
Abstract
Background:
Thyroid hormone (TH) synthesis is essential for the control of development, growth, and metabolism in vertebrates and depends on a sufficient dietary iodine intake. Importantly, both iodine deficiency and iodine excess (IE) impair TH synthesis, causing serious health problems especially during fetal/neonatal development. While it is known that IE disrupts thyroid function by inhibiting thyroid gene expression, its effects on thyroid development are less clear. Accordingly, this study sought to investigate the effects of IE during the embryonic development/differentiation of endoderm and the thyroid gland.
Methods:
We used the murine embryonic stem (ES) cell model of in vitro directed differentiation to assess the impact of IE on the generation of endoderm and thyroid cells. Additionally, we subjected endoderm and thyroid explants obtained during early gestation to IE and evaluated gene and protein expression of endodermal markers in both models.
Results:
ES cells were successfully differentiated into endoderm cells and, subsequently, into thyrocytes expressing the specific thyroid markers Tshr, Slc5a5, Tpo, and Tg. IE exposure decreased the messenger RNA (mRNA) levels of the main endoderm markers Afp, Crcx4, Foxa1, Foxa2, and Sox17 in both ES cell-derived endoderm cells and embryonic explants. Interestingly, IE also decreased the expression of the main thyroid markers in ES cell-derived thyrocytes and thyroid explants. Finally, we demonstrate that DNA methyltransferase expression was increased by exposure to IE, and this was accompanied by hypermethylation and hypoacetylation of histone H3, pointing to an association between the gene repression triggered by IE and the observed epigenetic changes.
Conclusions:
These data establish that IE treatment is deleterious for embryonic endoderm and thyroid gene expression.
Introduction
The thyroid hormones (TH) thyroxine and triiodothyronine are iodine-containing hormones in vertebrates and are the main hormones produced by the thyroid gland. TH synthesis is essential for the control of growth, development, and metabolism in vertebrates and depends on sufficient iodide intake (1).
Indeed, uptake of iodide is the limiting step in TH synthesis and is dependent on the activity of the sodium–iodide symporter (NIS) (2), located at the basolateral membrane of thyrocytes (3). Subsequently, iodide is transported through the apical membrane to the follicle colloid by a group of transporters including pendrin and anoctamin-1 (4,5). In thyroid follicular cells, thyroid peroxidase (TPO) uses H2O2 provided by dual oxidase 2 (DUOX2), to oxidize specific tyrosine amino acids within thyroglobulin (TG), and couples the iodotyrosines to generate the iodothyronines (6). The binding of thyrotropin (TSH) to its receptor (TSHR) expressed on the basolateral membrane of thyrocytes stimulates all the biochemical processes involved in TH synthesis and secretion (7).
An adequate level of iodine is essential for human biology. However, iodine is scarce in the environment, and iodine deficiency remains a major public health concern in many populations worldwide, causing several thyroid disorders (8,9) such as goiter and hypothyroidism at all ages and, in extreme cases, causing cretinism and mental retardation in newborns of mothers who are iodine deficient (10).
By contrast, some populations are exposed to high amounts of iodine (11). Iodine excess (IE) is also associated with hypothyroidism in susceptible patients, in addition to patients on antithyroid drug therapy and the concomitant use of potential goitrogens, such as lithium. Furthermore, IE is associated with hyperthyroidism in susceptible patients with risk factors including nontoxic or diffuse nodular goiter, latent Graves' disease, and long-standing iodine deficiency (12). Importantly, IE has been shown to disrupt thyroid function, as the trace element iodide is an extremely reactive anion (13).
According to the World Health Organization (WHO), adults should consume 150 μg of iodine per day to guarantee an adequate production of TH. However, in some countries, including Brazil and China, the concentration of urinary iodine in individuals can be higher than 300 μg/L (14,15).
The requirement of iodine consumption increases considerably during pregnancy and lactation, due to increased maternal TH production, increased renal clearance of iodine, and iodine transfer to the fetus (16). Accordingly, the WHO suggests a daily consumption of 200–250 μg iodine during these critical developmental periods to guarantee normal maternal and fetal TH synthesis (17). Therefore, maternal iodine intake should be carefully monitored, especially in iodine-deficient areas (18).
Although the deleterious effects of iodine deficiency to maternal and fetal thyroid function have been extensively reported (19), recent studies show that maternal consumption of high concentrations of iodine also impairs thyroid function in mothers and offspring (20 –23). In this context, we have recently demonstrated that maternal thyroid function is impaired by IE exposure during pregnancy and lactation (24). Additionally, this scenario also induces primary hypothyroidism in male offspring during adult life (25).
The fetal thyroid gland initially develops from the endoderm (26,27) at gestational day 24 in humans and at embryonic day 8.5 (E8.5) in mice/rats. Thyroid transcription factor 1 (Nkx2.1), thyroid transcription factor 2 (Foxe1), and paired box gene 8 (Pax8) are the major thyroid transcription factors involved in thyroid development and are expressed before or just after the first appearance of the thyroid diverticulum (28,29). Indeed, Nkx2.1 and Pax8 act coordinately in early thyroid gland embryogenesis, whereas Foxe1 is essential for thyroid migration. Beyond their role in thyroid development, the coordinated expression of thyroid transcription factors also maintains the differentiation of the thyroid gland through the regulation of the expression of genes that encode for proteins involved in TH synthesis (30).
We previously showed that IE exposure impairs the maternal and offspring thyroid function, although the effects of this treatment on fetal thyroid development are not known. Therefore, this study aimed to investigate the impact of IE treatment on thyroid-related gene expression in the embryonic development and differentiation of endoderm and the thyroid. Moreover, the participation of epigenetic mechanisms on these developmental processes was also evaluated.
