
Abstract
Background:
Sepsis can cause the nonthyroidal illness syndrome (NTIS), resulting in perturbed thyroid hormone (TH) signaling and reduced thyroxine (T4) levels. TH is a major regulator of muscle function, via its influence on mitochondria. This study aimed at evaluating the relationship between TH signaling, mitochondrial function, and the antioxidant defense system in the diaphragms of septic mice.
Methods:
Male C57Bl/6 mice were divided into two groups: cecal ligation and puncture (CLP) and sham. Twenty-four hours after surgery, plasma, diaphragms, and livers were collected. TH metabolism and responses were analyzed by measuring messenger RNA (mRNA) expression of Dio1 in the liver, and Thra, Thrb, Dio2, Slc16a10, and Slc16a2 (encodes MCT 10 and 8), in the diaphragm. T4 plasma levels were measured by radioimmunoassay. Damage to diaphragm mitochondria was assessed by electron microscopy and real-time polymerase chain reaction (qPCR), and function with oxygraphy. The diaphragm antioxidative defense system was examined by qPCR, analyzing superoxide dismutase (SOD) 1 (Sod1), mitochondrial superoxide dismutase (SOD 2; Sod2), extracellular superoxide dismutase (SOD 3; Sod3), glutathione peroxidase 1 (Gpx1), and catalase (Cat) expression. The effect of TH replacement was tested by treating the mice with T4 and triiodothyronine (T3) (CLP+TH) after surgery.
Results:
CLP mice presented reduced total plasma T4 concentrations, downregulated Dio1, and upregulated Il1b mRNA expression in the liver. CLP mice also displayed downregulated Thra, Thrb, Slc16a10, and Slc16a2 expression in the diaphragm, suggesting that TH signaling was compromised. The expression of Ppargc1a (encoding PGC1a) was downregulated, which correlated with the decrease in the number of total mitochondria, increase in the percentage of injured mitochondria, downregulation of respiratory chain complex 2 and 3 mRNA expression, and reduced maximal respiration. In addition, septic animals presented a three-fold increase in Ucp3 and G6pdh expression; downregulated Sod3, Gpx1, and Cat expression; and upregulated Sod2 expression, potentially due to elevated reactive oxygen species levels. The mitochondrial number and the percentage of injured mitochondrial were similar between sham and CLP+TH mice.
Conclusions:
Sepsis induced responses consistent with NTIS, resulted in mitochondrial damage and functional impairment, and modulated the expression of key antioxidant enzymes in the diaphragm. Thus, impaired diaphragm function during sepsis seems to involve altered local TH signaling, mitochondrial dysfunction, and oxidative stress defense.
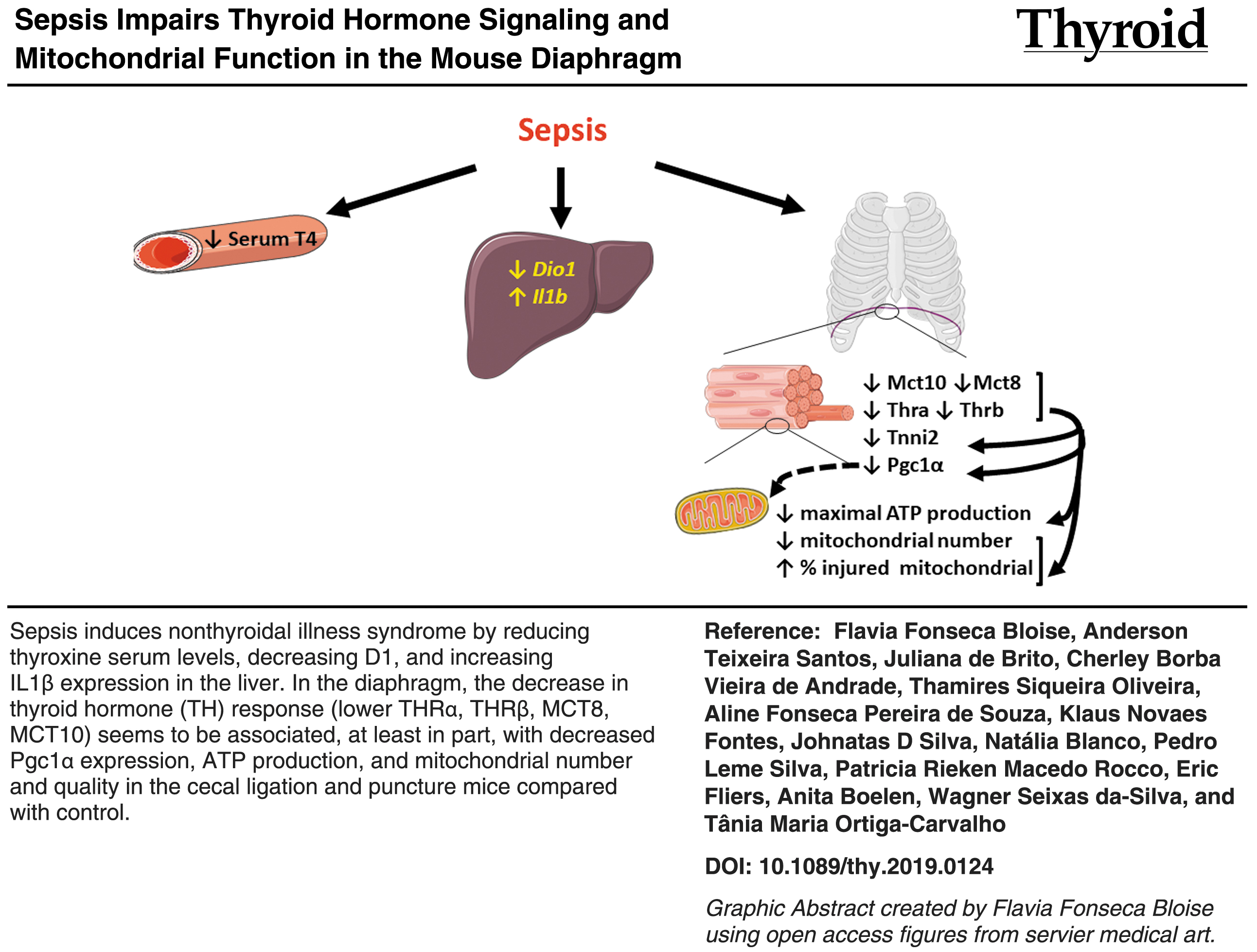
Color images are available online.
