
Abstract
Background:
Radioiodine refractory dedifferentiated thyroid cancer is a major clinical challenge. Anaplastic lymphoma kinase (ALK) mutations with increased ALK activity, especially fusion genes, have been suggested to promote thyroid carcinogenesis, leading to development of poorly differentiated thyroid carcinoma (PDTC) and anaplastic thyroid carcinoma. To determine the oncogenic potential of increased ALK activity in thyroid carcinogenesis in vivo, we studied mice with thyrocyte-specific expression of a constitutively active ALK mutant.
Methods:
Mice carrying a Cre-activated allele of a constitutively active ALK mutant (F1174L) were crossed with mice expressing tamoxifen-inducible Cre recombinase (CreERT2) under the control of the thyroglobulin (Tg) gene promoter to achieve thyrocyte-specific expression of the ALK mutant (ALKF1174L mice). Survival, thyroid hormone serum concentration, and tumor development were recorded. Thyroids and lungs were studied histologically. To maintain euthyroidism despite dedifferentiation of the thyroid, a cohort was substituted with levothyroxine (LT4) through drinking water.
Results:
ALKF1174L mice developed massively enlarged thyroids, which showed an early loss of normal follicular architecture 12 weeks after tamoxifen injection. A significant decrease in Tg and Nkx-2.1 expression as well as impaired thyroid hormone synthesis confirmed dedifferentiation. Histologically, the mice developed a carcinoma resembling human PDTC with a predominantly trabecular/solid growth pattern and an increased mitotic rate. The tumors showed extrathyroidal extension into the surrounding strap muscles and developed lung metastases. Median survival of ALKF1174L mice was significantly reduced to five months after tamoxifen injection. Reduced Tg expression and loss of follicular structure led to hypothyroidism with elevated thyrotropin (TSH). To test whether TSH stimulation played a role in thyroid carcinogenesis, we kept ALKF1174L mice euthyroid by LT4 substitution. These mice developed PDTC with identical histological features compared with hypothyroid mice, demonstrating that PDTC development was due to increased ALK activity and not dependent on TSH stimulation.
Conclusion:
Expression of a constitutively activated ALK mutant in thyroids of mice leads to development of metastasizing thyroid cancer resembling human PDTC. These results demonstrate in vivo that increased ALK activity is a driver mechanism in thyroid carcinogenesis.
Introduction
Thyroid cancer is the most common endocrine malignancy. Differentiated thyroid carcinoma (DTC) accounts for ∼90% of all thyroid cancers and shows an excellent prognosis. In contrast, poorly differentiated thyroid carcinoma (PDTC) and anaplastic thyroid carcinoma (ATC) are rare but have an unfavorable and often fatal prognosis (1 –4). PDTC accounts for a small proportion of all thyroid cancers with marked geographical differences: whereas PDTC is rarely seen in Japan (0.3%) and the United States (1.8%), it accounts for 4–6% of all thyroid cancers in Europe and Latin America (5 –7).
The tumors are usually widely invasive, extending to perithyroidal soft tissues in 60–70%. Regional lymph node involvement is found in 15–65%, and distant metastases, most commonly to the lung, are reported in 40–70% (5 –10). The overall 5-year survival of patients with PDTC is 60–70% and the 1-year survival of ATC is 10–20% (11). The poor prognosis is attributable to early mutations, the invasive and aggressive nature of the cancers, and, usually, refractoriness to radioiodine (3,12,13). Therefore, a main effort in thyroid cancer research is the identification of driver mutations in aggressive thyroid cancer amenable to targeted therapy (12,14,15).
A novel promising protein for targeted therapy could be the anaplastic lymphoma kinase (ALK). Oncogenic potential of ALK was first identified in neuroblastoma and nonsmall cell lung cancer (16 –18). A potential pathogenic role for ALK in development of thyroid cancer is based on the recent findings of ALK fusion genes in thyroid cancer without known driver mutations, for example, BRAF, RAS, or RET/PTC rearrangements.
Fusions of striatin (STRN), echinoderm microtubule-associated protein-like 4 (EML4), or Coiled-Coil Domain Containing 149 (CCDC149) genes with the kinase domain of the ALK gene were found in thyroid cancer samples (12,14,19 –21). In large-scale genetic studies, 4 ALK fusions were detected in 256 papillary thyroid carcinomas (PTCs), 3 in 35 PDTCs and 1 in 24 ATCs (14), 3 in 84 PDTCs and none in 33 ATCs (12), and 2 in 468 adult PTCs and none in 196 ATCs (21). In addition to ALK fusions, activating ALK point mutations has been reported in ATC (4,15). Fusion genes lead to increased expression and activity of the ALK. Consequently, expression of STRN-ALK transformed cells in vitro and induced tumor formation in subcutaneous rat PCCL3 cell xenografts (14).
The potential of ALK fusions to transform cells and the lack of overlap with other driver mutations in thyroid cancer samples suggested that increased ALK activity is an independent driver event in the development of thyroid cancer. To determine whether increased ALK activity is an independent driver event for thyroid cancer in vivo, we studied mice with thyrocyte-specific expression of the constitutively active ALK mutant F1174L (17,22). These mice develop an aggressive thyroid carcinoma with loss of follicular architecture and hormone synthesis, strap muscle invasion, and lung metastases, resembling human PDTC.