Materials and Methods
Animals
Wild-type CD1 mice were mated overnight, and pregnancy was confirmed by the presence of a vaginal plug, which was considered as E0.5. Embryos from timed pregnancies were collected, and embryonic endoderm and thyroid explants were isolated and cultured as described below.
Dissection and culture of embryonic ventral foregut endoderm explants were performed as follows: E8.5 embryos were removed from the uteri of the pregnant mice, transferred to dishes containing phosphate-buffered saline (PBS), and dissected free of decidua tissue, as previously described (31).
Briefly, under a dissecting microscope (magnification 60 × ), the yolk sac, optic lobes, and hindgut were carefully removed using electrolytically etched tungsten needles. To isolate the presumptive ventral endoderm, head, upper cardiac lobe, dorsal, and posterior tissues were dissected away from the ventral foregut. Extracted tissues were placed in the center of rat-tail collagen I-coated chambers (μ-slide 8 well; Ibidi GmbH) and incubated at 37°C, 5% CO2, for 2 hours to allow adherence to the matrix. Thereafter, the chambers were filled with pre-warmed RPMI medium supplemented with 10% fetal bovine serum, and the tissue explants were treated or not with NaI (10−6 M) for 24 hours to simulate IE.
Dissection and culture of embryonic thyroid explants were performed as follows: E14.5 embryos were removed from the uteri of pregnant mice and dissected in dishes containing PBS, as previously described (32). Briefly, the embryos were separated from the placenta, and the yolk sac and the amnion were removed. Using a stereomicroscope, the upper part of the head, the upper jaw, the ears, the neural tube, and the lower part of the embryos were removed.
The embryo portion containing the tongue and neck region was transferred to a new 10-cm plate, and the tissues posterior to the esophagus and trachea were excised. The trachea was then placed with its ventral side facing up and the thymus was removed. Then, the trachea containing the thyroid lobes was placed in collagen I-coated plates and cultured in Ham's F-12 Coon's medium supplemented with a six-hormone mixture (1 nM TSH, 10 μg/mL insulin, 10 ng/mL somatostatin, 5 μg/mL transferrin, 10 nm hydrocortisone, and 10 ng/mL glycyl-
All animal experimentation was performed in compliance with the European Community Law (86/609/EEC) and the Spanish Law R.D. (1201/2005) with the approval of the Ethics Committee of the Consejo Superior de Investigaciones Científicas, Spain.
Embryonic stem cell differentiation
Mouse R1 embryonic stem (ES) cells were cultured on mitomycin-inactivated mouse embryonic fibroblasts, as described previously (33). Cells were maintained in complete Glasgow's modification of Eagle's medium supplemented with 10% ES cell-certified fetal bovine serum, 0.1 mM nonessential amino acids, 1 mM sodium pyruvate, 0.1 mM 2-mercaptoethanol, 50 U/mL penicillin, 50 mg/mL streptomycin, and 1.000 U/mL leukemia inhibitory factor (Millipore, Billerica, MA). Embryoid bodies (EBs) were generated by culturing cells in hanging drops (1000 cells per drop) for up to 60 hours. Thereafter, EBs were collected and cultured for a further 60 hours in fresh medium supplemented with 50 ng/mL activin A (R&D Systems, Minneapolis, MN). Anterior foregut endoderm differentiation was achieved by next treating the EBs for 24 hours with 100 ng/mL noggin, a BMP antagonist, and 10 mM SB431542 (both from Sigma–Aldrich Chemical Co.), a selective inhibitor of endogenous activin A but with no apparent effect on BMP signaling (34,35).
In the studies performed to analyze the effect of IE on endoderm differentiation in vitro, the EBs successively treated with activin, noggin, and SB431542 and exposed or not to IE during the differentiation protocol were collected and total RNA/protein was extracted as described below.
To induce the differentiation of endoderm cells into thyrocytes, EBs were trypsinized and plated in new culture dishes. Twenty-four hours later, the cells were transiently transfected with Nkx2.1- and Pax8-expressing vectors (36,37) using Lipofectamine 3000 (Invitrogen–Life Technologies, Carlsbad, CA). After 48 hours of transfection, the cells were cultured with Ham's F-12 Coon's medium supplemented with 5% fetal calf serum and a six-hormone mixture (described above) for an additional 3 days in the presence or absence of IE (NaI 10−6 M).
The formation of spherical follicle-like structures (38) of endoderm cells transfected with Nkx2.1 and Pax8 was evaluated on Matrigel-embedded three-dimensional (3D) cultures. For this purpose, 7 × 103 cells were seeded into eight-well multiwell plates coated with 10% Matrigel (BD Biosciences, Franklin Lakes, NJ) and cultured with Ham's F-12 supplemented medium for 5 days. Follicle-like structure organization was then examined under a light microscope.
Flow cytometry analysis
Flow cytometry was used to evaluate the efficiency of endoderm and thyroid differentiation in vitro. Briefly, mouse embryonic stem cells-derived endoderm cells or transiently transfected endoderm cells were fixed in 2% paraformaldehyde for 20 minutes. Under permeabilized conditions, ES cell-derived endoderm cells were incubated with anti-SOX17 and anti-FOXA2 antibodies, while endoderm cells transiently transfected with Pax8- and Nkx2.1-expressing vectors were incubated with anti-PAX8 and anti-NKX2.1 antibodies. Thereafter, the cells were incubated with FITC-conjugated anti-rabbit, Alexa Fluor 568-conjugated anti-goat, and/or TRITC-conjugated anti-mouse antibodies (Supplementary Table S1). The fluorescence intensities of 104 cells were assayed on an Attune NxT Flow Cytometer (Thermo Fisher Scientific, Waltham, MA). Data were analyzed using Flowing Software 2.4.1 (Turku Center for Biotechnology, University of Turku, Finland).
