thyroid gland (16). This is in line with the gene expression data observed in this study and suggests that the sensitivity of an aging thyroid gland to TSH is decreased. This interpretation is further supported by elevated TSH concentrations in mice of advanced age, as shown here and as previously reported by others (17). Altered TSH efficacy on the aged thyroid gland has been reported in rats (18 –20), with possible differences in TSH secretion, bioactivity, and Tshr availability on the thyrocytes itself. However, the data reported here suggest a compensatory response rather than a causative role of the pituitary gland in aging.
Altered and organ-specific T3 supply in aged mice
Analysis of fT3 in serum and T3 in liver and heart showed that T3 content in the liver was lower in old compared with young animals while only marginal age differences were observed in serum and heart. Further, repression of positively TH regulated genes (e.g., hepatic Dio1) and induction of negatively regulated genes (hepatic Tbg) was found in liver of old animals, in line with low hepatic TH content. An age-dependent change in hepatic deiodinase activities with decreased Dio1 and increased Dio3 activity was previously reported in mice older than 24 months of age (17). However, in the experimental conditions used in this study, Dio1 activity did not differ in the liver of young and old PBS-treated animals. Surprisingly, significant changes in TH-regulated genes were found in old compared with young hearts, for example, induction of cardiac Myh7 and repression of Hcn2 expression. These findings suggest an organ-specific regulation of T3 supply during aging, which may be of relevance for the function of different tissues. Thus, these observations could also explain why TH excess is distinctly detrimental in older patients, resulting in increased cardiac morbidity and mortality (1,21,22).
Altered response to T4 challenge in aged mice
The debate whether to treat hypothyroidism in older adults is still ongoing (8,23,24). In addition, recent studies have emphasized the risk of potential adverse effects of overtreatment in older adults, who may be prone to an increased likelihood of T4-induced side effects and even increased mortality (25). Therefore, we asked whether acute organ response to T4 might differ between young and old mice. Several important observations were made: First, T4 serum concentrations correlate directly with organ T4 concentrations in young and old mice, strongly suggesting that the specific transport of T4 into liver and heart is not influenced by age. Second, conversion of T4 to T3 is decreased in old mice with lower systemic concentrations and tissue T3 content compared with young mice. Consequently, the ratios of T4/T3 were higher in serum, liver, and heart of old compared with young mice. Interestingly, the hepatic Dio1 activity measurements strongly indicate a role for Dio1 as a cause of this finding, as an increase after T4 injection was only noted in livers of young, but not old mice, illustrating the age-dependent difference of hepatic TH content. Third, since T3 increment is lower in serum and tissues after T4 injection in old compared with young mice, we also expected a decreased tissue response. This was confirmed for the pituitary, thyroid, and, to some extent, for the liver. Interestingly, in liver tissue the induction of the TH responsive gene Dio1, in contrast to activity, was significantly higher in old mice, possibly due to the impact of mild hypothyroidism on gene expression. After T4 injection, expression levels of TH responsive genes were equal in the hearts of old and young mice, while in the liver, pituitary, and thyroid gland fewer genes showed expression changes in old compared with young mice. This implies that aging may differentially impact TH response in various organs (26). These findings are in agreement with the notion that an aging organism is in a “low TH state.” Further, and importantly, old compared with young mice show a different organ-specific response to an acute TH challenge (Fig. 6B, C).
A shortcoming of our study is that we only determined the age effect at two time points (basal and 12 hours after acute T4 challenge), and that we cannot extrapolate on long-term effects on the organism. However, in previous studies, we have observed age-dependent differences on phenotypic traits of chronic hyper- and hypothyroidism in mice and think that they are of physiological relevance (26,27). A strength of our study is the characterization of very old mice of 23 months of age in a direct comparison to 3-month-old mice. Further, we comprehensively analyzed systemic and peripheral effects of TH concentrations in untreated and T4-treated mice and expanded the analysis by gene expression analyses of hepatic TH metabolism by Dio1.
In summary, the divergent systemic and organ-specific TH states and responses to T4 in old versus young mice support the clinical notion that TH substitution, while considered adequate at the serum level, may not be equally beneficial for all tissues, particularly in an aging organism. Further, age may impact local TH content and TH metabolism. Thus, our findings are in agreement with a narrower window for TH substitution in older adults and particularly careful clinical and biochemical monitoring of T4-substituted patients in this age group.
Footnotes
Acknowledgments
The authors are grateful to A. Jaeger, J. Göbel, S. Rehn, K. Strumann, and S. Kwapik for their dedicated technical support and thank the Imaging Core Facility Essen (IMCES) for support with the Olympus microscope. Further, they are thankful to Prof. Dr. S. Brandau and K. Bruderek (Department of Otorhinolaryngology, University Hospital Essen) for providing the Luminex 200 system for TSH measurement.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was funded by DFG FU 356/7-2 to D.F., MO 1018/2-2 to L.C.M., and ZW 221-2/1 to D.Z. in the framework of SPP 1629 THYROID TRANS ACT.
Supplementary Material
Supplementary Table S1
Supplementary Table S2