Introduction
Sepsis is a life-threatening condition associated with an unbalanced host response to infection (1). Septic patients frequently require expensive and intensive treatments, thus making sepsis a major public health concern (2 –4). It is also known that sepsis induces the nonthyroidal illness syndrome (NTIS) (5), and the severity of NTIS is associated with the mortality rate due to sepsis (6,7).
NTIS is characterized by a decrease in peripheral thyroid hormone (TH) serum levels while thyrotropin levels remain unchanged (8). NTIS can be viewed as part of the inflammatory response, and the changes in serum TH concentrations have been shown to modulate central and peripheral TH signaling and metabolism (8,9). Septic patients with lower triiodothyronine (T3) and thyroxine (T4) serum levels have a poor prognosis for a favorable outcome (5,10). In our previous work, it was found that sepsis, induced by cecal ligation and puncture (CLP) surgery, led to changes in TH serum levels and TH metabolism in the liver, which are also commonly observed during NTIS (11).
Further, in some cases, septic patients require mechanical ventilation, which can result in injury of the diaphragm. Further, sepsis can induce diaphragm weakness independent of mechanical ventilation (12). The mortality rate is greater in patients with increased diaphragm weakness, when compared with patients with a similar disease severity and without this condition (12). Currently, the mechanisms involved in septic-mediated diaphragm impairment are not known, and they require clarification.
Skeletal muscle function is directly and indirectly related to mitochondrial biogenesis and energy production (13). Further, TH is crucial to normal skeletal muscle physiology, since it is required for tissue regeneration and differentiation and is involved in the regulation of mitochondrial function. In addition, TH has been shown to be associated with the regulation of sarcolemmal Ca+2 concentrations (14). Interestingly, stimulation of a muscle cell line with lipopolysaccharide, a bacterial endotoxin, decreased mitochondrial function in a TH-dependent manner (15). Moreover, our group previously demonstrated that sepsis, induced by S. pneumoniae infection, decreased the responsiveness of the diaphragm to TH (15).
We, therefore, hypothesized that altered TH signaling induces changes in mitochondrial function and, consequently, impairs the diaphragm function of septic animals. The impact of sepsis on TH signaling, mitochondrial function, and antioxidant defense in the diaphragm were investigated by using real-time polymerase chain reaction (qPCR), electron microscopy, and oxygraphy in a CLP-induced sepsis model.
Materials and Methods
Ethics statement and animal experimentation
The project was approved by the local Animal Care Committee at the Federal University of Rio de Janeiro (CEUA-088/15), and it was registered with the Brazilian National Council for Animal Experimentation Control. Animal care was in compliance with the “Guide for the Care and Use of Laboratory Animals” established by the U.S. National Academy of Sciences (16).
Sepsis was induced by CLP and control C57Bl6 male mice that were two to three months old, weighing 24–26 g, and that were subjected to sham surgery, as previously described (11). Briefly, mice were anesthetized with ketamine (100 mg/kg; Cristália, Itapira, Brazil) and xylazine (10 mg/kg; Syntec) i.p. After anesthesia, the lower abdomen area was disinfected. Longitudinal incisions were performed in the lower quadrant of the abdomen and peritoneal cavity with scissors. Later, the cecum was located and exteriorized. The cecal ligation was performed below the ileocecal valve by using a cotton suture line. The cecum was perforated once between the ligation and the end of the cecum by using a 18G needle. A small amount of fecal material was exposed before reinsertion of the cecum in the abdominal cavity. The fascia, abdominal musculature, peritoneum, and skin were closed via simple running sutures by using a nylon suture line. Subsequently, 1 mL of sterile saline was administrated subcutaneously for volume substitution. The sham-operated mice (control) underwent the same procedure of CLP mice, excluding cecal ligation and perforation. Animals were housed in a temperature control environment (22°C) with a 12-h light/dark cycle and food and water ad libitum.
At 8 and 23 hours after CLP or sham operation, mice were scored for severity of sepsis (17). The following variable were evaluated: piloerection, alterations in gait, lethargy, alterations in respiratory rate, lacrimation, grip strength, body tone, respiration after manipulation, lack of exploratory behavior, and body temperature alterations; if present, the animal receives one point. The sum of total score reflected the severity of sepsis. Score 2–3 points mild sepsis, 4–7 moderated sepsis, and 8–10 severe sepsis. In our experimental condition, most of the CLP animals were ranked as moderate sepsis.
A total of 15 control (sham), 26 CLP mice were used; 8 CLP mice were treated with TH (CLP+TH); and six CLP and two CLP+TH mice died 6 and 18 hours after the surgery and were excluded from the experiments. TH (250 ng T3/g body weight plus 7.5 ng T4/g body weight in a volume of 100 μL; both from Sigma Aldrich) was administrated subcutaneously immediately, four and eight hours after the surgery, and control mice received saline. For each analysis, the number of animals used are indicated in the legends.
Animals were sacrificed under anesthesia with isoflurane 1.5–2.0% (Isoforine; Cristália), 24 hours after surgery between 9 and 11 am, to avoid changes in hormonal and gene expression due to circadian rhythms. Blood, liver, and diaphragm were collected, immediately frozen in liquid nitrogen, and stored at −70°C, until use. Freshly dissected diaphragms were used for the mitochondrial studies. Hormone determinations were performed with blood serum that was stored at −20°C.
Plasma total T4 measurement
Plasma total T4 concentration was measured by using a commercial radioimmunoassay kit (MP Biomedicals, CA), according to the manufacturer's recommendations. The limit of detection for T4 was 13 μM.
Total RNA extraction and qPCR
Total RNA from liver samples was extracted by using the TRIzol method, according to the manufacturer's protocol (TRIzol Reagent; Life Technologies, CA). Total RNA from diaphragm samples was extracted by using a combination of the TRIzol Reagent (Invitrogen, Carlsbad) and the Nucleospin RNA II kit (Macherey Nagel, Düren, DE). For cDNA synthesis, 800 ng of total RNA was reverse transcribed by using the High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, CA), according to the manufacturer's protocol. After cDNA synthesis, messenger RNA (mRNA) expression was evaluated by qPCR, using the HOT FIREPol Evagreen qPCR Supermix (Solis, Denmark) and the Master Cycler Realplex system (Eppendorf, Germany).
