
Abstract
Background:
Primary cilia (PC) are conserved structures in the adult thyroid gland of different mammals. It was recently described that in humans, PC are usually present as a single copy per follicular cell emerging from the follicular cell apex into the follicular lumen.
Methods:
To understand the role developed by PC in thyroid hormonogenesis better, their changes in different human functional thyroid diseases (diffuse toxic hyperplasia/Graves' disease [GD] and nodular hyperplasia [NH]/nodular goiter), in comparison to normal thyroid tissue, were investigated using immunofluorescence, morphometry, and electron microscopy analyses.
Results:
Significantly decreased ciliary frequencies were found in both NH (51.16 ± 11.69%) and GD (44.43 ± 23.70%) compared to normal thyroid tissue (76.09 ± 7.31%). Similarly, PC lengths were also significantly decreased in both NH (2.02 ± 0.35 μm) and GD (2.4 ± 0.48 μm) compared to normal glands (3.93 ± 0.90 μm). Moreover, in GD patients, hyperactive-follicle foci always showed diminished ciliary frequency and length compared to any other thyroid follicle pattern, independent of their thyroid status. Finally, in GD, the percentage of thyrocytes exhibiting PC in the “normal-appearance areas” was significantly lower in correspondence with the subsistence of signs of thyroid biosynthetic hyperactivity after long-term antithyroid drug treatment.
Conclusions:
The results suggest a direct relationship between ciliogenesis and both follicle activity and tissue heterogeneity in the functional pathology of the thyroid gland.
Introduction
Since their discovery by Zimmermann in 1898, primary cilia (PC) have been found in the vast majority of cell types in vertebrates, such as the renal tubule (1), bile duct (2,3), pancreatic cells (4), neurons (5,6), keratinocytes, fibroblasts, endothelial cells (7), and thyroid cells (8 –12). In the last decade, PC have emerged as key organelles in numerous cellular, physiological, and developmental processes (13 –15). In most models, PC act as extracellular sensory antennae associated with several important signaling pathways, for example wingless-Int-1, planar cell polarity, and Hedgehog pathways (16 –18). Alterations in PC structure or function have been reported to be responsible for several human diseases—so-called ciliopathies (19 –21).
Primary ciliogenesis is inversely correlated with cell-cycle progression, and PC have been proposed as negative regulators of cell division. PC are disassembled at the late S phase of the cell cycle and are considered organelle characteristics of cells in a differentiated state (22,23). Moreover, primary ciliogenesis requires an established apical membrane domain and shares several protein complexes also necessary for cell polarization and apical delivery (24,25).
It was recently described that PC are conserved structures in the adult thyroid gland of different mammals (humans, pigs, guinea pigs, and rabbits), where they are usually present as a single copy per follicular cell (12). The presence of PC in normal (Nthy-ori 3-1) and neoplastic (FTC-133 and 8505C) human thyroid cell lines has also been demonstrated, and their frequency has been reported to be lower in neoplastic compared to normal thyroid (NT) cells. In addition, defects in ciliogenesis have been described in malignant thyroid diseases, including papillary thyroid carcinoma (PTC) and Hürthle cell carcinoma (26).
The thyroid gland is a unique endocrine organ composed of follicles, with thyrocytes organized as an apicobasal-polarized epithelium surrounding a lumen in which their secretory product—thyroglobulin (Tg)—is stored extracellularly in large quantities. Tg iodination is carried out by a complex enzymatic machinery—named thyroxisome—located and restricted at the apical plasma membrane of thyrocytes (27,28) that produces moderate amounts of H2O2, which is essential for hormone synthesis, as well as other potentially toxic reactive oxygen species (29,30). Thyroid follicles, together with their adjacent capillaries, form so-called angiofollicular units (AFUs) that are considered the functional and morphological units of the thyroid gland, and are subjected to independent self-regulatory mechanisms that contribute to the heterogeneous nature of thyroid tissue (31 –34). In fact, in NT tissue, smaller active follicles displaying high columnar polarized epithelium coexist with larger hypofunctioning follicles surrounded by low cuboidal or flattened thyrocytes; this heterogeneity in size and activity are hallmarks of the human NT gland (35).
It is plausible to assume that in thyrocytes, PC, taking advantage of their ideal localization extending from the follicular cell apex into the follicular lumen, sense the colloid environment, and this sensory activity, coupled to specific intracellular downstream signaling pathways, contributes to the complex mechanism of thyroid hormones synthesis. Consequently, defects in ciliogenesis should be present in functional thyroid diseases such as diffuse toxic hyperplasia (Graves' disease [GD]) or nodular hyperplasia (NH; nodular goiter), which are characterized by a deregulation of hormone synthesis, changes in follicle integrity, and an altered proliferation rate.
Specifically, GD thyroid samples, from patients subjected to prior treatment with antithyroid drugs, show a highly marked variability from area to area, ranging from diffuse hyperplasia exhibiting tall columnar thyrocytes, papillary infoldings, and very little light stained colloid to zones displaying different degrees of regression of the hyperfunctioning changes characterized by flat cells and increased colloid stores (36,37). Nodular hyperplastic thyroid tissue also shows follicles that exhibit high heterogeneity in size and morphology, varying from small follicles with minimal amounts of colloid lined by high columnar thyrocytes to very large follicles containing abundant colloid lined by flat epithelium. These large follicles are considered the characteristic pattern of NH (38 –40).
Although PC are key organelles that have been associated with an increasing number of pathologies and whose presence in the surface of the thyrocyte has been known for decades (9 –12), there is a complete lack of information in the literature regarding the putative role that they could play in either the NT or in functional thyroid disease.
To understand the role of PC in thyroid hormonogenesis better, this study investigated PC in the normal human thyroid gland and in different functional thyroid diseases using both morphometrical and electron microscopy analysis. The identification of the molecular mechanisms underlying defective ciliogenesis in thyroid pathology could help in understanding the role played by PC in thyroid hormone biosynthetic activity and could shed light on the histopathological features of diseases affecting thyroid function.
Methods
Human thyroid specimens
Five human thyroid samples—three obtained from families with hereditary medullary thyroid carcinoma who underwent a prophylactic thyroidectomy, and two normal samples adjacent to resected PTCs—were used as NT glands. Ten NH or multinodular goiter tissues, and 10 diffuse toxic hyperplasia or GD samples, were obtained from patients undergoing thyroid surgery, diagnosed at the Department of Pathology of the Virgen Macarena University Hospital of Seville. All GD patients were subjected to prior treatment with antithyroid drugs (carbimazole). Tissue samples were collected with approval from the Research Ethics Committee of the Virgen Macarena University Hospital (C.P.-C.I. 1921). The clinicopathologic features of the patients are summarized in Tables 1 and 2.