Evaluation of endoderm cell apoptosis after IE treatment
Endoderm cells that were exposed or not to NaI (10−6 M) were analyzed by flow cytometry to assess apoptosis.
After treatment with noggin/SB431542, EBs were trypsinized and the cells were plated in new culture dishes. Twenty-four hours later, the supernatant was collected, and the adherent cells were trypsinized. Cells were centrifuged for 10 minutes at 1000 × g, washed with PBS, and then resuspended in 0.5 mL of annexin V binding buffer (10 mM HEPES, 140 mM NaCl, 2.5 mM CaCl2), containing 1 μg/mL Annexin V-APC (BD Biosciences) and 10 μg/mL propidium iodide (PI) (Sigma–Aldrich Chemical Co.). The cells were incubated for 15 minutes at room temperature in the dark. Subsequently, cells were analyzed by flow cytometry, using an Attune NxT Flow Cytometer with Attune NxT software, and at least 20,000 events were collected per sample. Data were analyzed using FlowJo software v 6.0 (FlowJo, Ashland, OR).
Gene expression analysis
Total RNA was extracted from endoderm and thyroid cells derived from EBs with Trizol (Invitrogen) and from embryonic endoderm and thyroid explants with the ReliaPrep RNA Cell Miniprep System (Promega, Madison, WI). Gene expression analysis of endoderm markers (Afp, Foxa1, Foxa2, Cxcr4, Gata4, and Sox17), thyroid markers (Slc5a5, Tshr, Tg, Tpo, Pax8, and Nkx2.1), and epigenetic markers (DNA methyltransferase [Dnmt]1 and Dnmt3) was evaluated by real-time quantitative polymerase chain reaction (RT-qPCR), as described previously (39), using the SYBR Green Master Kit (Stratagene, La Jolla, CA) on the Stratagene Mx3000P platform. Relative changes in gene expression were calculated using the 2−ΔΔCt method using Gapdh messenger RNA (mRNA) expression as an internal loading control. Melting curves were analyzed to confirm the amplification of a single PCR product.
The primers used in this study are listed in Supplementary Table S2.
Protein expression analysis
Total proteins were extracted in RIPA buffer (50 mM Tris [pH 8], 150 mM NaCl, 1% Nonidet P-40, 0.5% sodium deoxycholate, and 0.1% sodium dodecyl sulfate [SDS]) supplemented with a protease inhibitor cocktail (Roche, Basel, Switzerland). Proteins were resolved by 8–12% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE), transferred to nitrocellulose membranes, blocked, and subsequently incubated with specific primary and species-specific secondary antibodies diluted in Tris-buffered saline with 3% bovine serum albumin and 0.1% Tween 20, overnight at 4°C. Antibody reactivity was detected by chemiluminescence, and blot densitometry was analyzed using ImageJ Software (NIH). Antibodies used are listed in Supplementary Table S1.
DNA extraction and global DNA methylation analysis
DNA was extracted from tissues using the PureLink® Genomic DNA Kit (Invitrogen). Thereafter, global DNA methylation status was assessed with the Imprint® Methylated DNA Quantification Kit (Sigma–Aldrich Chemical Co.).
Histone modification
Post-translational modifications to histones were analyzed by Western blotting using specific primary antibodies to detect methylation (trimethyl-histone H3 [Lys27], H3K27me3) or acetylation (acetyl-histone H3, H3Ac) on histone H3. Antibodies used are listed in Supplementary Table S1.
Statistical analysis
All data are reported as mean ± standard error of the mean. Comparisons between two groups were made using two-tailed Student's unpaired t-test. Statistical analysis was performed with GraphPad Prism software (GraphPad, San Diego, CA). Differences were considered statistically significant at a p-value <0.05.
Results
IE impairs in vitro differentiation of endoderm cells
Previous studies have shown that combinatorial treatment of pluripotent stem cells with activin A, noggin, and SB431542 can direct their differentiation into endoderm (34,35). Using this differentiation protocol (Fig. 1A), we observed a significant decrease in the expression of the pluripotency cell markers Sox2 and Oct4, which are highly expressed in R1 ES cells (Fig. 1B). Furthermore, the successive exposure of EBs to activin, SB431542, and noggin significantly increased the expression of several endodermal markers, including Afp, Foxa1, Foxa2, Sox17, Gata4 (40,41), and Cxcr4 (42) (Fig. 1C). Flow cytometry analysis confirmed the directed differentiation of EBs, with almost 50% of the cells differentiated into endoderm cells as determined by the percentage of SOX17/FOXA2-positive cells (Fig. 1D). Of note, 65% of the cells presented at least one of the endoderm markers.
