
Abstract
Background:
Exosomes or small extracellular vesicles secreted from cells are nanovesicles with a diameter of 40–150 nm, which play a number of roles in both physiologic and pathologic processes. In Graves’ disease (GD), autoantibodies bind to the thyrotropin receptor (TSHR) on the surface of thyroid follicular epithelial cells and stimulate thyroid growth and thyroid hormone synthesis and secretion via cyclic adenosine monophosphate (cAMP) production. The present study aimed to confirm the existence of TSHR in exosomes secreted from thyroid cells and to define the role of TSHR exosomes in GD.
Methods:
Exosomes were isolated by differential centrifugation from the culture medium of the human thyroid follicular epithelial cell line (NTHY-ori 3-1) and thyroid carcinoma cell lines (8305C, 8505C, and FTC-133). TSHR expression in cell lysates and exosomes was evaluated by Western blot analysis. In order to study the function of TSHR exosomes, human embryonic kidney (HEK) 293 cells stably expressing TSHR (HEK/TSHR) were established. Using exosomes isolated from both HEK and HEK/TSHR cells, the binding capacity of the M22 human monoclonal autoantibody to TSHR exosomes and their effect on M22-mediated stimulation of cAMP production in HEK/TSHR cells were evaluated. As a positive control for the functional assay, human recombinant TSHR chimera protein capable of binding to TSH was used.
Results:
TSHR was detected in exosomes from cancer cells as well as normal epithelial cells. An in vitro binding assay showed that alkaline phosphatase-labeled M22 bound to TSHR exosomes in a dose-dependent manner. M22 dose-dependently stimulated intracellular cAMP production in HEK/TSHR cells. The addition of exosomes from HEK/TSHR cells but not those from parental HEK cells significantly ameliorated cAMP production stimulated by treatment with M22 in HEK/TSHR cells. A decoy effect similar to TSHR exosomes was observed for human recombinant TSHR chimera.
Conclusions:
The results suggest that exosomes expressing TSHR may be secreted from normal and cancerous thyroid cells. In the thyroid gland of patients with GD, TSHR exosomes may exert a decoy effect by sequestering autoantibody, thereby ameliorating autoantibody-mediated activation of thyroid function.
Introduction
The thyrotropin receptor (TSHR) is a glycoprotein hormone receptor that belongs to a subfamily of G protein-coupled receptors (1). TSHR is synthesized as a single-chain precursor with an intrinsic disulfide-bridged loop, which is cleaved within the loop, forming a mature type of receptor composed of two subunits (extracellular A-subunit and transmembrane and cytosolic B-subunit) linked by the disulfide bridge (2). In the thyroid gland, TSH secreted from the anterior pituitary binds to TSHR and plays a fundamental role in the regulation of thyroid cell proliferation and differentiation and thyroid hormone synthesis and secretion. In patients with Graves’ disease (GD), autoantibodies targeting the TSHR mimic the actions of TSH and cause thyroid overactivity characterized by high serum thyroid hormone levels and low serum TSH levels. The human monoclonal autoantibody to TSHR (M22) has been isolated from peripheral blood lymphocytes of a patient with GD, which acts as a powerful thyroid stimulator and inhibits binding of TSH to TSHR (3). A M22 monoclonal antibody-based TSHR antibody (TRAb) enzyme-linked immunosorbent assay kit is commercially available, and TRAb is used as a useful marker for diagnosing and monitoring GD. TSHR expression is preserved in adenomatous nodules and papillary carcinomas, and it is upregulated in the thyroid gland of patients with GD (4). The mechanism underlying the elevation of TSHR in GD, however, remains largely unknown (5).
Exosomes or small extracellular vesicles are nanovesicles secreted from cells with a diameter of 40–150 nm, and they contain biomolecules such as proteins, miRNAs, RNAs, and DNAs present in cells from which they were released (6,7). Thus, exosomes in body fluids such as blood and urine can be used as biomarkers for diseases such as cancer (8,9). On the other hand, exosomes play a variety of roles in cell-to-cell communication in both physiologic and pathologic conditions at the site where they are secreted and also at distant sites after their release into the circulation. For instance, ligands on the exosome surface alter intracellular signaling of recipient cells via binding to their receptors (10). Exosomes also influence the function of cells by transferring their cargo such as proteins and miRNAs (11).
