
Abstract
Background:
Mutations in the thyroid hormone (TH) transporter monocarboxylate transporter 8 (MCT8) lead to peripheral hyperthyroidism and profound psychomotor alterations in humans. Mice lacking Mct8 present peripheral hyperthyroidism but no gross neurological abnormalities due to brain compensatory mechanisms involving the enzyme deiodinase type 2 (Dio2).
Methods:
Here we have analyzed the endocrine and neurologic phenotype of mice lacking both Mct8 and Dio2 at three and six months of age. Thyroxine (T4) and 3,5,3′ triiodothyronine (T3) levels/content were measured by specific radioimmunoassays; motor skill performance was evaluated by the footprint, rotarod, four limb hanging wire, and balance beam tests; and brain histological analysis was performed by immunostaining for neurofilament and parvalbumin.
Results:
We have found that this mouse model presents peripheral hyperthyroidism and brain hypothyroidism. Interestingly, the severity of the brain hypothyroidism seems permanent and varies across regions, with the striatum being a particularly affected area. We have also found brain alterations at the histological level compatible with TH deficiency and impaired motor skills.
Conclusions:
These findings indicate the potential of Mct8/Dio2-deficient mice to represent a model for human MCT8 deficiency, to understand the mechanisms underlying its pathophysiology, and ultimately design therapeutic interventions for human patients.
Introduction
Mutations in the monocarboxylate transporter 8 (MCT8, SLC16A2) gene, located on the X chromosome, are associated with the orphan and rare disease Allan–Herndon–Dudley syndrome (AHDS) in males (1 –3). The MCT8 is highly specific in transporting thyroid hormones (THs), thyroxine (T4), and its nuclear active form 3,5,3′triiodothyronine (T3). MCT8-deficient patients present global developmental delay, profound intellectual disability, lack of speech, and poor communication skills, as well as severe neuromotor impairments with central hypotonia, progressive spastic quadriplegia, and dystonic movements (4 –13). These patients also present thyroid function test abnormalities, consisting of high serum T3, low T4, and reverse T3, and normal-to-slightly elevated thyrotropin (TSH) (2,3). Studies from mice and patient-specific induced pluripotent stem cells indicate that an impaired TH transport across the brain barriers, including the blood–brain barrier (BBB), is an important pathophysiological mechanism in MCT8 deficiency (14 –16) leading to decreased T4 and T3 content in the brain (17 –19). Indeed, strong evidence of brain hypothyroidism has been found in the brain of MCT8-deficient subjects at prenatal and postnatal ages (19).
Mct8 knockout (Mct8KO) mice were generated as a model for the AHDS. These mice faithfully replicate the alterations in the circulating TH concentrations of patients, with high T3 and low T4 (17,18). Unfortunately, they do not present gross neurological abnormalities (20); hence they are only a partial model of the disease. Despite this, they have been a useful tool to understand some aspects of the syndrome and to explore the differences between mice and humans. The current theory that explains why Mct8KO mice do not present gross neurological abnormalities, unlike patients, supports that increased deiodinase type 2 (Dio2) activity in the brain of Mct8KO mice (17,18,21) enhances local T4 to T3 conversion as a compensatory mechanism. In Mct8KO mice, T3 but not T4 transport is impeded at the BBB (14), therefore, it was postulated that T4 crosses the BBB and/or the blood–cerebrospinal fluid barrier in the absence of Mct8 through a transporter not present in humans. The organic anion transporting polypeptide 1c1 (Oatp1c1) transporter was proposed as a candidate to mediate T4 transfer into the mouse brain, as it is predominantly localized in capillary endothelial cells and in choroid plexus structures (22 –24) and most importantly, OATP1C1 expression is weak at the BBB endothelial cells of primate (25) and human (26) brain.
This hypothesis is supported by the phenotype observed in double knockout (KO) animals lacking both Mct8 and Dio2 proteins or Mct8 and Oatp1c1 transporters, as both animal models presented similar characteristics to hypothyroidism with decreased T3 content and altered expression of T3-target genes in the brain (21,27). Further characterization of double Mct8/Oatp1c1KO mice revealed that these animals present impaired motor skills with abnormal gait, poor coordination, and reduced grip strength; disturbed cerebellar development; reduced myelination; and compromised GABAergic interneurons. Moreover, they also replicate the patient's thyroid function tests (27) so they have been considered the animal model most closely reflecting human MCT8-deficient patients.
Even though Mct8/Oatp1c1KO mice somehow reflect human MCT8 deficiency, there are some limitations within this model. For instance, Oatp1c1 also transports other compounds including steroid hormone metabolites such as the conjugated sterol β-estradiol-17-β-glucuronide (28), so the phenotypic outcome of these animals could also be due to alterations in the transport of other molecules. Furthermore, mutations in the OATP1C1 transporter in humans have been related to brain hypometabolism, brain-specific hypothyroidism, and neurodegeneration (29). For these reasons, having an additional model of the syndrome would contribute to the understanding of the mechanisms that underlie MCT8 deficiency more robustly and it would be very useful to compare the effects of deficient TH signaling in the brain due to different conditions.
We propose the double Mct8/Dio2KO mice as an alternative and additional model for the AHDS. The advantage of this model versus the Mct8/Oatp1c1KO animal is that, as compensation by Dio2 activity in the absence of Mct8 is avoided in the brain, the observed phenotype will be only related to TH actions. This study has been performed in young adult and adult mice because, even though it is now clear that MCT8-deficient patients present neurological alterations before birth (19), and hence patients would benefit from prenatal treatment to prevent brain damage, most patients are diagnosed within the first year after birth (there are >320 affected individuals belonging to 148 families reported to date). In addition, many of the alterations found in hypothyroid rodents during the perinatal age are only transient and, therefore, the use of adult mice allows observing more persistent damage. Therefore, this study aims in the first instance to establish comparable conditions with existing patients to identify therapeutic targets, and to ultimately design therapeutic interventions that prevent or restore brain damage and simultaneously correct the thyrotoxic state in the periphery.
In this study we present an extensive analysis of Mct8/Dio2KO mice at three and six months of age. As previously reported, these mice replicate the peripheral hyperthyroidism characteristic of MCT8-deficient patients and presented a reduction in the cerebral T3 content (30), as has been found in the cerebral cortex of a 30th gestational week fetus with mutations in MCT8 (19). Moreover, these mice display persistent locomotor abnormalities and brain alterations at the histological level. Based on these findings we propose the Mct8/Dio2KO mouse as an alternative and additional model to understand human MCT8 pathophysiology and to test therapeutic strategies.
Materials and Methods
Ethics statement
All experimental procedures involving animals were performed following the European Union Council guidelines (directive 2010/63/UE) and Spanish regulations (R.D. 53/2013), and were approved by the ethics committee at Consejo Superior de Investigaciones Científicas (CSIC; approval number 162/17). All efforts were made to minimize suffering as indicated hereunder.