Materials and Methods
Animals and treatment
All animal experiments were approved by the Landesamt für Natur, Umwelt und Verbraucherschutz (LANUV) of North Rhine-Westphalia, Germany. A constitutively active ALK mutant (F1174L) was cloned into the Rosa26 locus. Expression of this ALK mutant is prevented by an upstream element containing a stop codon and a poly-A tail (hGH-pA; Supplementary Fig. S1A). This element is flanked by loxP sites (lox-stop-lox model). After excision of this stop element by Cre recombinase, expression of the ALKF1174L mutant is driven by the synthetic CAG promoter (17). In addition, the ALKF1174L transgene also encodes for luciferase downstream from an internal ribosomal entry site (IRES).
To achieve thyrocyte-specific expression of ALKF1174L, heterozygous ALKF1174L mice (ALKF1174L/+) were crossed with mice expressing Cre recombinase under control of a tamoxifen-inducible thyroglobulin promoter (TgCreERT2+/0) (23) (Supplementary Fig. S1A). TgCreERT2+/0;ALKF1174L/+ are referred to as ALKF1174L mice, and TgCreERT2+/0;ALK+/+ were used as controls, referred to as wild type (WT) mice. The background strain was C57BL/6. Mice were housed at 21°C ± 1°C and an alternating 12-hour light and 12-hour dark cycle. Standard chow (Sniff, Soest, Germany) and tap water were provided ad libitum, unless otherwise indicated. Tamoxifen was dissolved in corn oil (Sigma-Aldrich, St. Louis) at a concentration of 30 mg/mL.
At the age of 4–7 weeks and a body weight >15 g, ALKF1174L mice as well as WT littermates were injected intraperitoneally (i.p.) with 3 mg tamoxifen in 100 μL per mouse per day for 5 consecutive days (Supplementary Fig. S1A, B). Levothyroxine (LT4) substitution started 1 week after the first tamoxifen injection with tap water containing 133 ng/mL LT4 (renewed 3 times per week). LT4 stock solution was prepared at a concentration of 5 mg LT4 per 50 mL, 40 mM NaOH, and 0.2% bovine serum albumin (BSA) and stored in the dark at −20°C. The drinking water contained 130 ng/mL thyroxine (T4). Total T4 (TT4) serum concentrations in mice were measured every 30 days.
Genotyping and detection of recombination
Mice were genotyped using a standardized polymerase chain reaction (PCR) program and primers as follows: ALKF1174L-KI-rev 5′-CCCAAGGCACACAAAAAACC-3′, ALKF1174L-KI-fwd 5′-TGGCAGTCTAGGATCTG-3′, Cre-ERT2-rev 5′-TGGCAGCTCTCATGTCTCCAG-3′, Cre-ERT2-fwd 5′-TCAGAGATACCTGGCCTGGTC-3′. All primers were purchased from Eurofins MWG Operon (Ebersberg, Germany). Initial denaturation at 95°C for 5 minutes was followed by 40 cycles of 95°C for 30 seconds, 60°C for 15 seconds, and 72°C for 35 seconds. A final elongation step was carried out at 72°C for 3 minutes. Successful thyrocyte-specific Cre-mediated recombination was confirmed by PCR with the following primers: mALK-recom-rev: 5′- GCA CCA CGA AGT CAA CTG C -3′, ALKF1174L-fwd: 5′-TGG CAG TC TAG GAT CTG-3′, Rosa26-wt-rev: 5′-CAT GTC TTT AAT CTA CCT CGA TGG -3′; Rosa26-wt-fwd: 5′-CTC TTC CCT CGT GAT CTG CAA CTC C -3′(Supplementary Fig. S2).
In vivo bioluminescence imaging
ALK expression was also determined by detection of the coupled luciferase expression by in vivo imaging. Luciferase expression was coupled to ALKF1174L expression through an IRES. Mice were anesthetized with 1.75–2.5% isoflurane followed by an i.p. injection of 150 μg/g
Histology and immunostaining
Mice were euthanized with CO2 when predefined criteria were met, for example, loss of weight (≥20% body weight) or significant difficulty of breathing. A separate cohort of mice was euthanized at certain time points with CO2 to investigate cancer development. Blood was collected by cardiac puncture and serum was prepared as described previously (24). Directly after dissection, thyroid gland, brain, lung, and liver of each animal were macroscopically evaluated. Tissues were then fixed in 4% paraformaldehyde (phosphate buffered) and embedded in paraffin. Normal thyroid as well as tumor morphology was analyzed on 5-μm thick sections obtained from paraffin-embedded tissues stained with hematoxylin and eosin.
For Nkx-2.1 immunohistochemistry, endogenous peroxidases in thyroid sections were blocked with 3% hydrogen peroxide for 5 minutes. The primary antibody (1:50, Nkx-2.1, MA5-13961; Thermo Fisher Scientific, Inc., Waltham) was incubated for 30 minutes. The slides were incubated with horseradish peroxidase-labeled antimouse IgG for 30 minutes, 10 minutes with ZytoDAB, followed by hematoxylin (1:8; 5 minutes) staining. The samples were fixed with Immu-Mount (Thermo Fisher Scientific, Inc.). Tg and vimentin (Vim) expression was analyzed by immunofluorescence. Samples were blocked with 3% BSA/0.1% Triton/PBS for 30 minutes at room temperature (RT).
