
Abstract
Background:
Basic fibroblast growth factor (bFGF) has been implicated in the pathogenesis of Graves' orbitopathy (GO). It stimulates several processes, including hyaluronan synthesis, involved in orbital tissue volume expansion and may act synergistically with platelet-derived growth factor (PDGF)-BB. PDGF-BB is known to stimulate adipogenesis in orbital fibroblasts, but the effect of bFGF on adipogenesis in orbital fibroblasts is so far unknown. This study was conducted to determine whether (i) bFGF induces adipogenesis in orbital fibroblasts, (ii) bFGF and PDGF-BB together exert an additive or synergistic effect on adipogenesis, and (iii) treatment directed at bFGF- and PDGF-BB signaling may potentially be of interest for the treatment of GO.
Methods:
Orbital fibroblasts from GO patients and controls were cultured in adipocyte differentiation medium with or without bFGF and/or PDGF-BB at different concentrations. Adipogenesis was determined by Oil Red O staining and messenger RNA expression of the late adipocyte differentiation markers cell death-inducing DFFA-like effector C (CIDEC) and adiponectin (ADIPOQ). To demonstrate involvement of FGF-receptor and PDGF-receptor signaling, experiments were also conducted in the presence of dasatinib (inhibitor of PDGF-receptor) or nintedanib (inhibitor of PDGF-receptor and FGF-receptor).
Results:
bFGF significantly stimulated adipogenesis by orbital fibroblasts, as shown by increased Oil Red O staining and CIDEC and ADIPOQ expression after 14 days of differentiation. Furthermore, an additive effect of bFGF/PDGF-BB co-stimulation on adipogenesis was observed at the lowest concentration (12.5 ng/mL) of the growth factors tested. Nintedanib completely inhibited bFGF-, PDGF-BB-, and bFGF/PDGF-BB-induced adipogenesis, while dasatinib only fully abrogated PDGF-BB-induced adipogenesis.
Conclusion:
bFGF induces adipogenesis in orbital fibroblasts and as such may contribute to GO. The additive effect of bFGF and PDGF-BB on adipogenesis, along with the observed inhibitory effects of dasatinib and nintedanib, point at independent receptor-mediated effects. This supports the hypothesis that multi-target directed therapy might be more efficient in the treatment of GO.
Introduction
Graves' orbitopathy (GO), a potentially serious extrathyroidal manifestation of Graves' disease, is characterized by volume expansion of the orbital soft tissues surrounding the eyes. Typical clinical signs include proptosis, upper eyelid retraction, edema and erythema of the conjunctiva and eyelids, diplopia, and blindness in severe cases. Based on the European Group on Graves' Orbitopathy classification criteria, the severity of GO can be categorized into mild, moderate-to-severe, and sight-threatening GO. In sight-threatening cases, if left untreated, blindness may occur through dysthyroid optic neuropathy and/or corneal damage (1,2). This is of great concern because a significant portion of GO patients are refractory to therapy. Although the monoclonal antibody teprotumumab directed at the insulin-like growth factor 1 receptor (IGF-1R) showed unprecedented clinical potential in the treatment of GO (3,4), it can be expected that a substantial portion of patients will remain unresponsive to the very limited noninvasive and effective treatment options available today (5 –8). A better understanding of the pathophysiology underlying GO could reveal potential new targets for therapy that could complement existing therapies and possibly enable a specific patient-tailored approach in the treatment of GO.
Activation of orbital fibroblasts by inflammatory mediators, growth factors, and stimulatory autoantibodies directed against the thyrotropin receptor (TSHR) and possibly the IGF-1R derived from activated and infiltrated immune cells is at the heart of the pathogenesis underlying GO (9,10). The activated orbital fibroblasts in turn produce inflammatory cytokines and chemokines that further enhance local inflammation. In addition, the orbital fibroblasts proliferate, produce extracellular matrix components (especially the glycosaminoglycan hyaluronan), and differentiate into adipocytes (11 –15). All these processes contribute to the typical orbital tissue volume expansion and subsequent clinical signs.
Studies into factors and signal transduction pathways that drive de novo adipogenesis in GO remain relatively limited (9). Several factors involved in GO, including TSHR stimulatory autoantibodies, interleukin (IL)-1β, IL-6, prostaglandin D2 (PGD2), and platelet-derived growth factor (PDGF)-BB, stimulate adipogenesis by orbital fibroblasts, while for instance interferon (IFN)-γ and tumor necrosis factor (TNF)-α are reported to inhibit adipogenesis (2,9,16 –19). Multiple factors that affect adipogenesis will be present simultaneously in GO orbital tissue. Therefore, in order to provide additional insight into the pathogenesis of GO, the contribution of specific factors, as well as their interaction with other mediators involved in adipogenesis, requires further study.
Basic fibroblast growth factor (bFGF/FGF2) exerts its cellular effects by binding and activating receptors belonging to the receptor tyrosine kinase (RTK) family. It induces angiogenic, chemotactic, and mitogenic activity in several cell types and contributes to developmental processes, as well as tissue regeneration and repair processes following injury. On the other hand, excessive bFGF activity promotes fibrosis via excessive fibroblast activation (20,21). In GO, elevated bFGF serum levels have been reported, and immunohistochemical studies have revealed increased bFGF expression in orbital tissue, which was mostly confined to fibroblasts and adipocytes (22 –24). Although these data suggest a role for bFGF in the pathogenesis of GO, the potential cellular effector mechanisms by which bFGF could contribute have hardly been defined to date. Recently, it was demonstrated that bFGF stimulates proliferation and hyaluronan production by orbital fibroblasts (25). In relation to adipogenesis, bFGF was found to increase the adipogenic potential of several cell types, including mesenchymal stem cells, adipose-derived stem cells, and skeletal muscle progenitor cells (26 –30). Yet, bFGF has not been examined in relation to adipogenesis in GO so far. This is, however, important, especially since the combination of bFGF and PDGF-BB (another growth factor implicated in GO) was found to exert a synergistic stimulatory effect on hyaluronan and IL-6 production by orbital fibroblasts, while PDGF-BB also stimulates adipogenesis in orbital fibroblasts (18,25).
