
Abstract
Background:
Penta-brominated diphenyl ethers (PentaBDEs) are endocrine-disrupting chemicals that structurally resemble thyroid hormones and were widely used as flame retardants in household consumer products from 1975 to 2004. Polybrominated diphenyl ethers (PBDEs) cross the placenta, and evidence suggests that for many children, body burdens may peak during the toddler years. This study aimed to understand the impact of exposure timing by examining both pre- and postnatal exposure to BDE-47, the predominant penta-brominated diphenyl ether congener detected in humans, in relation to thyroid hormone parameters measured during early childhood.
Methods:
The Columbia Center for Children's Environmental Health Mothers and Newborns Study is a prospective birth cohort of African American and Dominican maternal–child pairs. Pregnant women were recruited from two prenatal clinics in Northern Manhattan and the South Bronx between 1998 and 2006. Participants included 158 children with (i) plasma PBDE concentrations measured at birth and in the toddler years (age 2–3 years), and (ii) serum thyroid parameters measured at three and/or five years of age. Outcomes included concentrations of serum thyrotropin, free thyroxine, and total thyroxine.
Results:
Children with high exposure to BDE-47 during the prenatal period (−17% [confidence interval −29 to −2]) or toddler age (−19% [confidence interval −31 to −5]) had significantly lower geometric mean thyrotropin levels compared to children with low BDE-47 exposure throughout early life. Associations with thyroxine were also inverse; however, they did not reach statistical significance at the p = 0.05 level. Sex-stratified models suggest associations with postnatal exposure may be stronger among boys compared to girls.
Conclusions:
The thyroid regulatory system may be sensitive to BDE-47 during pre- and postnatal periods.
Introduction
Endocrine-disrupting chemicals (EDCs) contribute substantially to human morbidity and are estimated to result in hundreds of billions in costs per year (1). EDCs are defined by their ability to cause changes in endocrine function and consist of several classes of chemicals with varying structures, actions, and endocrine targets (2). Many EDCs were introduced into U.S. commerce beginning in the 1970s to augment food packaging, personal-care products, and other household items (3). For example, following reports indicating improperly extinguished cigarettes were the leading cause of household fires (4), polybrominated diphenyl ethers (PBDEs) began to be used as flame retardants in electronics and household furnishings in 1975 (5). PBDEs were used as three technical mixtures known as penta-brominated diphenyl ether (PentaBDE), octa-brominated diphenyl ether, and deca-brominated diphenyl ether, each comprised of several congeners. PentaBDEs, which are estimated to make up 90% of the human body burden, were primarily applied to products containing polyurethane foam, including couches, car seats, carpet padding, and other upholstered items (6). PentaBDEs were often used in large volumes. For example, reports indicate they comprised up to 3% by weight of the polyurethane foam contained within a couch (7). During manufacturing, PBDEs are not chemically bonded to base polymers and thus have a propensity to migrate into the indoor environment and settle in house dust (8). Human exposure occurs primarily through incidental ingestion of dust, placing young children at risk for elevated exposure due to their frequent hand to mouth behavior and often close proximity to the floor (5,9 –11). Owing to their lipophilic properties, PBDEs have long half-lives (PentaBDE congeners: 1.6–6.5 years) (12) and are known to penetrate the fat-soluble placenta, as well as partition into breast milk (13,14). It is estimated that >46,000 tons of PentaBDEs were used in North America until their phase-out in 2004, leading to nearly ubiquitous exposure and body burdens that are the highest in the world (15,16). Despite the phase-out, exposure to PBDEs continues because of their resistance to environmental degradation and ongoing release from consumer products that are infrequently replaced (15).
PBDE congeners consist of a diphenyl ether backbone around which varying numbers of bromine atoms are attached (5). This molecular structure closely resembles that of the halogenated (iodine) thyroid hormones triiodothyronine (T3) and thyroxine (T4) (17), supporting the putative interaction of PBDEs with thyroid hormone transport proteins, receptors, and/or degradation enzymes (18). Thyroid hormones bind to receptors in nearly every organ in the human body and play critical roles in the regulation of growth, metabolism, and brain development (19). In vitro research indicates hydroxylated metabolites of BDE-47, the predominant congener detected in humans, markedly inhibit the capacity of T3 to bind with receptors (20), and evidence from animal research suggests PBDE exposure alters thyroid hormone homeostasis (18) as well as other thyroid-dependent processes. For example, research conducted in Xenopus laevis has demonstrated BDE-47 exposure arrests thyroid-dependent metamorphosis of tadpoles into froglets and disrupts thyroid hormone-related gene expression in the brain (21,22). Likewise, research conducted in avian and murine models has consistently demonstrated associations between prenatal exposure to PentaBDE congeners, including BDE-47, with reductions in circulating levels of T4 (reviewed by Costa et al. and Birnbaum and Staskal) (18,23).