Animal models and experimental design
All mice were housed at the Instituto de Investigaciones Biomédicas “Alberto Sols” under temperature- and light-controlled conditions at 22°C ± 2°C on a 12:12 light–dark cycle with ad libitum access to food and water. Experiments were carried out in wild type (Wt), Mct8-deficient (Mct8KO), Dio2-deficient (Dio2KO), and double Mct8- and Dio2-deficient (Mct8/Dio2KO) male mice. Wt and Mct8KO mice were originally produced by Dumitrescu et al. (17) and Dio2KO mice were initially produced by Schneider et al. (31) and a colony was established at our animal facility in the same C57BL/6J genetic background. For the experiments, Wt and Mct8KO littermates were obtained by backcrossing Mct8 −/+ females with Mct8 +/y males, and Dio2KO and Mct8/Dio2KO male mice were obtained from Mct8 +/y Dio2 −/− males and Mct8 −/+ Dio2 −/− females. Of note, in this breeding strategy, Dio2 −/− progenitors present high T4 plasma levels that could potentially affect the Dio2KO and Mct8/Dio2KO progeny. The Mct8 and Dio2 genotypes were confirmed by PCR of tail DNA as described (21).
Studies were conducted in three- and six-month-old Wt, Mct8KO, Dio2KO, and double Mct8/Dio2KO mice. At three months of age, there were no differences in the weight of the different genotypes (Wt = 24.78 ± 1.787 g; Mct8KO = 23.38 ± 2.446 g; Dio2KO = 25.00 ± 3.742 g; Mct8/Dio2KO = 22.50 ± 3.749 g); however, at six months of age, Mct8/Dio2KO animals had lower body weight than Wt mice (Wt = 30.40 ± 2.011 g; Mct8KO = 28.63 ± 1.408 g; Dio2KO = 33.13 ± 2.416 g; Mct8/Dio2KO = 26.75 ± 1.581 g).
For motor task analysis, Wt (n = 10, from 5 L), Mct8KO (n = 8, from 5 L), Dio2KO (n = 8, from 4 L), and Mct8/Dio2KO (n = 10, from 4 L) mice at 3 months of age and Wt (n = 10, from 5 L), Mct8KO (n = 8, from 5 L), Dio2KO (n = 8, from 4 L), and Mct8/Dio2KO (n = 9, from 4 L) mice at six months of age performed the tests described in the “Motor Tasks Assessment” section.
For histological analyses of the brain, Wt (n = 4, from 3 L), Mct8KO (n = 4, from 3 L), Dio2KO (n = 4, from 3 L), and Mct8/Dio2KO (n = 4, from 3 L) mice at 3 months of age and Wt (n = 4, from 3 L), Mct8KO (n = 4, from 3 L), Dio2KO (n = 4, from 3 L), and Mct8/Dio2KO (n = 4, from 3 L) mice at six months of age were anesthetized with ketamine (75 μg/g body weight) and medetomidine hydrochloride (1 μg/g body weight) and transcardially perfused with 4% paraformaldehyde in 0.1 M phosphate buffer (PB). Brains were removed, postfixed overnight in the same fixative solution, cryoprotected in 30% sucrose, and cut into 25 μm free-floating sections in the coronal plane on a cryostat.
For hormonal determinations in plasma and tissues, Wt (n = 9 from 6 L), Mct8KO (n = 9 from 6 L), Dio2KO (n = 7 from 5 L), and Mct8/Dio2KO (n = 10 from 5 L) mice at three months of age, and Wt (n = 10 from 5 L), Mct8KO (n = 8 from 5 L), Dio2KO (n = 8 from 4 L), and Mct8/Dio2KO (n = 9 from 4 L) mice at six months of age were anesthetized with ketamine (75 μg/g body weight) and medetomidine hydrochloride (1 μg/g body weight) and transcardially perfused with saline to remove blood from tissues before their collection. Before perfusion, blood was extracted by retroorbital collection and used for the determination of T4 and T3 plasma concentrations. Tissues (cerebral cortex, striatum, and gastrocnemius muscle) were harvested.
For morphometric analysis of the thyroid gland, Wt (n = 4 from 1 L) and Mct8/Dio2KO (n = 4 from 2 L) mice at six months of age were perfused with 4% paraformaldehyde in 0.1 M PB. Thyroid glands attached to a section of trachea were postfixed overnight at 4°C in the same solution, washed in phosphate-buffered saline (PBS), and embedded in paraffin. Sections of thyroid glands (8 μm) were stained with hematoxylin and eosin.
Hormonal determinations in plasma and tissues
High specific activity 125I -T3 and 125I-T4 (3000 μCi/μg) were labeled with 125I (NEZ033A; Perkin Elmer) using (3 –5)-T2 (D0629; Sigma) and T3 (T2877; Sigma) as substrates, respectively, as previously described (32,33). One minor modification was the separation of the labeled products, which was done by ascending paper chromatography for 16 hours in presence of butanol–ethanol–ammonia 0.5N (5:1:2) as solvent. The 125I -T3 and 125I-T4 were eluted and kept in ethanol at 4°C.
T3 and T4 were extracted from individual 80 μL aliquots of plasma with methanol (1:6), evaporated to dryness, and taken up in the radioimmunoassay buffer for determinations. T3 and T4 extraction from tissues (cerebral cortex, striatum, and gastrocnemius), as well as determinations of T3 and T4, was performed as previously described (34,35) with the dynamic range being 0.4–100 pg T3/tube and 2.5–320 pg T4/tube.
Motor tasks assessment
Footprint test
The gait of the mice's was evaluated using the footprint test as previously described (36). The hind and the fore paws were painted with red and black nontoxic waterproof paint, respectively, and the animals were required to run along a tunnel (20 × 20 × 70 cm) lined with paper, with a dark goal box at the end of the tunnel to encourage the mouse to run toward a dark and safe environment. Measurements for three-step cycles were averaged, considering a cycle as the distance from one pair of hind prints to the next. Footprints at the start and the end of the tunnel were excluded from the analysis as they correspond to the initiation and termination of the movement. The front base width, hind base width, forelimb stride length, and hind limb stride length were analyzed.
Rotarod test
Motor coordination and balance were evaluated with the accelerating-rotarod test as previously described (36). Sixty minutes before the test, animals were subjected to an initial training period under constant velocity. For the accelerated rotarod test, mice were placed on a rotarod (Ugo Basile, Italy) that accelerated from 4 to 44 rpm in three minutes and was maintained at 44 rpm for two further minutes. The latency of the mice to fall off the rod was recorded over the maximum observation period of five minutes. Mice were tested for five consecutive days with three trials per day with a 20 minutes intertrial interval. Data from three trials were averaged.
Four limb hanging wire test
Muscle strength was evaluated using the hanging wire test (37). Mice were placed on the top of a wire cage lid quadrant (14 × 14 cm) that was inverted horizontally 50 cm above a surface with bedding. The latency to fall was recorded over a maximum period of 120 seconds. Mice were tested for two consecutive days in one trial per day. Data from both days were averaged.
