
Abstract
Background:
Hyalinizing trabecular tumor (HTT) is a rare thyroid neoplasm with a characteristic trabecular growth pattern and hyalinization. This lesion has been the subject of long-term controversy surrounding its genetic mechanisms, relationship to papillary thyroid carcinoma (PTC), and malignant potential. Due to the presence of nuclear features shared with PTC, HTT frequently contributes to a false-positive cytology, which hampers patient management. The goal of this study was to apply genome-wide sequencing analyses to elucidate the genetic mechanisms of HTT and its relationship to PTC.
Methods:
Whole-exome, RNA-Seq, and targeted next-generation sequencing analyses were performed to discover and characterize driver mutations in HTT. RNA-Seq results were used for pathway analysis. Tissue expression of GLIS3 and other proteins was detected by immunohistochemistry. The prevalence of GLIS fusions was studied in 17 tumors initially diagnosed as HTT, 220 PTC, and 10,165 thyroid fine-needle aspiration samples.
Results:
Using whole-exome and RNA-Seq analyses of the initial three HTT, no known thyroid tumor mutations were identified, while in-frame gene fusion between PAX8 exon 2 and GLIS3 exon 3 was detected in all tumors. Further analysis identified PAX8–GLIS3 in 13/14 (93%) and PAX8–GLIS1 in 1/14 (7%) of HTT confirmed after blind pathology review. The fusions were validated by Sanger sequencing and FISH. The fusions resulted in overexpression of the 3′-portion of GLIS3 and GLIS1 mRNA containing intact DNA-binding domains of these transcription factors and upregulation of extracellular matrix genes including collagen IV. Immunohistochemistry confirmed upregulation and deposition of collagen IV and pan-collagen in HTT. The analysis of 220 PTC revealed no PAX8–GLIS3 and one PAX8–GLIS1 fusion. PAX8–GLIS3 was prospectively identified in 8/10,165 (0.1%) indeterminate cytology fine-needle aspiration samples; 5/5 resected fusion-positive nodules were HTT on surgical pathology.
Conclusions:
This study demonstrates that GLIS rearrangements, particularly PAX8–GLIS3, are highly prevalent in HTT but not in PTC. The fusions lead to overexpression of GLIS, upregulation of extracellular matrix genes, and deposition of collagens, which is a characteristic histopathologic feature of HTT. Due to unique genetic mechanisms and an indolent behavior, it is proposed to rename this tumor as “GLIS-rearranged hyalinizing trabecular adenoma.”
Introduction
Hyalinizing trabecular tumor (HTT) is a rare neoplasm derived from thyroid follicular cells that shows no invasion and has a characteristic trabecular growth pattern and hyalinization. This lesion has been the subject of long-term controversy surrounding its genetic mechanisms, relationship to papillary thyroid carcinoma (PTC), and malignant potential.
The first detailed description of this tumor in the modern era was provided by Carney et al., who reported in 1987 a series of 11 tumors with distinct histopathological appearance and named them “hyalinizing trabecular adenoma” (1). All of these tumors had a prominent trabecular pattern, abundant intra-trabecular hyalinized stroma, and nuclear features overlapping with those of PTC, but they were well circumscribed or encapsulated, had no invasion, and demonstrated no tumor recurrence or metastasis on a 10-year mean follow-up (1). A year later, the same tumor was described by Bronner et al. in a series of nine cases as “paraganglioma-like adenoma of the thyroid” (2).
In 1991, a study of nine thyroid tumors with trabecular pattern and hyalinization was reported, and one of those cases had a lymph node metastasis at presentation (3). Although the latter case was diagnosed on incisional biopsy only, the authors argued that tumors with such appearance may have malignant behavior and should be designated as “hyalinizing trabecular tumor.” Several subsequent studies reported rare tumors with trabecular appearance and hyalinization that were associated with blood vessel or tumor capsule invasion and distant metastasis, as well as areas of classic PTC merging with this tumor (4 –6). Together with the prominent nuclear features of PTC seen in the tumor cells, these reports argued against the benign nature of hyalinizing trabecular adenoma. As the controversy persisted, the 2004 World Health Organization (WHO) classification placed this tumor in a separate category of tumors with low malignant potential and designated it as “hyalinizing trabecular tumor” (7). The same approach was followed by the most recent 2017 WHO classification of tumors of endocrine organs (8).
The biological relationship of this tumor to PTC was suggested by two publications reporting that HTT frequently harbor RET/PTC rearrangements (9,10), a characteristic genetic feature of PTC. However, these reports raised a controversy by themselves due to concerns about the histologic criteria for tumor selection and use of highly sensitive assays for RET/PTC detection prone to false positivity (11). A subsequent study revealed no RET/PTC rearrangements in a series of 18 HTT analyzed by reverse transcription polymerase chain reaction (RT-PCR) (12). Furthermore, these tumors were found to harbor no BRAF or RAS mutations, which are highly prevalent in classic type and follicular variant PTC (12 –18), pointing to a possibility that HTT is a distinct tumor and not a variant of PTC.
The controversy was further fueled by the fact that HTT cells exhibit pronounced nuclear features of PTC, including nuclear pseudoinclusions, and are frequently diagnosed as either PTC or suspicious for PTC in fine-needle aspiration (FNA) cytology samples (19).
As a result, over the last 15 years, this lesion has been considered a borderline tumor with low malignant potential rather than a benign adenoma. This is despite the fact that no adverse outcome has ever been documented in a tumor with this morphology lacking invasive features at presentation (20).
The goal of this study was to apply genome-wide molecular analyses based on next-generation sequencing (NGS) to elucidate the genetic mechanisms of HTT and investigate its relationships with PTC.
Methods
Study samples
Snap-frozen tissue and formalin-fixed paraffin-embedded (FFPE) tissue samples collected at the Department of Pathology at the University of Pittsburgh Medical Center (UPMC) were retrospectively studied following approval by the University of Pittsburgh Institutional Review Board (IRB). Additional fresh-frozen tissue samples from HTT and corresponding normal thyroid were collected at the MD Anderson Cancer Center with IRB approval. Finally, retrospective analysis of genomic data from 10,165 consecutive FNA samples from thyroid nodules with indeterminate cytology routinely tested by ThyroSeq v3 Genomic Classifier (GC) was performed. Clinical and surgical follow-up was collected on 14 resected HTT and eight FNA samples positive for GLIS fusions.
Pathology review
Glass slides from tumors initially diagnosed as HTT at the Department of Pathology at the UPMC were reviewed by three thyroid pathologists (R.R.S., S.I.C., and Y.E.N.) blinded for the results of molecular analyses. All tumors were classified according to the 2017 WHO classification of endocrine tumors (8). In case of discrepancy of diagnostic opinion, cases were discussed again and a consensus diagnosis was reached.
Whole-exome sequencing
The library construction was performed using the SeqCap EZ System (NimbleGen, Madison, WI) according to the manufacturer's instructions. Briefly, genomic DNA was sheared, the size was selected to roughly 300 bp, and the ends were repaired and ligated to specific adapters and multiplexing indexes. Fragments were incubated with SeqCap biotinylated DNA baits after ligation-mediated PCR, and the hybrids were purified using streptavidin-coated magnetic beads. After amplification of ≤18 PCR cycles, the libraries were sequenced on the HiSeq 3000 (Illumina, San Diego, CA), using 100 bp pair-ended reads.