Widespread research supports classification of PBDEs as developmental neurotoxicants (11,24 –27), with disruption of thyroid hormones as a leading putative mechanism underlying observed relationships (18,28). However, despite convincing evidence from in vitro and animal research, results from studies investigating PBDEs in relation to thyroid hormone function in humans include a mix of negative, positive, and null associations (29 –34). Notably, previous studies have measured thyroid hormone parameters in maternal blood (29,30,34) collected during pregnancy or parturition, cord blood (29,32,34), or infant blood (31,32) collected within hours to weeks of birth—periods when transient yet substantial endocrine system changes occur, including profound alterations to the thyroid regulatory system (35). It is plausible that inconsistencies across previous human studies are partially attributable to misclassification introduced by the timing of thyroid hormone measurement. The present study addresses this limitation by examining plasma PBDE concentrations in samples collected at birth and during the toddler years (age 2–3 years) in relation to thyrotropin (TSH), free T4, and total T4 levels measured in serum samples collected during early childhood (age 3–5 years). Based on results from animal research, it was hypothesized that prenatal exposure to BDE-47 would be associated with lower T4 levels at birth (reviewed by Costa et al. and Birnbaum and Staskal) (18,23). It was further hypothesized that these associations would act through a programming pathway, leading to effects that persist throughout childhood. Because of the typically higher exposure of young children compared to fetuses (via direct interaction with the external environment), it was also hypothesized that children with low prenatal exposure but high postnatal exposure would show evidence of a dysregulated thyroid regulatory system during childhood. This is the largest prospective study to examine both pre- and postnatal exposure to PBDEs in relation to thyroid endpoints.
Methods
Study sample
The sample includes a subset of participants enrolled in the Columbia Center for Children's Environmental Health (CCCEH) Mothers and Newborns birth cohort, which recruited African American and Dominican women from New York City between 1998 and 2006. Women were ineligible for study participation if they were outside the ages of 18–35 years, initiated prenatal care after the 20th week of pregnancy, had a multiple pregnancy, used tobacco products or illicit drugs, had diabetes, had hypertension, or were HIV positive. Women were considered fully enrolled if a maternal or umbilical cord blood sample was collected at the child's delivery. During pregnancy and at each postnatal study visit, a bilingual research worker conducted a structured interview to collect information about socio-demographic and life-style factors. At delivery, study staff collected umbilical cord blood; at follow-up visits at the ages of two, three, and five years, a pediatric phlebotomist collected child venous blood. All samples were transported to the CCCEH laboratory immediately following collection, where the buffy coat, packed red blood cells, and plasma were separated and frozen at −70°C. Additional details describing the cohort design, recruitment, and follow-up have been previously published (36 –38).
At delivery, 727 mothers remained eligible and were fully enrolled in the cohort. At the two-, three-, and five-year follow-up visits, 566 (78%), 562 (77%), and 551 (76%) maternal–child pairs remained in the study, respectively. At these postnatal follow-up visits, blood was collected from 92–98% of children. PBDE concentrations were measured in all available stored cord plasma samples and in early childhood samples among children with follow-up data (cord plasma n = 327, 2-year follow-up n = 43, 3-year follow-up n = 102, or both 2- and 3-year follow-up n = 13). Thyroid hormone parameters were measured in 185 of the 327 children with a measure of PBDE concentrations at three years of age (n = 112), five years of age (n = 35), or both three and five years of age (n = 38; Fig. 1). The study protocol was approved by the Institutional Review Board of Columbia University Medical Center. It was determined at the Centers for Disease Control and Prevention (CDC) that the agency was not engaged in human subject research. Before each study visit, mothers gave written informed consent for herself and for her child.