The present study hypothesized that exosomes secreted from thyroid follicular epithelial cells may express TSHR and that TSHR on exosomes may exert a decoy effect on M22-mediated thyroid cell activation in GD by sequestering M22. To prove this hypothesis, first the study examined whether exosomes isolated from thyroid cell lines contain TSHR. Then, the study generated a cell line stably expressing TSHR as a model for thyrocytes in GD where TSHR is upregulated. Lastly, the effect of TSHR exosomes on M22-mediated stimulation of intracellular cyclic adenosine monophosphate (cAMP) production in the stably transfected cells was studied using exosomes isolated from these cells as well as from parental cells.
Methods
Cell culture
The NTHY-ori 3-1 human thyroid follicular epithelial cell line immortalized by SV-40 was purchased from the European Collection of Cell Cultures (Salisbury, United Kingdom). The human embryonic kidney (HEK) 293 cell line, the 8505C poorly differentiated papillary thyroid adenocarcinoma cell line, the 8305C anaplastic thyroid carcinoma cell line, and the FTC-133 follicular thyroid carcinoma cell line were from the RIKEN Cell Bank (Wako, Saitama, Japan). NTHY-ori 3-1 and 8305C cells were cultured in RPMI 1640 supplemented with 10% fetal bovine serum (FBS; Sigma–Aldrich, St. Louis, MO). HEK and 8505C cells were cultured in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% FBS. FTC-133 cells were cultured in DMEM/F-12 with 10% FBS. All cell cultures were maintained in a humidified incubator at 37°C under 5% CO2.
Generation of HEK cells stably expressing TSHR
Human TSHR cDNA was amplified by polymerase chain reaction (PCR) using KOD polymerase (Toyobo, Osaka, Japan) from a human thyroid gland cDNA library (Takara Bio, Shiga, Japan). The nucleotide sequences of the primer pair were as follows: forward 5′-gtc aGA ATT Caa ata gcc ccg agt ccc gtg-3′ and reverse 5′-gtc aGT CGA Cgt tcc cct acc att gtg agt-3′, where capital letters indicate EcoRI and SalI sites, respectively. After digestion, PCR products were ligated into the PCI-neo vector (Promega, Madison, WI). The inserted cDNA sequence was verified by direct sequencing. HEK cells were transfected with the TSHR expression plasmid along with pBAsi-hU6 Pur DNA (Takara Bio) using FuGENE HD (Promega). After selection under puromycin (2 μg/mL), individual clones were picked, expanded, and tested for TSHR expression. A clone with the highest TSHR expression (HEK/TSHR) was used.
Isolation of exosomes by differential centrifugation
Cells seeded on a 100 mm dish were cultured for 72 hours in medium containing 10% exosome-deprived FBS. Exosomes were isolated from the conditioned medium, as previously described (12). Briefly, the medium was centrifuged at 2000 g for 10 minutes to eliminate cells. Then, the supernatant was centrifuged at 12,000 g for 30 minutes to remove debris. After filtration through a 0.22 μm polyvinylidene difluoride (PVDF) membrane, exosomes were pelleted by ultracentrifugation at 110,000 g for 70 minutes. Pellets were re-suspended in phosphate-buffered saline (PBS) and stored at −80°C until use.
Western blot analysis
Total cell lysates were prepared in ice-cold lysis buffer (1% Igepal CA-630, 1% sodium deoxycholate, 0.1% sodium dodecyl sulfate [SDS], 150 mM NaCl, 25 mM Tris-HCl; pH 7.6) containing protease inhibitor cocktail. Cell lysates and exosomes were subjected to electrophoresis on SDS-polyacrylamide gels and transferred to PVDF membranes. After blocking in 5% skim milk, membranes were hybridized with goat polyclonal antibody against TSHR (diluted 1:200, sc-7816; Santa Cruz Biotechnology, Santa Cruz, CA) or rabbit monoclonal antibody against CD9 (diluted 1:1000, #13174; Cell Signaling Technology, Beverly, MA) and then with a horseradish peroxidase-linked secondary antibody. After washing, bound proteins were visualized using an ECL Prime Western blotting detection system (GE Healthcare, Little Chalfont, United Kingdom). Human thyroid membrane extract obtained from Abcam (Cambridge, United Kingdom) was used as a positive control for the detection of TSHR.
Binding of autoantibody to exosomes
Exosomes isolated from HEK and HEK/TSHR cells were incubated in 50 mM sodium bicarbonate/carbonate buffer (pH 9.6) overnight at 4°C on a 96-well microplate (MaxiSorp; Thermo Fisher Scientific, Waltham, MA). After washing with PBS with Tween 20 (PBS-T), blocking was performed in Tris-buffered saline with 1% bovine serum albumin for one hour at room temperature. After washing with PBS-T, exosomes attached to the well were incubated for one hour at room temperature with alkaline phosphatase-labeled M22 (Fuji Rebio, Tokyo, Japan). After washing with PBS-T, alkaline-phosphatase substrate CDP-Star (Thermo Fisher Scientific) was added, and chemiluminescence was determined using an EnVision multilabel reader (PerkinElmer, Waltham, MA).