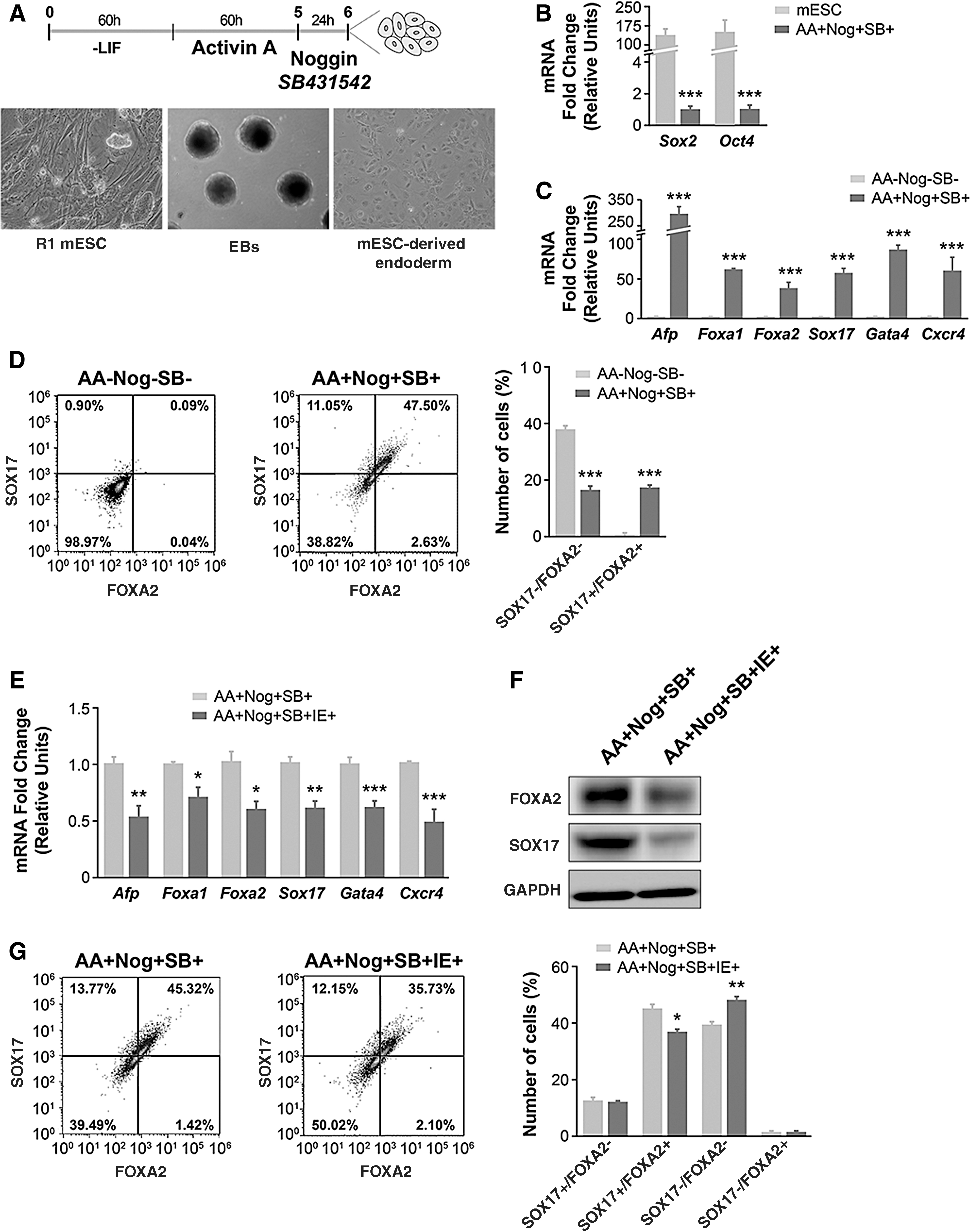
IE impairs endoderm differentiation and gene/protein expression. (
Once the endoderm culture protocol was established, we exposed cultures to IE (10−6 M NaI) to study its effect on endoderm gene expression. As shown in Figure 1E, the gene expression of all the endoderm markers analyzed was significantly lower in cultures exposed to IE from the beginning of the differentiation protocol than in control cultures. In addition to the reduction in mRNA expression (Fig. 1E), IE treatment also reduced the protein abundance of SOX17 and FOXA2 as measured by Western blotting (Fig. 1F) of ES cell-derived endoderm cells. Similarly, flow cytometry analysis confirmed the decrease in the percentage of Sox17/Foxa2-positive cells (Fig. 1G). Overall, these data unequivocally show that IE has a negative effect on gene expression of endoderm-specific genes.
Of note, this reduction in the expression of endoderm markers was not related to IE-induced apoptosis, as the profile of annexin V/PI double-stained cells with or without IE treatment was very similar (Supplementary Fig. S1).
IE alters gene expression in embryonic endoderm explants
To assess the effect of IE on endoderm differentiation markers in the early embryo (four to six somites), we isolated the presumptive ventral endoderm from E8.5 embryos (Fig. 2A) during tissue patterning, when endoderm markers are expressed. Exposure of E8.5 endoderm explants to IE for 24 hours led to a significant decrease in the mRNA abundance of several endoderm markers (Fig. 2B). Interestingly, the expression of the transcription factors Nkx2.1 and Pax8, which are master thyroid differentiation factors in thyroid gland development (43), was also significantly decreased in endoderm explants exposed to IE (Fig. 2C).