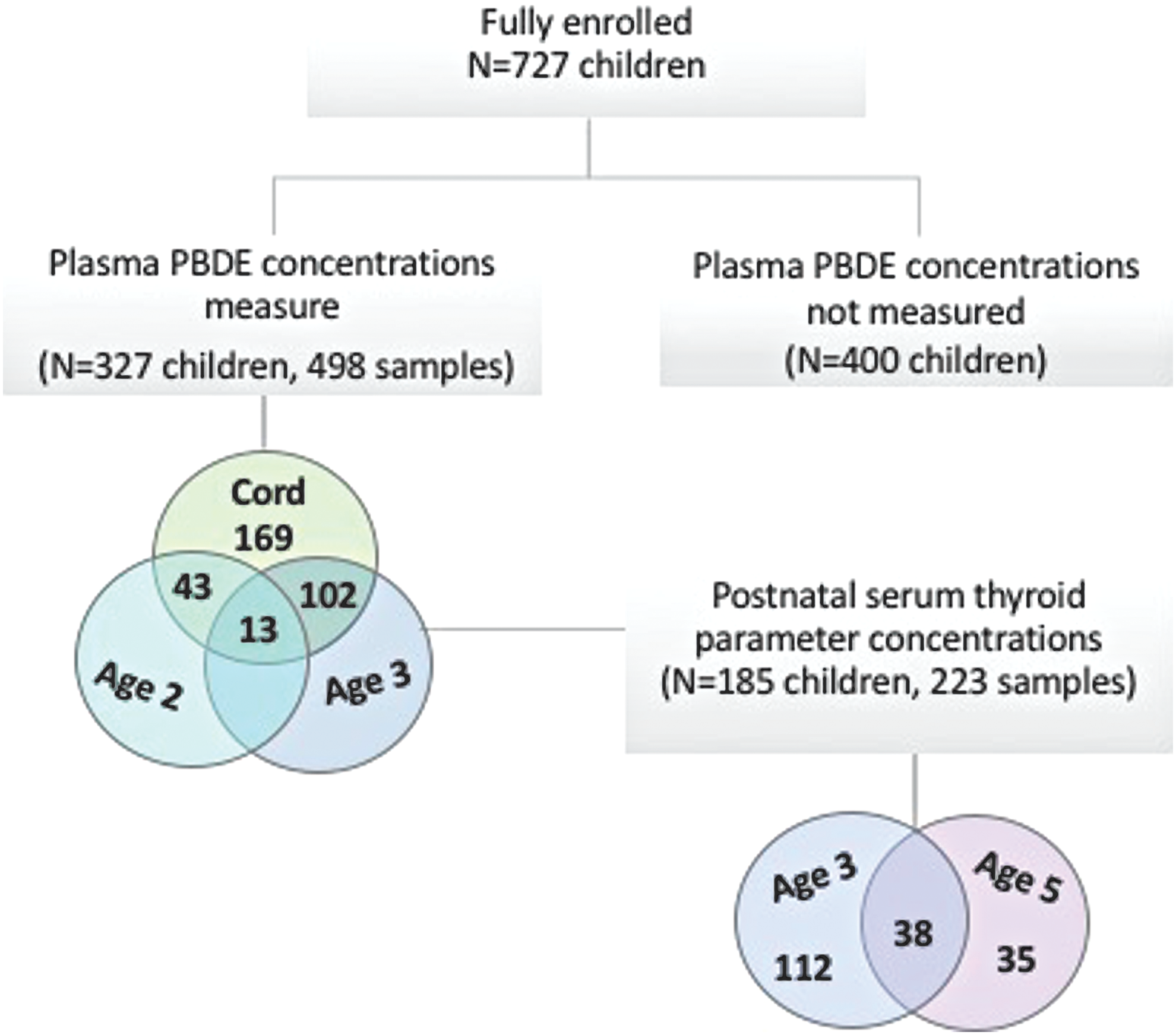
Flow diagram of participant selection from the Columbia Center for Children's Environmental Health Mothers and Newborns birth cohort study. Color images are available online.
PBDE analysis
The CDC's Persistent Organic Pollutants Biomonitoring Laboratory measured PBDE concentrations in umbilical cord and venous plasma samples. Detailed analytic methods are available elsewhere (39,40) and information pertaining to analysis of PBDEs in this cohort has been previously described (9,41). Briefly, samples were fortified with internal standards followed by automated liquid–liquid extraction using a Gilson 215 liquid handler (Gilson, Inc., Middleton, WI). Final analytical determinations were made by gas chromatography isotope dilution high-resolution mass spectrometry using a DFS instrument (Thermo Fisher Scientific, Bremen, Germany). Each analytical batch was comprised of method blanks (n = 3), quality-control samples (n = 3), and study samples (n = 26). All reported data were subtracted from the median concentration detected in method blank samples. Co-extracted lipids were removed using a silica gel/silica sulfuric acid column with automation on a Rapid Trace SPE workstation (Biotage, Uppsala, Sweden), and total cholesterol and triglycerides were determined on a Roche Hitachi 912 Chemistry Analyzer (GMI, Inc., Ramsey, MN). Total cord blood lipid levels, including unmeasured free cholesterol and phospholipids, were estimated by summation of individual lipid components using an umbilical cord blood–specific formula (total cord blood lipids = 2.657 × total cord blood cholesterol + cord blood triglycerides +0.268, in g lipids/L plasma; A. Sjodin, pers. commun., November 2016). Child total blood lipids were estimated using the short formula developed by Phillips et al. (42). PBDEs were examined as a lipid standardized variable in all models (ng/g lipid).
Thyroid hormone analysis
TSH, free T4, and total T4 were measured in child serum samples by the Clinical and Epidemiologic Research Laboratory at Boston Children's Hospital. All analyses were performed by automated immunoassay using a competitive electrochemiluminescence detection system (Roche Diagnostics, Indianapolis, IN). The lowest detection limits were 0.005 μIU/mL, 0.26 pmol/L, and 5.4 nmol/L for TSH, free T4, and total T4, respectively. Day-to-day imprecision values ranged from 1.8% to 5.4% for 0.09–3.96 μIU/mL of TSH, from 3.5% to 6.6% for 8.75–50.70 pmol/L of free T4, and from 3.0% to 6.9% for 33.4–237 nmol/L of total T4. Maternal iodide concentrations, an essential substrate for thyroid hormone biosynthesis (43), were measured in maternal spot urine samples collected during the third trimester. Before performing statistical analyses, iodide was adjusted for specific gravity to control for variation in urinary dilution.
Statistical analysis
The study focused on BDE-47 (percent detected: 80%), which was the only congener for which cord plasma concentrations were detectable in >50% of samples; concentrations in child plasma (age 2–3 years) were detectable in 99% of samples. Consistent with other studies, in the samples, cord plasma BDE-47 concentrations were moderately to highly correlated with the other primary congeners that comprise the PentaBDE formulation (Spearman's ρ: BDE-99, 0.83; BDE-100, 0.76; BDE-153, 0.47; p < 0.01). As previously described (41), a distribution-based approach was used to multiply impute values for plasma BDE-47 concentrations below the sample-specific limit of detection (LOD), which is determined by the sample's volume and lipid content.