Samples were incubated with primary antibodies for Tg (1:1000, ab156008; Abcam) and Vim (1:100, D21H3; Cell Signaling, Danvers) dissolved in 1% BSA/0.1% Triton/PBS overnight at 4°C in a humidified chamber and protected from light. Appropriate Cy3-labeled secondary antibodies were dissolved in 1% BSA/PBS (1:200). Samples were then incubated with a secondary antibody for 1 hour at RT. Sections were counterstained with Hoechst 33342 (1:1000 in PBS; Thermo Fisher Scientific, Inc.) for 5 minutes at RT to visualize nuclei. Images were taken with an ELYRA PS.1 LSM710 confocal microscope (Zeiss, Germany). Pseudocolor imaging (green) was used to visualize Tg.
Mouse thyroid sections were reviewed by two experienced thyroid pathologists blinded to the genotype. For detection of Ki-67, tissue sections were incubated with a rabbit anti-Ki-67 antibody (1:400, RBK027; Zytomed Systems, Germany) and positivity was detected with the ZytoChem-Plus HRP Kit (ZytoChem-Plus HRP Kit, Broad Spectrum; HRP008DAB; Zytome Systems, Germany). A total of 500–1000 tumor cells were counted from 10 representative images of each stained tumor. Ki-67 index was calculated by dividing Ki67-positive by Ki67-negative epithelial cells.
Immunoblot analysis
Thyroid tissues (WT thyroids n = 2, ALKF1174L thyroids n = 2) were homogenized in radioimmunoprecipitation assay lysis buffer (150 mM NaCl, 50 mM Tris–HCl, 1% NP-40, 0.5% sodiumdesoxycholate, 0.1% sodium dodecyl sulfate (SDS), 2 mM ethylenediaminetetraacetic acid) with PhosSTOP and cOmplete Protease Inhibitor Cocktail (Sigma-Aldrich) for whole protein lysates. A total of 20 μg of protein were separated by SDS-polyacrylamide gel electrophoresis and blotted onto a polyvinyl difluoride membrane (Roti-Fluoro PVDF; Roth). Membranes were blocked with 5% BSA (Sigma-Aldrich) in Tris-buffered saline with 150 mM NaCl, 0.1% Tween 20, and pH 7.4 for 1 hour at RT.
The following primary antibodies were used: p44/42 MAPK (Erk1/2) (1:1000, #4695S; Cell Signaling), phospho-p44/42 MAPK (1:1000, #4370T; Cell Signaling), phospho-AKT (S473) (1:1000, #9271S; Cell Signaling), phospho-AKT (T308) (1:1000, #2965S; Cell Signaling), and AKT (1:1000, #9272; Cell Signaling). They were incubated overnight at 4°C. Antirabbit IgG HRP linked (1:2000; #7074S Cell Signaling) served as second antibody. The signals were detected with a VersaDocMP4000 (BioRad) and the band densities were determined with Image Lab software (BioRad).
Thyroid function tests
Serum thyrotropin (TSH) was measured with a sensitive heterologous disequilibrium double-antibody precipitation radioimmunoassay (25). TT4 was measured by ELISA (DRG Instruments, Marburg, Germany).
Statistical analyses
Data were analyzed with GraphPad Prism6. Data are presented as mean ± standard error of the mean unless otherwise specified. p < 0.05 was considered statistically significant.
Results
Thyroid-specific ALKF1174L expression in ALKF1174L/+ mice
Tamoxifen-inducible Tg-specific Cre mice (Tg-CreERt+/0) were crossed with heterozygous lox-stop-lox ALKF1174L/+ mice to generate heterozygous Tg-specific Cre mice either heterozygous for mutant ALKF1174L (Tg-CreERt+/0; ALKF1174L/+) or homozygous for WT ALK (Tg-CreERt+/0; ALK+/+) (Supplementary Fig. S1A). To induce expression of the mutant ALK allele, ALKF1174L mice were injected with tamoxifen (Supplementary Fig. S1B). WT littermates expressing Tg-specific Cre were also injected.
No transgene effect was observed without tamoxifen injections, and activation of Cre by tamoxifen in WT littermates did not affect thyroid morphology or function. In addition, we used in vivo imaging of tamoxifen-injected ALKF1174L and WT mice, as an IRES genetically couples luciferase expression to ALKF1174L expression. A bioluminescence signal was detected only in thyroids of tamoxifen-injected ALKF1174L mice but not in WT mice (Supplementary Fig. S1C). Successful and thyrocyte-specific recombination was confirmed by PCR (Supplementary Fig. S2).