Bioinformatics analysis was performed using a custom protocol developed for tumor and paired normal and tumor-only analysis (21). Briefly, after conversion of base call files to FASTQ files using bcltofastq (Illumina), sequences were aligned to the human reference genome (GRCh37.p13; GCF_000001405.25) using BWA MEM (22) and encoded into a BAM (binary sequence alignment) format using Samtools (23). Post alignment, the BAM files were sorted, indexed, de-duplicated, and subjected to local indel realignment and base quality score recalibration using GATK (24). Subsequently, variant calling was performed using Varscan2 (25) for single nucleotide variants (SNV) and short indel detection and Scalpel (26) for large indel detection. Variant calls from both callers were integrated, normalized, annotated (27,28), and prioritized based on reported minor allele frequency in population databases (29,30), variant location (coding vs. non-coding), in silico prediction algorithms (31,32), and public somatic and germline variant databases (33 –35). Somatic mutation density analysis was performed for samples with tumor and paired normal and included all non-synonymous SNV and small indels in the coding regions of the genes. Copy number and loss of heterozygosity analysis was performed using FACETS (36) and CNVkit (37) algorithms.
RNA-Seq analysis
For RNA-Seq (transcriptome) analysis, libraries were prepared using the Illumina TruSeq™ RNA Exome Sample Preparation Kit v1, according to the manufacturer's protocol. Cluster generation and paired-end sequencing were performed on an Illumina HiSeq2500 using a HiSeq Paired-End (PE) Rapid Cluster Kit v2 and HiSeq Rapid SBS Lit v2 (Illumina).
For detection of gene fusions, the sequencing reads were collected, and a quality filtering was applied based on Q-score (38). The Chimerascan algorithm (39), which utilizes Tophat aligner (40), was used. The fusions detected with Chimerascan were filtered using custom filtering algorithm that utilizes the distance between breakpoints and various cancer databases, such as Atlas (41), CIS (RTCGD) (42), and CHCG (43). The junctions and the individual exon expression levels were used to visualize the fusion point.
For gene expression analysis, the filtered high-quality reads from HiSeq sequencing system were aligned to human genome (hg19-GRCh37) using TopHat aligner (40), and the number of reads mapped to each gene was calculated using RSeM (44) and featureCounts (45) tools. From the read counts, the differential expression analysis was performed using the edgeR package (46). The genes with a fold change >2 with a p-value of <0.05 were selected as differentially expressed genes and used in pathway analysis using various databases such as KEGG (47), Reactome (48), and DAVID (49). The GO biological process analysis was conducted using DAVID (49). The fold changes in GLIS3 and GLIS1 expression between GLIS-rearranged HTT and normal thyroid tissue were also analyzed using transcript counts (TPM) with the fold-change detection pipeline (50).
Targeted NGS
Targeted NGS analysis was performed to genotype surgically removed tissue samples and FNA samples from thyroid nodules with indeterminate cytology using the ThyroSeq v3 GC assay, as previously described (51). The assay uses targeted amplification‐based NGS technology to detect genomic alterations in 112 thyroid-related genes, including PAX8–GLIS3 fusion by sequencing DNA and RNA on the Ion GeneStudio S5 System (Thermo Fisher Scientific, Waltham, MA) according to the manufacturer's protocol. Torrent Suite software v5.2.2 (Thermo Fisher Scientific) and software developed in house (Variant Explorer v2) were used for data analysis and interpretation (51).
RT-PCR and Sanger sequencing analysis
RNA from frozen and FFPE tissues was reverse transcribed with the High Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA) and by SuperScript IV VILO Master Mix (Invitrogen, Carlsbad, CA), respectively. RT-PCRs were conducted by HotStarTaq DNA Polymerase (Qiagen, Valencia, CA), using the primers reported in Supplementary Table S1. The RT-PCR products were sequenced in both directions using the BigDye Terminator Kit and an ABI 3130xl DNA Sequencer.
Fluorescence in situ hybridization
Following deparaffinization and rehydration, 4 μm FFPE sections were used to hybridize with commercially available probes from Empire Genomics LLC for PAX8 (green), GLIS3 (orange), and GLIS1 (orange) to detect PAX8–GLIS3 or PAX8–GLIS1 fusions. An additional PAX8 probe (green) was generated from BAC clone RP11-155L11 (Empire Genomics LLC, Buffalo, NY). Microscopy was performed using a Leica SP5 TCS 4D confocal laser scanning fluorescence microscope (Leica Camera, Wetzlar, Germany) with digital image capture.
Immunohistochemical analysis
After deparaffinization and rehydration, 4 μm FFPE sections were incubated with primary rabbit antibodies (anti-GLIS3, 1:50, NBP2-33787, Novus Biologicals, Centennial, CO; anti-pan-Collagen, 1:100, PA1-85324, Thermo Fisher Scientific; and anti-Collagen IV, 1:100, PA1-28534, Dako) for 14 hours at 4°C. Following the washes, secondary HRP-conjugated anti-rabbit antibodies (1:100; P0448; Dako, Agilent, Santa Clara, CA) were applied. Visualization was performed with DAB Peroxidase Substrate Kit (SK-4100; Vector Laboratories, Burlingame, CA).
Results
Identification of GLIS3 fusion in HTT using NGS
To search for driver genetic alterations in HTT, first whole-exome sequencing (WES) of DNA from three snap-frozen HTT samples and two corresponding normal tissues was performed. The analysis revealed no driver point mutations or indels known for thyroid or other cancer types and no potential novel driver mutations (Supplementary Table S2). Overall, the analysis showed a low somatic mutation density (M = 0.039 non-synonymous mutations/Mb) in the two tumors with corresponding normal tissue available. In addition, the WES analysis demonstrated absence of significant somatic DNA copy number alterations in these tumors (Supplementary Fig. S1).
Next, RNA-Seq (transcriptome) analysis of the three snap-frozen HTT samples and one corresponding normal tissue sample was performed. It detected an in-frame fusion between exon 2 of the PAX8 gene and exon 3 of the GLIS3 gene in all three HTT samples, while no fusion was identified in corresponding normal thyroid tissue (Supplementary Table S3). The fusion is expected to be a result of an inter-chromosomal rearrangement between the PAX8 gene located on chromosome 2q14.1 and GLIS3 on chromosome 9p24.2 (Fig. 1A). The fusions were confirmed in all HTT samples by RT-PCR and Sanger sequencing (Fig. 1 B). RT-PCR also showed the presence of alternative transcripts between PAX8 exon 2 and GLIS3 exon 4, which were not, however, in-frame. In addition to the PAX8–GLIS3 fusion, two HTT samples revealed the presence of several intra-chromosomal fusions involving the 2q11.2–2q14.1 chromosomal region adjacent to the location of PAX8 (Supplementary Table S3), providing evidence for multiple chromosomal breaks occurring in this region on 2q.