The sequences for each primer pair are shown in Table 1, in summary for analyses TH responsiveness and metabolism: Dio1, Dio2, Thra1, Thrb1, Slc16a2, Slc16a10, Tnni2, and Hr; for systemic inflammation: Il1b; to investigate mitochondrial physiology: Ppargc1a, Opa1, and Dnm1l; Ucp3; G6pdh to analyze the respiratory chain: Ndufb8, Sdhb, Uqcrc2, and Cox4i1; to investigate reactive oxygen defenses: superoxide dismutase (SOD) 1 (Sod1), mitochondrial SOD2 (Sod2), extracellular SOD3 (Sod3), glutathione peroxidase 1 (Gpx1), and catalase (Cat). The reference genes are listed next. Quantification of mRNA expression was calculated by using the standard curve method and corrected by the geometric mean of the following reference genes: Phctr and Pib for the diaphragm (control 358.9 ± 26.7 and septic 388.8 ± 71.6, p = 0.6), Gapdh and Rpl0 for the liver (control 1032 ± 220 and septic 1115 ± 131, p = 0.6), and Vdac for mitochondrially encoded genes (control 1633 ± 94 and septic 1239 ± 262, p = 0.2).
Primer Pair's Sequences
The best reference genes were selected according to their Cq, amount values, low variance, and no significant differences between groups. The qPCR efficiency was between 90% and 100%. PCR cycles were as follows: denaturation for 12 minutes at 95°C, 40 cycles of 15 seconds at 95°C, 30 seconds of annealing temperature (65°C or 60°C), 30 seconds at 72°C, and the following melting program. Genomic DNA contamination and qPCR quality were evaluated by using intron-spanning primers, reverse transcriptase-negative samples, and melting curve analyses from each reaction.
Skeletal muscle high-resolution respirometry
Three milligrams of diaphragm was washed in chilled BIOPS buffer (2.77 mM CaK2EGTA, 7.23 mM K2EGTA, 20 mM imidazole, 20 mM taurine, 6.56 mM MgCl2, 5.77 mM ATP, 15 mM phosphocreatine, 0.5 mM dithiothreitol, and 50 mM K-MES, pH 7.1); the tissues were permeabilized with cold BIOPS buffer supplemented with 50 μg/mL saponin, for 40 minutes with gentle agitation. The samples were then transferred to Mir05 buffer (110 mM sucrose, 60 mM K-MES, 0.5 mM EGTA, 3 mM MgCl2, 20 mM taurine, 10 mM KH2PO4, 20 mM K-HEPES, and 1 g/L bovine serum albumin, pH 7.1) and maintained in this buffer for at least 10 minutes under mild agitation.
Oxygen consumption was measured with an O2K (Oroboros Instruments GmbH, Innsbruck, Austria) in Mir05 buffer. Basal O2 consumption was recorded for at least five minutes followed by drug administration in the following order: 5 mM pyruvate and 5 mM malate, 3 mM ADP, 10 μM cytochrome c, 10 mM succinate, two pulses of 1 μg/mL oligomycin, 600 nM Carbonyl cyanide-4-(trifluoromethoxy) phenylhydrazone, and 10 mM KCN. All data were recorded after O2 consumption stabilized.
Transmission electron microscopy and mitochondrial quantification
Fragments of the diaphragm were processed and analyzed qualitatively and quantitatively by using a JEM1011 microscope (JEOL). Briefly, the fragments were washed in PBS, fixed in 0.1 M sodium cacodylate buffer (pH 7.2) containing 2.5% glutaraldehyde for 24 hours, and washed three times, 10 minutes each, in the same buffer. The tissue was postfixed in 0.1 M sodium cacodylate buffer (pH 7.2) containing 1% osmium tetroxide (OsO4), dehydrated with increasing acetone concentrations (30%, 50%, 70%, 90%, and 100%), and embedded in Poly/Bed812 resin (Ted Pella, Inc., Redding, CA).
After polymerization, ultrathin sections were obtained and contrasted with uranyl acetate-lead citrate. For ultrastructural evaluations, sections were collected in three grids with a distance of 7 μm between them. A total of 10 images (70 nm) were acquired from random fields in each grid. The total number of mitochondria, as well as the number of healthy and injured mitochondria were quantified to obtain the percentage of injured mitochondria (18). Digital micrographs were captured by using an AMT XR80 CCD digital camera (Advanced Microscopy Techniques, Woburn, MA, EUA) at 12,000 × magnification
Statistical analysis
The Kolmogorov
Results
Sepsis due to CLP induces NTIS
The CLP procedure decreased total serum T4 levels by 33% (p < 0.01) when compared with sham-operated control animals (Fig. 1A). In addition, liver Dio1 mRNA expression was decreased by 42% (Fig. 1B, p < 0.05), while liver ll1b mRNA expression increased eight-fold (Fig. 1C, p < 0.01), when compared with controls. Liver Il1b is an acute-phase response marker and a T4 decrease associated with a decrease in liver Dio1 are hallmarks of NTIS. Thus, CLP induced systemic inflammation and NTIS.

Serum T4 levels and gene expression confirm that acute CLP is a nonthyroidal illness syndrome model. Total serum T4 (
Expression of Thra1 and Slc16a2 is reduced during sepsis in the diaphragm
Based on a previous study, demonstrating the involvement of Thra1, Thrb, Dio2, Slc16a10, and Slc16a2 in TH signaling (19), the expression of these genes was investigated in the diaphragms of septic and control mice. As shown in Figure 1D–H, the expression of Thra1, Thrb, Slc16a10, and Slc16a2 was reduced by 78% (p < 0.05), 71% (p < 0.0001), 70% (p < 0.05), and 55% (p < 0.0001), respectively, in the septic diaphragm, while Dio2 expression was unaffected. In summary, skeletal muscle TH receptors (Thra1 and Thrb) and transporters (Slc16a10 and Slc16a2) were decreased, suggesting a decreased responsiveness to TH action in the diaphragm. In addition, liver Dio1 is highly sensitive to TH and can be used to evaluate TH signaling. Thus, the investigation of TH responsive genes could help to elucidate this hypothesis.