Thyrocyte-specific ALKF1174L expression leads to development of thyroid tumors resembling human PDTC
ALKF1174L mice developed massive goiters, which could be monitored in vivo by bioluminescence imaging (Fig. 1A). Goiters reached a volume up to 20-fold larger than that of WT thyroids with an irregular structure (Fig. 1B). This massive thyroid growth is probably the first consequence of constitutive activation of ALK and, subsequently, the MAPK/ERK and PI3K/AKT signaling pathways. Compared with thyroids from WT mice, thyroids from ALKF1174L mice showed increased phosphorylation of ERK and AKT at T308, but not at S473 (Fig. 1C). Histologically, ALKF174L thyroids showed partial follicular destruction and an increased number of perifollicular fibroblasts 12 weeks after tamoxifen injection.
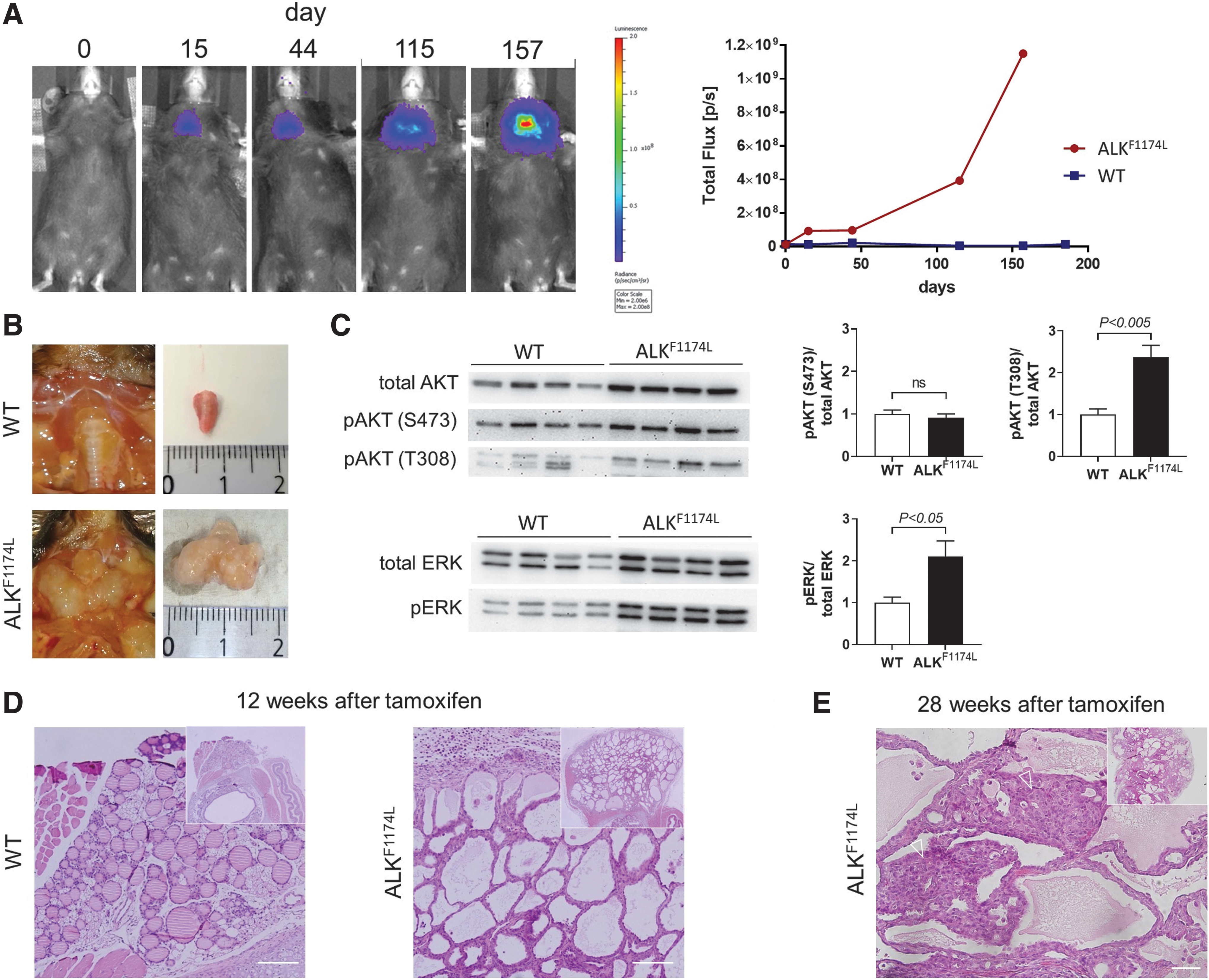
Features of ALKF1174L-positive thyroid glands. (
Furthermore, numerous follicles showed a considerable reduction or complete lack of colloid (Fig. 1D). Twenty-eight weeks after tamoxifen injection, the thyroids of ALKF1174L mice developed epithelial tumors with a solid/trabecular pattern (Fig. 1E). The tumor cell nuclei showed raisin-like folding of the nuclear membrane, resembling “convoluted nuclei” of human PDTC (8). Occasionally, mitotic figures were found, and the mitotic rate was >4/10 high power fields (HPF). Overall, the animal tumor morphology with a solid/trabecular growth pattern, convoluted tumor cell nuclei, and increased mitotic rate (>4/10 HPF) resembled human PDTC, meeting the so-called Turin consensus proposal criteria of PDTC (8), which have been included as an integral part of the PDTC definition in the recently published WHO classification of thyroid gland tumors (11).