Therefore, this study examined the effect of bFGF on adipogenic differentiation by orbital fibroblasts obtained from GO patients during the active and inactive disease stage and controls. In addition, the interaction between bFGF and PDGF-BB on adipogenesis by GO orbital fibroblasts was also examined.
Methods
Cell culture
Orbital tissue specimens were obtained from eight euthyroid GO patients during orbital decompression surgery. Four of the patients underwent surgery during the active disease stage (clinical activity score [CAS] was 5/7 in two of these patients and 7/7 in the other two patients) and four during the inactive disease stage (CAS all 0). Furthermore, orbital tissue was obtained from four control patients without inflammatory and thyroid disease who underwent surgery for cosmetic reasons. Orbital fibroblast cultures were established from orbital tissues, as described before (31). Once fibroblast monolayers were obtained, cultures were passaged serially after gentle treatment with trypsin/EDTA (Gibco, Paisley, United Kingdom) and cultured in Dulbecco's modified Eagle's medium (DMEM; Gibco) supplemented with 10% fetal calf serum (FCS; BioWhittaker, Verviers, Belgium) and antibiotics (penicillin and streptomycin; BioWhittaker) (32). Orbital fibroblasts used for experiments were between the 6th and 13th passage. All orbital tissues were obtained at the Rotterdam Eye Hospital (Rotterdam, the Netherlands) after obtaining informed consent and in accordance with the principles of the Declaration of Helsinki. The study was approved by the local ethics committee from the Erasmus University Medical Center (Medical Ethics Committee Erasmus MC) and the ethics committee from the Institutional Research Board of the Rotterdam Eye Hospital (research protocol: 2007-01).
Adipocyte differentiation
Orbital fibroblasts were seeded at 7.5 × 104 cells/well onto 24-well plates or at 4.0 × 105 cells/well onto six-well plates for Oil Red O (ORO) staining or RNA isolation, respectively. Cells were allowed to adhere overnight in DMEM 10% FCS in a humidified 5% CO2 incubator at 37°C. Adipocyte differentiation was induced, as described before (33). In brief, the culture medium was changed into adipocyte differentiation medium (DM) consisting of 1:1 serum-free DMEM/Ham's F12 (HyClone, Logan, UT) supplemented with antibiotics, 33 μM biotin (Sigma–Aldrich, St. Louis, MO), 17 μM pantothenic acid (Sigma–Aldrich), 1 μM insulin (Sigma–Aldrich), 10 μg/mL transferrin (Merck, Darmstadt, Germany), 0.2 nM triiodothyronine (T3; Sigma–Aldrich), 0.2 μM carbaprostaglandin (PGI2; Cayman Chemical Company, Ann Arbor, MI), and 10 μM rosiglitazone (Sigma–Aldrich) for 14 days. In the first three days, the DM was supplemented with 1 μM dexamethasone (Sigma–Aldrich) and 0.1 mM isobutylmethylxantine (IBMX; Sigma–Aldrich). The differentiation period was continued for 14 days during which the DM was refreshed every three to four days.
Adipocyte differentiation was compared between orbital fibroblasts cultured in DM in the absence or presence of recombinant human bFGF (12.5, 25, or 50 ng/mL; 233-FB; R&D Systems, Abingdon, United Kingdom), PDGF-BB (12.5, 25, or 50 ng/mL; 220-BB; R&D Systems), or a combination of bFGF (12.5 or 25 ng/mL) and PDGF-BB (12.5 or 25 ng/mL). Adipogenesis was assessed by ORO staining and mRNA expression analysis of the adipocyte associated molecules cell death-inducing DFFA-like effector c (CIDEC) and adiponectin (ADIPOQ).
ORO staining
ORO working solution was freshly prepared by mixing 6 mL 0.5% ORO (Sigma–Aldrich) stock solution in absolute isopropanol with 4 mL milli-Q and filtering through a 0.45 μm filter (Whatman, Dassel, Germany). Adipocyte differentiation cultures on 24-well plates were washed with 1 mL phosphate-buffered saline (37°C, pH 7.4) per well and stained by adding 500 μL ORO working solution for 10 min at room temperature. Subsequently, the culture plates were washed twice with 1 mL distilled water to remove excess ORO. Stained fibroblasts were visualized using an Axiovert 100 light microscope (Zeiss, Oberkochen, Germany), and photographed at 100 × and 250 × magnification using an AxioCam MR5 (Zeiss).
For quantification, the ORO staining was eluted from the cells with 500 μL absolute isopropanol on ice. The optical density (OD) was measured with the VersaMax ELISA microplate reader (Molecular Devises, Sunnyvale, CA) at 490 nm, as described before (18).
Real-time quantitative polymerase chain reaction analysis
After 14 days of adipocyte differentiation, mRNA was isolated using a GenElute Mammalian Total RNA Miniprep Kit (Sigma–Aldrich) and converted into cDNA, as described before (32). Transcript levels of CIDEC and ADIPOQ were determined by real-time quantitative polymerase chain reaction (QuantStudio 5; Thermo Fisher Scientific, Waltham, MA), normalized to the control gene Abelson (ABL) and expressed as fold induction relative to unstimulated cells. Primer–probe combinations used are listed in Supplementary Table S1.
Effect of FGF receptor and PDGF receptor inhibition on adipogenesis
To demonstrate involvement and to differentiate between FGF-receptor (FGFR) and PDGF-receptor (PDGFR) signaling in adipogenesis, orbital fibroblasts were stimulated with bFGF (12.5 ng/mL) or PDGF-BB (12.5 ng/mL) alone or in combination in adipocyte DM. This was performed in the absence or presence of 0.5 μM of the tyrosine kinase inhibitor (TKI) nintedanib (exhibits high inhibitory capacity for FGFRs and PDGFRs: BIBF 1120; S1010; molecular weight 539.62; Selleckchem, Houston, TX) or the TKI dasatinib (exhibits high inhibitory capacity for PDGFRs but not FGFRs: S1021; molecular weight 488.01; Selleckchem) (25,34).