Sepsis affects mitochondrial function
Ppargc1a is a T3 responsive gene of interest. Interestingly, the protein encoded by the Ppargc1a gene, PGC1α, is important for regulating transcription and stimulating mitochondrial biogenesis and function (14). In addition to other tissues, Ppargc1a is also expressed in skeletal muscle and has been used to confirm reduced TH signaling (20). Ppargc1a expression was reduced by 90% in the septic mice, when compared with controls (Fig. 2A, p < 0.01).
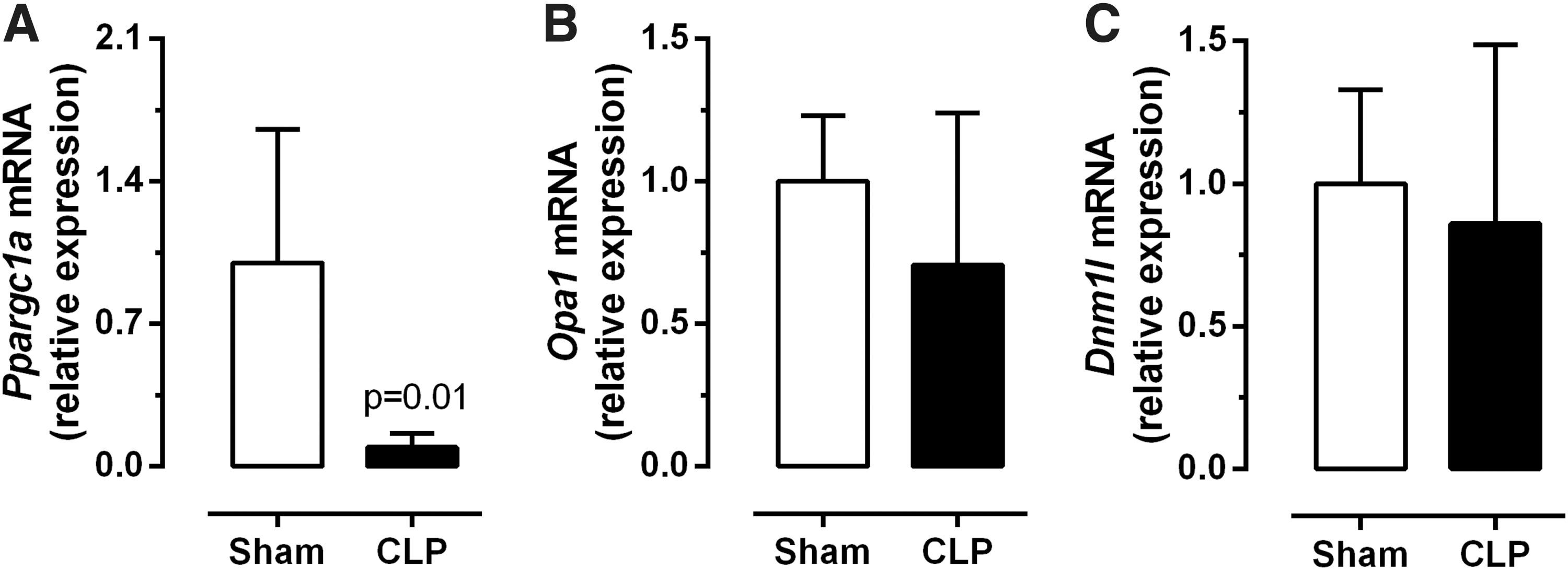
Mitochondrial dynamics in the diaphragm. Expression of Ppargc1a (
Given the reduced Ppargc1a mRNA expression levels in the diaphragm, markers of mitochondrial fusion (Opa1) and fission (Dnm1l) processes were also investigated, so as to characterize mitochondrial physiology in more detail. No significant differences were observed for the expression of Opa1 (Fig. 2B) or Dnm1l (Fig. 2C) in the diaphragms of septic or control animals.
On the other hand, the number of mitochondria was significantly reduced in the septic group, when compared with controls (Fig. 3A–E, p = 0.01). In addition, there was a detectable increase in the size of the mitochondria from septic animals; the increased organelle size was caused by mitochondrial swelling (Fig. 3A–D, F, p = 0.01). In fact, mitochondrial swelling is associated with reduced organelle function and increased permeability (21). Indeed, when quantifying mitochondrial damage in the diaphragm of septic and control mice, it was found that a greater percentage of injured organelles were detected in the septic animals (Fig. 3G).