Baseline Characteristics of the Patients
TSH-R, thyroid stimulating hormone receptor; ND, not determined/no data; TR-X, total thyroidectomy.
Clinical Parameters of Graves' Disease Patients
fT4, free thyroxine; TSH, thyrotropin.
Thyroid glands were fixed in 10% neutral-buffered formalin, embedded in paraffin using the standard procedure, sectioned at 4–5 μm thickness, and mounted on silane-coated glass slides. Consecutive tissue sections were stained with hematoxylin and eosin (H&E) for histological diagnosis and then visualized to select thyroid tissue with a characteristic appearance to perform immunofluorescence (IF).
Ultrastructural studies
Samples for transmission electron microscopy (TEM) and scanning electron microscopy (SEM) were obtained from the same two previous patients with normal-appearing thyroid adjacent to thyroid carcinoma, and from three and five of the previous patients with NH and GD, respectively.
For the TEM studies, pieces were fixed in 2.5% glutaraldehyde in 0.1 M cacodylate buffer (pH 7.2), post-fixed in 1% osmium tetroxide, dehydrated in acetone, and embedded in Spurr, as previously reported (12). Ultrathin sections were photographed with a Zeiss Libra 120 transmission electron microscope.
Similarly, for the SEM studies, the samples were fixed in the same glutaraldehyde solution for a minimum of five days and post-fixed in 1% osmium tetroxide. After dehydration, specimens were dried with the critical point method using CO2, sputter coated with vacuum-evaporated gold and photographed with a Zeiss EVO scanning electron microscope, as previously reported (12).
Double IF staining
The double IF staining was carried out according to the procedure previously reported (12). In brief, the sections were dewaxed in xylene and hydrated through graded alcohols. An antigen retrieval step using EnVision Flex Target Retrieval Solution High pH (DM828; Dako) was performed in a heating instrument, PTLink (Dako). After applying washing solution and after nonspecific blocking with 10% normal donkey serum, the primary antibody, a monoclonal anti-acetylated α-tubulin (SigmaAldrich) that labels the axoneme was applied. The slides were then incubated with Cy3-labeled donkey anti-mouse IgG secondary antibody (Jackson ImmunoResearch Laboratories). After washing, the slides were incubated with polyclonal rabbit anti-E-cadherin antibody (Santa Cruz BiotechnologyUSA) and subsequently with Cy2-labeled donkey anti-rabbit IgG antibody (Jackson ImmunoResearch Laboratories). DAPI was added for nuclei counterstaining, and the slides were coverslipped with Dako Fluorescent Mounting Medium (S3023). Different controls for specificity of the IF technique were performed.
The samples were observed under a fluorescence microscope (Olympus BX50) equipped with a scientific digital camera (Hamamatsu ORCA-03G). All image files were processed using Image-Pro-Plus v7.0 (Media Cybernetics) to create composite RGB micrographs, enhance contrast, and obtain measurements.
Morphometrical analysis
Analysis of PC frequency
To evaluate the frequency of PC in the different functional thyroid groups, 10–20 micrographs per case at 200 × magnification were morphometrically assessed using a software processing and image analysis (Cell* Imaging Software). First, in every photograph, a ranking of the thyroid follicles according to their histological appearance and their size (internal perimeter) was established: (1) rounded follicles of different sizes: small-sized follicles (<50 μm), medium-sized follicles (50–100 μm), large-sized follicles (100–500 μm), and giant follicles (>500 μm, with abundant colloid); (2) follicles with papillary infoldings; and (3) hyperfunctioning follicles, which exhibited a tall follicular epithelium and scanty colloid. The average thyroid follicles examined per patient was as follows: in the NT group, a minimum of 35 follicles per thyroid section (15 small-sized follicles, 15 medium-sized follicles, and at least five large-sized follicles) were assessed; in the NH group, a minimum of 50 follicles of different sizes per section was examined, including giant follicles; and in the GD group, in which the heterogeneity among follicles was pronounced after antithyroid treatment, a minimum of 60 follicles of every pattern per gland was analyzed. Second, the ciliary frequencies in follicular cells were assessed by analyzing the relative number of cilia protruding from the apical surface of the epithelium versus the number of nuclei in adequately oriented sections of those thyroid follicles. To this end, all visible cilia that were connected to the thyrocyte apical surface and oriented toward the colloid were measured, independently of their angle or apparent orientation. In total, the presence of PC in the current study was evaluated in 1300 thyroid follicles and 43,000 thyrocytes: 5000 thyrocytes (an average of 1000 per case) from NT glands, 18,000 thyrocytes (an average of 1800 per case) from NH samples, and 20,000 thyrocytes (an average of 2000 per case) from GD samples.
Analysis of PC length
PC lengths were morphometrically assessed in 150 composite micrographs acquired using Image-Pro-Plus v7. with a 40 × , UPlanFl N.A. = 0.75 objective. To minimize oblique sectioned ciliary length underestimation, cilia were measured at all relative angles or orientations, and only those lacking connection to the follicular epithelium surface or not reaching the colloid were excluded from the analyses. In brief, the length of PC was evaluated in at least 100 follicular cells per case, with a total of 2727 cilia being measured, which corresponded to the different histological patterns established in the different groups.
Statistical analysis
The percentage of thyrocytes exhibiting PC and the axonemal lengths in the different kinds of thyroid follicles of each group were measured and expressed as the mean ± standard deviation. In addition, the same procedure was applied to study differences between NH and GD compared to NT. Statistical differences were tested by either one-way analysis of variance followed by the corresponding post hoc test or the Mann–Whitney rank sum test. p-Values <0.05 were accepted as significant.