Results
Effect of bFGF and PDGF-BB on lipid accumulation in orbital fibroblasts
A dose-dependent effect of bFGF-induced adipogenesis in orbital fibroblasts was demonstrated by a very clear accumulation of larger-sized fat droplets and an increase in the number of adipocytes (Fig. 1). In line with a previous observation, PDGF-BB (50 ng/mL) induced adipogenesis in orbital fibroblasts (18), which also appeared to be dose dependent, as this effect declined with decreasing concentrations of PDGF-BB (Fig. 1). Quantification of the ORO staining confirmed a statistically significant increase of lipid accumulation in orbital fibroblasts differentiated in the presence of all the concentrations of bFGF or PDGF-BB tested. The effect of both bFGF and PDGF-BB on adipogenesis was concentration dependent, although the increase in lipid accumulation observed between 25 and 50 ng/mL of bFGF, as well as between 25 and 50 ng/mL of PDGF-BB, did not reach statistical significance (Fig. 2). No difference in adipogenic response to bFGF or PDGF-BB was observed between orbital fibroblasts obtained from patients during active or inactive stage of disease (data not shown), or between orbital fibroblasts from GO patients and controls (Fig. 1 and Supplementary Figs. S1 and S2) Therefore, further experiments were only performed with GO orbital fibroblasts and growth factor concentrations of 12.5 and 25 ng/mL.
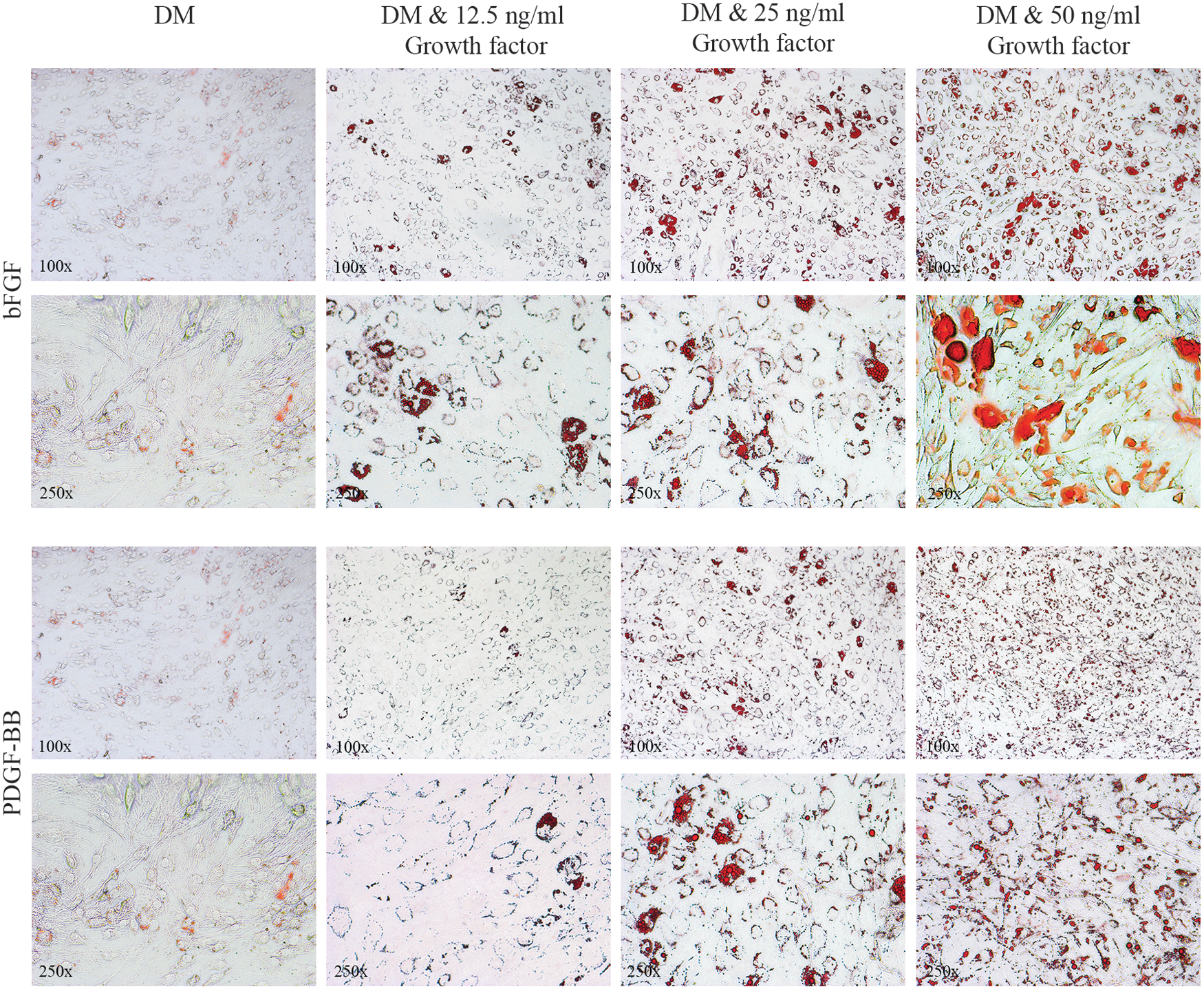
Graves' orbitopathy (GO) orbital fibroblasts cultured for 14 days in adipocyte differentiation medium (DM), DM +12.5 ng/mL basic fibroblast growth factor (bFGF) or platelet-derived growth factor (PDGF)-BB, DM +25 ng/mL bFGF or PDGF-BB, or DM +50 ng/mL bFGF or PDGF-BB. Fibroblasts were photographed at 100 × (first and third row) and 250 × (second and fourth row) magnification after staining with Oil Red O (ORO). Representative GO orbital fibroblast cultures are depicted.