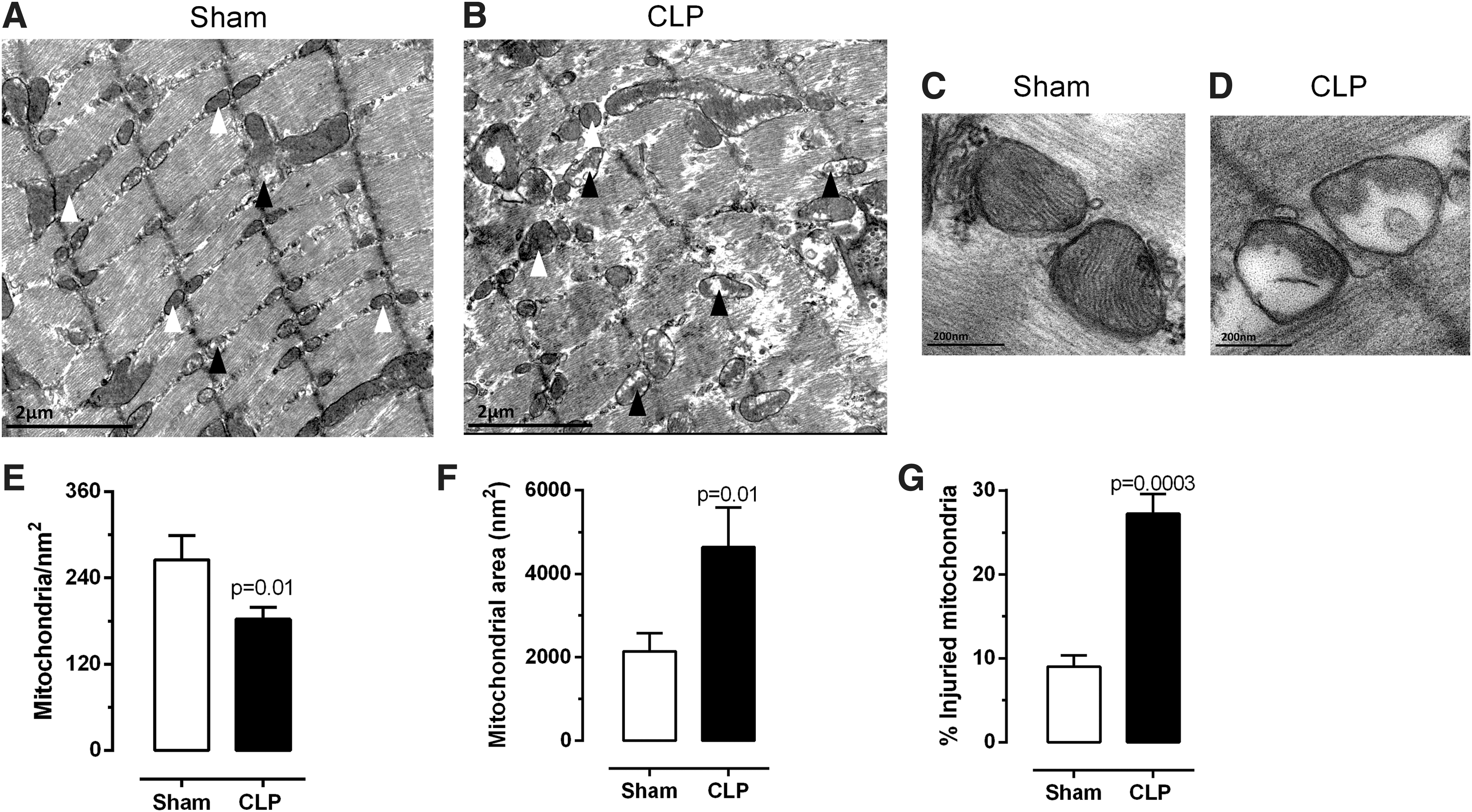
Mitochondrial quantification in the diaphragm. TEM photomicrographs of sham (
As shown in Figure 4, the physiology of mitochondria from the permeabilized diaphragms of septic and control mice was measured by HRR (Fig. 4A). Basal rates of respiration, considering complex I substrate and oxygen consumption coupled to ADP phosphorylation after succinate addition, did not differ between septic and control mice (Fig. 4B, C). On the other hand, the maximal respiration (in the presence of FCCP) was significantly reduced by 31% in the septic group (Fig. 4D, p = 0.04). In line with these results, mRNA expression levels of genes encoding for proteins related to complex II and III (Sdhb and Uqcrc2, respectively), in the respiratory chain, were clearly downregulated (Fig. 5).
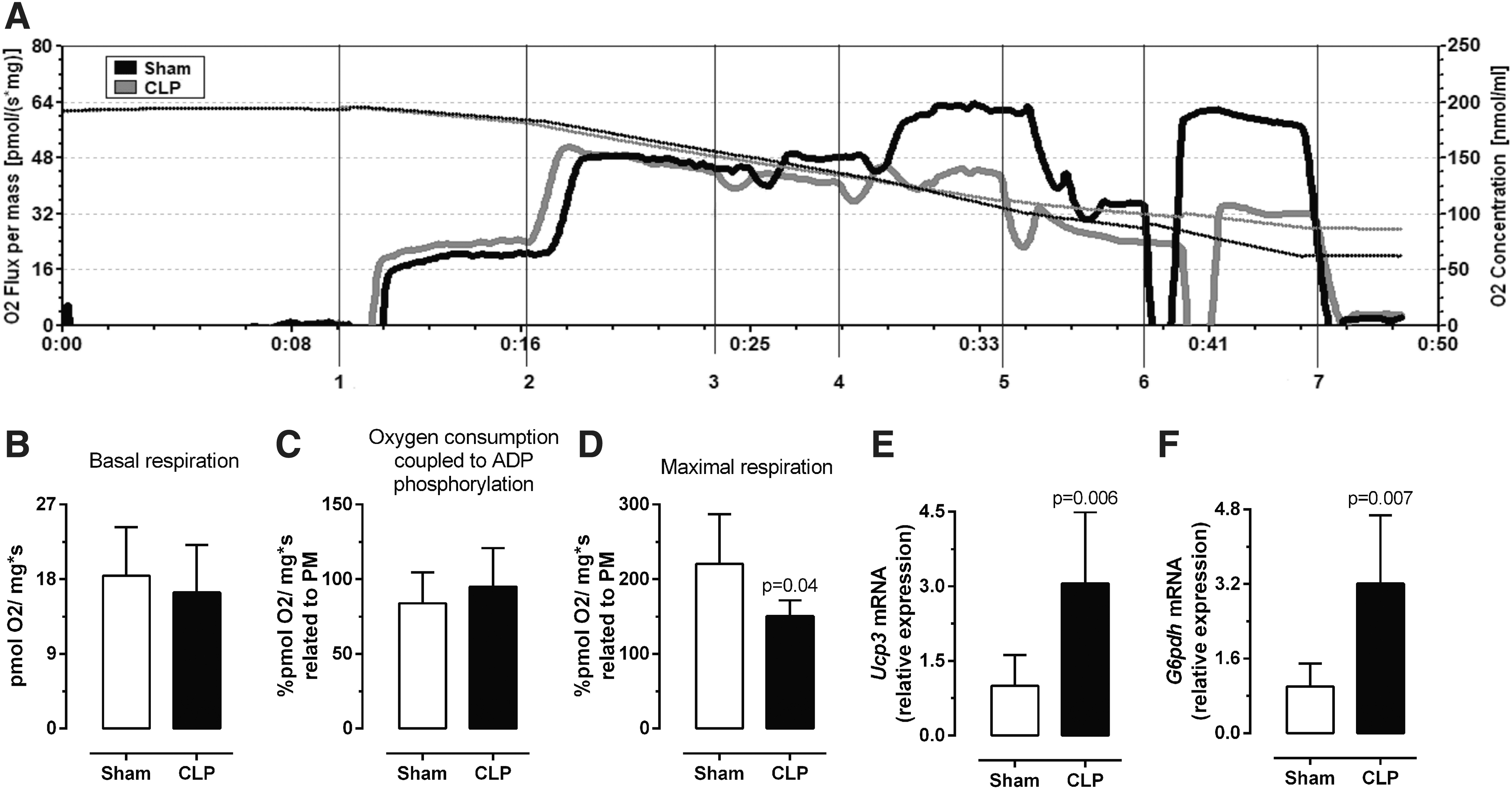
Diaphragm mitochondrial function. HRR in the diaphragm. (

Mitochondrial respiratory chain. Expression of complex I (Ndufb8—
The decrease in maximal respiration was not associated with a significant difference in oxygen consumption coupled to ADP phosphorylation. This could be related to a change in the proton gradient. As proton leaking is commonly associated to uncoupling protein activity, we investigated the expression of the premier uncoupling protein in skeletal muscle UCP3 (Ucp3). In addition, Ucp3 mRNA expression in the diaphragm was increased three-fold in the septic mice when compared with controls (Fig. 4E, p = 0.006).