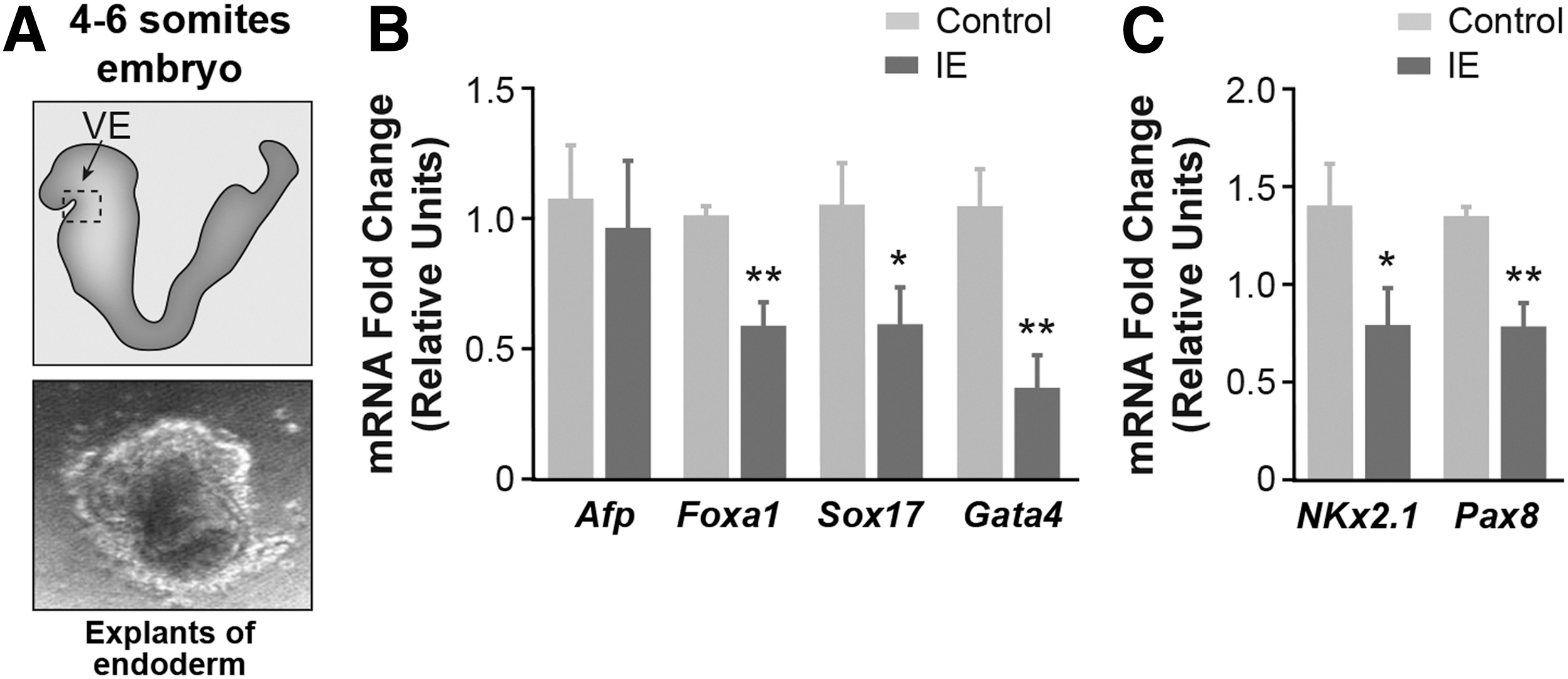
IE alters the gene expression in embryonic endoderm explants obtained from embryonic day 8.5 (E8.5) CD1 mice. (
IE induces epigenetic modifications in endoderm cells and endoderm explants
We previously showed that maternal exposure of rats to IE throughout pregnancy and lactation leads to an increase in the expression of Dnmt1 and Dnmt3 in the thyroid of adult male offspring (25). Thus, we analyzed the mRNA expression of Dnmt1 and Dnmt3 in EB-differentiated endoderm cells subjected or not to IE. As shown in Figure 3A, IE treatment significantly increased the expression of Dnmt1 mRNA without affecting the levels of Dnmt3a/b. Given that an increase in DNA methylation is commonly linked to a reduction of transcription, the repression of the endoderm markers expression observed with IE might be associated with epigenetic events. A similar result was found for embryonic endoderm explants exposed to IE for 24 hours (Fig. 3B). In agreement with the increased expression of Dnmt1 mRNA, global DNA methylation was significantly higher (threefold) in IE-exposed embryonic endoderm explants than in controls (Fig. 3C).
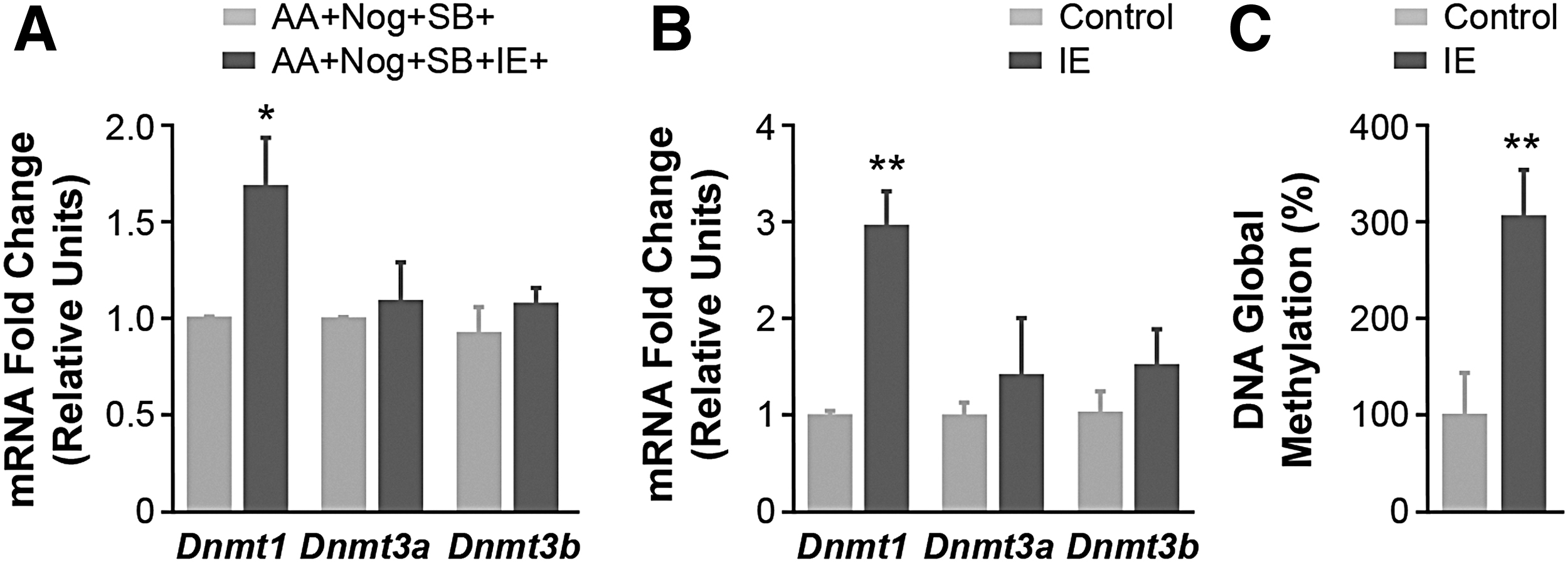
IE treatment induces epigenetic modifications in endoderm cells and embryonic explants. (
IE impairs in vitro and ex vivo differentiation of the thyroid gland
Having established a protocol for the directed differentiation of definitive endodermal cells, we next attempted to generate thyroid follicular cells by transiently overexpressing the instructive transcription factors PAX8 and NKX2.1, in combination with TSH treatment (Fig. 4A).