ORO quantification of GO orbital fibroblasts (N = 8; n = 4 from active disease and n = 4 from inactive disease). After 14 days of differentiation in the absence or presence of bFGF
Effect of bFGF and PDGF-BB on CIDEC and ADIPOQ mRNA expression
bFGF stimulation for 14 days was associated with enhanced CIDEC and ADIPOQ mRNA expression. No clear dose-dependent effect of bFGF on mRNA expression levels of these genes was observed (Fig. 3). Stimulation with PDGF-BB also resulted in enhanced mRNA expression of CIDEC and ADIPOQ, which tended to be less than that observed with bFGF, although not at a statistically significant level (Fig. 3).
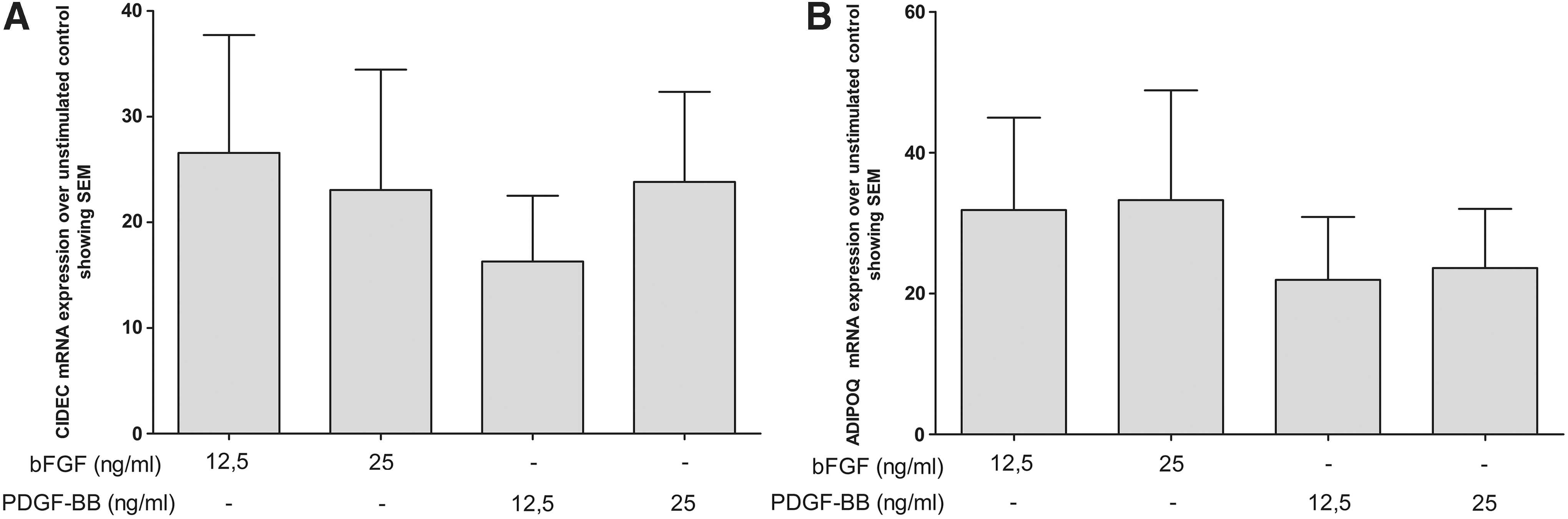
Expression of (
Co-stimulatory effect of bFGF and PDGF-BB on adipogenesis in GO orbital fibroblasts
The combination of bFGF (12.5 ng/mL) and PDGF-BB (12.5 ng/mL) stimulated adipogenesis more strongly compared to that induced by either of these factors alone at this concentration, and appeared to be additive in nature (Figs. 4 and 5). Stimulation with 25 ng/mL of bFGF or 25 ng/mL of PDGF-BB alone induced adipogenesis more strongly compared to stimulation with 12.5 ng/mL of these growth factors alone. However, no further enhancing effect on adipogenesis was observed upon co-stimulation with a combination of bFGF (25 ng/mL) and PDGF-BB (25 ng/mL) compared to stimulation with the individual growth factors at this concentration (Figs. 4 and 5).