The increase in UCP3 would suggest increased uncoupling, not observed in the HHR. UCP3 is also related to reactive oxygen species (ROS) defense (22). Thus, we suggest that the impact on UCP3 could be linked to ROS. In view of this possibility, we analyzed another gene related to oxidative phosphorylation and ROS, G6pdh, and found a significant increase in G6pdh mRNA expression in the septic diaphragm (Fig. 4F, p = 0.007).
Sepsis modulates ROS defenses in the diaphragm
To determine whether sepsis alters the expression of genes involved in defending the cell against ROS, the mRNA expression of soluble Sod1, Sod2, Sod3, Gpx1, and Cat was analyzed. Although Sod1 expression was unchanged (Fig. 6A), the expression levels of Sod3, Gpx1, and Cat were significantly decreased in the diaphragm of septic mice (Fig. 6C–E, p = 0.02). In contrast, Sod2 expression was three-fold higher in septic animals (Fig. 6B, p = 0.0015).
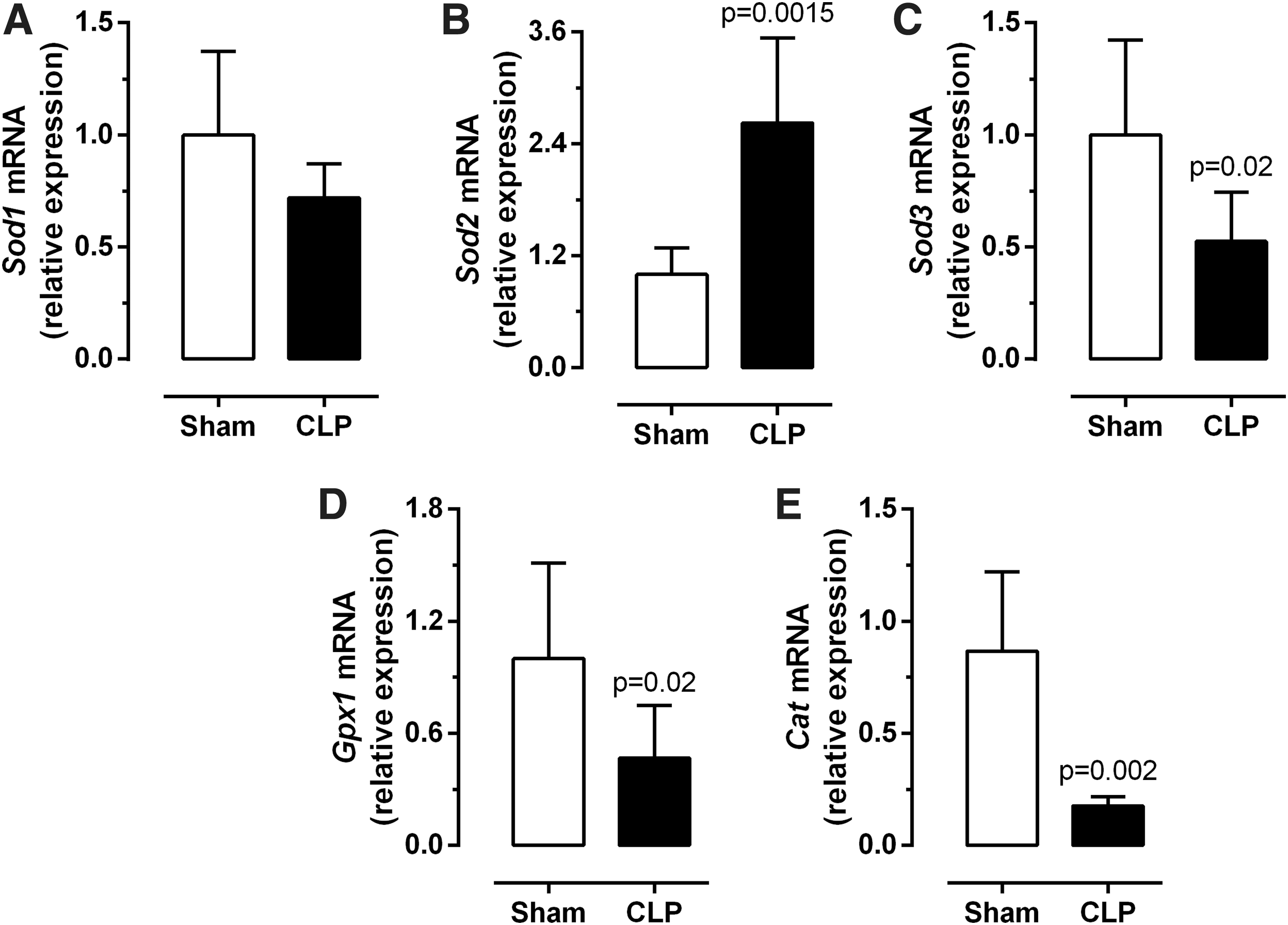
Diaphragm reactive oxygen species defense and generation. Diaphragm Sod1 (
TH replacement therapy improves mitochondrial quality
To evaluate whether TH administration could affect sepsis outcome and improve mitochondrial physiology, we investigate the survival and the sepsis severity (Table 2). After TH treatment, the percentage of animals with no signs of distress was 33.3% in the CLP+TH group compared with 8.7% in the CLP group. The comparison between the animals with no signs of distress (score 0–1) with moderate sepsis (score 4–7) in the CLP and CLP+TH groups did not reach significance (p-value of 0.1). TH treatment did not change the percentage of animal survival (CLP 79% and CLP+TH 75%, Table 2).
Sepsis Survival Rate and Severity
CLP, cecal ligation and puncture; TH, thyroid hormone.