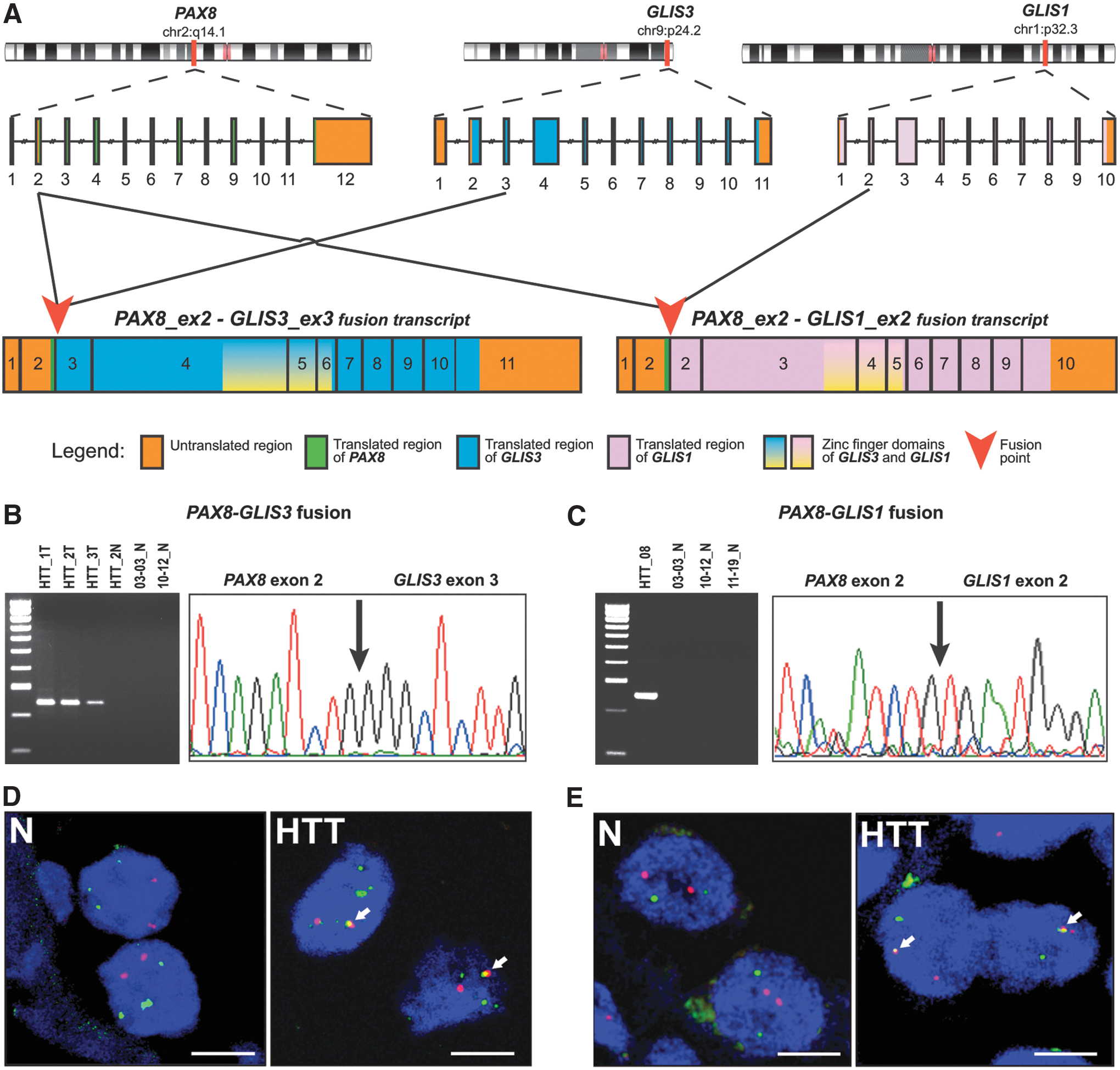
Structure and confirmation of PAX8–GLIS3 and PAX8–GLIS1 fusions identified by RNA-Seq analysis in hyalinizing trabecular tumor (HTT) samples. (
Analysis of an expanded cohort of HTT cases
To determine the prevalence of PAX8–GLIS3 fusion in these tumors, 14 additional samples initially diagnosed as HTT were studied (Table 1). All samples were analyzed by RT-PCR, Sanger sequencing, and a 112-gene targeted NGS panel (ThyroSeq v3 GC) that detects PAX8–GLIS3 and other gene fusions in addition to mutations and copy number alterations. Seven of those samples that had sufficient RNA quantities were studied by RNA-Seq. Overall, 10/14 samples were found to be positive for the same fusion between exon 2 of the PAX8 gene and exon 3 of GLIS3. No BRAF, RAS, or RET/PTC mutations or other driver fusions were identified in these samples. However, out of four PAX8–GLIS3 fusion-negative cases, one was positive for the HRAS p.Q61R mutation, two were positive for multiple copy number alterations, and one sample was negative for all genomic alterations by targeted NGS analysis but revealed a PAX8–GLIS1 fusion on RNA-Seq (Fig. 2). In the latter case, the fusion point of the chimeric transcript was between exon 2 of the PAX8 gene and exon 2 of GLIS1 (Fig. 1A). PAX8 is located on chromosome 2q14.1 and GLIS1 on chromosome 1p32.3, indicating that the fusion is a result of an inter-chromosomal rearrangement. The fusion was confirmed by RT-PCR and Sanger sequencing (Fig. 1C). Analysis of eight normal thyroid tissues available for these cases by targeted NGS revealed no GLIS fusions or other mutations, indicating that all identified molecular alterations are somatic events (Fig. 2).