Results
Histological aspects
As the existence of a correlation between follicular size and functional activity is generally accepted (with smaller follicular sizes considered more active, and the largest follicular sizes lined by flattened cells considered hypofunctioning follicles), in the present study, a ranking of thyroid follicles according to their size and appearance was established. In the NT group, a mixture of normal-rounded follicles of different sizes was observed, with small- and medium-sized follicles having more abundance than large follicles (Fig. 1A). In the NH group, however, follicles exhibited conspicuous differences in size, ranging from small with minimal colloid to very large colloid lakes, so-called giant follicles, which were typically lined by flattened epithelium (Fig. 1B). Finally, in the GD group, as all patients received some form of therapy before resection, the heterogeneity in follicular pattern was rather characteristic (Fig. 1C). In addition to normal-rounded follicles of different sizes, abundant areas with hyperplastic papillae and focal areas with hyperfunctioning follicles appeared, adding to the variable presence of lymphocytic infiltrates and follicles in apoptosis (Fig. 1C). As expected, no PC were identifiable in thyroid sections stained by H&E.
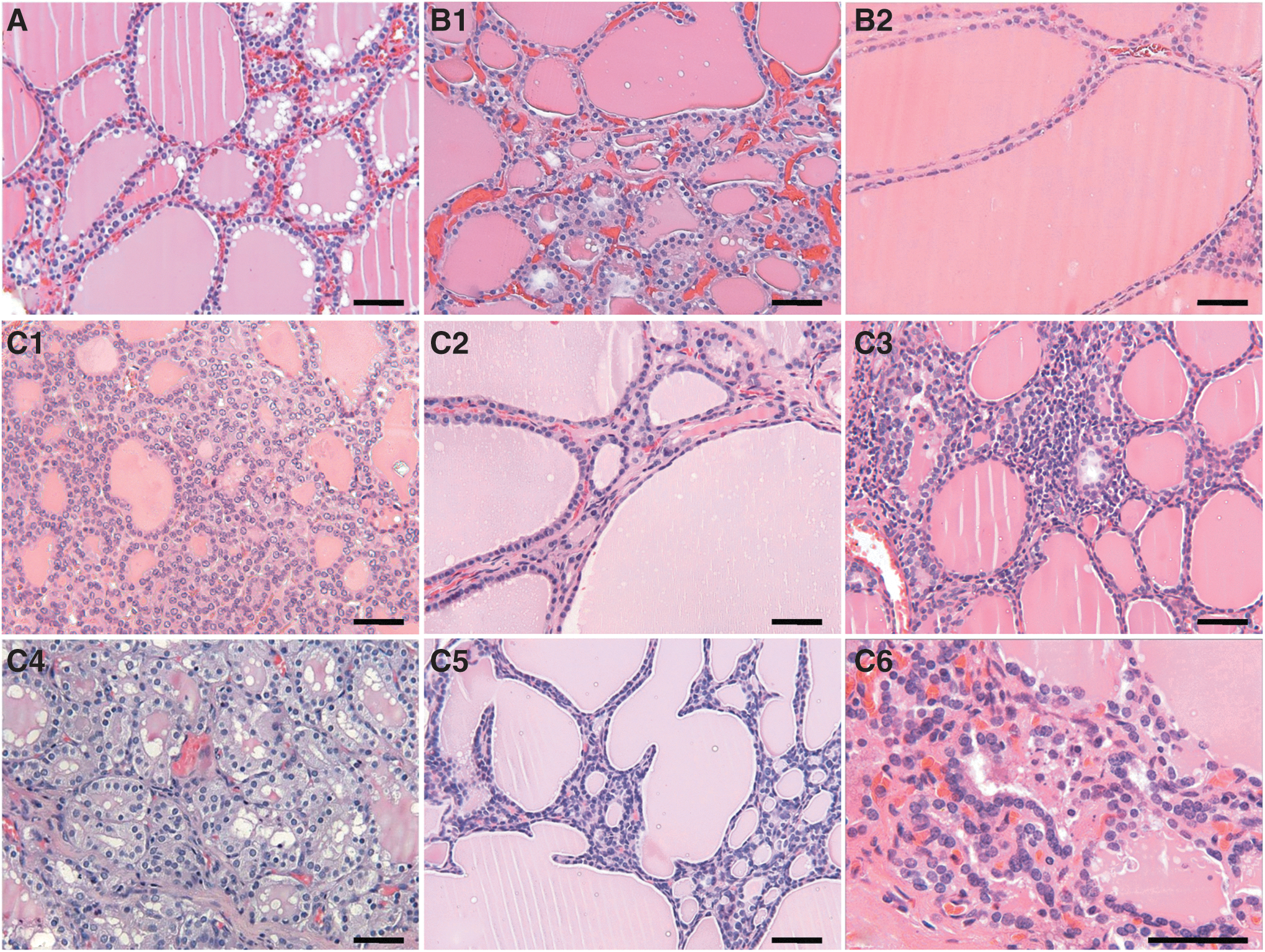
Typical morphological aspects of thyroid follicles in (
PC detection by immunofluorescence
The presence of PC was detected in the different thyroid sections using acetylated α-tubulin antibody and Cy3 labeling, which mark the axoneme in red. To confirm their location and frequency, an E-cadherin antibody and Cy2 label were used, which mark the epithelial cell perimeter in green. Generally, PC emerged from the apical surface of thyrocytes and entered into the colloid perpendicularly.
In NT glands, thyrocytes exhibiting PC were observed in all follicles. When the same material was analyzed at a higher magnification, almost every follicular cell harbored a unique PC (Fig. 2). In tangential sections of the thyroid follicles, PC were seen to emerge mainly from the central area of the apical surface of every thyrocyte (Fig. 2E).