Latent class growth analysis (LCGA) was performed using the SAS Proc Traj procedure (44) to estimate trajectories of BDE-47 concentration between birth and three years. LCGA is a group-based modeling technique that empirically clusters individuals with a shared temporal pattern of change for a given characteristic (i.e., change in PBDE concentration over early life) (45). Before estimating trajectories, continuous BDE-47 concentrations (ng/g lipid) were log10-transformed to improve normality of the distribution, and non-detected concentrations were replaced with the sample-specific mean value across the 10 imputed data sets. Models were iteratively tested with varying numbers of groups (n = 2–5) and shapes (linear–cubic), and the optimal number of trajectories was determined based on: (i) visual confirmation of distinct trajectories, each of which comprised >10% of the data; (ii) evaluation of the Bayesian Information Criterion; and (iii) evaluation of the average posterior probability of group membership. Additional details describing LCGA model fitting are provided in Supplementary Table S1.
Multivariable linear regression was used to examine associations between trajectories of BDE-47, treated as a categorical variable, and thyroid hormone parameters collected between three and five years. The generalized estimating equations (GEE) approach was used with an exchangeable working correlation to account for repeated thyroid measures within a child over time. An exchangeable working correlation was selected based on evaluation of the empirical correlation matrix, which did not indicate an autoregressive relationship, as well as evaluation of the quasi-likelihood information criterion. In all models, TSH, free T4, and total T4 were expressed as continuous variables, and TSH was log10-transformed to improve normality of the distribution. Models including an interaction term between age and BDE-47 trajectory did not indicate that the association between PBDEs and thyroid parameters significantly varies by age at blood collection. Separate models were further examined for thyroid hormone parameters measured at three and five years of age in sensitivity analyses.
Intra-individual thyroid hormone concentrations decrease with age (46). Therefore, the study a priori included exact age at blood draw as a time-varying covariate. Directed acyclic graphs (DAGs) were constructed based on substantive knowledge and previously published research to identify the minimal set of covariates sufficient to estimate the unconfounded effect of PBDEs on thyroid hormone parameters, which included only race/ethnicity (Supplementary Fig. S1). The set of potential confounders considered included: sex, race/ethnicity (African American/Dominican), date of birth, gestational age (in weeks), birth weight (in grams), prenatal environmental tobacco smoke exposure (yes/no, as previously described) (47), breastfeeding history (<12 weeks/≥12 weeks), parity (nulliparous/multiparous), relationship status (unmarried/married or with the same partner for seven years or more), maternal age (in years), material hardship (none/unable to afford food, clothing, or housing), maternal education (less than high school/high school or equivalent), and maternal employment (employed/not employed). All variables relating to the mother or household were collected during the prenatal period, and variables relating to the delivery were extracted from hospital medical records. Information on breast-feeding history was collected at 3-, 6-, 12-, 24-, and 36-month follow-up visits. In sensitivity analyses, the influence of covariate selection was further evaluated by examining a priori (age at blood draw only) and fully adjusted models.
Given sex differences in the incidence of many thyroid-related diseases (48,49), potential effect modification by child sex was explored using cross-product terms and sex-stratified models. The influence of maternal iodide status during pregnancy was examined by stratifying participants by the pregnancy-specific threshold for population iodine sufficiency (≥150 μg/L) and examining models within each stratum (50). Finally, to compare the results with findings from other cohort studies, the GEE approach was used to examine associations between plasma BDE-47 concentrations treated as a continuous, log10-transformed variable measured in cord blood or three-year-old blood in relation to repeated thyroid hormone parameters. Statistical analyses were performed using SAS v9.4 (SAS Institute, Cary, NC) or RStudio v0.99.891 and constructed DAGs using DAGitty v2.3 (51).
Results
Table 1 presents characteristics of the study population. All maternal–child pairs were African American (45%) or Dominican (55%), and at delivery 37% of mothers had less than a high school education, 76% were not in a stable relationship, and 36% reported experiencing material hardship. Socio-demographic and life-style characteristics were similarly distributed between children included in the analysis and those excluded due to missing PBDE or thyroid hormone data, with the following exceptions: the excluded sample had a higher proportion of Dominican participants (68% vs. 55%; p < 0.01) and fewer nulliparous mothers (42% vs. 51%; p = 0.04), and on average newborns had lower birth weights (mean difference: 119 g; p < 0.01). The difference in birth weight may reflect racial/ethnic variation between the included and excluded samples, as African American newborns weighed on average 148 g less than Dominican infants (p < 0.01). Among included children, BDE-47 concentrations (at birth, 2 years of age, and 3 years of age) and thyroid hormone parameters (at 3 and 5 years of age) were not significantly associated with parity or birthweight. At two and three years of age, BDE-47 concentrations were significantly higher among children included in the analysis compared to excluded children (p = 0.05), which likely reflects that included children were more likely to be born earlier during the enrollment period (i.e., prior to the 2004 phase-out of BDE-47) to allow time to age into later blood draws (9).
Participant Characteristics, BDE-47 Concentrations, and Thyroid Parameter Levels Among Maternal–Child Pairs Included in the Analysis (n = 185)
Geometric mean.
Included and excluded significantly different at p = 0.05.
BDE, brominated diphenyl ether; ETS, environmental tobacco smoke; T4, thyroxine; TSH, thyrotropin.