T3 responsiveness was investigated by determining the expression of Tnni2, Klf9, and Hr, a positively regulated gene. CLP significantly decreased Tnni2 and Hr mRNA expression in the diaphragm compared with sham-treated mice. However, Tnni2 expression in the CLP+TH was not significantly different compared with sham or CLP animals. It is important to notice that Klf9 and Hr expression increased in the CLP+TH group compared with controls (Supplementary Fig. S1).
Next, we investigated the Ppargc1a mRNA expression levels in the diaphragm, the total mitochondrial number, and the percentage of injured mitochondria (Fig. 7). The CLP-induced decrease of Ppargc1a expression is not different in TH-treated mice versus untreated mice (Fig. 7D). However, the impact of sepsis on the total number of mitochondria and percentage of healthy organelles improved after TH treatment (Fig. 7A–C, E, F). In addition, we investigated whether TH treatment would improve complex II and III expression, which was downregulated by sepsis. TH treatment in CLP mice did not change Sdhb (complex II) and Uqcrc2 (complex III) mRNA expression compared with CLP mice and the sham group (Fig. 7G, H).
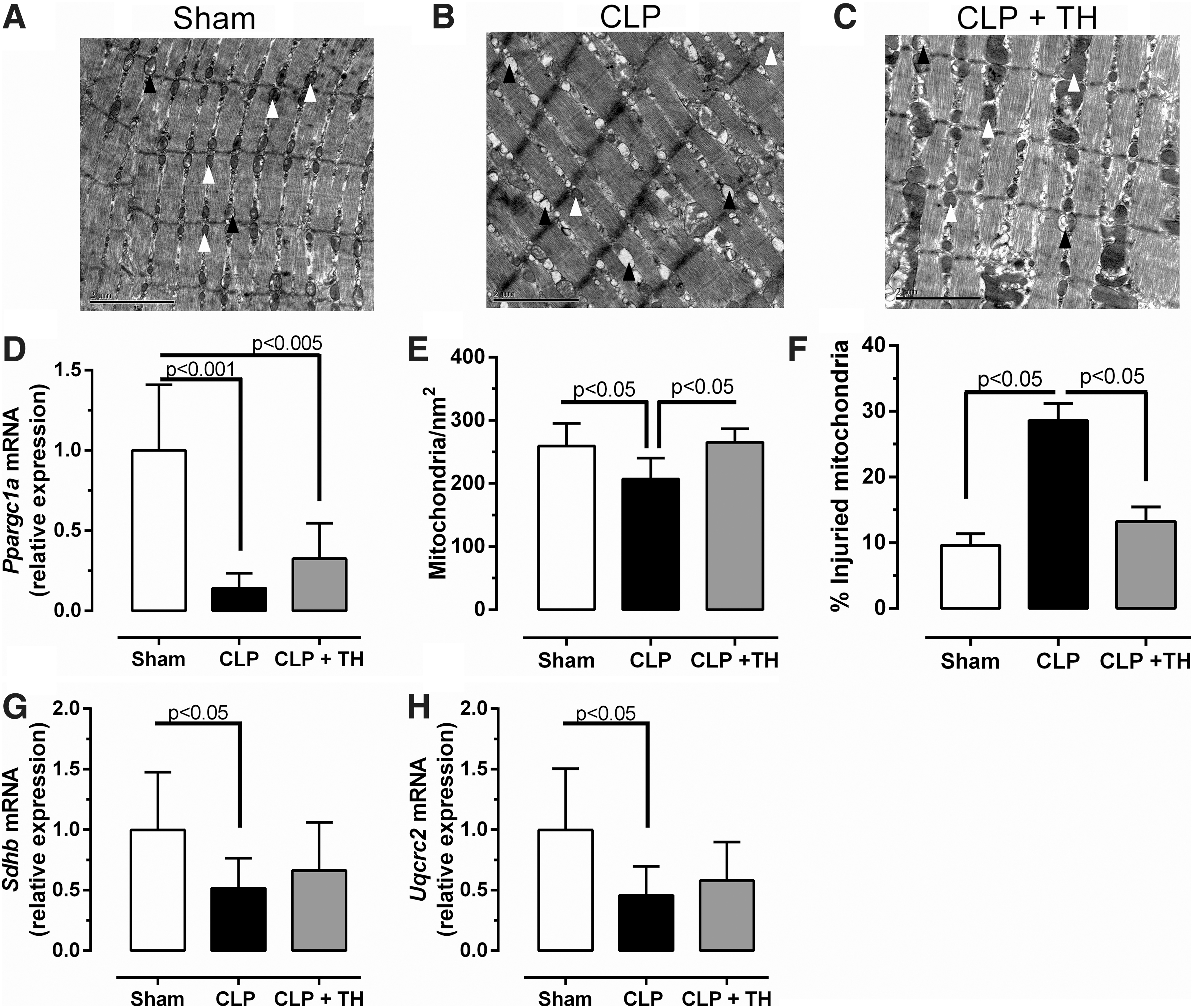
TH replacement improves mitochondrial parameters. TEM photomicrographs of sham (
Discussion
This study demonstrates that CLP-induced sepsis promotes the development of NTIS since we observed a decrease in T4 serum levels and liver Dio1 expression, hallmarks of NTIS, in our murine model (Fig. 1). In addition, CLP-induced sepsis impairs TH signaling in the diaphragm (Fig. 2 and Supplementary Fig. S1). The results also demonstrate a decrease in mitochondrial function and an imbalance in the ROS defense system in the diaphragms of septic mice (Figs. 3–6). TH treatment in CLP mice increased the expression of Hr and Klf9, T3 responsive genes that are positively regulated (Supplementary Fig. S1), and improved mitochondrial parameters (Fig. 7).
Taken together, the results suggest that the decrease in TH responsiveness in the diaphragm impairs mitochondrial function, and it reduces oxidative phosphorylation capacity and ROS defense. It is plausible that both factors are related to diaphragm weakness, which is often present in critically ill septic patients.
Normal muscle TH levels are necessary for stimulating the function and activity of skeletal muscle mitochondria (14). For example, TH has been implicated in the stimulation of oxidative phosphorylation, mitochondrial biogenesis fission, and mitophagy (23). The reduction in total plasma T4 levels and the observed decreases in Thra1, Thrb, Slc16a10 (MCT10), and Slc16a2 (MCT8) expression levels suggest that the TH availability of the diaphragm is reduced. Further, it is tempting to speculate that the observed decrease in TH availability could perturb and/or impair mitochondrial function in the diaphragm.