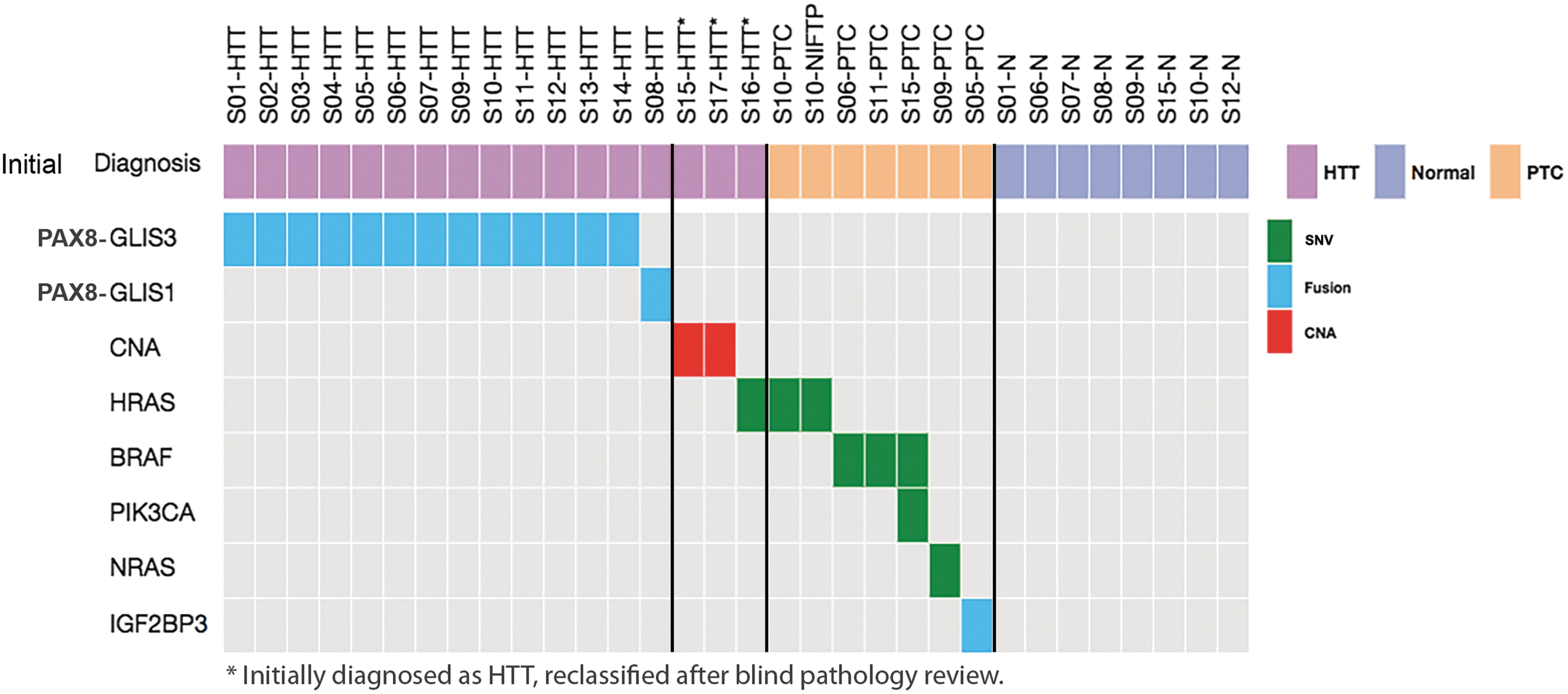
Summary of molecular alterations detected among 17 tumors initially diagnosed as HTT and in co-existing other tumors and normal thyroid tissue. PTC, papillary thyroid carcinoma; CNA, copy number alterations.
Characteristics of Tumors Initially Diagnosed as HTT
F, female; M, male; HTT, hyalinizing trabecular tumor; PTC, papillary thyroid carcinoma; CNA, copy number alterations.
Next, to confirm the fusions at the chromosomal level, fluorescence in situ hybridization (FISH) was performed with DNA probes corresponding to the PAX8 and GLIS3 and PAX8 and GLIS1 chromosomal regions. The hybridized tumor cell nuclei demonstrated one pair of fused signals in the majority of cells, confirming the occurrence of inter-chromosomal rearrangements leading to PAX8–GLIS3 and PAX8–GLIS1 fusions (Fig. 1D and E).
Histopathological review of tumors
Histological slides from all 17 cases with the initial diagnosis of HTT were reviewed by three thyroid pathologists blinded to the results of molecular analysis. The histologic diagnosis of HTT was confirmed in 14 cases. All of these tumors were positive for a GLIS rearrangement, including 13 (93%) PAX8–GLIS3 fusions and one (7%) PAX8–GLIS1 fusion (Table 1). The tumors were well circumscribed and had a thin or moderately thick fibrous capsule as shown in Figure 3A (For digital images of all tumors, see
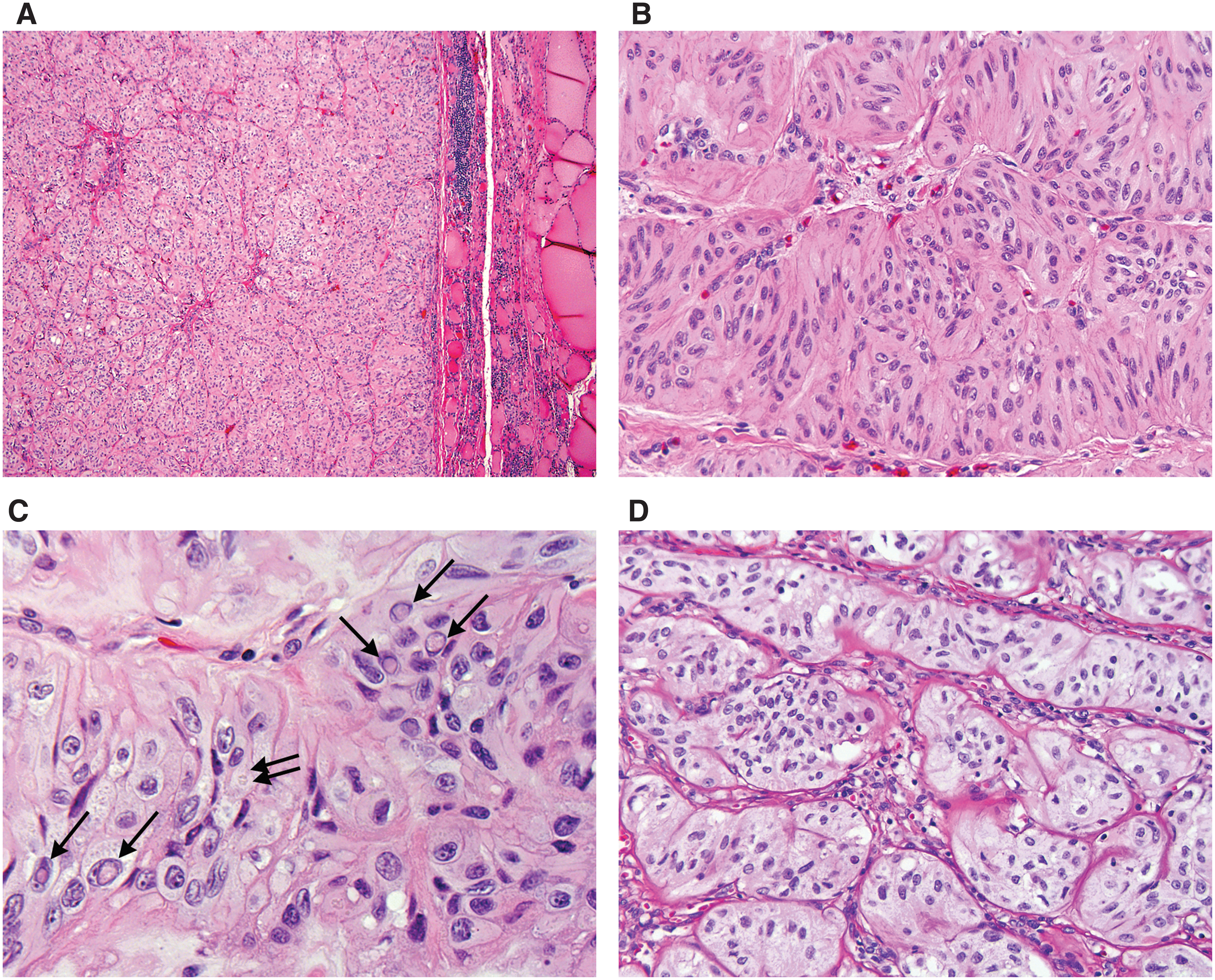
Histopathologic features of HTT carrying PAX8–GLIS3 (
In three tumors with the initial diagnosis of HTT, this diagnosis was not confirmed on blind review. These tumors had a smooth, noninvasive border with variably thick capsules and a vague trabecular architecture with some stromal hyalinization, which prompted the initial diagnosis of HTT, but lacked a pronounced trabecular architecture, intra-trabecular hyalinization, and abundance of nuclear pseudoinclusions expected in HTT (Fig. 4; see
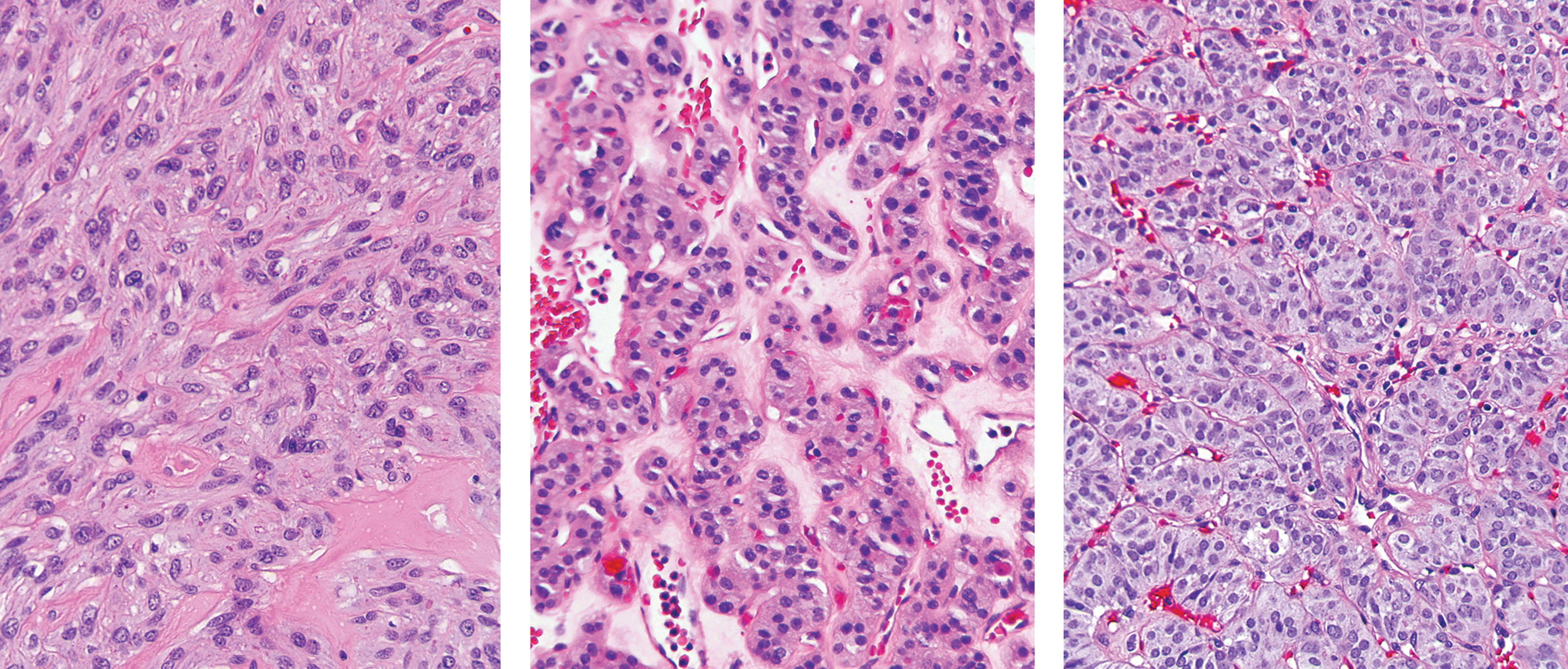
Histopathologic features of GLIS fusion-negative tumors initially diagnosed as HTT and reclassified on blind review by a panel of thyroid pathologists. Left: This tumor (S15-HTT) composed of mostly spindle cells arranged in solid sheets with focal stromal hyalinization was reclassified as follicular adenoma with spindle cells. Center: This tumor (S16-HTT) with predominantly microfollicular architecture, cells with abundant granular cytoplasm, and stromal hyalinization was reclassified as Hürthle cell adenoma. Right: This tumor (S17-HTT) with solid and trabecular architecture and no hyalinization but moderately developed nuclear features of PTC was reclassified as solid variant of PTC. See full digital images of representative tumor sections at