BDE-47 was detected in 80% of cord plasma samples and 99% of toddler plasma samples. The lower detection frequency in cord blood is consistent with results from an independent cohort based in New York City (BDE-47: 81%) and a cohort based in Baltimore (BDE-47: 90%), which both measured PBDE concentrations in cord blood (52,53). As expected, plasma BDE-47 concentrations were significantly lower at birth (14.2 ± 0.9; n = 327) compared to the toddler years (2 years of age: GM±GSE = 37.8 ± 5.8, n = 56, paired t-test using log10-transformed BDE-47 in ng/g lipids, t = −4.07, p = 0.0002; 3 years of age: GM ± GSE = 32.0 ± 3.1, n = 115, paired t-test using log10-transformed BDE-47 in ng/g lipid, t = −6.90, p < 0.0001). BDE-47 in cord plasma correlated poorly with BDE-47 in child plasma measured at two years of age (ρ = −0.03, p = 0.82) or three years of age (ρ = 0.09, p = 0.36). However, BDE-47 measured at two years of age was strongly correlated with BDE-47 measured at three years of age (ρ = −0.79, p < 0.01). Exposure percentiles for each group are provided in Supplementary Table S2. As illustrated by Figure 2A, the best-fitting LCGA model revealed three trajectories of BDE-47 exposure characterized by (i) persistent low (34%), (ii) high-decreasing (28%), and (iii) low-increasing (38%) plasma concentrations across early childhood.
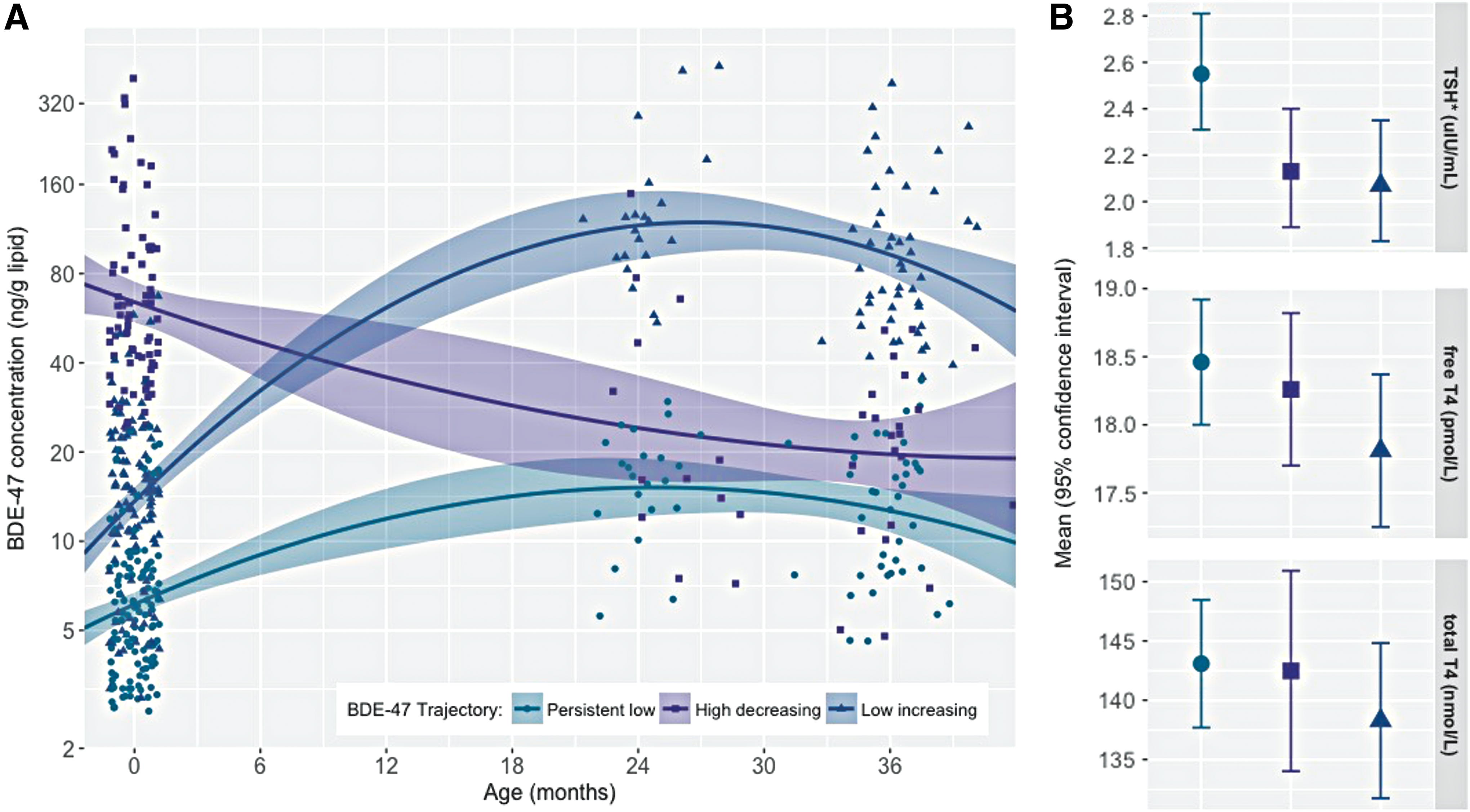
(
In models examining BDE-47 trajectories in relation to serum thyroid parameters measured between three and five years, children assigned to the high-decreasing or low-increasing trajectory had 17% [confidence interval (CI) −29 to −2] and 19% [CI −31 to −5] lower geometric mean TSH levels compared to children assigned to the persistent low trajectory, respectively. Associations between each of these trajectories and T4 levels (free and total) were also inverse. However, they did not reach statistical significance at the p = 0.05 level (Table 2, which presents estimates from GEE models, and Figure 2B, which plots adjusted mean thyroid parameter concentrations stratified by BDE-47 trajectory). Results from fully adjusted models, as well as models examining thyroid hormones at three and five years of age separately, did not substantially deviate from these results (see Supplementary Tables S3 and S4). Models examining cord plasma BDE-47 as a continuous variable are presented in Figure 3 and Supplementary Table S5.