Balance beam test
Balance was evaluated as described (38). Mice were placed on the center of a horizontal wooden bar (0.9 cm wide × 50 cm long) 40 cm above a surface with bedding. The latency to fall was recorded over a maximum period of 40 seconds and two trials were performed in one single day. The activity on the bar was rated as [0] if the mouse remained in the center of the bar or if it moved but it did not reach the ends; and [1] if the mouse reached one of the ends of the bar.
Histological analyses
Immunohistochemical procedures were the same for antiparvalbumin (Pvalb, 1:1500, 28 P3088; Sigma-Aldrich) and antineurofilament 220KD (Nefh, 1:300, MAB5266; Millipore) antibodies. Free-floating sections were incubated with 3% hydrogen peroxide and 10% methanol in PBS at RT for 15 minutes to block endogenous peroxidase activity. Afterward, nonspecific antibody binding was prevented by blocking the tissue in PBS containing 0.1% triton X-100, 4% bovine serum albumin (BSA, A4503; Sigma), and 5% normal horse serum (S-2000; Vector Laboratories) at RT for one hour. Then, tissue sections were incubated with the primary antibodies at 4°C overnight in PBS containing 0.1% Triton X-100, 4% BSA, and 1% normal horse serum. The sections were subsequently washed in PBS, and incubated for one hour at RT with biotinylated secondary antibodies (Vector Laboratories) at a 1:200 dilution in PBS containing 0.1% Triton, 4% BSA, and 1% normal horse serum. The immune signal was amplified using the Avidin-Biotin Complex (Ultra-Sensitive ABC Peroxidase Staining Kit, 32050; Thermo Scientific) and developed with 0.5 mg/mL diaminobenzidine (D5637; Sigma) and 0.01% hydrogen peroxide. The sections were mounted on glass slides, dehydrated, cleared in xylene, and coverslipped with Depex (18243; Serva). To avoid methodological differences, brain tissue from Wt, Mct8KO, Dio2KO, and Mct8/Dio2KO animals of the same age was processed in parallel. Negative controls omitting the primary antibody were run in parallel and no staining was observed in control experiments with any of the antibodies used.
Quantification of the intensity of immunostaining for Pvalb and Nefh was performed in the motor and somatosensory cortex and, in the basal ganglia, in the caudate-putamen using four animals for each genotype at three months of age. For motor and somatosensory cortex, three representative sections were chosen per region, and three microphotographs were randomly captured in each section using a 40 × oil-immersion objective. For studies in the caudate-putamen, three representative sections were chosen, and four microphotographs were randomly captured in each section using a 20 × objective. All the images were captured with an Olympus DP70 digital camera joined to an Olympus BX51 microscope, using the CAST stereological software package (Visiopharm, Hørsholm, Denmark, and Olympus España). Images were collected in TIFF format and transformed to 8-bit gray using the image analysis software ImageJ (Wayne Rasband, National Institutes of Health). The median gray value of the pixels from the entire photograph was automatically assigned by ImageJ software. These data were corrected by the tissue background signal and transformed to percentage relative to mean value of the Wt animals (which is represented as 100%).
For morphometric analysis of the thyroid gland, four thyroid slices from each animal (four Wt and four Mct8/Dio2KO) were used to study the size of the thyroid follicles by NIH Image J software. In each slice, we quantified the area of 50 follicles.
Data analyses
Data are expressed as mean ± SE. Differences between means were obtained by one-way analysis of variance and the Bonferroni's post hoc test to correct for multiple comparisons or by the Student's t-test to compare values between two groups using the GraphPad software. Significant differences are represented as *p < 0.05, **p < 0.01, and ***p < 0.001.
In the balance beam test, differences in the activity on the bar between Wt and Mct8/Dio2KO mice were calculated by 2 × 2 contingency tables and the Fisher's exact test.
The rotarod test was analyzed by two general linear models (one for three-month-old and another for six-month-old animals) using “latency to fall” as the dependent variable and, as independent variables, genotype (Wt, Mct8KO, Dio2KO, and Mct8/Dio2KO) and day (first, second, third, fourth, and fifth day of test) with the SPSS statistics 19 package. For comparison of values between Wt and Mct8/Dio2KO animals, Student's t-test was used and significant differences are represented as # p < 0.05, ## p < 0.01, and ### p < 0.001.
Results
Mct8/Dio2KO mice present abnormal TH levels/content in plasma and tissues and larger thyroid follicles
Three- and six-month-old Mct8/Dio2KO mice present peripheral TH alterations, showing high T3 (3-fold at three months of age and 1.5-fold at six months of age) and low T4 (nearly 2-fold at three months of age and 1.5-fold at six months of age) plasma levels in comparison with Wt mice. All these data are consistent with previous findings in one- and three-month-old Mct8/Dio2KO mice (30,39), similarly to single Mct8KO animals and resembling the peripheral situation of MCT8-deficient patients (Fig. 1A). Indeed, three- and six-month-old Mct8KO mice present high T3 (2.7-fold) and low T4 (2-fold) plasma levels, as previously reported (17,18,40). Dio2KO mice displayed unaltered T3 and high T4 (1.3-fold at both ages) plasma levels, as already described (36,41). Although TSH levels were not assessed in this study, they have been found to be elevated in Mct8KO, Dio2KO, and Mct8/Dio2KO mice by others (17,30,31,40).