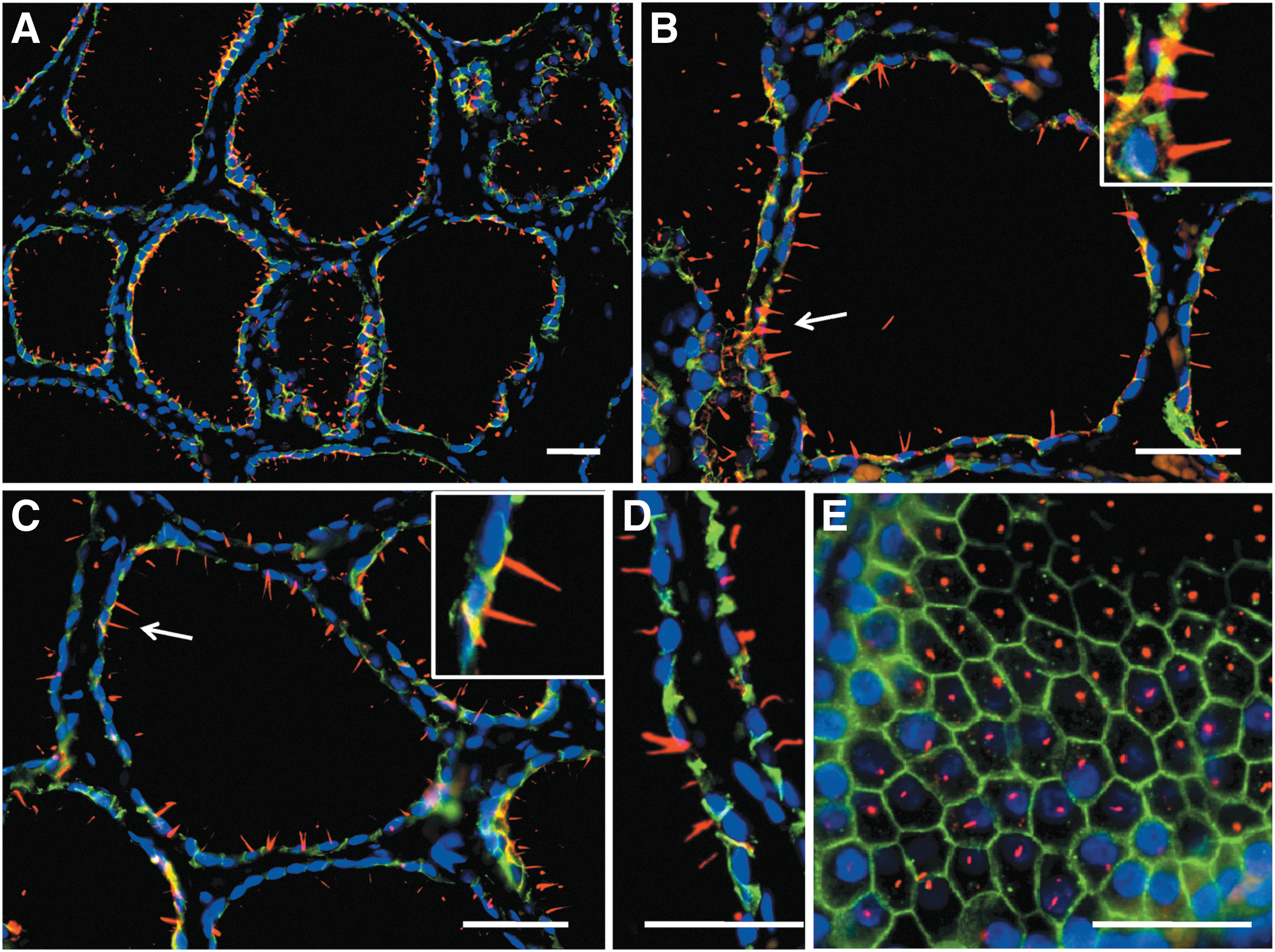
Distribution of primary cilia (PC) in NT glands using double immunofluorescence (E-cadherin, green; acetylated α-tubulin, red; nuclear counterstaining with DAPI, blue). Thyroid follicles exhibit numerous and easily recognizable PC oriented toward the colloid. Virtually every follicular cell displays at least one cilium (
In NH, both the number of follicular cells showing PC and the average PC length considerably decreased, independently of the follicular size (Fig. 3). In contrast, in GD, the apparent frequency of thyrocytes presenting PC varied considerably among follicles, in accordance with the rather follicular heterogeneity inherent to diffuse toxic hyperplasia after antithyroid treatment (Fig. 4). Consequently, a rigorous morphometrical study was performed to evaluate the PC frequencies in the thyrocytes of the different functional thyroid groups objectively.
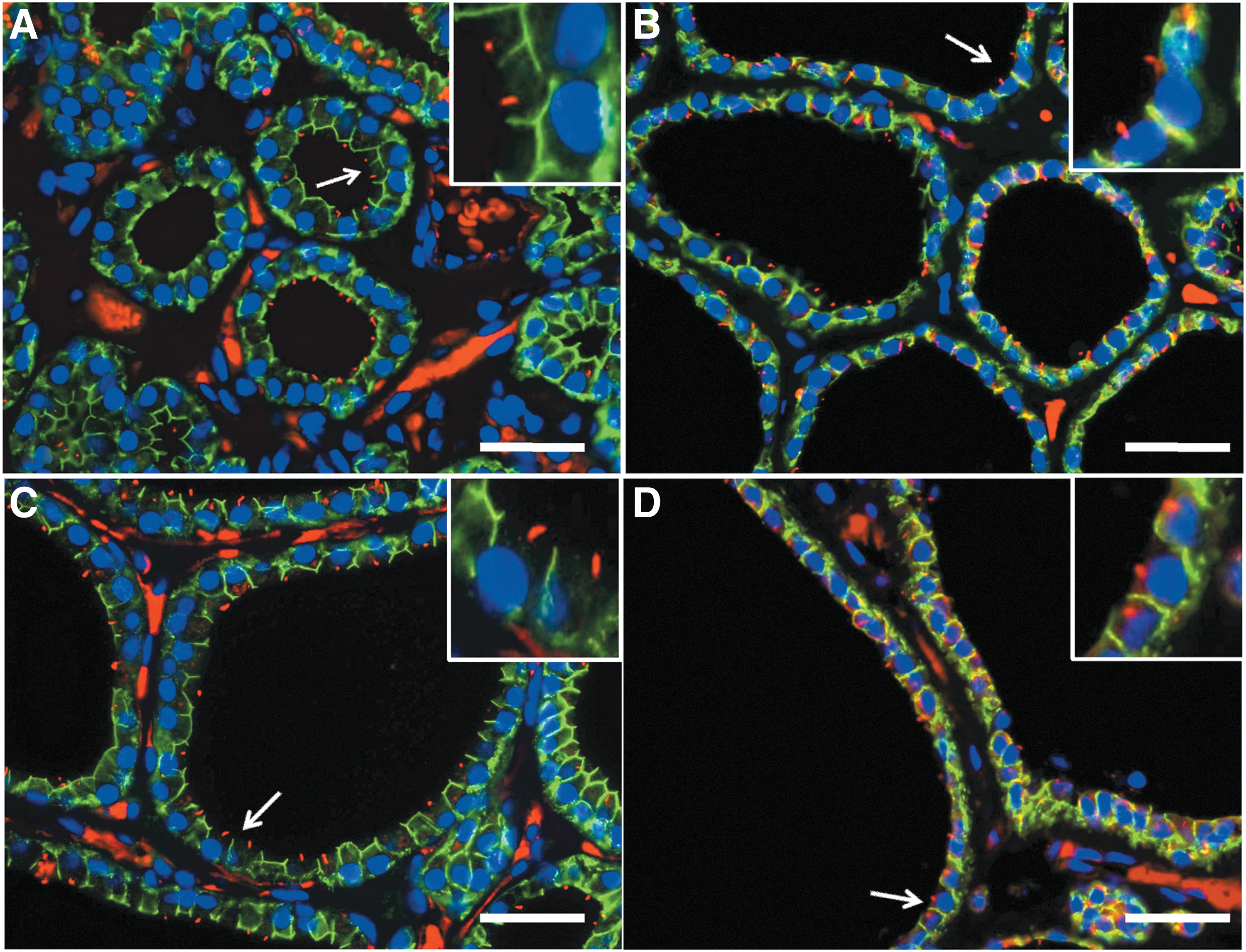
Distribution of PC in nodular thyroid hyperplasia using double immunofluorescence (E-cadherin, green; acetylated α-tubulin, red; nuclear counterstaining with DAPI, blue). PC are observed in small follicles (

Distribution of PC in GD using double immunofluorescence (E-cadherin, green; acetylated α-tubulin, red; nuclear counterstaining with DAPI, blue). PC are readily identified in small follicles (
Morphometrical analysis of PC frequency
In NT glands, 76.09 ± 7.31% of the thyrocytes showed PC, with non-statistically significant differences among follicles of different sizes (Fig. 5A).