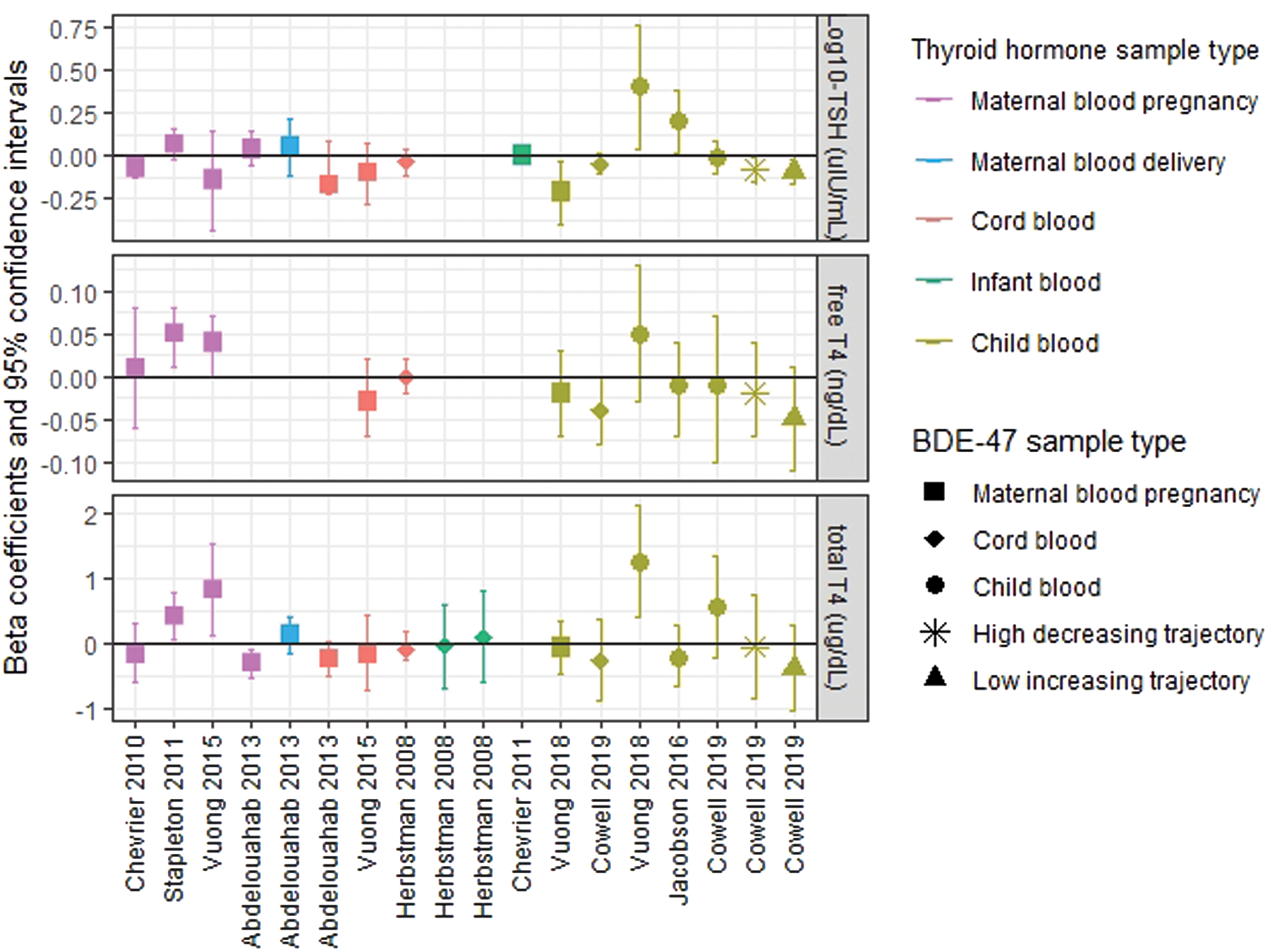
Results from multiple linear regression models examining associations between log10BDE-47 (ng/g lipid) and thyroid hormone parameters (log10TSH: μIU/mL; free thyroxine [T4]: ng/dL; total T4: μg/dL) measured as continuous variables reported by nine North American birth cohort studies. Herbstman et al. (32) and Stapleton et al. (33) applied a natural-log transformation to BDE-47 and TSH rather than a log10-transformation. Stapleton et al. (33) additionally natural-log transformed free T4. To facilitate comparison of the present results to others, final models expressing free T4 and total T4 were reanalyzed in units of ng/dL and μg/dL, respectively. Abdelouahab et al. (29) modeled free T4 measured in cord blood on a pmol/L basis. therefore, these results were excluded from the figure to accommodate the y-axis scale. Supplementary Table S8 presents summary data for BDE-47 and thyroid hormone parameters measured by each study. Color images are available online.
Age- and Ethnicity-Adjusted Associations Between BDE-47 Trajectories (ng/g Lipid) and Serum Thyroid Parameters
Data shown as β [CI]. 185 children and 223 observations.