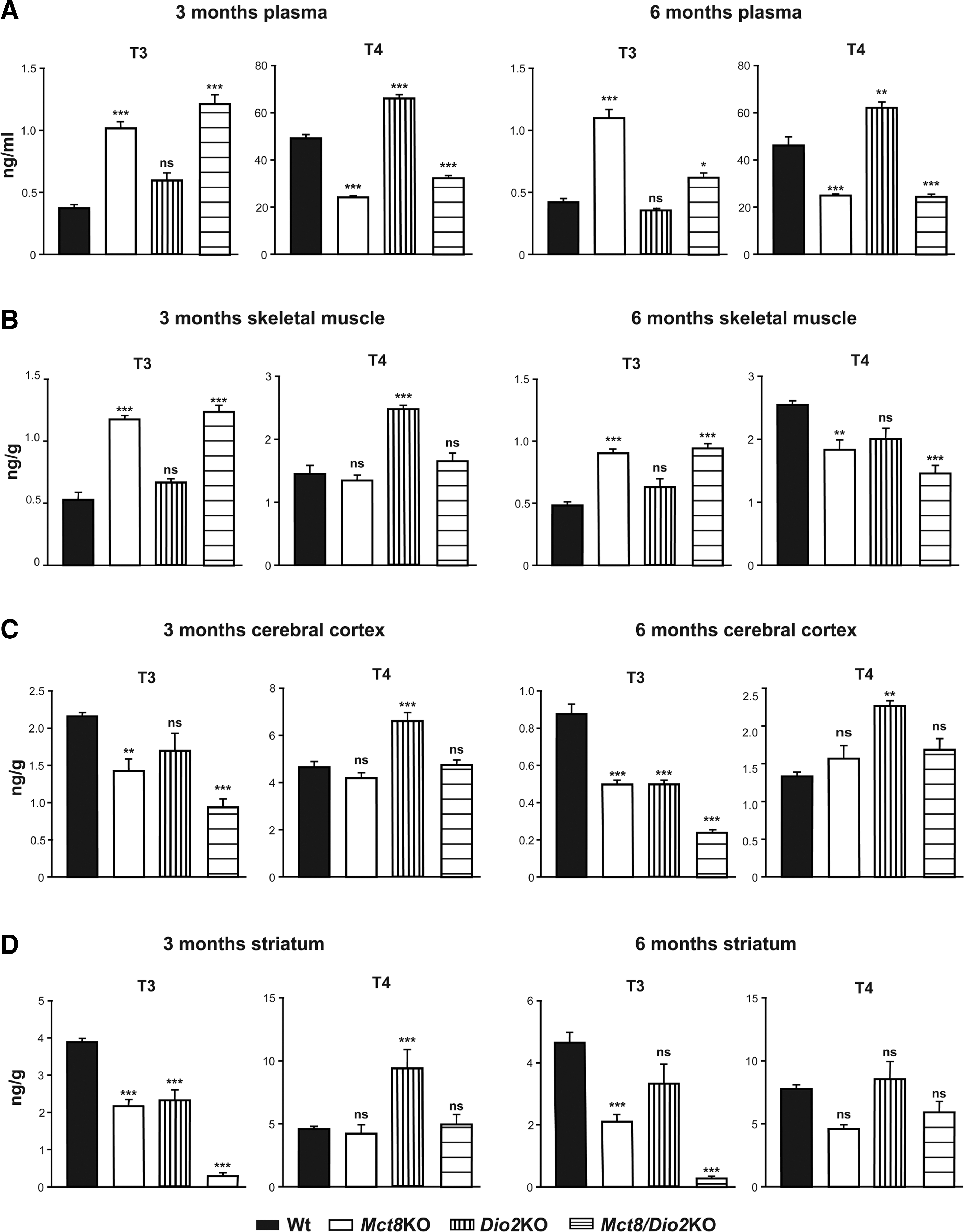
Thyroid hormone levels in plasma and tissues.
Owing to the deleterious effects that the peripheral hyperthyroidism causes on the muscle of MCT8-deficient patients, we measured TH content in the skeletal muscle. We found that three- and six-month-old Mct8KO mice present high T3 content in the skeletal muscle (2.5-fold at three months and 2-fold at six months of age in comparison to Wt) as previously reported in three-month-old mice (42,43), and no alterations in T4 content in the skeletal muscle at three months of age as recently found (43). Interestingly, we observed that there is a 1.3-fold reduction in the skeletal muscle content of T4 in six-month-old Mct8KO animals, indicating a change in the muscular TH content with age in comparison with Wt values. Dio2KO mice displayed high T4 content in the skeletal muscle (1.3-fold) at three months of age but T4 content was not altered in six-month-old Dio2KO mice, consistent with our previous findings (36). Most importantly, double Mct8/Dio2KO mice presented high T3 content in the skeletal muscle (threefold at three months of age and twofold at six months of age), no alterations in the T4 content at three months of age but, as observed in the single Mct8KO animals, decreased T4 content (1.7-fold) in the skeletal muscle of six-month-old Mct8/Dio2KO mice in comparison with Wt animals, again indicating variations in the TH content of the skeletal muscle with age (Fig. 1B).
In the cerebral cortex, three- and six-month-old single Mct8KO mice presented a 1.5-fold reduction in the T3 content as previously found (17,18) and no change in the content of T4. Dio2KO mice presented a 1.25-fold decrease in the cerebral cortex content of T3 at three months of age (although this decrease was not statistically significant), a 1.70-fold reduction in the T3 content at six months of age, and a 1.5-fold increase in the content of T4 in the cerebral cortex at both ages, consistent with previous findings (36,41). Double Mct8/Dio2KO mice presented a strong decrease in the content of T3 in the cerebral cortex at both ages (2.25-fold at three months and >3.5-fold at six months of age) and no variations in the content of T4 (Fig. 1C).
In the striatum, another brain region that contains the caudate-putamen and the globus pallidus, Mct8KO mice exhibited a decrease in the content of T3 at three (1.75-fold) and six months of age (2.25-fold) and a slight decrease in the content of T4 at six months of age that was not statistically significant. Three-month-old Dio2KO mice presented a 1.5-fold increase in the content of T4 in this region. Finally, double Mct8/Dio2KO displayed a severe decrease in the content of T3 in the striatum at three and at six months of age (13- and 18-fold, respectively) and no changes in the content of T4 (Fig. 1D).
When we studied the morphology of the thyroid gland in six-month-old Mct8/Dio2KO animals, we found larger follicular areas (1.7-fold) in comparison with Wt animals (Fig. 2), as previously described for several mice models lacking functional Mct8 protein expression (30,40,44).
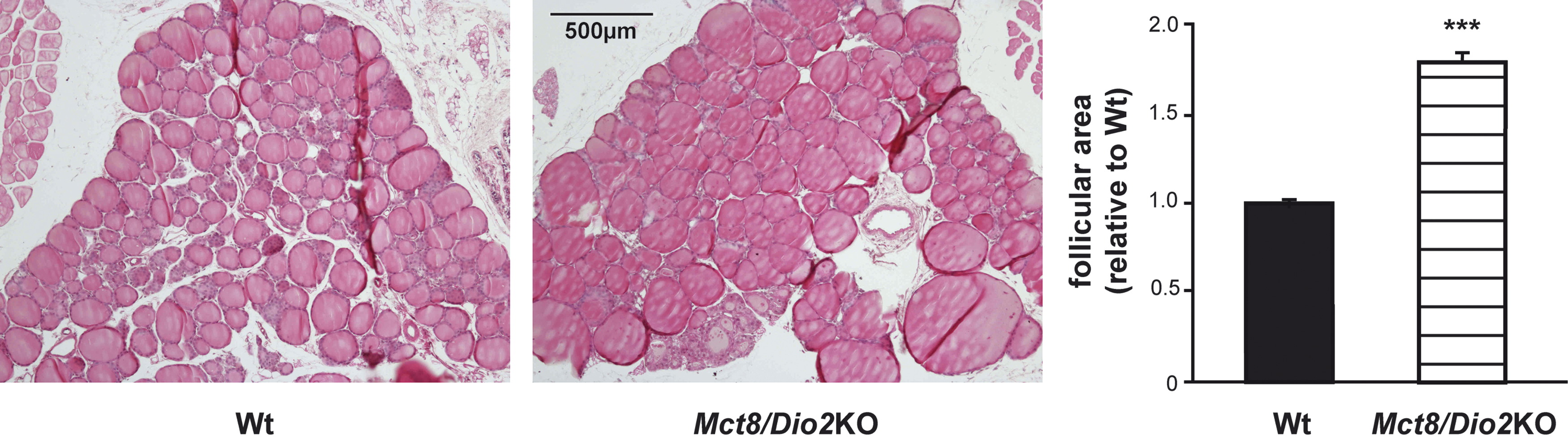
Thyroid gland histology of Wt and Mct8/Dio2KO mice. Thyroid gland sections from Wt (n = 4) and Mct8/Dio2KO mice (n = 4) at six months of age were stained with hematoxylin and eosin. The area of 50 follicles was measured in four slices for each animal. Data represent the mean of the follicle size (±SEM) and are expressed relative to Wt. ***p < 0.001 determined by Student's t-test.