As a result, among tumors possessing all classical microscopic features of HTT, all 14 had GLIS fusions.
Functional consequences of GLIS rearrangements
The identified rearrangements involve fusions of PAX8 with either the GLIS3 or GLIS1 genes. PAX8 is a paired box transcription factor, which is highly expressed in differentiated thyroid follicular cells and required for normal thyroid development and function (52). GLI-similar 1 and 3 (GLIS1 and GLIS3) belong to a family of the GLI-similar zinc finger transcription factors that can act either as activators or repressors of gene transcription, and in thyroid, GLIS3 is known to be an important regulator of thyroid hormone biosynthesis (53,54). GLIS1 and GLIS3 share a highly homologous DNA-binding domain that consists of five zinc finger motifs (55). The DNA-binding domain is coded by exons 4–6 of GLIS3 and exons 3–5 of GLIS1. The PAX8–GLIS3 and PAX8–GLIS1 fusions juxtapose exon 3 of GLIS3 or exon 2 of GLIS1 in-frame and downstream of exon 2 of PAX8. As a result, the chimeric transcripts should be regulated by the PAX8 gene promoter and preserve the zinc-finger containing DNA-binding domains of both GLIS genes.
The predicted structure of PAX8–GLIS3 and PAX8–GLIS1 transcripts was confirmed by RNA-Seq data. They showed high levels of expression of exons 3–10 of GLIS3 in the chimeric PAX8–GLIS3 transcript in tumors compared to the normal thyroid tissue (Fig. 5A). PAX8 had a similar level of expression in the tumor and normal samples. Overall, compared to normal thyroid tissues (n = 16), the PAX8–GLIS3-positive HTT (n = 7) showed a 6.8-fold increase (range 3.6- to 58.4-fold) in GLIS3 mRNA expression (Fig. 5B). Similarly, a tumor carrying a PAX8–GLIS1 fusion showed a 20.2-fold increase in GLIS1 mRNA compared to normal thyroid tissues. Further, overexpression of the GLIS3 protein was demonstrated in HTT tissue sections by immunohistochemistry with an antibody to the C-terminus region of human GLIS3, with immunoreactivity observed both in the tumor cell nuclei and the cytoplasm (Fig. 5C).