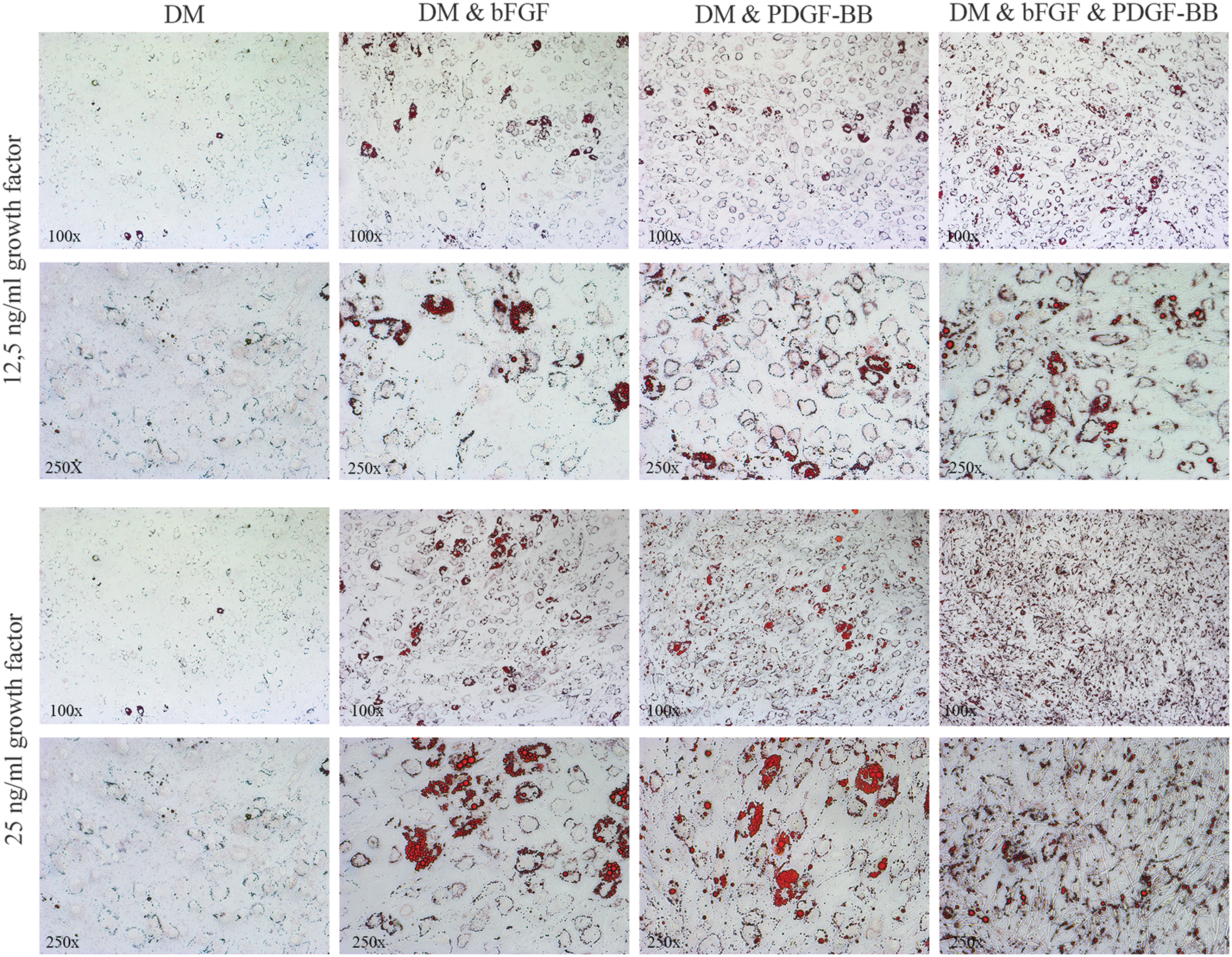
GO orbital fibroblasts cultured for 14 days in DM in the absence or presence of bFGF, PDGF-BB, or a combination of bFGF and PDGF-BB. The upper two rows depict stimulation with a concentration of 12.5 ng/mL for each growth factor. The lower two rows depict stimulation with a concentration of 25 ng/mL for each growth factor. Fibroblasts were photographed at 100 × (first and third row) and 250 × (second and fourth row) magnification after staining with ORO. Representative GO orbital fibroblast cultures are depicted.
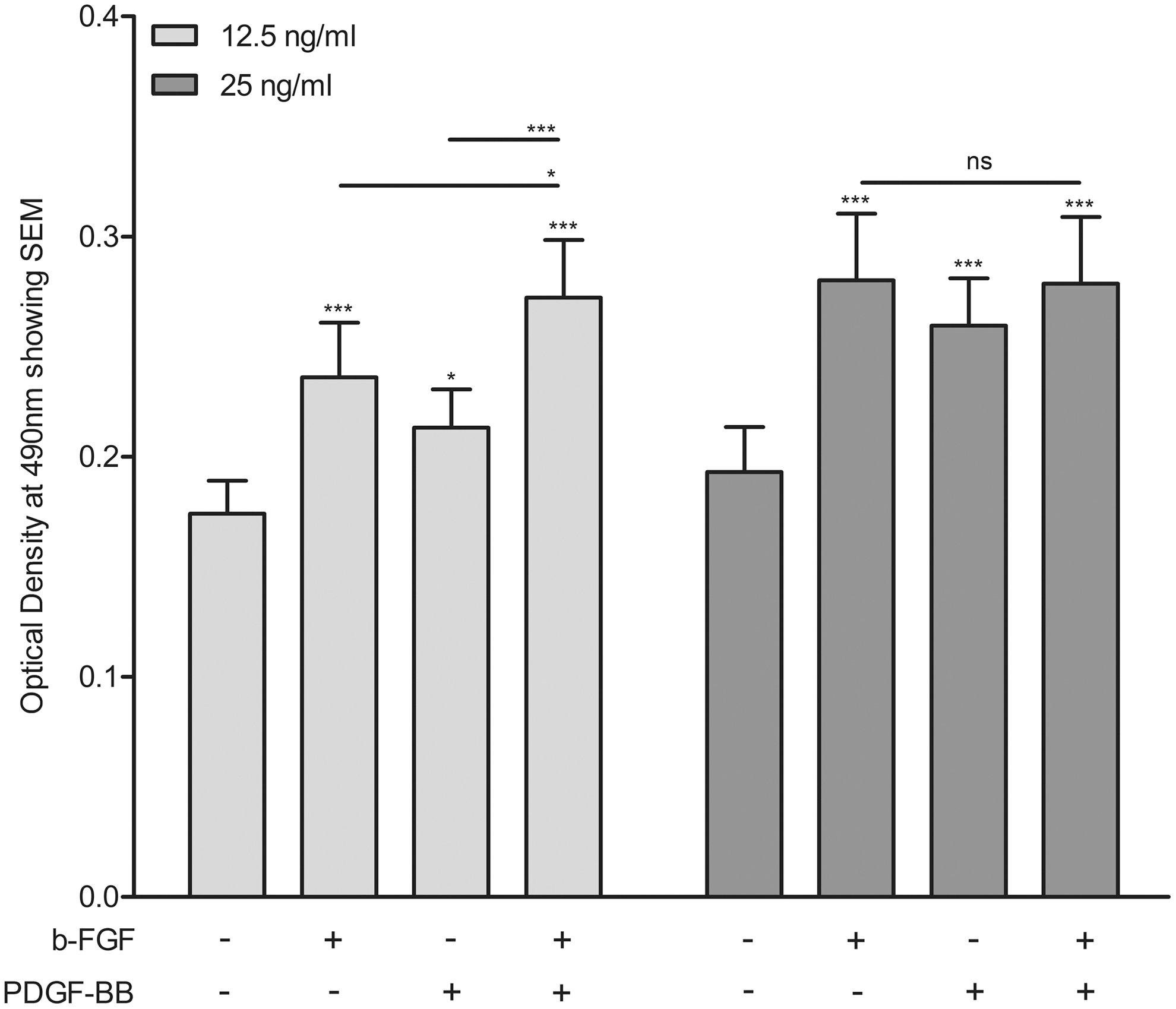
ORO quantification of GO orbital fibroblasts (N = 8; n = 4 from active disease and n = 4 from inactive disease). After 14 days of differentiation in the absence or presence of bFGF (12.5 or 25 ng/mL), PDGF-BB (12.5 or 25 ng/mL) or a combination of bFGF and PDGF-BB. Subsequently, the ORO was eluted, and the ORO-containing eluate was analyzed by spectrophotometry at 490 nm. Data are presented as the mean OD. Error bars indicate SEM. Statistical analysis was done using repeated-measures ANOVA followed by Tukey's multiple comparison test. *p < 0.05; ***p < 0.0001.