Mct8/Dio2KO mice present pronounced motor skill impairments
First, we evaluated the ability of Wt, Mct8KO, Dio2KO, and double Mct8/Dio2KO mice to perform motor tasks exploring gait, coordination, balance, and muscle strength. As age can be an influential factor in these tests, and taking into account that most patients are diagnosed late after birth, these studies were performed in young adult mice (three months of age) and adult mice (six months of age). We found no abnormalities in Mct8KO animals as previously reported (20); however, we found profound motor skill impairments in Mct8/Dio2KO mice.
The gait of the animals was studied by the footprint test. Mct8/Dio2KO mice exhibited an altered gait pattern in comparison with Wt animals with increased hind limb stride width, and decreased forelimb and hind limb stride length. Interestingly, forelimb stride width was no different from that of Wt animals. These observations were similar at three and six months of age. Mct8KO animals exhibited gait parameters similar to those of Wt mice and Dio2KO presented decreased forelimb stride length only at three months of age (Fig. 3).
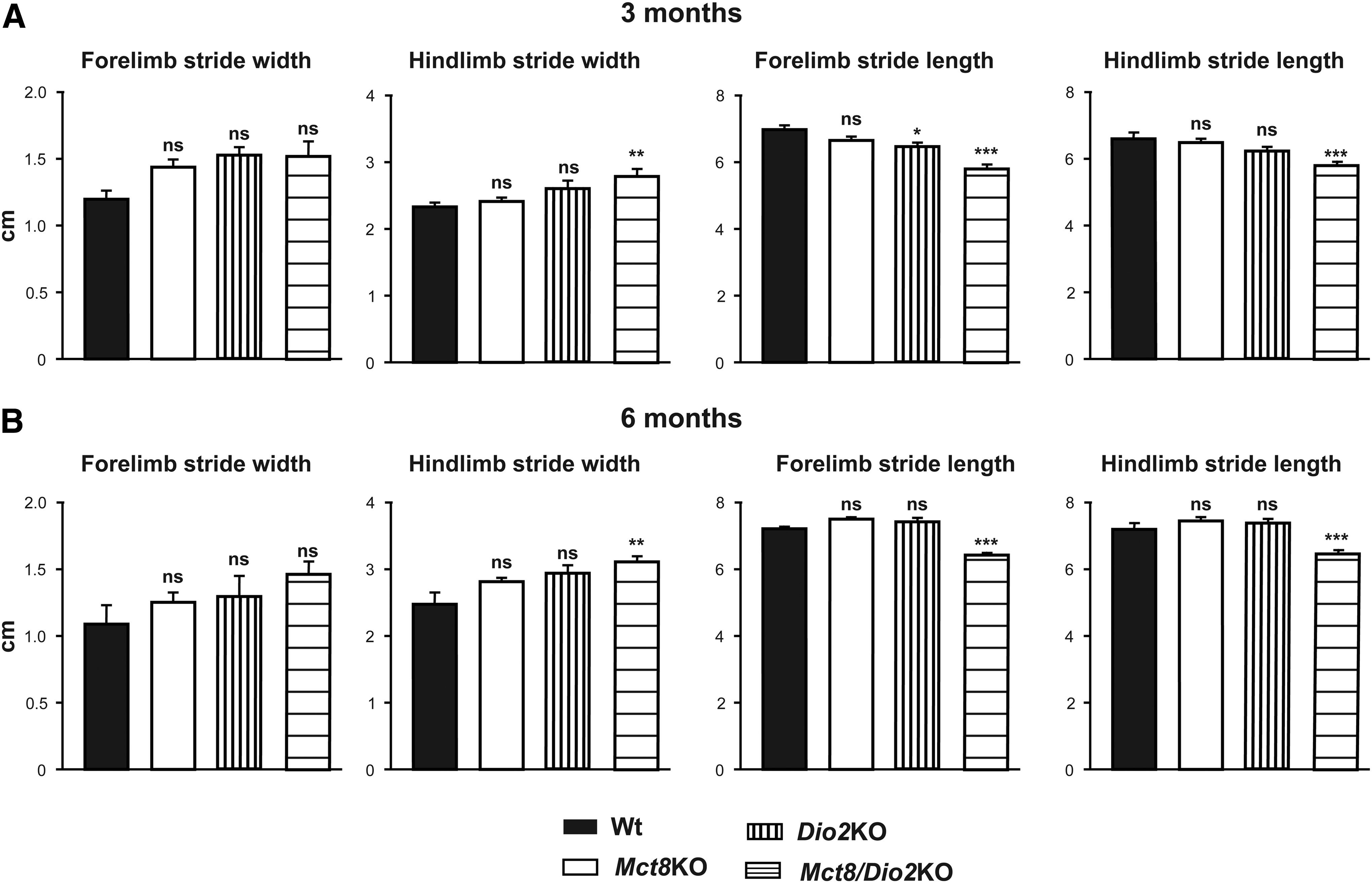
Abnormal gait in Mct8/Dio2KO mice.
To further characterize the locomotor behavior of Mct8/Dio2KO mice, we studied their performance in the accelerating rotarod test. The ability of the animals to stay on the accelerating rod was tested for five consecutive days at three and six months of age. Three-month-old Mct8/Dio2KO animals tended to fall from the rotarod earlier than the mice with the other genotypes, but the latency to fall off the rod was only significantly decreased during the first day of the test. There were no differences in the locomotor performance of three-month-old Wt, Mct8KO, and Dio2KO mice. At six months of age, Mct8/Dio2KO mice showed pronounced alterations in their performance in the rotarod test, as the latency to fall off the rod was much lower in Mct8/Dio2KO mice than in Wt animals in every day of the test. At this age, neither Mct8KO nor Dio2KO animals showed any statistically significant differences in the performance in the rotarod test in comparison with that of Wt mice (Fig. 4A). The alterations in rotarod performance observed in six-month-old Mct8/Dio2KO animals in comparison with Wt animals are unlikely to be related to the low body weight of Mct8/Dio2KO animals, since six months old Mct8/Dio2KO mice have increased body weight in comparison with younger mice (e.g., three-month-old), and their performance in the rotarod is much more impaired than at three months of age.