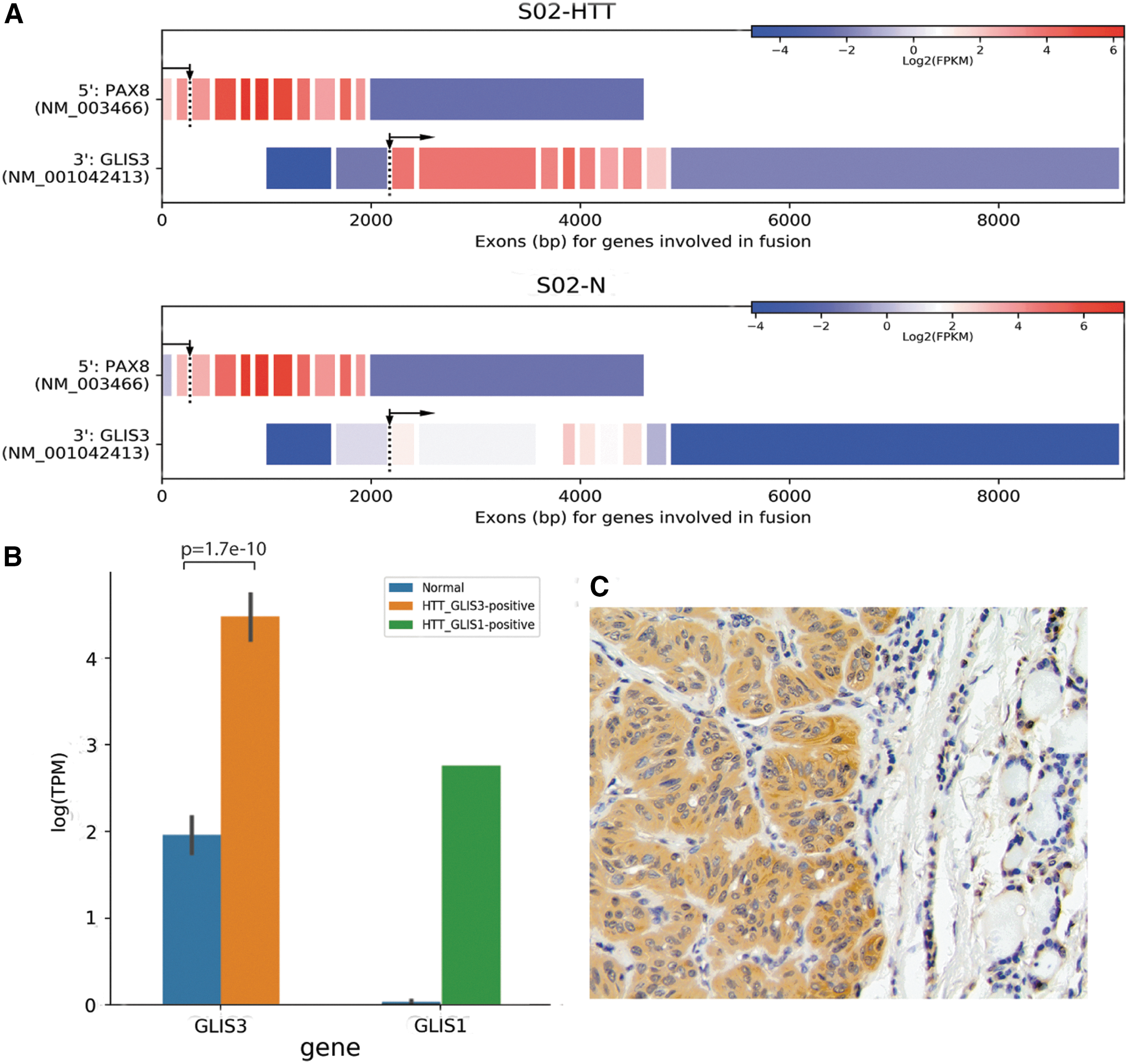
Consequences of PAX8–GLIS fusions on expression of GLIS3 and GLIS1. (
To obtain further insights into the impact of PAX8–GLIS3 fusion on thyroid cell function, we used the RNA-Seq gene expression data from GLIS-fused HTTs and normal thyroid tissues to perform pathway analysis using several pathway enrichment programs (KEGG, Reactome, and Gene Ontology (GO) biological process analysis; Supplementary Table S4A–CS). The analysis revealed dysregulation of multiple pathways, with extracellular matrix–related (ECM-related) and thyroid hormone biosynthesis and ion transmembrane transport (TH-related) pathways being among the top pathways dysregulated in tumors with PAX8–GLIS3 as compared to normal thyroid tissue (Fig. 6A and Supplementary Table S5). A number of extracellular matrix-related genes, including the majority of collagen genes, such as COL4A1, COL5A3, COL5A2, COL15A1, and COL1A2, were expressed at significantly higher levels in HTT as compared to normal thyroid tissue (Supplementary Table S5 and Supplementary Fig. S2). Overexpression and deposition of collagens in the areas corresponding to intra-trabecular hyalinization seen in HTT was confirmed by immunohistochemistry with collagen IV and a pan-collagen antibodies (Fig. 6B and C).
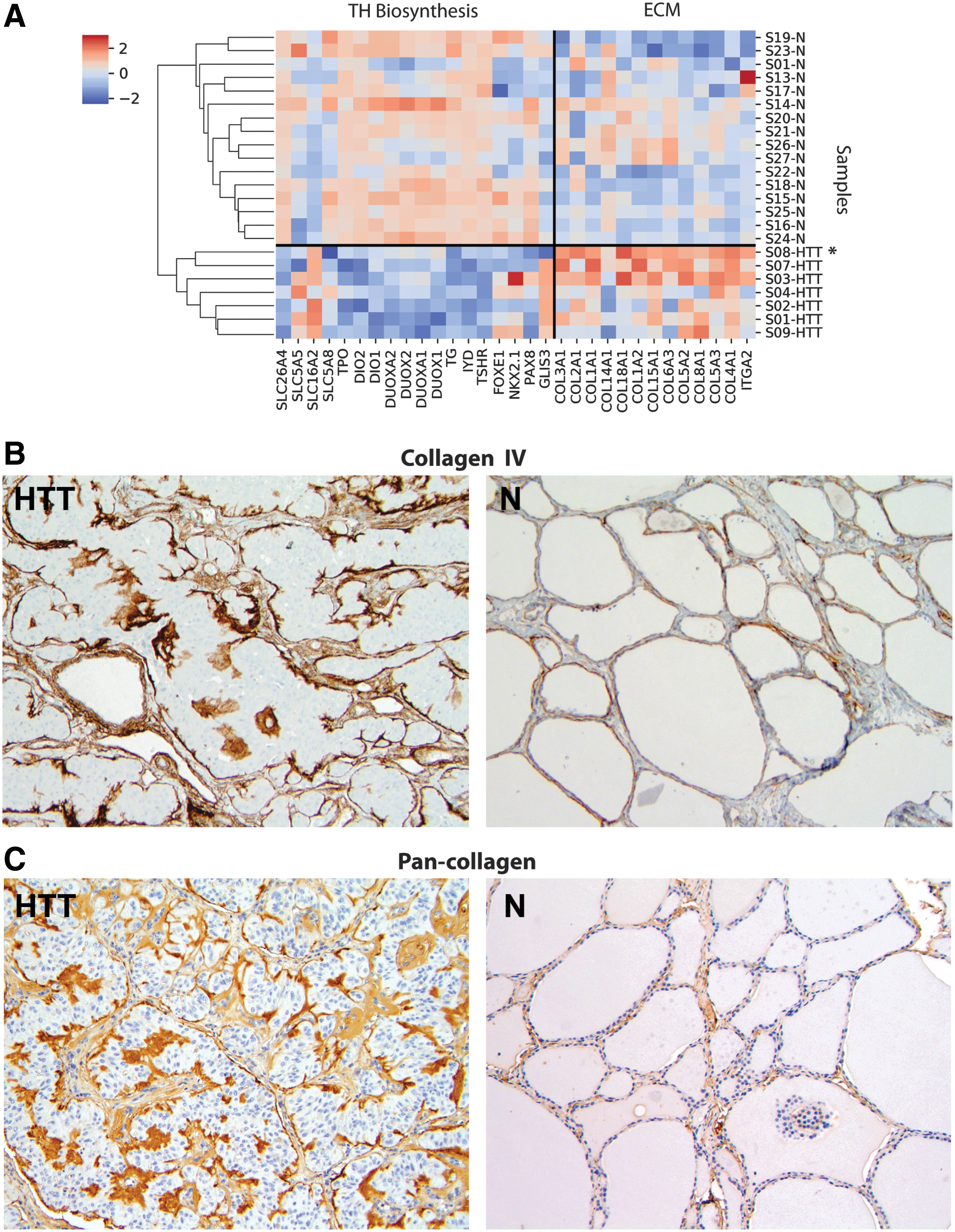
Dysregulation of extracellular matrix (ECM)-related and thyroid hormone biosynthesis and ion transmembrane transport-related pathways in GLIS-rearranged HTT. (
Among genes related to thyroid hormone biosynthesis, SLC16A2 (MCT8) and SLC5A5 (NIS) were significantly upregulated and TSHR, TPO, and deiodinase genes (DIO1, DIO2) were downregulated (Supplementary Table S5 and Supplementary Fig. S3).