T3 treatment did not improve mechanical ventilation parameters in patients admitted to the ICU with NTIS (24). However, our data demonstrate improvement of mitochondrial parameters after treatment with a combination of T4 and T3 starting at the beginning of sepsis (Fig. 7). This is in contrast to the study of Bello et al. (24), who started treatment when respiratory dysfunction was already established. However, it is important to point out that our survival data do not suggest an improvement in sepsis outcomes (CLP 75%, CLP+TH 79%; Table 2). Thus, TH treatment can improve parameters associated with diaphragm physiology, but it may not improve survival in septic patients.
The oxidation of glucose to obtain energy leads to pyruvate formation in the cytoplasm, which is completely oxidized inside the mitochondria via the citric acid cycle. Concomitantly, nicotinamide adenine dinucleotide and flavin adenine dinucleotide are reduced but the electron transfer chain promotes their re-oxidation. As a consequence, these redox processes provide energy, in the form of a proton gradient across the inner mitochondrial membrane, which defines the mitochondrial membrane potential. Thus, ATP synthesis is coupled to the return of protons to the mitochondrial matrix through the FoF1-ATPase enzyme, which then uses the proton motive force to synthesize ATP (25).
The observed decrease in the number of mitochondria (Fig. 3A–E) is probably a consequence of increased mitochondrial damage (Fig. 3G) and both phenomena may lead to decreased maximal respiration (Fig. 4D). To better understand the mechanism involved in the decreased maximal phosphorylation, we investigated the genes associated with the four complexes of the electron transfer chain. There was a downregulation in Sdhb (complex II) and Uqcrc2 (complex III) expression in the diaphragms of septic mice. Complexes II and III are critical for maintaining electron flux in the respiratory chain and oxygen consumption, and the observed decrease in gene expression suggests that sepsis impairs diaphragm mitochondria physiology. TH treatment improved Sdhb and Uqcrc2 expression, suggesting that sepsis may affect mitochondrial respiration via reduction of TH signaling.
Mitochondrial uncoupling also decreases the proton gradient across the inner mitochondrial membrane. The premier uncoupling protein in skeletal muscle is UCP3. The observed increase in UCP3 expression in the skeletal muscle of septic animals is in agreement with previous studies (26,27). The role of the UCP3 uncoupling protein is controversial in the literature (28 –30). Most of the authors consider UCP3 to be a mitochondria uncoupling protein with low activity (22). However, it is possible that increased UCP3 levels are reducing mitochondrial coupling, which can also influence the oxidative phosphorylation capacity. The decrease in maximal phosphorylation observed in the CLP group is not likely to be only associated with the change in skeletal muscle UCP3 expression. Previous studies with septic Ucp3(−/−) mice showed that these mice did not present any differences in the mitochondrial membrane potential, when compared with septic wild-type mice, and the data presented here do not show differences in oxygen consumption coupled to ADP phosphorylation between groups (Fig. 4C) (22,31). Previous studies have also suggested that the upregulation of UCP3 in muscle is part of a defense mechanism employed to protect the cell against oxidative damage (32,33). To further investigate mitochondrial ROS generation and defense recruitment, we measured G6pdh expression, which is linked to increased mitochondrial ROS (34), and the mitochondrial superoxide dismutase (Sod2). In addition to the increase in Ucp3 expression, there was also an increase in G6pdh and Sod2 expression, and a decrease in Sod1, Sod3, Gpx1, and Cat expression in the diaphragms of septic mice. The upregulation of Ucp3, G6pdh, and Sod2 suggests that there is an increase in mitochondrial ROS production. It should also be noted that the downregulation of genes encoding for cytoplasmic antioxidant enzymes (Sod3, catalase, and glutathione peroxidase) could be related to the observed increase in cellular damage susceptibility.
It has been demonstrated that TH augments the expression of Ppargc1a and that the inflammatory response inhibits Ppargc1a expression (35 –37). The gene product, PGC1α, is primarily responsible for mitochondrial biogenesis and ROS defense (38 –40). The reduction in Ppargc1a expression in the diaphragm of septic animals could be due to increased proinflammatory cytokine expression or reduced TH action. We previously showed that stimulation of a muscle cell line with a bacterial endotoxin decreased mitochondrial function, which was associated with reduced Dio2 expression (15). Unfortunately, we could not measure diaphragm TH levels and, therefore, could not demonstrate a causal role for decreased TH concentrations in the Ppargc1a decrease in the diaphragm. Since PGC1α is capable of stimulating Sod1, Sod2, Sod3, Cat, and Gpx1 expression (37,41), the observed decrease in Sod3, Gpx1, and Cat expression is consistent with the observed decrease in Ppargc1a expression, and might be an indirect effect of TH. The increase in Sod2 expression could also be a compensatory mechanism used to protect cells from mitochondrial ROS-mediated damage.
This study demonstrates that acute sepsis promotes the development of NTIS and decreases the TH availability in the mouse diaphragm. This was accompanied by an imbalance in the antioxidant defense system, which appears to have severely impacted the mitochondrial physiology in the diaphragm. In conclusion, the results provide evidence that maintenance of the mitochondrial physiology in the diaphragm during sepsis is associated with TH response.
Footnotes
Acknowledgments
The authors thank the Biology Institute from the Fluminense Federal University for allowing them to use the transmission electron microscope (JEM 1011), and they acknowledge technical support from the biologist Alan Moraes, during the image acquisition. They are also very grateful to Juliana Pena Gonçalvez and Hannaily Ribeiro, for basic research support.
Author Disclosure Statement
The authors declare that there are no conflicts of interest.
Funding Information
This research was supported by the Brazilian Society of Endocrine and Metabolism—Thyroid department (SBEM), Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ, CNE 2015 E26/203.190/2015 and CNE E26/203.095/2016), Conselho Nacional de Pesquisa e Desenvolvimento (CNPq, 304667/2016-1), and Coordenação de Aperfeiçoamento Pessoal de Nível Superior (CAPES, 01). F.F.B. was a recipient of The Domestic Post Doctorate Program (PNPD, CAPES). A.T.S. was a recipient of the PhD program (CNPq).
Supplementary Material
Supplementary Figure S1