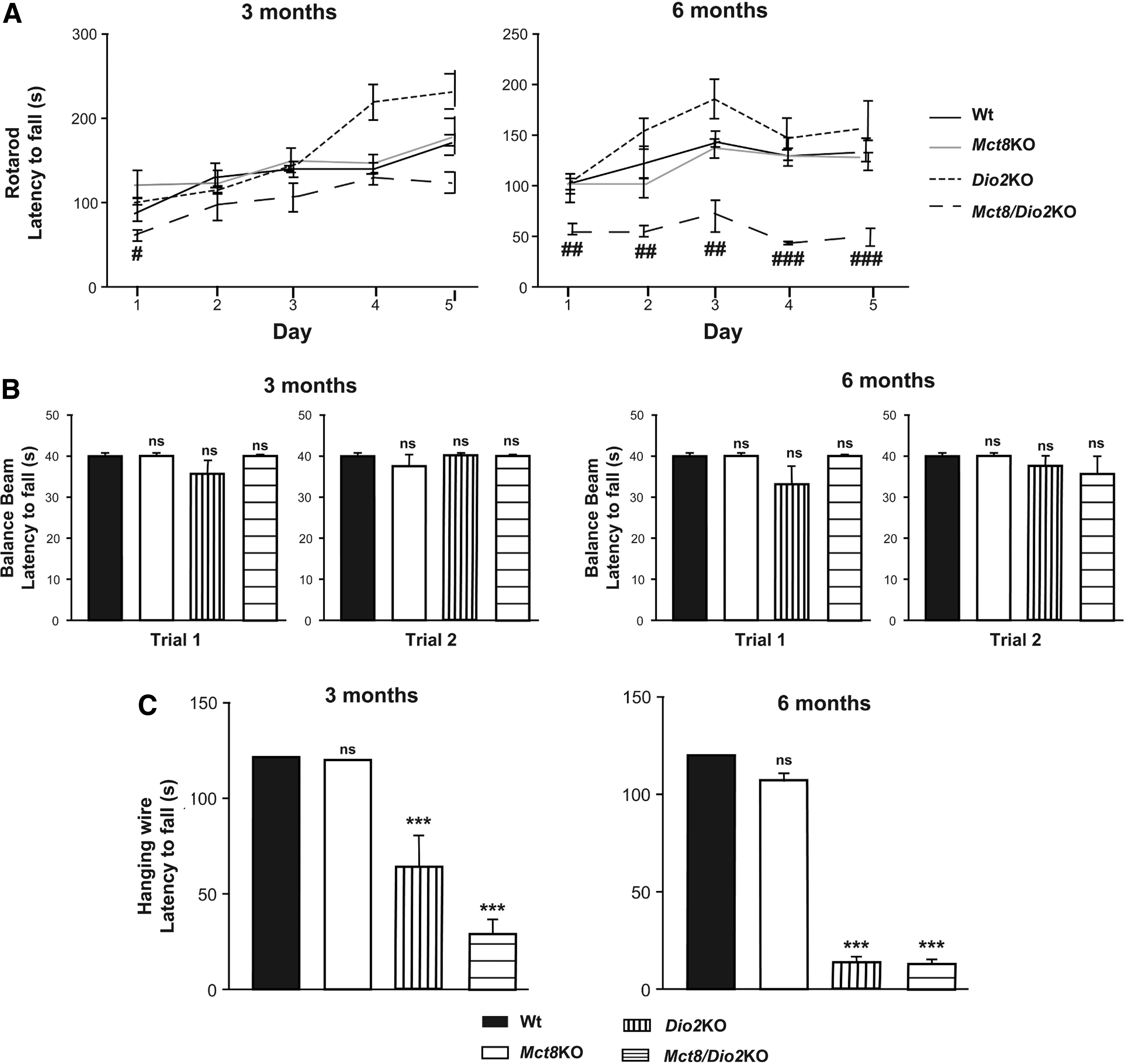
Motor skill impairments in Mct8/Dio2KO mice.
Equilibrium is not affected in Mct8/Dio2KO mice as can be seen from the balance beam test. The latency to fall off the beam was not different in Wt, Mct8KO, Dio2KO, and Mct8/Dio2KO animals at three and six months of age neither in the first nor the second trial (Fig. 4B). The activity on the bar was not different between Wt and Mct8/Dio2KO animals. At three months of age, 70% and 50% of Wt and Mct8/Dio2KO animals, respectively, reached the end of the bar on the first trial (p = 0.6499) and, in the second trial, 90% of the Wt mice and 60% of Mct8/Dio2KO animals arrived to the end of the bar (p = 0.3034.). At six months of age, on the first trial, 90% and 55% of Wt and Mct8/Dio2KO animals reached the end of the bar (p = 0.1409) and, in the second trial, 100% of the Wt animals and 66% of Mct8/Dio2KO animals walked to the end of the bar (p = 0.0867).
Finally, we explored muscle strength in the four limb hanging wire test. Mct8/Dio2KO mice displayed severe alterations in muscle strength in comparison with their matched age control Wt animals. While Wt and Mct8KO animals were able to hold from the grid for the entire test period (120 seconds), the mean holding latency value was 28.70 seconds for three-month-old Mct8/Dio2KO mice and 13.63 seconds for six-month-old Mct8/Dio2KO mice. Grip strength was also severely impaired in Dio2KO mice with a mean holding latency value of 64.19 seconds for three-month-old mice and 14.43 seconds for six-month-old animals (Fig. 4C).
Mct8/Dio2KO mice present brain alterations at the histological level
The only study that has explored the neuropathology of the syndrome by analyzing brain tissue sections from MCT8-deficient subjects published so far revealed serious alterations in brains of a 30th gestational week male fetus and an 11-year-old boy. Among these alterations, there was a loss of Pvalb expression and impaired axonal maturation (19).
To assess whether Mct8/Dio2KO mice replicate some of these alterations, we extensively characterized the expression of Pvalb and neurofilament 220KD (Nefh) in brain slices of Wt, Mct8KO, Dio2KO, and Mct8/Dio2KO mice at three and six months of age. We focused the analysis on the cerebral cortex (motor and somatosensory cortex) and the basal ganglia (caudate-putamen and substantia nigra) due to the significant impairments observed in these regions.
As shown in Figure 5, at three months of age, the expression of the calcium-binding protein Pvalb was abnormal in the cerebral cortex of Mct8/Dio2KO animals, which displayed a severe decrease in the expression of this protein in the motor and the somatosensory cortex in comparison with those in Wt animals (55.6% ± 5.93%, p = 0.0049 and 56.4% ± 0.80%, p = 0.0002, respectively, as compared with 100% for the Wt group). The expression of Pvalb was also decreased in these regions in the Mct8KO mice, although to a much lesser extent compared with Mct8/Dio2KO animals (65.4% ± 6.09%, p = 0.0084 and 75.1% ± 2.37%, p = 0.0013). Dio2KO mice also showed a significant reduction at the motor cortex (70.5% ± 2.54%, p = 0.0059) and especially at the somatosensory cortex (54.6% ± 3.78%, p = 0.0001). In the basal ganglia, immunostaining of Pvalb was strongly decreased in the caudate-putamen and the substantia nigra of Mct8/Dio2KO mice when compared with those in Wt animals (59.4% ± 6.53%, p = 0.0015 for caudate-putamen). The expression of this protein was also reduced in the basal ganglia of Mct8KO and Dio2KO animals, although this reduction was not as strong as in Mct8/Dio2KO mice (76.8% ± 4.35%, p = 0.0129 and 75.2% ± 3.28%, p = 0.0041, respectively).