Percentage of follicular cells exhibiting PC in the different patterns of thyroid follicles in NT (
In thyroid sections of NH, the percentage of thyrocytes exhibiting PC decreased to 51.16 ± 11.69%, with non-statistically significant differences among follicles of different sizes (Fig. 5B). When these data were compared with those found in NT glands, a highly significant difference appeared (p < 0.001; Fig. 6).
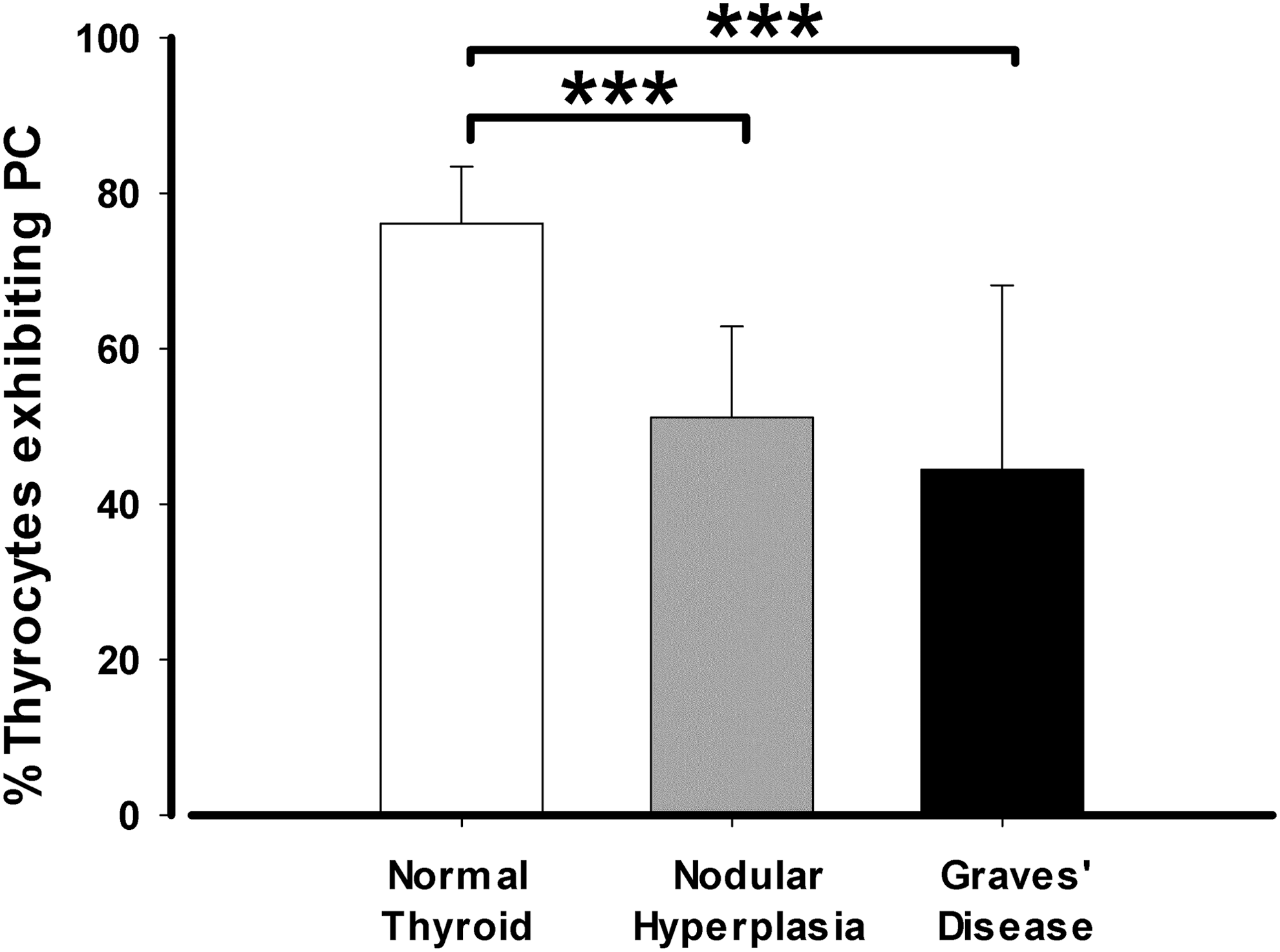
Comparison of the percentage of follicular cells exhibiting PC among NT, NH, and GD. The frequency of cilia is higher in NT, followed by NH and then GD. The comparisons among groups are significant. The results are presented as percentage of follicular cells presenting PC, and are expressed as the mean ± SD. Data were compared using one-way ANOVA multiple-comparison versus control. ***p < 0.001.