Loss of thyroid-specific markers confirms dedifferentiation of thyrocytes
In contrast to thyroids from WT mice, which showed homogeneous Tg staining (Fig. 2A, left), ALKF1174L thyroids displayed partial loss of Tg staining (Fig. 2A, right). The tumors showed, similar to human PDTCs, a significantly reduced Tg expression. Similarly, decreasing expression of Nkx-2.1 demonstrated the gradual dedifferentiation of thyrocytes (26). Whereas WT thyroids were positive for Nkx-2.1 in almost all follicular cells, Nkx-2.1 expression in ALKF1174L thyroids was severely diminished 28 weeks after tamoxifen injection and even further reduced with increasing animal age (Fig. 2B). In ALKF1174L-positive mice, Nkx-2.1 positivity was only preserved in the few follicle-forming thyrocytes.

Loss of thyroid-specific differentiation markers and signaling pathway activation in ALKF1174L thyroids. (
Cancer aggressiveness is particularly enhanced by endothelial–mesenchymal transition (EMT) that allows a polarized epithelial cell to undergo phenotypical changes, enabling it to assume a mesenchymal cell phenotype, including an augmented migratory capacity (27,28). We, therefore, analyzed EMT by staining sections from ALKF1174L thyroids for Vim, a protein of the cytoskeleton of mesenchymal cells and a specific marker for EMT (27,28). Although WT thyroid sections were completely negative for Vim staining, the number of Vim-expressing cells was increased in ALKF1174L thyroids (Fig. 2C). This finding confirms EMT and supports the ALK-driven dedifferentiation of thyroid tissue.
Thyroid-specific ALKF1174L expression leads to development of an aggressive metastatic thyroid carcinoma
To evaluate tumor aggressiveness, we analyzed thyroid sections from ALKF1174L mice for signs of local invasion and screened peripheral organs (lung, liver, and brain) for metastases. We found invasive tumor infiltrating the surrounding cervical strap muscle tissue (Fig. 3A). The high mitotic rate of the PDTC-like tumors was confirmed by a Ki67 positivity of 26% (Fig. 3B). In addition to local invasive tumor growth, the tumors developed lung metastases (Fig. 3C).