Altered Parvalbumin expression in three-month old Mct8/Dio2KO mice. Brain coronal sections of Wt (n = 4), Mct8KO (n = 4), Dio2KO (n = 4), and Mct8/Dio2KO (n = 4) mice were immunostained with anti-Pvalb. Representative images showing the immunostaining pattern in different regions of the cerebral cortex (motor and somatosensorial cortex) and basal ganglia (caudate-putamen and substantia nigra). Pvalb immunostaining intensity was strongly decreased in all the regions in Mct8/Dio2KO mice in comparison with Wt mice. Sections of Mct8KO and Dio2KO mice also showed a reduction in the staining pattern but much less pronounced than in Mct8/Dio2KO animals. Pvalb, Parvalbumin.
As shown in Figure 6, at three months of age, there was a distinctive staining pattern for Nefh in Mct8/Dio2KO mice. The expression of this protein was markedly reduced in the cerebral cortex of Mct8/Dio2KO mice, especially at the somatosensory cortex (63.4% ± 6.47%, p = 0.0090 and 51.6% ± 3.90%, p < 0.0001, for motor and somatosensory cortex, respectively). At the basal ganglia, there was also a reduction in the intensity of expression of Nefh in the caudate-putamen and the substantia nigra in Mct8/Dio2KO animals in comparison with Wt (77.7% ± 3.99%, p = 0.0276 for caudate-putamen). In Mct8KO and Dio2KO mice, the expression of Nefh in the cerebral cortex was also decreased, although not as severely as in double Mct8/Dio2KO mice (78.6% ± 5.71%, p = 0.0454 and 78.9% ± 5.93%, p = 0.0216 for Mct8KO and 78.5% ± 4.59%, p = 0.0278 and 81.7% ± 2.81%, p = 0.0062 for Dio2KO mice). However, the expression of Nefh in basal ganglia was very similar to control Wt animals (84.2% ± 7.20%, p = 0.1554 for Mct8KO and 92.8% ± 3.23%, p = 0.3664 for Dio2KO animals in the caudate-putamen).

Altered neurofilament 220KD expression in three-month old Mct8/Dio2KO mice. Representative images of brain coronal sections of Wt (n = 4), Mct8KO (n = 4), Dio2KO (n = 4), and Mct8/Dio2KO (n = 4) mice immunostained with anti-Nefh. Mct8/Dio2KO mice showed a markedly reduced staining especially in the motor cortex and substantia nigra. Mct8KO and Dio2KO mice also showed a decreased immunostaining intensity in the cerebral cortex (motor and somatosensory cortex) but to lesser extent than Mct8/Dio2KO mice. In basal ganglia (caudate-putamen and substantia nigra), Mct8KO and Dio2KO mice showed a Nefh-staining pattern similar to Wt animals.
At six months of age, the immunostaining pattern for Pvalb and Nefh was very similar to that observed in three-month-old mice. In general, there was a strong decrease in the expression of these two proteins in the double Mct8/Dio2KO mice in comparison with Wt mice in all studied regions, and this decrease was much less pronounced in the Mct8KO or Dio2KO mice (data not shown).
Discussion
Owing to the severe alterations that arise from mutations in the TH transporter MCT8, much effort has been put into understanding the causes that underlie such alterations and into developing successful therapeutic approaches. As data available from humans are limited (45,46), the generation of appropriate models and instruments is essential to achieve these goals. The lack of the perfect model is hindering our understanding of MCT8 deficiency; therefore, the use of a comparative approach combining several models will be the best option to tackle this problem.
Regarding mouse models, single Mct8KO mice are only a partial model of the disease as they do not reproduce the neurological alterations present in humans. Nevertheless, this model has provided much of the current knowledge regarding MCT8 deficiency. To begin with, Mct8KO mice have been useful to prove that despite having high serum T3 concentrations, there is a state of brain hypothyroidism with reduced T3 content (17,18), and alterations in the expression of some T3-dependent genes (21). Moreover, thanks to elegant studies using Mct8KO mice, it was first suggested that the pathogenesis associated with MCT8 deficiency arises from an impaired T3 transport across the BBB (14). The double Mct8/Oatp1c1KO mouse seems to be a better model for the AHDS as it presents peripheral hyperthyroidism and brain alterations compatible with brain hypothyroidism. However, the fact that Oatp1c1 transports other molecules (28) and that OATP1C1 deficiency in humans has been associated with neurological alterations (29) offers some limitations to consider the Mct8/Oatp1c1KO mouse as the ultimate model for the AHDS and highlights the need for alternative and additional models that support and complement each other. In this study, we have characterized the Mct8/Dio2KO mouse as an alternative mouse model for the AHDS. We have found that young adult and adult Mct8/Dio2KO animals replicate the situation in MCT8-deficient patients with peripheral hyperthyroidism with increased plasma T3 levels and a state of brain hypothyroidism with decreased T3 content. In addition, they also present brain alterations at a histological level and display motor skill impairments compatible with the psychomotor abnormalities in MCT8-deficient patients. This model, however, also presents certain limitations as it is deprived of Dio2, when patients present increased Dio2 activity in the brain, and does not fully replicate the low T4 content in the brain of MCT8-deficient patients (19).
The three Mct8-deficient mouse models (Mct8KO, Mct8/Oatp1c1KO, and Mct8/Dio2KO), like MCT8-deficient patients, present increased T3 plasma levels that lead to thyrotoxicosis due to an excess of T3 in peripheral tissues. Indeed, studies in three-month-old Mct8KO mice have demonstrated that these mice have hypermetabolism with increased total energy expenditure (42) and that, despite lacking Mct8, there is a hyperthyroid state of the kidneys (47) and the skeletal muscle (42). Double Mct8/Oatp1c1KO mice have been recently found to also present a thyrotoxic state in skeletal muscle (43), and we now provide evidence that Mct8/Dio2KO mice also exhibit a hyperthyroid state of the skeletal muscle at three and six months of age. Regarding T4 content in the skeletal muscle, we have found that three-month-old Mct8KO and Mct8/Dio2KO mice present normal T4 values despite low T4 circulating levels consistent with previous data in Mct8KO mice (42) and Mct8/Oatp1c1KO mice (43) of approximately the same age. Intriguingly, T4 content was low in six-month-old Mct8KO and Mct8/Dio2KO mice, suggesting that there are variations in the content of THs with age. These findings are relevant as the skeletal muscle is an important TH target (48) and the Mct8 transporter seems to play an important role in muscle regeneration (43). Analysis of these three mouse models will provide further evidence regarding the peripheral effects of MCT8 deficiency at different ages.