Decoy effect of TSHR exosomes on autoantibody-mediated cAMP production
HEK/TSHR cells were seeded onto 48-well type 1 collagen-coated plates (Corning, New York, NY) and cultured overnight. Cells were then preincubated in serum-free medium (HyClone SFM4HEK293; GE Healthcare Hyclone, Chicago, IL) for four hours. During the last 30 minutes of the preincubation, reactions between M22 autoantibody and exosomes isolated from HEK or HEK/TSHR cells were performed in serum-free medium at 37°C for 30 minutes. Immediately after the reaction, preincubation medium was replaced with the medium. After incubation for 30 minutes, cells were lysed, and the intracellular cAMP level was determined using a Cyclic AMP Select EIA kit according to the manufacturer's protocol (Cayman Chemical, Ann Arbor, MI). As a positive control, Chinese Hamster Ovary (CHO) cell line-derived human TSHR ectodomain (Met22 to Gly413) chimera protein (8950-TR-050, R&D Systems, Minneapolis, MN) was used, which was shown to bind to human recombinant TSH.
Statistical analysis
Statistical differences were assessed by one-way analysis of variance followed by Tukey's test. All statistical analyses were performed using R v 3.4.1 (The R Foundation for Statistical Computing, Vienna, Austria). A p-value of <0.05 was considered statistically significant.
Results
Presence of TSHR in exosomes secreted from normal and cancerous thyroid cells
To examine whether exosomes expressing TSHR are secreted from thyroid cells, exosomes were isolated from the cell culture medium of normal thyroid follicular epithelial cell line (NTHY-ori 3-1) and thyroid carcinoma cell lines (8505C poorly differentiated papillary adenocarcinoma, 8305C anaplastic carcinoma, and FTC-133 follicular carcinoma) by differential centrifugation. In the case of NTHY-ori 3-1 cells, TSHR as well as an exosomal marker protein, CD9, were enriched in exosomes compared to cell lysates (Fig. 