CI, confidence interval.
No significant sex differences were observed in the proportion of children assigned to each trajectory. Likewise, thyroid hormone parameters did not significantly differ between girls and boys at three or five years of age (Supplementary Table S6). In sex-stratified models (see Supplementary Fig. S2 and Supplementary Table S7), the inverse association observed between the low-increasing BDE-47 trajectory (vs. persistent low trajectory) and childhood thyroid parameters (TSH and free T4) was augmented among boys (percent change TSH: −30 [CI −45 to −11]; p-interaction: 0.12; unit change free T4: −1.18 [CI −2.18 to −0.20]; p-interaction: 0.21) and attenuated among girls (percent change TSH: −8 [CI −26 to 13.3]; unit (nmol/L) change free T4: −0.19 [CI −1.19 to 0.82]). While the interaction terms did not reach statistical significance at the p = 0.05 level, these findings suggest sex may modify the association between postnatal BDE-47 exposure and thyroid hormone parameters. Given the relatively small sample size for investigating interactions, it will be important that these findings are replicated by other research groups.
Specific-gravity adjusted urinary iodide concentrations among the 115 mothers with an available urine sample ranged from 45.4 to 425.9 μg/L; 27% of mothers had a concentration below the pregnancy-specific threshold for population iodine sufficiency (150 μg/L). In age, ethnicity, and specific gravity adjusted models, no significant interaction was detected between BDE-47 and maternal urinary iodine status, treated as a continuous or categorical variable (<150 vs. ≥150 μg/L) for any thyroid hormone parameter.
Discussion
Compared to children with low cord plasma BDE-47 concentrations (GM ± GSD = 5.8 ± 0.4 ng/g lipid) that remained low throughout early childhood (GM ± GSD = 13.8 ± 1.2 ng/g lipid), children with high prenatal exposure (GM ± GSD = 66.6 ± 6.1 ng/g lipid) that decreased after birth had significantly lower circulating TSH levels measured between three and five years of age. TSH is a key effector and stimulus of the hypothalamic–pituitary–thyroid (HPT) axis, which maintains circulating thyroid hormone levels around an intra-individual set point. Briefly, low levels of circulating T3 and T4 stimulate the pituitary gland to release TSH, which in turn stimulates the thyroid gland to produce and secrete T3 and T4 (43,54). Evidence from animal models and human clinical studies suggests the set point around which this negative feedback mechanism responds may be partially determined during gestation (55 –58). The present findings suggest prenatal exposure to PBDEs may program a “reactive HPT axis” phenotype such that less TSH is required to stimulate production and release of adequate T4.
Children with low cord plasma BDE-47 concentrations (GM ± GSD = 13.8 ± 1.2 ng/g lipid) that increased during the toddler years (GM ± GSD = 106.9 ± 9.2 ng/g lipid) also had significantly lower TSH concentrations between three and five years of age compared to children with low cord plasma BDE-47 concentrations (GM ± GSD = 5.8 ± 0.4 ng/g lipid) that remained low during the toddler years (GM ± GSD = 13.8 ± 1.3 ng/g lipid). Interestingly, the magnitude of this association was stronger among boys, despite the finding of no significant difference in BDE-47 concentrations or TSH levels between girls and boys. Boys with high exposure during the toddler years also showed significantly lower free and total T4 levels compared to boys with persistent low BDE-47 exposure. The finding of depressed T4 levels is consistent with research conducted in murine models, which have consistently found PBDE exposure to be associated with reduced serum T4 levels (59). Putative mechanisms underlying this finding include PBDE interference with thyroid hormone transport and metabolism. For example, research conducted in mice suggests PBDEs induce upregulation of thyroid hormone metabolizing enzymes, resulting in enhanced clearance of T4 (60), and both animal and in vitro studies suggest PBDEs or their hydroxylated metabolites may bind and displace T4 from protein transporters, thereby disrupting circulating levels (18).
Several birth cohort studies (n > 100) have investigated cross-sectional associations between prenatal exposure to PBDEs and thyroid hormone parameters measured in maternal or cord blood collected during pregnancy, delivery, or early postnatal life with a combination of positive, negative, and null findings (Fig. 3 and Supplementary Table S8) (29 –34). Results from these studies are inconsistent and difficult to compare to the present findings, given variation in the measurement of thyroid hormones during developmental periods when normal fluctuations in HPT axis homeostasis occur. For example, an estrogen-induced elevation of thyroid-binding globulin and placental production of chorionic gonadotrophin triggers maternal T4 levels to increase sharply and TSH levels to fall during the first trimester of pregnancy, and during delivery, a stress and cold-evoked surge in TSH occurs in the newborn, followed by a reflexive increase in T4 over the next 24–48 hours (35,43,61).
Similar to studies focused on pregnancy and infancy, results from research investigating cross-sectional associations between PBDEs and thyroid dysregulation during childhood are inconsistent and difficult to compare due to differences in study design and variation in both the distribution of PBDE concentrations and congeners detected. Specifically, of the eight studies identified that examined postnatal exposure to PBDEs and thyroid parameters, three focused on special populations with unusually high exposure levels (i.e., children living and working near electronics recycling facilities in China) (62 –64), two detected unusually low PBDE concentrations for unexplained reasons (65,66), and one was conducted among a small sample (n < 30) of older children (aged 14–18 years) (67). Among 80 children admitted to a hospital for a non-endocrine-related disease between one and five years of age, Jacobson et al. detected a significant positive association between BDE-47 (ng/g lipid) and TSH but no significant associations with total or free T4 or T3, reverse T3, or T3 uptake (68). A number of factors could underlie the differences in the findings, including study design (cross-sectional vs. prospective), variation in age at time of thyroid parameter measurement (1–5 years vs. 3–5 years), or differences in the source populations (i.e., general population recruitment vs. hospital-based recruitment).