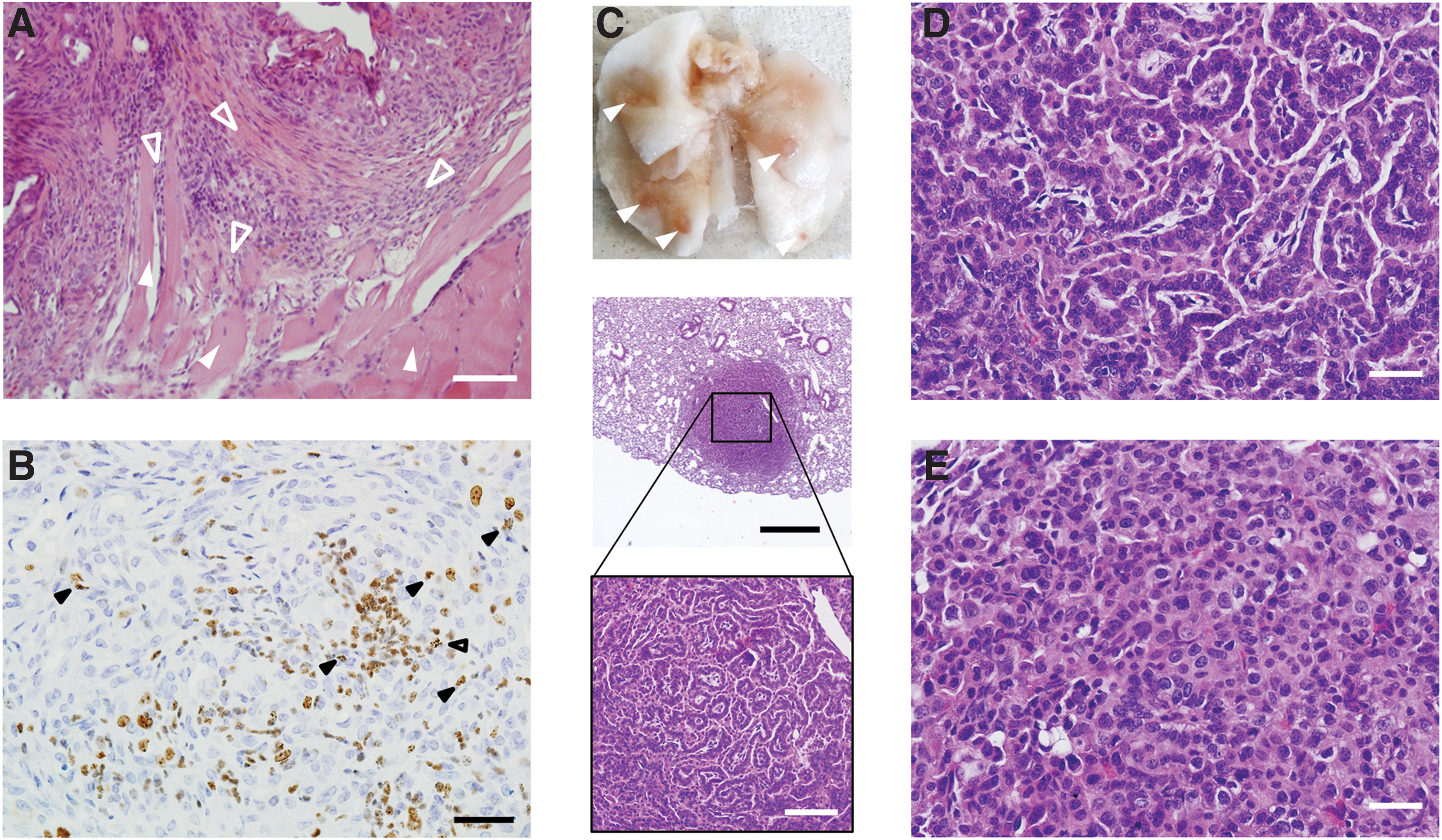
Invasive growth and pulmonary metastasis of PDTC-like thyroid tumors. (
Of note, lung metastases occurred as early as 24 weeks after tamoxifen treatment in ALKF1174L mice. The lung metastases showed morphological similarity to the primary tumor with a solid/trabecular growth pattern, convoluted tumor cell nuclei, and an increased mitotic rate (Fig. 3D, E). No metastases were found in brain and liver. The aggressive behavior of the ALKF1174L-dependent thyroid carcinomas was associated with a severely reduced survival of ALKF1174L mice. Whereas all WT mice survived the observation period of 60 weeks without complications, the median survival of ALKF1174L mice was severely reduced to only 20 weeks after tamoxifen injection.
Hypothyroidism is not a prerequisite for thyroid cancer development, but appears to promote progression
Considering the loss of follicular structure as well as the apparent dedifferentiation of ALKF1174L thyroids, we tested whether thyroid hormone synthesis was impaired. TT4 serum concentrations were significantly decreased in ALKF1174L mice compared with WT mice (Fig. 4). Consequently, the TSH serum concentration was severely increased in ALKF1174L mice (Fig. 4). In summary, destruction of the follicular structure and dedifferentiation led to overt hypothyroidism in ALKF1174L mice. TSH stimulates thyrocyte and thyroid cancer growth.
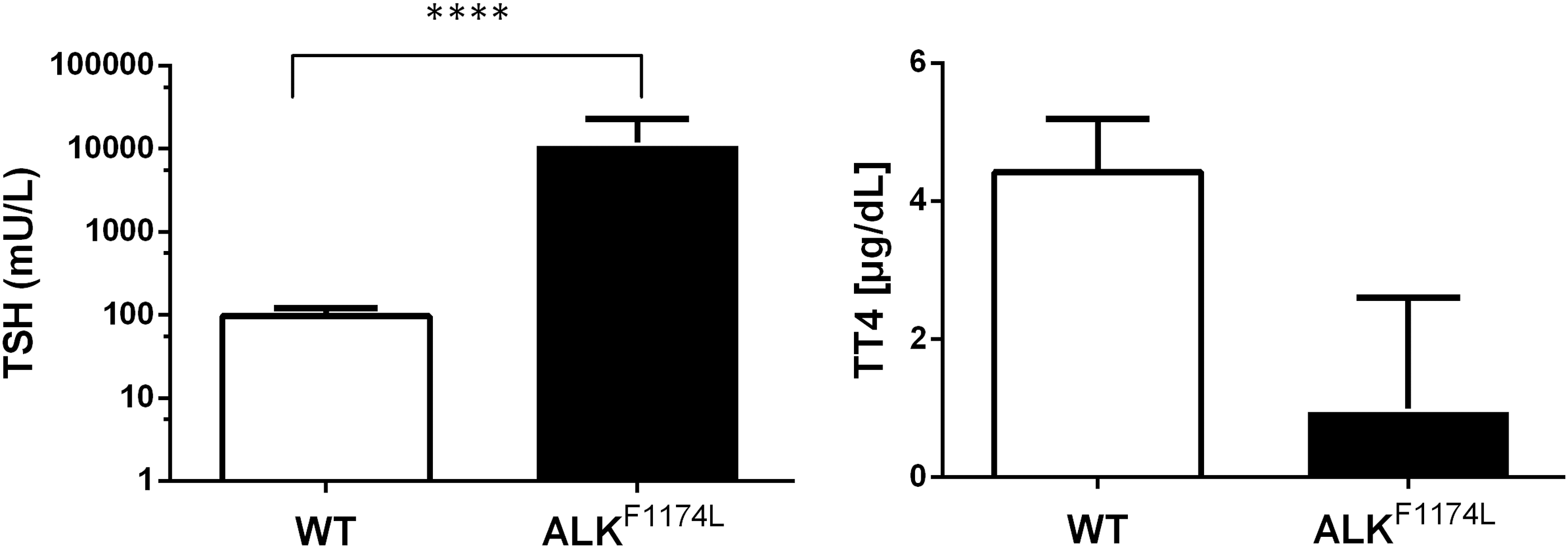
Thyroid-specific ALKF1174L expression leads to overt hypothyroidism. Serum concentration of TT4 was significantly decreased in ALKF1174L mice (black column) compared with WT control mice (open column). A significantly higher serum concentration of TSH in ALKF1174L mice confirmed hypothyroidism. (WT n = 5 and ALKF1174L n = 13; unpaired t-test). TSH, thyrotropin; T4, thyroxine; TT4, total T4.