1A). Similarly, the levels of TSHR in exosomes from thyroid carcinoma cell lines were higher than those in cell lysates. The abundance of CD9 and TSHR were different among thyroid carcinoma cell lines (Fig. 1B). These results suggest that both normal and cancerous thyroid cells secrete exosomes expressing TSHR.
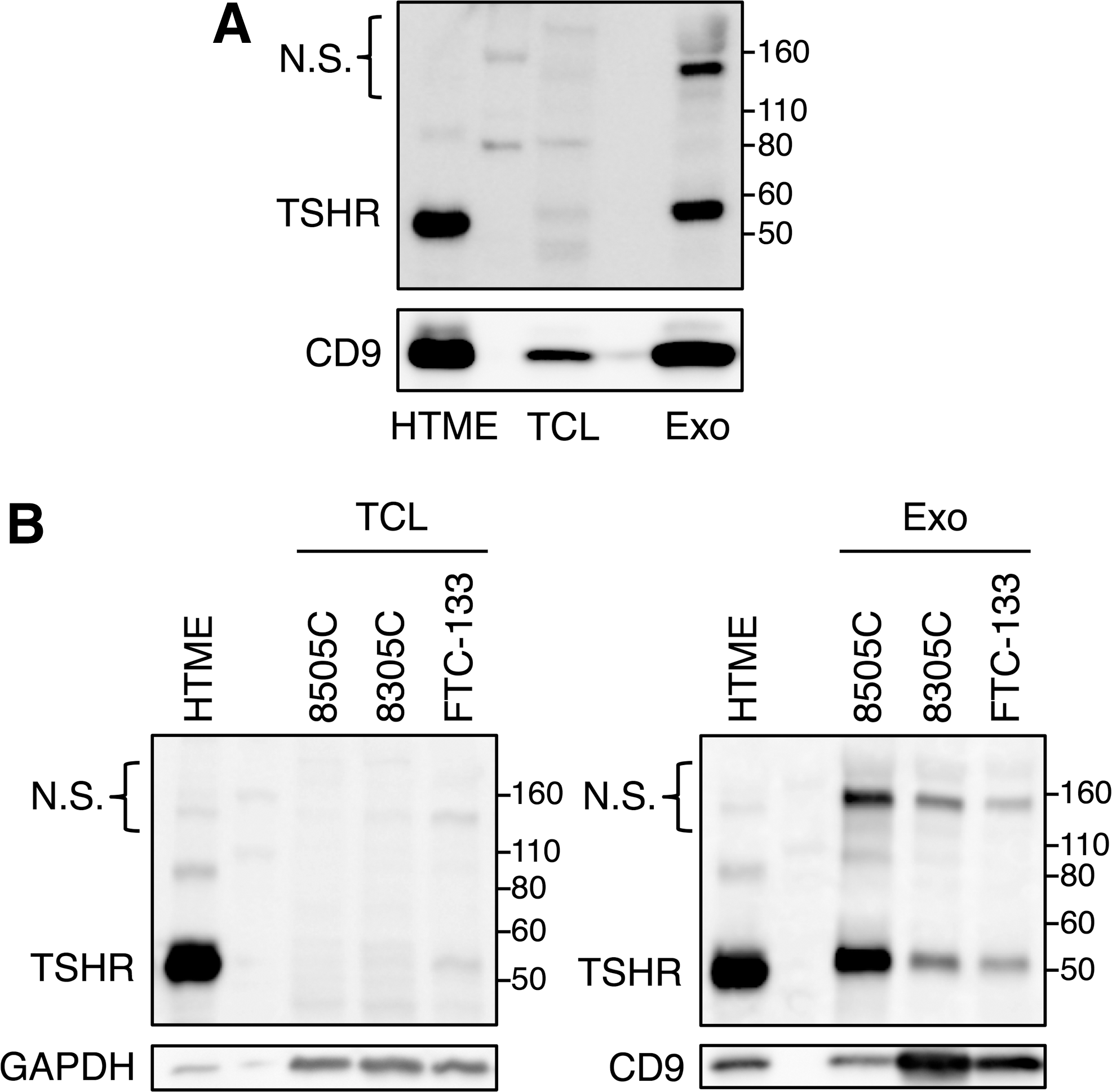
Expression of TSHR in exosomes isolated from normal and cancerous thyroid cells. Exosomes were isolated from the culture medium of a thyroid follicular epithelial cell line (NTHY-ori 3-1) and thyroid carcinoma cell lines (8505C poorly differentiated papillary adenocarcinoma, 8305C anaplastic carcinoma, and FTC-133 follicular carcinoma) by differential centrifugation. Cell lysates and exosomes of NTHY-ori 3-1 cells (
Establishment of HEK/TSHR cells
In order to examine the function of TSHR exosomes in GD, HEK/TSHR cells were generated as a model for thyrocytes in GD where TSHR is upregulated. As shown in Figure 2A, the TSHR levels in both cell lysates and exosomes were increased in HEK/TSHR cells compared to parental HEK cells. TSHR was detectable in exosomes from HEK cells but was markedly increased in those from HEK/TSHR cells. In the following experiments, therefore, exosomes from HEK/TSHR cells were used as TSHR exosomes and those from HEK cells were used as control exosomes expressing a trace amount of TSHR.
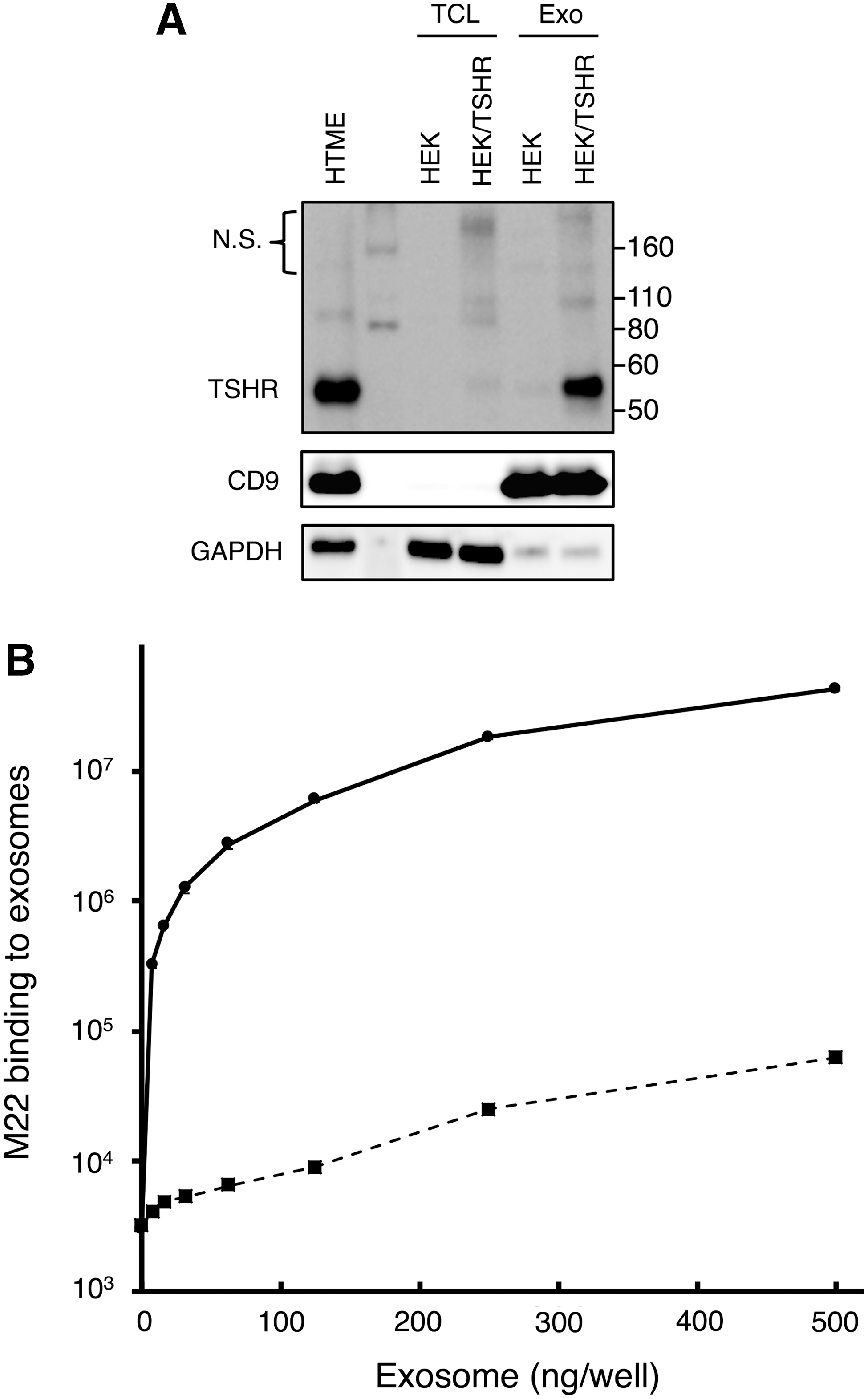
Establishment of HEK cells stably expressing TSHR and binding characteristics of M22 autoantibody to TSHR exosomes. HEK 293 cells stably expressing TSHR were developed (HEK/TSHR).
Binding of M22 autoantibody to TSHR exosomes