Effect of receptor inhibition on adipogenesis
bFGF-induced adipogenesis was significantly reduced by nintedanib (0.5 μM) up to the level of DM alone, while dasatinib (0.5 μM) slightly, but not significantly, inhibited bFGF-induced adipogenesis. PDGF-BB-induced adipogenesis was significantly inhibited up to the level of DM by nintedanib as well as dasatinib (Figs. 6 and 7), which is in line with previous findings of nintedanib and dasatinib inhibition of PDGF-BB induced proliferation and hyaluronan and IL-6 production (25). The combined effect of bFGF and PDGF-BB on adipogenesis was significantly reduced by nintedanib up to the level of DM alone. Dasatinib also significantly inhibited adipogenesis induced by co-stimulation with bFGF and PDGF-BB. Yet, this inhibition was not complete and only reached levels slightly below the adipogenesis observed with bFGF stimulation alone (Figs. 6 and 7).
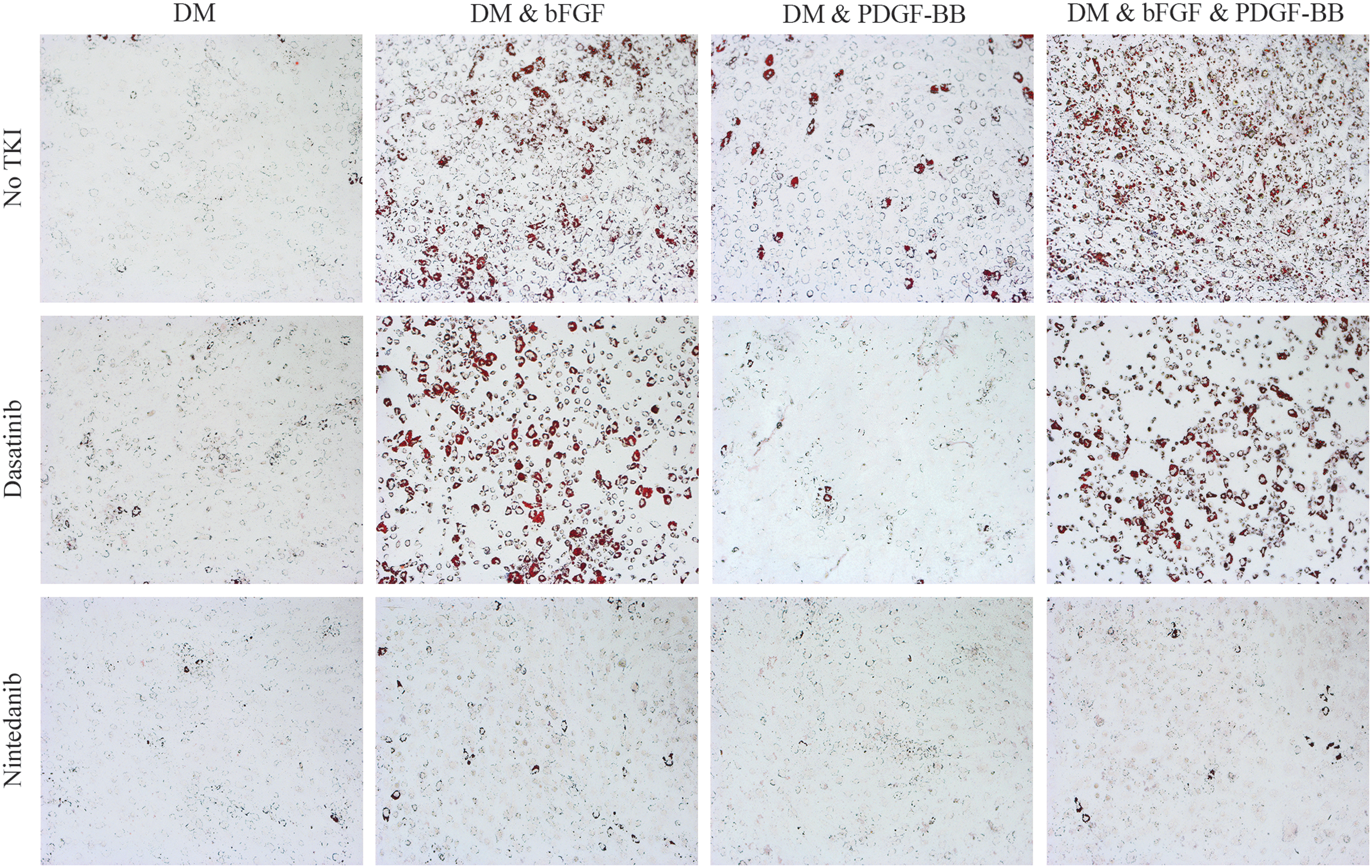
GO orbital fibroblasts cultured for 14 days in DM, in the absence or presence of bFGF (12.5 ng/mL), PDGF-BB (12.5 ng/mL), or a combination of bFGF and PDGF-BB (both at 12.5 ng/mL), either without or with 0.5 μM dasatinib or 0.5 μM nintedanib. Cells were photographed at 100 × magnification. Representative GO orbital fibroblast cultures are depicted.