In thyroid sections of GD, the ciliary frequencies decreased even further to 44.43 ± 23.70%, with the high standard deviation being a consequence of the variety of follicular patterns (Fig. 5C). The average value was statistically significant in comparison to that of NT (p < 0.001; Fig. 6). In relation to intra-group differences in the percentage of thyrocytes presenting PC among the various histological patterns characteristics of GD, no significant differences were found between rounded follicles of different sizes (49.71.0 ± 4.60%) and papillary follicles (45.31 ± 19.89%). However, a significant difference was found when those patterns were compared to hyperfunctioning follicles (16.01 ± 4.86%; p < 0.001; Fig. 5C).
Furthermore, in GD, the percentage of PC-exhibiting thyrocytes in “normal rounded-follicle areas” was markedly different among cases, consistent with the persistence of signs of thyroid biosynthetic hyperactivity after long-term antithyroid drug treatment. Thus, PC frequency in those “normal areas” was significantly lower in patients who exhibited any signs of hyperactivity (undetectable thytrotropin [TSH] levels, remaining hyperactive follicle foci, or high serum thyroxine [T4] levels either during the treatment follow-up or prior to surgery) compared to patients showing no signs of hyperactivity (Fig. 7A).
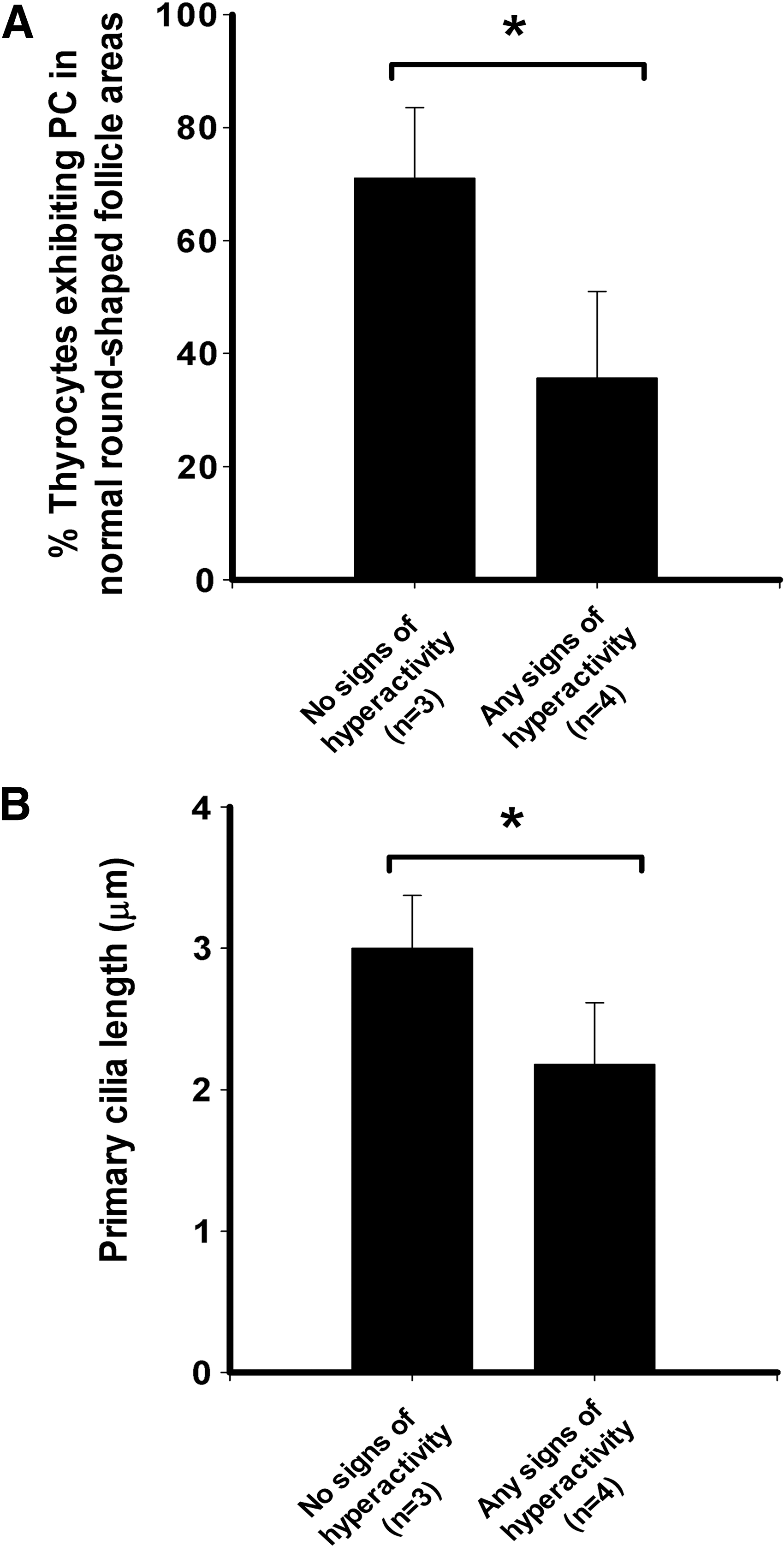
Comparison of percentages of follicular cells exhibiting PC (
Analysis of PC length
The length of PC showed characteristics alterations according to the pathological functional state of the thyroid gland (Fig. 8). Specifically, in NT glands, the normal thyrocytes showed the longest PC, with an average length of 3.93 ± 0.90 μm and a maximal length of 5.04 μm. In contrast, thyrocytes in NH exhibited the shortest PC, with an average length of 2.02 ± 0.35 μm and a maximal length of 3.24 μm, and the data were statistically significant in comparison to controls (p < 0.001). Finally, in GD, the average length of PC reaches a value of 2.42 ± 0.48 μm, with a maximal length of 3.49 μm, which was significant when compared to NT (p < 0.001). In all groups, no intra-group differences among follicles of different sizes were detectable. Nevertheless, it is necessary to specify that in the GD group, as in hyperfunctioning follicles PC are rarely observed, most PC in this group were measured in the rest of the thyroid follicles, implying a favorable bias in the final length of the PC, which would mean that the true length could be even lower.