It was hypothesized that TSHR exosomes secreted from thyroid follicular epithelial cells may exert a decoy effect by sequestering autoantibody in GD. To examine the binding of the M22 human monoclonal autoantibody to TSHR exosomes, an in vitro binding assay was performed. Alkaline-phosphatase-labeled M22 was used to detect its binding to TSHR on exosomes immobilized on microplate wells. Exosomes from both HEK/TSHR and HEK cells showed a dose-dependent increase in the signal intensity. The M22 binding to exosomes from HEK/TSHR cells was higher than that from HEK cells by >1000-fold (Fig. 2B), which was consistent with markedly lower TSHR expression in HEK exosomes (Fig. 2A). Non-labeled M22 inhibited the binding of labeled M22 to HEK/TSHR exosomes in a dose-dependent manner, demonstrating specific binding of M22 to TSHR on exosomes (Supplementary Fig. S1). These results indicate that TSHR exosomes are capable of binding to M22 autoantibody.
Inhibition of M22-mediated stimulation of cAMP production by TSHR exosomes
Next, the study examined the effect of TSHR exosomes on M22-mediated stimulation of cAMP production in HEK/TSHR cells. First, the temporal changes in intracellular cAMP production in response to M22 were tested. The intracellular cAMP levels increased in a dose-dependent manner and peaked at 30 minutes after stimulation with M22 (Fig. 3A). No cAMP increase was observed in the absence of M22. Second, the cAMP levels were measured 30 minutes after stimulation with M22 (100 ng/mL) in the presence of HEK/TSHR and HEK exosomes (5, 20, and 50 μg/mL). The results showed that HEK/TSHR exosomes significantly inhibited M22-stimulated cAMP production in a dose-dependent manner (Fig. 3B). Lastly, the inhibitory effect of a human TSHR ectodomain chimera protein, which was shown to bind to human recombinant TSH, on M22-mediated stimulation of cAMP production was tested. Similar to the case of TSHR exosomes, the chimera added to the culture medium dose-dependently reduced M22-stimulated cAMP production (Fig. 3C). Taken together, these results suggest that TSHR exosomes may sequester M22 and inhibit M22-mediated stimulation of cAMP production, thereby exerting a decoy effect.