Only one other study has prospectively examined associations between both pre- and postnatal BDE-47 concentrations and thyroid hormone levels measured during early childhood. Using multiple informant models, Vuong et al. detected significant inverse associations between maternal log10-BDE-47 concentrations measured during pregnancy and ln-TSH measured at three years of age (β = −0.20 [CI −0.38 to −0.03]) among 158 maternal–child pairs living in Cincinnati, Ohio (69). Also consistent with our finding of a trend toward a stronger association between the early prenatal high BDE-47 trajectory and TSH among girls, Vuong et al. found that inverse associations between prenatal BDE-47 and ln-TSH were only statistically significant among girls. Vuong et al. found that inverse associations between prenatal BDE-47 and ln-TSH were only statistically significant among girls. While the present study did not detect evidence of a sex-specific effect between prenatal BDE-47 and free T4, Vuong et al. found a significant inverse association only among boys (69). Additionally, in contrast to the present observation of inverse associations between postnatal BDE-47 and ln-TSH, the researchers detected significant positive associations between serum BDE-47 measured at two years of age (n = 71) but not one (n = 77) or three years of age (n = 71), and both ln-TSH and total T4 measured at three years of age (69). Results of sex interactions with postnatal exposure were not reported.
Despite evidence indicating (i) sex-specific associations between PBDEs and thyroid gland function in birds (70), (ii) interactions with sex hormone receptors in fish and rodents (71 –73), and (iii) altered sex hormone levels in children (74) and pregnant women (75), few studies have investigated sexually dimorphic effects of PBDEs on thyroid hormone disruption. Specific mechanisms underlying the present observation of stronger associations with postnatal exposure among boys compared to girls are unknown. However, the high degree of overlap between the HPT axis and the hypothalamic–pituitary–gonadal axis, which regulates circulating sex hormone levels, suggests disruption in one system may have downstream consequences for the other (76). For example, evidence suggests hypogonadism in hypothyroid men reflects a hypothyroidism-induced blunted pituitary response to gonadotropin-releasing hormone secreted by the hypothalamus (77). Further, male reproductive organs, including the prepubertal testes, are thyroid-responsive tissues (78), and animal studies have demonstrated that experimentally induced hypothyroidism results in testicular damage, decreased testosterone concentrations, and arrest of sexual maturity (79,80). Research designed to investigate the effect of PBDEs on overlapping pathways central to both thyroid and reproductive hormone homeostasis is needed to understand more fully the mechanisms underlying the current sex-specific findings.
In addition to the longitudinal design, the present study has several strengths. First, PBDE concentrations were comparable to other geographically and temporally similar birth cohorts and reflect general population exposure (52,53). Additionally, the relatively large sample size allowed many potential confounders to be examined, as well as effect modification by child sex and maternal iodide status during pregnancy to be explored. Unfortunately, it was not possible to evaluate selenium, which is known to be an important determinant of thyroid status (81). Additional limitations include the lack of T3 levels, thyroid-binding protein levels, and PBDE metabolite data, which structurally resemble endogenous thyroid hormones more closely than parent congeners (18). Finally, thyroid parameters were analyzed by immunoassay, which may be affected by variation in serum thyroid-binding protein levels (43).
Overall, the findings suggest the thyroid regulatory system may be sensitive to disruption by PBDEs during both the pre- and postnatal periods. Pregnant women and young children should minimize exposure to these EDCs. While research on PBDE exposure intervention studies is limited, results from observational research suggest several behavioral modifications for reducing contact with dust may be effective in limiting PBDE exposure, including wet or damp mopping the home (9,41), wiping down plastic toys (82), vacuuming with a HEPA filter and/or wearing a dust mask while vacuuming, washing hands frequently (83), avoiding hand-to-mouth behaviors (82) (i.e., thumb sucking, nail biting), and purchasing flame-retardant-free furniture and furnishings, which can be readily identified in the United States by examining product tags.
Footnotes
Acknowledgments
We thank Dr. Lori Hoepner for assistance with data management and Pat Vuguin for review of manuscript drafts. This work was supported by National Institutes of Health (grant numbers R01ES021806, R01ES013543, P50ES009600, T32ES023772 and T32ES007322) and Environmental Protection Agency FP-91779001.
The findings and conclusions in this report are those of the authors and do not necessarily represent the official position of the Centers for Disease Control and Prevention. Use of trade names is for identification only and does not imply endorsement by the CDC, the Public Health Service, or the U.S. Department of Health and Human Services. This publication was developed under STAR Fellowship Assistance Agreement no. FP-91779001 awarded by the U.S. Environmental Protection Agency (EPA). It has not been formally reviewed by the EPA. The views expressed in this publication are solely those of the study authors.
Author Disclosure Statement
The authors report no conflicts of interest in this work.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
Supplementary Table S5
Supplementary Table S6
Supplementary Table S7
Supplementary Table S8