Molecular profiling of synchronous PTC
HTTs are known to co-occur in thyroid glands that, not infrequently, also have PTC and other nodular lesions (20). Indeed, in the series of HTT cases, six coexisting PTC and one noninvasive follicular thyroid neoplasm with papillary-like nuclear features (NIFTP) present in the same or opposite thyroid lobe were identified. These tumors were genotyped for the known thyroid tumor alterations using a 112-gene targeted NGS panel (51). All synchronous PTC and a NIFTP case were negative for GLIS fusions. However, they were all positive for other alterations, including the BRAFV600E mutation in two PTC, a BRAFV600E and PIK3CA in one PTC, an NRASQ61R mutation in one PTC, a THADA–IGF2BP3 fusion in one PTC, and an identical HRASG13R mutation in both a PTC and a NIFTP coexisting in the same thyroid gland (Fig. 2).
Prevalence of GLIS fusions in PTC and thyroid nodules
In order to evaluate the prevalence of PAX8–GLIS3 and PAX8–GLIS1 fusions in PTC, 220 samples of PTC by RT-PCR were studied. This set of tumors included 111 freshly frozen samples of unselected PTC and 109 FFPE samples of preselected aggressive PTC. In addition, The Cancer Genome Atlas (TCGA) data on 484 PTC tested for fusions using RNA-Seq were reviewed (56). No PAX8–GLIS3 fusion was found among the 220 tumor samples (Supplementary Figs. S5 and S6) and in the TCGA study (Table 2). However, a PAX8–GLIS1 fusion was identified in 1/111 tumors from the unselected PTC cohort (Supplementary Figs. S4 and S5). No PAX8–GLIS1 fusion was found among 109 samples from the aggressive PTC cohort or 484 PTC in TCGA study. Histologic slides from the tumor positive for PAX8–GLIS1 were reviewed and revealed an encapsulated tumor with predominantly solid and microfollicular architecture and only a focal trabecular pattern composed of cells with moderately developed nuclear features of PTC but highly abundant nuclear pseudoinclusions seen in focal areas (Supplementary Fig. S4). Despite a somewhat unusual appearance, the diagnosis of PTC was confirmed. Interestingly, the microscopic appearance of the tumor, as well as patient demographics, were identical between this case and a thyroid tumor positive for PAX8–GLIS1 fusion in the expanded TCGA PanCancer Atlas study (tumor case # TCGA-BJ-A291, contributed from the University of Pittsburgh Medical Center), suggesting that it may represent the same tumor.
Prevalence of PAX8–GLIS3 and PAX8–GLIS1 Fusions in the Current Study and TCGA Cohort
As reported by TCGA Research Network (56).
The same case was reported as PAX8/GLIS1 positive in TCGA PanCancer Atlas study (see
TCGA, The Cancer Genome Atlas.
Finally, taking advantage of the fact that PAX8–GLIS3 fusion is detectable by the ThyroSeq v3 GC panel, the prevalence of this fusion was evaluated in 10,165 consecutive FNA samples from thyroid nodules with indeterminate cytology. PAX8–GLIS3 was detected in eight (∼0.1%) of the nodules. Surgical follow-up was available for five of these patients, and all five nodules carrying PAX8–GLIS3 were pathologically diagnosed as HTT after excision.
Clinical features and follow-up for patients with HTT
All 14 patients with HTT were female, with a mean age of 57 years (range 31–75 years). The mean size of resected HTT was 1.6 cm (range 0.6–3.5 cm). Preoperative ultrasound was performed for six patients; five (83%) HTT nodules were solid/hypoechoic, and three (50%) were hypervascular. FNA cytology was reported as positive for malignancy (Bethesda VI) in one case, suspicious for malignancy (Bethesda V) in two cases, follicular neoplasm/suspicious for follicular neoplasm (Bethesda IV) in two cases, and atypia of undetermined significance/follicular lesion of undetermined significance (Bethesda III) in one case. Clinical follow-up was available for 9/14 patients with GLIS fusion-positive HTT. There were no tumor recurrences after a mean follow-up of 65 months (range 4–181 months). Radioactive iodine was administered to two patients and recommended to one patient for concurrent PTC.
Discussion
This study uncovered the underlying genetic mechanism of the thyroid neoplasm known as HTT by demonstrating that gene fusions involving the PAX8 and GLIS genes represent a genetic hallmark of these tumors. In fact, these fusions, typically PAX8–GLIS3, were identified in 100% of tumors that had fully developed histological features of HTT.
Further, the study shows that these tumors have a quiet genome with a low number of somatic mutations and no significant DNA copy number alterations. Furthermore, these tumors have no mutations characteristic of PTC and all of its variants, such as BRAF and RAS point mutations and RET/PTC and other gene fusions. Reciprocally, PTC were found to carry no PAX8–GLIS3 fusions in a large and well annotated cohort of tumors in this study (n = 220) and in TCGA tumor cohort (n = 484) (56). Similarly, none of the PTC co-occurring in the same thyroid gland with HTT had a GLIS fusion, but instead they carried BRAFV600E and other alterations characteristic of PTC. However, a PAX8–GLIS1 fusion, which was detected in one HTT, was also found in a tumor that had some microscopic features of HTT but carried a consensus diagnosis of PTC. Overall, the contrasting molecular profiles of HTT and PTC, including tumors arising in the same thyroid gland, argue strongly against biological similarity between these two tumor types.