Another important finding of this study is that there seems to be selective sensitivity to Mct8 deficiency across brain regions, as previously suggested (49). In the cerebral cortex, we have found a decrease of T3 in three- and six-month-old Mct8KO animals to ∼60% of the content in Wt animals, while in the striatum, T3 content decreased to 55% and 40% of the Wt values at three and six months of age, respectively. This indicates that the striatum is more sensitive to Mct8 deficiency than the cerebral cortex. This finding was more robust in double Mct8/Dio2KO animals. In the cerebral cortex, there was a decline in the content of T3 to ∼40% and 25% of the Wt values at three and six months of age, respectively, and in the striatum there was a decrease to 8% and 5% of the Wt content at three and six months of age, respectively. The striatum includes the caudate-putamen and the globus pallidus, which are the main components of the basal ganglia, responsible mainly for motor control and also for motor learning, executive functions, and behaviors and emotions (50). MCT8-deficient patients have been shown to have important lesions in the putamen by MRI studies (11), so our results could have important implications for the understanding of the syndrome and its treatment. Owing to the poor control movement of MCT8-deficient patients, these findings could point to the basal ganglia as an important target for therapeutic treatments.
To continue with the brain, these three Mct8-deficient mouse models present a state of brain hypothyroidism with decreased T3 content. Cerebral T3 content in Mct8KO and Mct8/Dio2KO mice has already been discussed and double Mct8/Oatp1c1KO mice have been shown to have a decrease of T3 to ∼10% of the levels in Wt animals in the forebrain at P21. The state of brain hypothyroidism leads to alterations at the histological level to different extent in each model. We have found very mild alterations with a slight reduction in the expression of Pvalb in Mct8KO animals as previously shown (20). In this study, the authors characterized the immunohistochemical expression of Pvalb in Mct8KO mice for the first time and they concluded that there were no alterations in the number or distribution of Pvalb-positive interneurons.
Like in Mct8/Oatp1c1KO mice (27), we have detected important alterations in the expression of Pvalb in the brain of Mct8/Dio2KO mice at three and six months of age. There was reduced expression of Pvalb in the motor and the somatosensory cortex and in the basal ganglia. This is consistent with the reduced T3 content found in the cerebral cortex and the striatum of Mct8/Dio2KO mice and with previous studies that have detected alterations in Pvalb immunoreactivity under TH insufficiency conditions (51 –53). Most importantly, this finding is in agreement with observations in the brain of an MCT8-deficient subject wherein PVALB expression was not detected in the cerebral cortex (19). In this same study, the expression of neurofilaments immunoreactivity was remarkably reduced or even absent in the cerebral cortex of a MCT8-deficient subject. In this study, we have found reduced immunoreactivity of Nefh in the cerebral cortex, the caudate-putamen, and the substantia nigra of Mct8/Dio2KO mice. As in the case of Pvalb, this is in agreement with the reduced T3 content detected in the cerebral cortex and the striatum of Mct8/Dio2KO mice and with previous studies describing alterations in neurofilament immunoreactivity under TH deficiency (54 –56). These findings might have important implications as the reduction in Pvalb immunoreactivity suggests that there are profound alterations in the inhibitory interneurons that mediate feedforward inhibition in thalamocortical, interlaminar, and interareal circuits (57), and the decrease in Nefh immunostaining indicates that there are important neuronal maturation impairments (58).
We have also found important motor skill impairments in Mct8/Dio2KO mice. These alterations have also been described in Mct8/Oatp1c1KO animals and include variations in global gait pattern, reduced grip strength, and decreased motor coordination. Interestingly, while Mct8/Oap1c1KO mice exhibit abnormalities in the rotarod performance in every day of the test at three months of age, Mct8/Dio2KO showed an altered performance pattern that was only statistically significant on the first day of training at this age. However, rotarod performance of six-month-old Mct8/Dio2KO mice showed severe impairments in every day of test. This raises the question about the underlying differences between developmental stages and between the mouse genotypes that may explain the differential rotarod performance.
Studies in MCT8-deficient subjects show that patients present important brain alterations already from prenatal stages (19). It is, therefore, indispensable to study these animal models in perinatal stages to unravel the mechanisms underlying MCT8-deficient brain development and to design appropriate therapeutic strategies. Mct8/Dio2KO mice have decreased expression of some T3-dependent genes at postnatal day 3 in the brain (59), but further analysis should be performed. This is especially important as the role of MCT8 in the perinatal period is still unclear. Single Mct8KO mice present with cerebral hyperthyroidism during the perinatal period (60,61). Moreover, Mct8-deficient zebrafish present neurological and behavioral deficiencies very early in development without changes in the expression of the T3-target genes Klf9 and Nrgna, suggesting that MCT8 might have a TH-independent role that induces neurological deficiencies in the perinatal period (62). The use of the current available mouse models and additional models including human patient-derived induced pluripotent stem cells (16) and nonmammalian vertebrates such as zebrafish (63), chicken (64), or even Xenopus (65) will be necessary to explore this and other aspects of the syndrome and to provide additional insights into the human disease.
In summary, this study expands the phenotypic characterization of Mct8/Dio2KO mice as an additional and alternative model for human MCT8 deficiency. Three- and six-month-old Mct8/Dio2KO mice display abnormal circulating TH levels with low T4 and high T3 resembling the human condition, a state of brain hypothyroidism with reduced T3 content varying across brain regions, alterations at the histological level and locomotor impairments. Importantly, the observed alterations persist and even aggravate with age. In addition, this mouse model has already been reported to present changes in the expression of T3-target genes in the brain of juvenile (21) and perinatal (59) animals. Indeed, Mct8/Dio2KO mice have already been used to test possible therapeutic interventions for MCT8 deficiency (39), but the findings presented here offer additional markers to expand the markers allowing to assess the impact of therapeutic interventions.
Further characterization of this and other models at different developmental stages will provide further insights into the molecular underpinnings of developmental brain disorders under TH deficiency conditions and will offer additional tools to develop therapeutic strategies to prevent and/or restore brain damage in MCT8-deficient patients.
Footnotes
Acknowledgments
We thank Dr. Maria Jesus Obregon for her valuable assistance with thyroid hormone determinations, María Camino de Lucas for animal care, and Dr. Juan Bernal for critical reading of the article and helpful discussions. We thank Dr. Valerie Anne Galton and Dr. Samuel Refetoff for the generous gift of Dio2KO and Mct8KO mice, respectively.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by grants from the Spanish Plan Nacional de I+D+i (Grant No. SAF2017-86342-R to A.GF), the Sherman Foundation (Grant No. OTR02211 to A.GF and S.BL), and the Center for Biomedical Research on Rare Diseases (Ciberer), Instituto de Salud Carlos III, Madrid, Spain. The cost of this publication has been paid, in part, by FEDER funds.