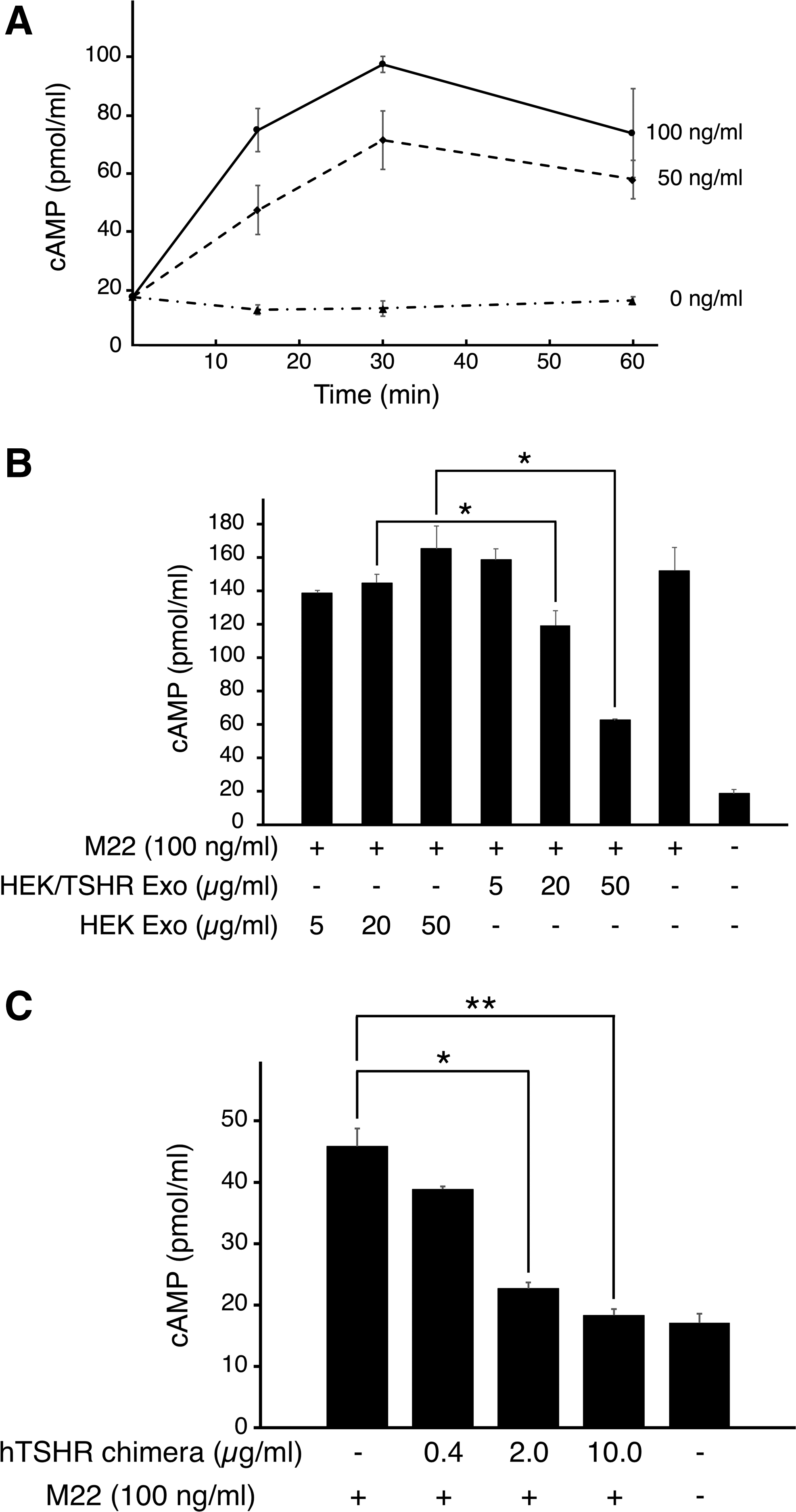
Effects of TSHR exosomes on M22-mediated stimulation of intracellular cAMP production. (
Discussion
The present study shows that TSHR is present in exosomes secreted from the NTHY-ori 3-1 human follicular epithelial cell line and the 8505C, 8305C, and FTC-133 thyroid carcinoma cell lines. Using exosomes isolated from HEK/TSHR and HEK cells as TSHR exosomes and control exosomes, respectively, the study demonstrates that TSHR exosomes may bind to and sequester M22 autoantibody and thereby inhibit M22-stimulated cAMP production in HEK/TSHR cells generated as a model for thyrocytes in GD.