To test whether stimulation by TSH is required for thyroid cancer development in ALKF1174L mice, we directly compared ALKF1174L mice with and without LT4 substitution through drinking water starting at the time point of the tamoxifen injections. LT4 substitution prevented development of hypothyroidism and the mice remained euthyroid with unchanged TT4 throughout the experiment (Supplementary Fig. S3). These animals developed thyroid cancers with identical histological features as mice without LT4 substitution. The median survival was longer in mice on LT4 substitution (32 vs. 18 weeks, p < 0.001; Fig. 5), whereas the final tumor weight did not differ between the 2 groups (375 ± 204 mg without LT4 vs. 449 ± 237 mg with LT4, not significant).
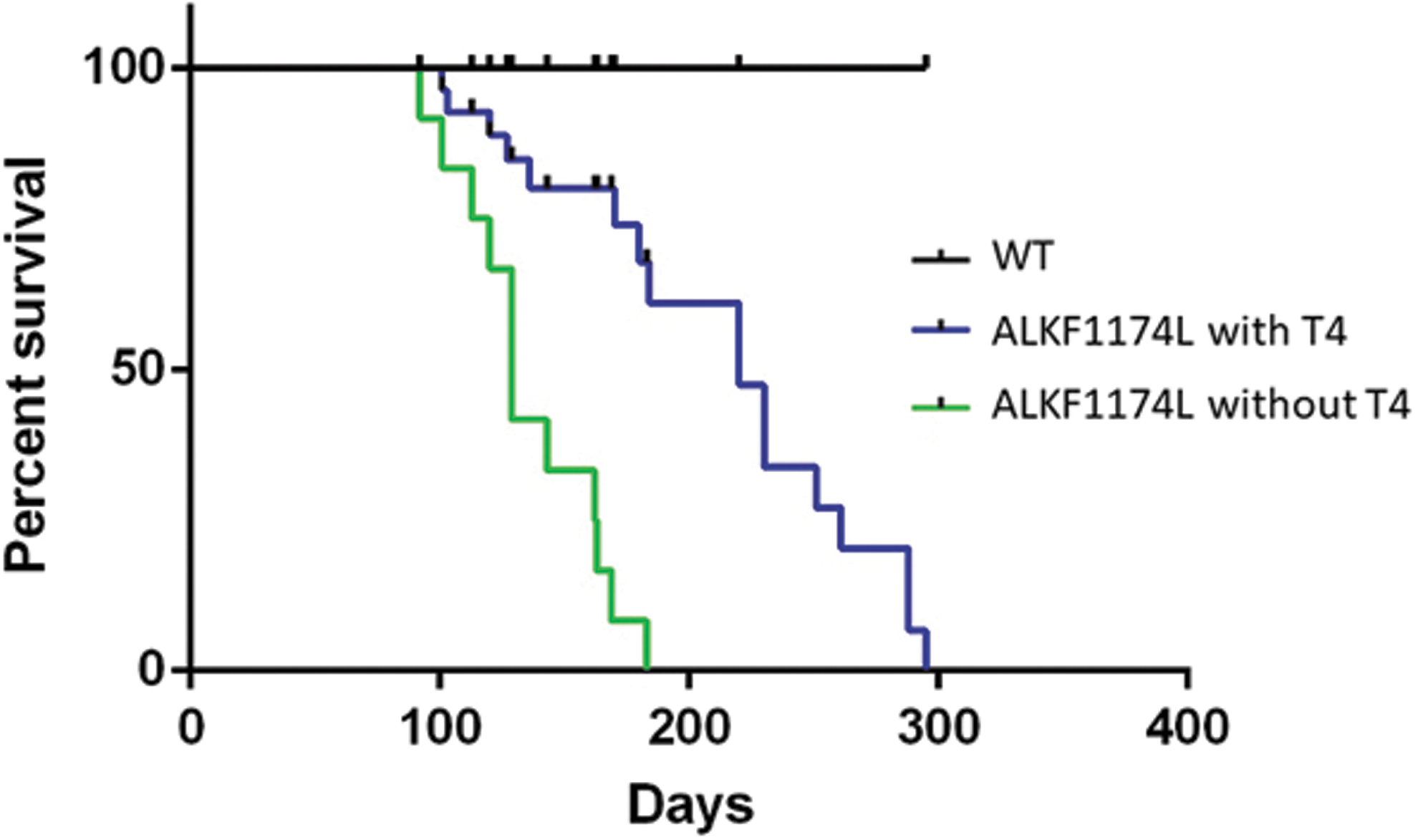
Median survival is reduced in ALKF1174L mice. Kaplan–Meier analysis of ALKF1174L without (n = 12) and with (n = 17) T4 substitution and WT (n = 13 with and without [n = 14] T4 substitution) mice shows a severely reduced median survival of ALKF1174L mice to 18 weeks after induction of ALKF1174L expression through tamoxifen, whereas all WT mice survived the observation period. Compared with untreated mice, substitution with T4 lead to a significantly longer survival (32 weeks, p < 0.001).