This study demonstrates that fusions of GLIS3 or GLIS1 genes to PAX8, formed as a result of inter-chromosomal rearrangements, lead to the generation of chimeric transcripts and strong overexpression of the 3′ portions of the GLIS genes with the intact DNA-binding zinc finger domains of these transcription factors. Further, the study found that the PAX8–GLIS3 fusion-positive tumors show consistent upregulation of expression of extracellular matrix–related genes, including several collagen genes such as COL4A1, coding for the alpha-1 subunit of collagen IV found primarily in the basal lamina. Excessive production and deposition of collagen IV and other collagens in these tumors was confirmed by immunohistochemistry. This is likely responsible for producing extensive hyalinization, which results in the characteristic microscopic appearance and name of these tumors. How exactly GLIS regulates the expression of extracellular matrix–related genes remains to be clarified, but such an effect of GLIS is not entirely unexpected. Indeed, GLIS3 knockout in thyroid cells of mice has been previously shown to lead to significant downregulation of the extracellular matrix genes (54). Abnormal intracellular and extracellular deposition of basement membrane material was demonstrated in these tumors before by electron microscopy and immunohistochemistry using a collagen IV antibody (57,58), although the mechanisms responsible for this phenomenon were not known.
Dysregulation of the thyroid hormone biosynthesis pathway in HTT carrying GLIS fusions was also observed, which is also expected based on the known role of GLIS3 in regulating thyroid cell development and function (54). However, specific genes belonging to this pathway were variably affected, as some of them (SLC16A2/MCT8, SLC5A5/NIS) were upregulated, whereas many other genes coding for proteins involved in thyroid hormone synthesis either were downregulated or had levels of expression similar to normal thyroid tissue.
Whereas the role of GLIS3 in thyroid cell function is well established, little is known about the physiological function of GLIS1 in thyroid cells. GLIS1 and GLIS3 belong to a subfamily of Krüppel-like zinc finger transcription factors that regulate transcription of a variety of genes in physiological and pathological conditions (59). In the thyroid, GLIS3 has been shown to be an important regulator of thyroid hormone synthesis, with loss of GLIS3 function by mutation associated with the development of congenital or neonatal hypothyroidism (53,60). Similarly, strong downregulation of the expression of SLC5A5/NIS and other iodide transporters was observed in thyroid glands of GLIS3-deficient mice (54). The known high homology (∼94%) between the zinc finger DNA-binding domain of GLIS3 and GLIS1 (55) and findings in this study of similar effects of overexpression of these two genes through gene fusion on dysregulation of signaling pathways including thyroid hormone biosynthesis raise the question whether GLIS1 is also involved in regulating certain aspects of thyroid cell function.
In this series of patients and among 118 tumors reported in the largest study to date (20), tumors with histopathologic features of HTT that lack invasion did not have regional or distant metastases at presentation, and patients experienced no tumor recurrence or any other adverse effects on follow-up. Furthermore, no GLIS fusions were found in a series of 109 aggressive PTC, including those with distant metastases or extensive local invasion. This suggests that in the absence of invasion, GLIS fusion-positive thyroid neoplasms currently known as HTT are benign, as initially suggested by Carney et al. (1). Therefore, these tumors should be considered adenomas and classified as “GLIS-rearranged hyalinizing trabecular adenoma.”
The change of the tumor name from more ambiguous “tumor” to “adenoma” should reflect more accurately the benign nature of this disease, but it does not exclude the existence of a malignant counterpart of this tumor. In fact, it would closely recapitulate the deeply engraved nomenclature approach in medical practice to thyroid adenoma/thyroid carcinoma tumors that have similar overall morphology but are separated based on the absence or presence of invasion. Single cases of invasive tumors with trabecular growth and hyalinization have been reported, and some manifested with distant metastases (4,5,20). Such tumors can be designated as “hyalinizing trabecular carcinoma,” as proposed before (5). However, it remains unclear whether these invasive tumors carry GLIS fusions, and at this point in time, both possibilities should be considered. If invasive tumors are found to carry GLIS fusions, it would support the designation of these tumors as “GLIS-rearranged hyalinizing trabecular carcinoma” and provide strong evidence for a biological similarity between these tumors and existence of progression of these tumors with characteristic hyalinization and trabecular growth from adenoma to carcinoma. Alternatively, if invasive tumors with similar histology are found to be GLIS-negative but instead carry genetic alterations in classical PTC genes, this would indicate that they represent a peculiar type of PTC and suggest that GLIS-positive tumors are likely nonprogressive benign neoplasms.
There is also a possibility that tumors carrying GLIS3 and GLIS1 fusions have a different predisposition to malignant transformation. Indeed, current data suggest that progression of PAX8–GLIS3-positive tumors would be a very rare event, as these fusions were not identified in >700 carefully curated PTC in TCGA cohort (56) and this study. However, a single case of PAX8–GLIS3 has been reported in a 1 cm classical PTC that was intrathyroidal and had no invasion (61). The reported mRNA fusion point between exon 2 of PAX8 and exon 4 of GLIS3 appears to be different to the one identified in tumors in this study and apparently resulted in an out-of-frame fusion. Furthermore, the histological appearance of the tumor was not illustrated, precluding evaluation for the microscopic features of HTT. The probability of malignant transformation may be higher for tumors with PAX8–GLIS1 fusion, which was found with low prevalence in HTT, but also one tumor classified as PTC. This tumor had some microscopic features of HTT, particularly a very high number on nuclear pseudoinclusions, but overall had the histological appearance of PTC.
Finally, this study demonstrates that GLIS fusions can be detected preoperatively in thyroid FNA samples, and among nodules with indeterminate cytology, primarily Bethesda III and IV, PAX8–GLIS3 was found in ∼0.1% of cases. The prevalence of these fusions in Bethesda V and IV nodules may be even higher, as HTT in more than half of the cases yield a cytological diagnosis of PTC or suspicious for PTC (19,62,63). In fact, due to well expressed nuclear features of PTC, HTT commonly pose a diagnostic difficulty for thyroid cytology and lead to false-positive cytology diagnoses triggering total thyroidectomy (62).
Interestingly, among nodules with a prospectively detected PAX8–GLIS3 fusion that underwent surgery, 5/5 were diagnosed as HTT on surgical pathology. While the number of cases with prospectively accumulated follow-up is low and further studies are required to reliably establish the probability of malignancy in nodules carrying PAX8–GLIS3 and PAX8–GLIS1 fusions, the results of this study suggest that thyroid lobectomy is likely to be an appropriate initial surgical approach for such nodules. Future studies are required to determine if clinically significant cancers carrying these fusions exist, which will help to determine if surgical treatment is needed at all for thyroid nodules carrying PAX8–GLIS3 or any type of GLIS fusion.
Footnotes
Acknowledgments
We thank Mrs. Jessica Tebbets for her help in preparation of this manuscript and for collection of follow-up information. This work was supported in part by the NIH grants R01CA181150 and P50CA097190. Sample collection was supported in part by the UPCI Tissue and Research Pathology/Health Sciences Tissue Bank shared resource which is supported in part by award P30CA047904 and by a generous gift from William and Susan Johnson.
Author Disclosure Statement
M.N.N. and Y.E.N. receive compensation from their employer in connection with ThyroSeq test offered through CBLPath/Sonic Healthcare USA; they own intellectual property related to the ThyroSeq test. The remaining authors have nothing to disclose.