Upon TSH binding, TSHR preferentially couples to the Gsα subunit, resulting in activation of adenylyl cyclase and an increase in intracellular cAMP levels. Internalized TSHR co-localizes with early endosomes, and a majority is recycled to the cell surface (13,14), while TSH is degraded in the lysosomes. Inward budding of early endosomes leads to multivesicular body formation containing intraluminal vesicles. Exosomes are released into the extracellular milieu when multivesicular bodies fuse with the outer cell membrane (15). Thus, TSHR exosomes may be generated from the endocytic pathway. On the other hand, microvesicles (MVs) arise from the outward budding and fission of the plasma membrane (16). It has been reported that an endocrine disruptor, dichlorodiphenyltrichloroethane, alters the membrane raft constitution and inhibits TSHR internalization, leading to a release of MVs containing TSHR (17). In this study, however, MVs were eliminated from exosome samples by centrifugation at 12,000 g. TSHR exosomes were secreted from thyroid carcinoma cell lines, as well as normal follicular epithelial cells, which was consistent with a previous report showing TSHR expression in thyroid carcinoma and normal tissues (4).
Chazenbalk et al. reported that a secreted recombinant TSHR ectodomain variant neutralizes TSHR autoantibodies present in the sera of GD patients (18). The present study shows that TSHR molecules on exosomes bind to TSHR autoantibody as well, and that TSHR exosomes exert a decoy effect. A decoy effect has been previously demonstrated for exosomes secreted from breast cancer cells. Specifically, exosomes expressing human epidermal growth factor receptor 2 (HER2) bind to a therapeutic monoclonal anti-HER2 antibody, herceptin, thereby allowing continued tumor cell proliferation (19). The present results suggest that TSHR exosomes secreted from thyroid follicular epithelial cells may neutralize or attenuate overstimulation by TSHR autoantibodies in thyroid glands of GD patients. It may also be possible that TSHR exosomes secreted from thyroid carcinoma cells bind to TSH and affect TSH-dependent proliferation.
This study demonstrates a decoy effect mediated by TSHR exosomes on M22-stimulated cAMP production using HEK/TSHR cells and exosomes from HEK/TSHR and HEK cells. Initially, NTHY-ori 3-1 cells were treated with M22, and intracellular cAMP levels were measured, but the response was not strong enough to study the decoy effect, presumably because of limited expression of TSHR (20). It was also found that the yield of exosomes from the cell line was too low to perform functional studies. Indeed, TSHR expression in the thyroid gland was shown to be increased in GD compared to control (4). In order to mimic the pathologic conditions found in GD, cells stably overexpressing TSHR were therefore established and used. Since autoantibodies but not TSH, which is the physiologic ligand for TSHR, stimulate thyrocytes in GD, HEK/TSHR cells were treated with M22.
There have been a few reports that examined blood exosomes in patients with thyroid carcinoma. To the authors’ knowledge, no study has investigated blood exosomes secreted from the thyroid glands of GD patients. It is likely that TSHR exosomes secreted from thyroid follicular epithelial cells may be released into the circulation and will therefore be present in the blood, although TSHR exosomes have yet to be detected successfully in the serum of GD patients. Development of a highly sensitive immunoassay would allow the detection of TSHR exosomes in the blood. It is tempting to speculate that TSHR exosomes may reach other organs and tissues and play critical roles in the manifestation of extrathyroidal symptoms of GD. On the other hand, a previous report using a genetically-engineered spontaneous mouse model of GD demonstrated that extrathyroidal TSHR antigen is likely to aggravate rather than attenuate hyperthyroidism (21). It is also well known in other diseases that removing autoantibodies, for example by plasmapheresis, only transiently reduces antibody levels and is followed by a rebound in antibody secretion (22). It is therefore possible that TSHR exosomes in the blood might exacerbate hyperthyroidism. Further research is needed to clarify the systemic roles of TSHR exosomes.
In conclusion, evidence is provided for the presence of TSHR exosomes secreted from normal and cancerous thyroid cell lines, and it is proposed that TSHR exosomes may exert a decoy effect by binding to and sequestering autoantibodies, thereby attenuating autoantibody-mediated activation in GD patients (Fig. 4).