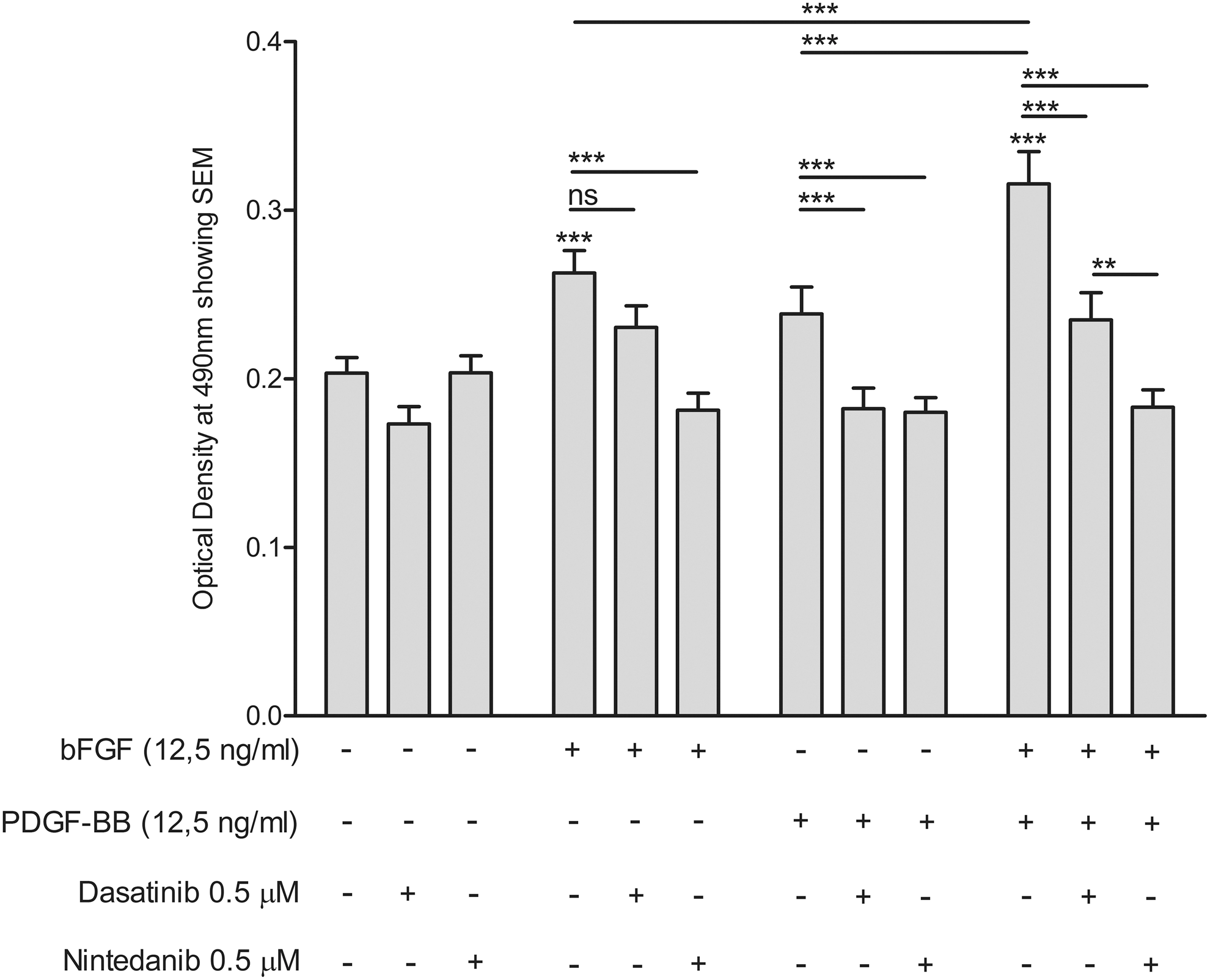
ORO quantification of GO orbital fibroblasts (N = 8; n = 4 from active disease and n = 4 from inactive disease). After 14 days of differentiation in the absence or presence of bFGF (12.5 ng/mL), PDGF-BB (12.5 ng/mL), or a combination of bFGF and PDGF-BB (both at 12.5 ng/ml), in either the absence or presence of dasatinib (0.5 μM) or nintedanib (0.5 μM). GO orbital fibroblasts were stained with ORO. Subsequently, the ORO was eluted, and the ORO-containing eluate was analyzed by spectrophotometry at 490 nm. Data are presented as the mean OD. Error bars indicate SEM. Statistical analysis was done using repeated-measures ANOVA followed by Tukey's multiple comparison test. **p < 0.01; ***p < 0.0001.
Discussion
Orbital tissue-volume expansion in GO is determined by several components, including excessive production of the glycosaminoglycan hyaluronan by orbital fibroblasts and the differentiation of orbital fibroblasts into adipocytes (adipogenesis). Soluble molecules present within the orbital tissue from GO patients are considered to stimulate these processes by orbital fibroblast activation (9). Elevated levels of bFGF have been found in orbital tissue and serum from GO patients (23,24). A pathological contribution of bFGF to GO has been attributed to the stimulation of hyaluronan production and proliferation by orbital fibroblasts (22,25). The current study demonstrates for the first time that bFGF also stimulates adipogenesis of orbital fibroblasts. In contrast to previous findings for the induction of hyaluronan and IL-6 production by orbital fibroblasts, no synergistic effect of bFGF and PDGF-BB on adipogenesis was observed. Rather, an additive effect on adipogenesis at the lowest growth factor concentrations tested was observed. The data further support the notion that bFGF might represent a growth factor that contributes to orbital tissue expansion in GO.