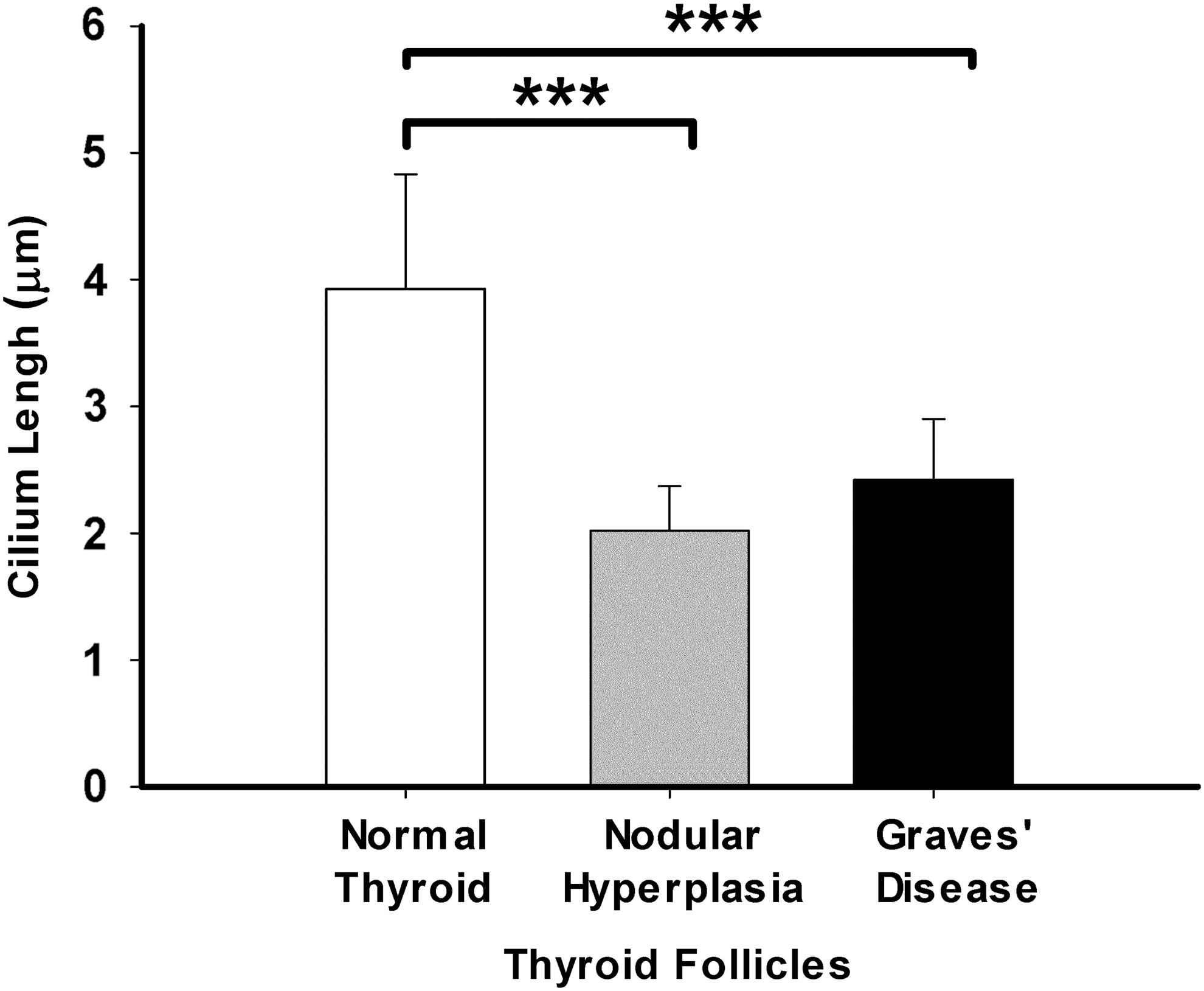
Quantitative changes in the PC length among NT, NH, and GD. NT show the longest PC of all groups, followed by thyrocytes in GD and thyrocytes from NH, which exhibit the shortest PC. The differences among groups were statistically significant. The results are expressed as the mean ± SD. ***p < 0.001.
Finally, and similarly to what is described above for PC frequency, axonemal lengths were also significantly lower in the “normal areas” adjacent to hyperactive follicle foci in those GD cases that showed hyperfunctioning signs (Fig. 7B).
Ultrastructure of PC
TEM analysis of specimens of NT glands revealed a sporadic detection of PC, located on the apical surface of follicular cells protruding into the colloid (Fig. 9A1). Usually, only one PC per cell was observed. Proximally, the cilium ended in a typical basal body, in whose proximity a centriole was observed (Fig. 9A2). When NT samples were observed by SEM, follicular cells showed a polyhedral outline with four to seven sides. The cell surfaces, which were generally convex, presented numerous microvilli and, emerging among them from the geometric center of the thyrocyte, one PC protruding into the colloid (Fig. 10A).

Ultrastructure of NT gland (

Histological aspects of NT gland (
In NH thyroid glands, PC were detected more sporadically than in controls when observed by TEM, likely as a consequence of the larger size of most thyroid follicles (Fig. 9B). The differences in follicular size were striking when observed by SEM (Fig. 10B1). However, PC were easily distinguished, preferably in medium-sized follicles, although they tended to disappear when the thyrocytes acquired a squamous-like appearance (Fig 10B2 and B3).
In GD, the concurrent use of TEM and SEM confirmed the heterogeneity among follicles. There were astounding images of follicles bearing papillae (Fig. 10C1–C2), or complex mosaics of follicular surfaces with variable morphology, such as the presence of rather dome-shaped cells and numerous microvilli (Fig. 10C3 and C4). Despite that complexity, PC could be observed emerging among microvilli sporadically by TEM (Fig. 9C), and frequently by SEM (Fig. 10C3–C4), although not as evident as in NT glands.
Discussion
The present study shows differences in the frequency, distribution, and morphology of PC in human thyroid functional pathology compared to NT glands. Thyroid tissue from both GD and NH showed lower ciliary frequencies and shorter axonemal lengths when tissue samples were analyzed as a whole and compared to the control group. Moreover, in the context of the pronounced tissue heterogeneity characteristic of NT itself and particularly of functional thyroid pathology, the different follicular patterns also exhibited changes in ciliogenesis, with a generally lower frequency of PC in zones with altered follicles compared to those areas where normal-appearance follicles predominate, with PC almost absent in hyperfunctioning areas of GD. Finally, when lower ciliary frequencies and axonemal lengths were noticeable in the apparently normal areas trapped in GD thyroid samples, a poorer response to antithyroid drug treatment was observed.