Thus, also in euthyroid mice, increased ALK activity is sufficient to induce thyroid cancer, whereas hypothyroidism with increased TSH stimulation appears to promote tumor progression.
Discussion
ALK mutations and gene rearrangements have been described in PTC, PDTC, and ATC (12,14,15). Because ALK fusions increase cell proliferation and transformation in vitro and in xenografts, increased ALK activity is considered a driver mechanism for thyroid cancer development. We studied whether increased ALK activity with subsequently increased PI3K and MAPK/ERK signaling in thyrocytes leads to development of thyroid cancers in vivo and, if so, which type of cancer. Increased thyrocyte-specific expression of a constitutively ALK mutant in ALKF1174L mice led to the development of thyroid cancers with features closely resembling human PDTC.
The pathogenic potential of ALK fusion genes depends on two main features, increased expression and ligand-independent ALK activation. Whereas WT ALK expression in thyrocytes is low, expression of the STRN-ALK and EML4-ALK fusion genes is significantly higher, on average 55-fold that of WT ALK (14,29), due to the active promoter of the upstream fusion partner. The upstream fusion partner also provides a dimerization domain that leads to ligand-independent activation of the ALK domain and increased signaling with increased MAPK activation.
In the mouse model used here, the mechanism of increased expression and activity of ALK differs from fusion genes. First, in STRN-ALK and EML4-ALK fusion genes, the active promoter that increases expression of the chimeric gene is provided by the upstream fusion partner, whereas in the ALKF1174L mouse an active synthetic promoter ensures increased ALK expression. Second, in STRN-ALK fusion proteins, ligand-independent ALK activation is mediated by a dimerization domain provided by the upstream fusion partner, whereas in the ALKF1174L mouse, an activating mutation leads to ligand-independent ALK activation. Notwithstanding these mechanistic differences between mouse model and patients, the ALKF1174L mouse demonstrates in vivo that increased ALK activity with subsequent MAPK and PI3K signaling pathway activation is a driving event in development of thyroid cancer, with a phenotype consistent with PDTC.
According to the WHO classification (11), human PDTC is defined as a follicular cell neoplasm that shows limited evidence of follicular cell differentiation and is morphologically and behaviorally intermediate between differentiated (follicular and papillary) and anaplastic carcinomas. The histopathological criteria for PDTC, as listed in the Turin consensus proposal (8), are a solid, trabecular, and/or insular growth pattern, convoluted tumor cell nuclei, tumor necrosis, and/or an increased mitotic rate (>3/10 HPF).
Furthermore, human PDTC shows decreased expression of Tg, diminished expression of Nkx-2.1, and a Ki67 index of 10–30% (11). The thyroid tumors of ALKF1174L mice showed a solid/trabecular growth pattern, convoluted nuclei, an increased mitotic rate, reduced Tg and Nkx-2.1 expression, a high Ki67 index and extrathyroidal extension into cervical strap muscles and lung metastases. Thus, the thyroid carcinomas found in ALKF1174L mice meet the criteria developed for the diagnosis of human PDTC.
Dedifferentiation with reduced Tg expression and loss of the regular follicular structure led to severe hypothyroidism and a significant increase in TSH in ALKF1174L mice. TSH is a stimulus of thyroid cancer growth. Such a stimulatory effect of TSH had been demonstrated in TRβPV/PV mice that develop PTC only when thyrotropin receptor signaling is intact (30). In contrast, hypothyroidism alone for more than a year leads to thyroid growth in WT mice, but not to development of metastatic thyroid cancer. These observations led to the conclusion that stimulation of growth by TSH is necessary but not sufficient for PTC development. Similarly, in thyroid-specific BRAFV600E mice, complete TSH receptor knockout appeared to attenuate PTC development but did not prevent it (31,32). Euthyroid BRAFV600E mice still developed PTC (33).
These mouse models suggest that the presence of an oncogenic mutation is a prerequisite for development of thyroid cancer, while additional stimulation by TSH may promote cancer progression. Our results are in agreement with this hypothesis. Hypothyroid as well as LT4-substituted ALKF1174L mice developed PDTC, demonstrating that PDTC development is due to expression of the constitutively active ALK mutant and not to prolonged TSH stimulation. Yet, the survival time was reduced in hypothyroid mice, indicating that TSH stimulation does promote thyroid cancer growth.
Increased ALK activity, mainly due to ALK fusion genes, had been suggested as a driving event in the development of dedifferentiated thyroid carcinoma. The development of PDTC in the mouse model with increased ALK activity presented here strongly supports this hypothesis with in vivo data.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by a young investigator grant to H.K. (Stiftung Universitätsmedizin Essen) and Mercator Research Center Ruhr (An-2015-0054) and Deutsche Forschungsgemeinschaft (Mo 1018/3-1) grants to L.C.M.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3