bFGF stimulated adipogenesis in a concentration-dependent manner. A positive correlation between serum bFGF levels and CAS has been described (22). Furthermore, bFGF can be detected in the orbital tissue of almost half of GO patients with a CAS ≥5, indicative of an active disease stage (24). Importantly, this study found a similar pro-adipogenic effect of bFGF in orbital fibroblasts isolated from patients with active disease, patients with inactive disease, and control patients. This suggests that the capacity of orbital fibroblasts to differentiate into adipocytes when encountering bFGF does not significantly alter with disease or disease progression, although bFGF levels may differ during the course of GO disease. On the other hand, this implies that bFGF might contribute to various pathological processes during the different disease stages, depending on local inflammatory milieu in the orbital tissue. The active disease stage of GO is associated with predominant infiltration of T-helper 1 (Th1)-cells and, accordingly, elevated levels of typical Th1 cytokines, including IFN-γ, which can exhibit an inhibitory effect on adipogenesis in orbital fibroblasts. One could speculate that the adipogenic potency of bFGF might therefore be limited during this disease stage (19,35). Progression of GO to the inactive disease stage is associated with increased orbital presence of Th2 T cells and associated cytokines (35). Therefore, bFGF might be involved in skewing the pathology toward adipocyte formation at a certain stage of progression from active to inactive disease. Further studies that determine the effect of bFGF, as well as other growth factors and cytokines, in a more disease stage–related environment are required to predict in vivo effects of these factors better, for instance in more complex in vitro orbital fibroblast culture systems or alternatively in whole orbital tissue cultures from GO patients (36) or a preclinical GO animal model (37).
The adipogenesis-enhancing effect of bFGF was similar to that induced by PDGF-BB, which was previously found to enhance adipogenesis in orbital fibroblasts (18). Adipogenesis is regulated by a complex and tightly controlled temporal gene transcriptional program. CIDEC and ADIPOQ expression is associated with late adipogenic differentiation and is involved in the formation and maintenance of mature adipocytes (38 –44). bFGF as well as PDGF-BB enhanced CIDEC and ADIPOQ mRNA expression in GO orbital fibroblasts, thus confirming adipocyte differentiation, as observed by ORO staining. Induction of CIDEC and ADIPOQ appeared slightly lower upon PDGF-BB stimulation than upon bFGF stimulation. These data suggest that both bFGF and PDGF-BB induce adipogenesis in GO orbital fibroblasts and that differences in kinetics may exist.
An additive effect of bFGF/PDGF-BB co-stimulation on adipogenesis was only observed for the lowest concentration (12.5 ng/mL) of growth factor tested. This might suggest that the higher concentrations of bFGF and PDGF-BB used in this study already skew to maximal lipid accumulation individually. Alternatively, the assays were performed in the presence of the peroxisome proliferator-activated receptor gamma agonist rosiglitazone and thus a rather strong pro-adipogenic environment (33,45,46). The possibility cannot be excluded that this culture condition resulted in an underestimation of the pro-adipogenic effect of bFGF, PDGF-BB, or a combination thereof on adipogenesis.
The combined effect of bFGF and PDGF-BB on adipogenesis was fully abrogated by nintedanib, a TKI with high inhibitory potency for FGFRs and PDGFRs. In contrast, the TKI dasatinib that exhibits high inhibitory capacity for the PDGFRs but not the FGFRs only inhibited adipogenesis slightly below the level of that induced by bFGF alone. These findings corroborate the additive effects that bFGF and PDGF-BB exert on adipogenesis in GO orbital fibroblasts and are indicative for bFGF and PDGF-BB driving independent routes of adipogenesis. Although not statistically significant, dasatinib slightly reduced adipogenesis under baseline culture conditions (in the absence of growth factors), as well as bFGF-induced adipogenesis (Fig. 7). This might be caused by off-target effects of dasatinib on the Src-family tyrosine kinases, which are involved in adipogenesis (47,48). On the other hand, it cannot be excluded that bFGF stimulation is associated with PDGFR transactivation (49) or that bFGF stimulation is associated with induction of autocrine PDGF signaling that is inhibited by dasatinib.
Importantly, the present data also illustrate that although a combination of growth factors might synergistically enhance certain aspects of GO pathogenesis (e.g., hyaluronan and cytokine production, as described for bFGF + PDGF-BB) (25), this is not necessarily the case for every pathogenic mechanism involved in GO. Therefore, studies exploring the effect that combinations of mediators involved in GO pathogenesis exert on orbital fibroblast activity are of paramount importance to increase our understanding of this complex disease. However, so far, the number of such studies remains limited (9).
This study along with previous observations (9,25) indicate that FGF and PDGF signaling pathways are potentially interesting treatment targets in GO. Although not yet clinically available, several possibilities can be thought of for targeting these signaling pathways, including bFGF and PDGF-B neutralizing antibodies, recombinant soluble receptors, aptamers, and dominant negative ligands targeting their respective receptors (50 –55). Alternatively, one might consider using a TKI such as nintedanib to interfere with the RTK, although it should be noted that considerable adverse effects are to be expected with these classes of therapeutics (56,57). These options indicate potential future therapeutic possibilities capable of interfering not only with bFGF and PDGF-BB signaling but also several other factors that will be revealed as the pathogenesis of GO is further unraveled. An individualized therapy treating patient-specific dominant pathological factors might be the best overall approach in treating GO.
In conclusion, this study shows that bFGF is a strong pro-adipogenic factor for orbital fibroblasts in vitro and as such may be involved in the pathogenesis of GO. As indicated by this as well as previous studies, interactions between different growth factors, cytokines, autoantibodies, and receptors may play an important role in GO pathogenesis (25,58 –61). Understanding such potential interactions not only enhances our pathological knowledge on this disease, but could also lead to improved treatment approaches, for instance by a multiple-target approach.
Footnotes
Acknowledgment
This research was partly funded (SV, WAD) by the Ratchadapisek Sompoch Endowment Fund (2017), Chulalongkorn University (760001-HR).
Author Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Table S1