In contrast to motile cilia, PC are rarely described in the evaluation of pathology specimens because PC are not readily identifiable with standard stains such as H&E. Therefore, the first report in which the presence of PC was analyzed in thyroid pathology by using alternative methods, such as SEM and TEM, was not published until 1987 by Nesland et al. (41). These authors qualitatively appreciated that almost every follicular cell possessed PC in the NT as well as in most goiters, but PC occurred less frequently in GD. Additionally, PC showed a gradual decrease from normal glands compared to adenomas, well-differentiated thyroid carcinomas, and anaplastic carcinomas (41). Later, the presence of PC was reported in the thyroid gland of different mammals, as well as in different human thyroid cell lines, using both double IF and electron microscopy (12). The use of IF to identify PC provided the tools to analyze more extensive areas from paraffin-embedded thyroid samples, added to the additional advantage of applying quantitative methods to assess changes in PC frequency and length objectively, as was later done by Lee et al. (26) who studied the distribution of PC in different thyroid pathology and cell lines. Specifically, they described decreased frequencies of ciliated follicular cells from Hürthle cell carcinoma and the oncocytic variant of PTC, but reported no differences in ciliogenesis in the conventional variant of PTC, follicular carcinoma, Hashimoto's thyroiditis, and benign NH when compared to NT tissue. In the present study, however, after evaluating >1300 thyroid follicles and 43,000 thyrocytes, a high degree of intra- and inter-case variability is shown in the frequency and length of PC in functional thyroid pathology.
The present study found shorter average PC lengths than those reported before using confocal microscopy (3.93 ± 0.90 μm vs. 7.3 ± 1.2 μm) (12). Due to epifluorescence microscopy limitations related to measuring PC correctly that are not perfectly oriented in the horizontal plane, average PC length are likely underestimated, so that the results should be interpreted as a comparison between one functional group to another. In this sense, the average PC lengths presented here are comparable to those reported by Lee et al. (26) using a similar epifluorescence based morphometric approach.
As previously shown, thyroid PC are located at the apical surface of almost every thyrocyte protruding into the follicular lumen (12). Taking advantage of this unique localization, the ciliary membrane might possess specific receptors that could sense in some specific way the level of iodinated Tg stored in the colloid, the oxidative charge associated with the outer surface of the plasma membrane, or other processes taking place in the follicular lumen related with the proteolysis of Tg and consequently with thyroid hormone synthesis (27). These sensory activities would be coupled in coordination with the biosynthetic process through specific intracellular downstream signaling pathways. In line with this context, Szumska et al. recently found evidence that trace amine-associated receptor 1 (Taar-1), a G-protein-coupled receptor, is confined at the cilial extension of thyroid epithelial cells, where it could mediate rapid membrane-initiated effects of thyronamines, the endogenous decarboxylated and deiodinated relatives of the classical thyroid hormones T4 and T3 (42). They proposed that thyronamines can activate Taar-1 by acting from within the follicular lumen as part of a nonclassical mechanism of thyroid autoregulation. Furthermore, using Taar1-deficient mice, this same group also demonstrated that thyroid epithelia are characterized by a vesicular localization of the TSH receptor, which suggests that Taar1 signaling is necessary to maintain normal TSH receptor signaling (43).
According to Colin et al. (44), hyper- and hypofunctioning follicles differ in the presence and/or the amount of dense colloid-containing globules where older molecules of Tg are stored. Under increased TSH stimulation, this compact form of colloid, present in resting hypofunctioning AFUs, can be fragmented and metabolized into soluble Tg molecules to synthesize thyroid hormones (45 –48). Solubilization of Tg is extracellularly facilitated by cysteine proteinases such as cathepsins B, K, and L, which are proteolytically active at the luminal surface of thyrocytes (49 –51). Interestingly, Qatato et al. have demonstrated that Taar-1 is involved in the regulation of cathepsin-mediated proteolytic processing of Tg, establishing a definitive relationship between Taar1 localization at the cilial membrane and thyroid hormone synthesis (43).
Defective ciliogenesis might be associated with failure in many processes of thyrocyte function, such as loss of apicobasal polarization of the follicular epithelium, increased intrafollicular oxidative stress, hyperstimulation, or increased proliferation rate, depending on the different outcomes of thyroid disease, and on the genetic or biochemical background of the patient. Moreover, because of the complex biosynthetic mechanism of thyroid hormones, thyrocyte activity is associated with marked apical membrane turnover (exo- and endocytosis), so that ciliary dynamics could likely be affected in hyperactive epithelial cells (52 –54). Furthermore, PC are highly dynamic structures that disassemble and reassemble coupled with cell division (55,56). Thus, recent evidence shows that dysregulation of the cell cycle, as it occurs in cancer cells for example, can be associated with ciliary loss (26,57,58). Accordingly, shorter axonemal average lengths and perhaps lower ciliary frequencies could be due to increased cilium dynamics associated with either high division rates and/or high apical membrane turnover intrinsic to thyroid hyperplasia, as illustrated by hyperplastic foci characteristic of GD samples. The present study found shortened PC and lower PC frequencies in HN and GD groups when compared to NT. However, with regard to PC lengths, differences not only affected average but also maximal axonemal extension. These results support the hypothesis that altered ciliogenesis is present in functional thyroid pathology and may be responsible for the observed ciliary loss and shortened axonemal lengths. In this context, it was observed that when the “normal-appearance areas” of GD thyroid samples showed decreased ciliogenesis, it was accompanied by signs of residual thyroid hyperactivity after long-term antithyroid drug treatment.
In conclusion, the present study shows that differences in thyroid primary ciliogenesis are associated with functional pathology and moreover with thyroid tissue heterogeneity. The observations suggest that the contribution of PC to thyroid function, including their role in the complex mechanism of thyroid hormone biosynthesis, in the self-regulation of the AFUs, and eventually in the origin of functional and neoplastic disease, is an attractive hypothesis that requires further investigation.
Footnotes
Acknowledgments
This work was supported by grants from the Consejería de Innovación, Ciencia y Empresa (refs. CTS-439/2017). The authors thank Mr. John Brown for corrections to the English language, Mr. Marcos Ortega for histological sample processing, and the Microscopy Service of CITIUS (General Research Services, Seville University, Spain) for technical assistance with electron microscopy studies.
Author Disclosure Statement
No competing financial interests exist.