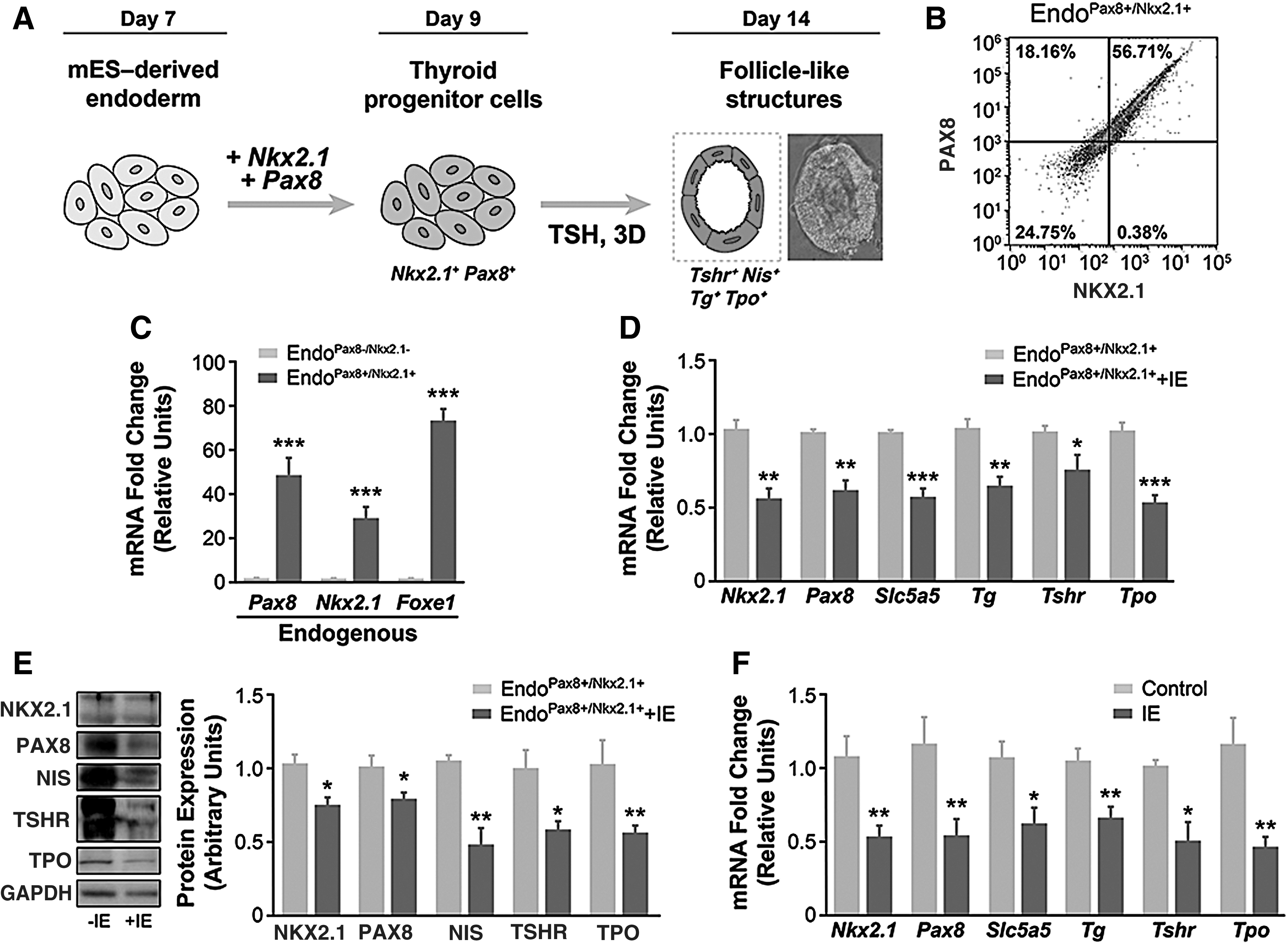
IE impairs thyrocyte gene expression. (
The efficiency of the transient transfection protocol was confirmed by flow cytometry analysis. Indeed, as shown in Figure 4B, almost 60% of the EB-derived endoderm cells showed expression of both transcription factors 48 hours after transient transfection. Importantly, the transfection of Nkx2.1- and Pax8-expressing vectors increased the expression of the endogenous forms of both transcription factors as well as the expression of Foxe1 (Fig. 4C). We also tested the effect of 3D Matrigel culture on the organization of follicle-like structures derived from endoderm-transfected cells, finding structures reminiscent of those found in the thyroid, confirming the efficiency of the differentiation protocol (Fig. 4A).