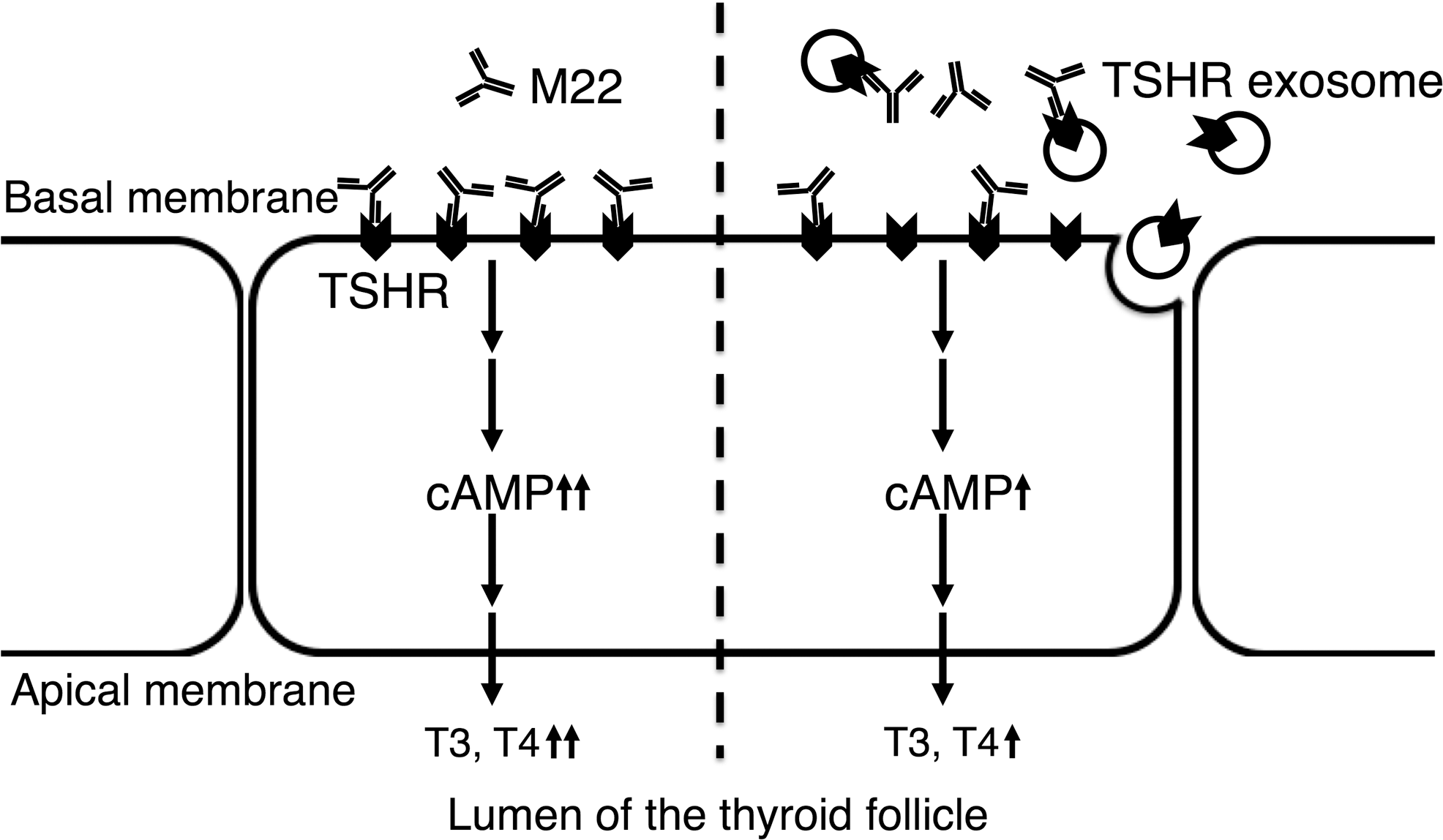
Schematic illustration of the decoy effect of TSHR exosomes on M22-mediated stimulation of intracellular cAMP production. In GD, M22 autoantibody binds to TSHR expressed on thyrocytes and stimulates intracellular cAMP production, leading to their activation including thyroid hormone synthesis and secretion (left). In the proposed model (right), thyrocytes secrete exosomes expressing TSHR, which bind to and sequester M22, resulting in a decrease of M22-mediated stimulation of cAMP production (decoy effect).
Footnotes
Author Disclosure Statement
The authors declare that they have no competing interests.
Supplementary Material
Supplementary Figure S1