To evaluate the influence of IE exposure on thyroid-specific gene expression following endoderm cell differentiation to thyrocytes by transient overexpression of NKX2.1 and PAX8 (48 hours), we analyzed the gene expression of the thyroid markers Nkx2.1, Pax8, Slc5a5, Tg, Tshr, and Tpo in cultures subsequently exposed or not to 10−6 M NaI for 3 days, in the presence of TSH. As shown in Figure 4D, exposure of endoderm cells to IE significantly decreased the mRNA content of the thyroid markers. This was also reflected in the protein content of NKX2.1 and PAX8, and of the thyroid differentiation markers, NIS, TSHR and TPO in thyrocytes differentiated from endoderm cells (Fig. 4E and Supplementary Fig. S2). A similar pattern of gene expression was obtained in embryonic thyroid explants from E14.5 mice exposed to IE for 48 hours (Fig. 4F).
IE induces epigenetic mechanisms in thyrocytes and embryonic thyroid explants
Finally, we examined some epigenetic processes in EB-derived endoderm cells transiently expressing PAX8 and NKX2.1. Results showed that IE treatment significantly increased the mRNA expression of Dnmt1 and Dnmt3a in thyrocytes derived from ES cells, whereas Dnmt3b mRNA expression remained unchanged (Fig. 5A). Furthermore, IE treatment also increased the methylation of histone H3 lysine 27 and decreased the acetylation of histone H3 in these cells (Fig. 5B). Likewise, exposure of E14.5 embryonic thyroid explants to IE for 48 hours also augmented their mRNA expression of Dnmt1, Dnmt3a, and Dnmt3b (Fig. 5C) and increased global DNA methylation (Fig. 5D).

IE exposure induces epigenetic modifications in thyrocytes differentiated from ES cell-derived endoderm cells and in embryonic thyroid explants. (
Discussion
The data presented here suggest that IE inhibits endoderm and thyroid gene expression, a mechanism that seems to be associated with epigenetic changes during embryonic life.
Exposure of the thyroid gland to IE acutely decreases TH synthesis and secretion, a transient physiological response known as the acute Wolff–Chaikoff effect (44). Thereafter, an “escape” from the acute Wolff–Chaikoff effect usually occurs, which seems to be mediated by a decline in the expression and activity of NIS, reducing the intracellular levels of iodide (45,46). However, this autoregulatory mechanism does not fully develop until the 36th week of gestation in humans (47). Accordingly, the deleterious impact of IE exposure is evidently more severe in the fetus than in the adult.
The treatment dose used in this study aimed to mimic the levels of IE exposure that are observed in human populations. Indeed, previous epidemiological studies have demonstrated that the serum iodine concentration in humans is 5–6 × 10−7 M (48 –50). A similar concentration is also observed in pregnant and lactating women living in adequate iodine areas (51,52). Of note, pregnant women living in areas with more than adequate iodine areas present with a serum iodine concentration of ∼8 × 10−7 M (53). Furthermore, studies using experimental rat models demonstrate that the serum iodine concentration in control animals is approximately 4–5 × 10−7 M (54,55). Accordingly, the iodide dose used in the present study is about twice the usual serum iodine concentration observed in individuals exposed to adequate iodine condition (rats or humans) and is similar to that observed in pregnant women living in more than adequate iodine areas.
The intrauterine environment plays a pivotal role in the early gene expression programming of adult diseases through epigenetic mechanisms (56 –58). Along this line, our previous data in rats demonstrated that maternal exposure to IE reduces the mRNA and protein content of important thyroid transcription factors in adult offspring, which correlates with decreased expression of proteins involved in TH synthesis (25). This is consistent with data showing that impairment of thyroid transcription factor expression, by mutation or epigenetic modification, induces thyroid dysgenesis (59,60). Our present data reinforce the notion that the repression of Pax8 and Nkx2.1 is a consequence of IE exposure during embryonic life.
The thyroid gland is the first endocrine gland to develop in the fetus and relies on an adequate maternal iodine supply to produce TH. Although the development of the fetal hypothalamic–pituitary–thyroid (HPT) axis occurs independent of the maternal equivalent, it depends on the maternal–placental system for the adequate supply of iodide (61). Therefore, both iodine deficiency and IE might trigger deleterious effects on the development of the HPT axis in the fetus.
In this context, preliminary data from our group indicate that the expression of thyroid genes is significantly reduced in postnatal day 1 rats that are exposed to IE during the intrauterine period, which is in agreement with data reporting the development of goiter in the fetuses of mothers exposed to high iodine intake (62,63). It is important to mention that in patients with partial DUOX2 defects, IE is reported to be potentially protective for thyroid function at birth (64,65). Nevertheless, the data presented herein suggest that, in the absence of mutations or defects in the activity of proteins related to TH synthesis, the early exposure to high concentrations of iodine is detrimental, to both endoderm and thyroid development and differentiation.
The mechanism by which iodine elicits the repression of instructional transcription factors is unknown. Moreover, it is intriguing to speculate how this effect can take place in endoderm as, to the best of our knowledge, the existence of NIS in this embryonic layer is yet to be demonstrated. That being said, it is well accepted that other transporters including chloride channels can transport iodide when cells are exposed to IE (1), and such transporters have been described in the endoderm (66). It is also important to highlight that iodide is a scavenger of reactive oxygen species (ROS), which themselves can have an effect on endoderm, as described in differentiated induced pluripotent stem cells (67). Furthermore, some actions of iodide are due to its capacity as a reducing agent (68), similar to other halogenated compounds.
This raises the question of whether the observed effects on genetic and epigenetic processes is specific for iodide or whether it is due to biochemical changes induced by halogen excess, a possibility that remains to be explored in futures studies. However, it is known that iodide has a greater reducing capacity than bromide (69).
We recognize that our study was based on in vitro and ex vivo models, however, the results corroborate the findings of IE-induced impairment of thyroid gene expression/function previously described in rats and in humans.
Endoderm is one of the three major germ layers formed during in vivo vertebrate development, giving rise to a variety of organs including thyroid, liver, lungs, and pancreas (70 –72). Accordingly, several molecular and morphological events occur in the endoderm at E8.5, and tissue programs are activated by different inductive signals mainly regulated by transcriptional activators including FOXA1, FOXA2, GATA4, GATA6, and SOX17 (40,73 –75). These “pioneer” factors have the ability to open chromatin through high-affinity DNA binding sites and by interacting with histones H3 and H4 through their C-terminal domains (76), facilitating the binding of additional transcription factors.
In the present study, we successfully induced endoderm differentiation following an established protocol in mouse ES cells (34,35,77,78). We found that IE treatment significantly decreased the expression of several endoderm transcription factors in this model, and these results were confirmed in IE-treated endoderm explants from mouse embryos. Therefore, in addition to the presumptive deleterious effects of IE exposure on thyroid development, this treatment might also impair the development/differentiation of other endoderm-derived organs. Indeed, our previous studies showed that fetus absorption was increased in iodine-treated pregnant rats, suggesting that the normal embryonic development is impaired by IE exposure (25).
In addition to the expression of endoderm-specific transcription factors, it has been well described that at E9.5, a group of endoderm cells co-expressing four transcription factors (NKX2.1, PAX8, FOXE1 and HHEX), detaches from the primitive pharynx and migrates to the neck (30). At E14.5, these transcription factors control the expression of the thyroid differentiation mRNAs that encode for the proteins involved in TH biosynthesis and in thyroid follicle formation (43).
Thus, a decline in the expression of thyroid differentiation genes in IE-exposed animals could be explained by the early diminished expression of Pax8 and Nkx2.1, as observed in this study. Indeed, the lower abundance of PAX8 and NKX2.1 in endoderm explants, thyroid explants, and also in thyrocytes derived from mouse ES cells suggests that the reduction of thyroid gene expression observed in male rats during adult life is related to an early reprogramming of thyroid transcription factor expression during embryonic life.
PAX8 and NKX2.1 directly interact and synergistically activate thyroid-specific transcription even in nonthyroid cells (79 –81). In agreement with this, through transient transfection of Pax8- and Nkx2.1-expressing vectors, we successfully generated thyroid cells from ES cell-derived endoderm cells. Indeed, besides their expression of thyroid differentiation markers, these cells were able to form follicle-like structures in 3D cultures. Our protocol was efficient, as shown by the expression of both transcription factors in almost 60% of the transfected cells. Also, the method used was less time-consuming and aggressive to ES cells than other protocols since no additional selection with antibiotics was necessary (82). Finally, the concomitant overexpression of Pax8 and Nkx2.1 was accompanied by their endogenous expression, suggesting an autoinduction mechanism of these transcription factors on their own expression during thyroid development.
Our data indicate that IE treatment, both in vitro and ex vivo, triggers epigenetic mechanisms, reported here as increased DNA methyltransferase expression, enhanced DNA methylation, and increased methylation and reduced acetylation of the histone H3 N-terminal tail. In general, these epigenetic modifications are related to transcriptional repression (83,84) and, therefore, could be associated with the repression of endoderm and thyroid gene expression during embryonic life.
Finally, our in vitro and ex vivo results strengthen the hypothesis that the inhibitory effects of IE exposure for gene expression of the fetal thyroid are directly triggered by iodide and do not depend on the alterations of maternal TH levels. As discussed above, the redox state has been related to modifications in the cellular epigenome (85), and previous studies have demonstrated that IE can also increase ROS production in thyrocytes (39,46,67,86). Therefore, the epigenetic changes observed here could also be mediated by the IE-induced oxidative stress in the in vitro and ex vivo models. This needs to be investigated in future studies.
Importantly, it has been proposed that developmental gene programming works as a “predictive adaptive response.” This hypothesis implies that the gene expression regulation in the fetus is part of a physiological adjustment that occurs in the intrauterine environment to guarantee the survival of the offspring in a similar postnatal environment to their mothers (87). Therefore, the IE-induced decrease of thyroid gene expression might be related to an adaptive response in the offspring to an iodine-rich environment. Conversely, the transfer of IE-programmed animals to an iodine-sufficient or iodine-deficient environment could induce hypothyroidism, which was observed in our previous study (25), since these animals showed decreased expression of the genes that encode for proteins involved in TH synthesis.
In summary, our data suggest that IE directly affects the endoderm and thyroid during embryonic life and induces epigenetic mechanisms that could repress the expression of instructive signals in these tissues. Moreover, since endoderm markers were repressed by IE exposure, a direct role of this treatment on additional endoderm-derived organs (such as liver, pancreas, lung) cannot be ruled out. Finally, these results reinforce the existence of a predictive adaptive response triggered by the maternal intrauterine environment.
Footnotes
Acknowledgments
We are grateful to Dr. Kenneth McCreath for helpful comments and English correction, to Dr. Roque Bort (Instituto de Investigación Sanitaria La Fe, Valencia, Spain) for providing aliquots of SOX17 and FOXA2 antibodies, and to Javier Pérez (Instituto de Investigaciones Biomédicas, Madrid, Spain) for the artwork.
Author Disclosure Statement
The authors have nothing to disclose.
Funding Information
This study was supported by grants from Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) to C.S.-N. (12/24391-6, 15/16695-3, 16/18517-8) and to M.T.N. (13/05629-4); and grants from Ministerio de Economía, Industria y Competitividad (Spain; MINECO) (SAF2016-75531-R) and from Comunidad Madrid (Spain; B2017/BMD-3724 TIRONET CM2) to P.S.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Table S1
Supplementary Table S2