
Abstract
Background:
GLIS3 (GLI-Similar protein 3) is a transcription factor involved in several cellular processes. Homozygous mutations in the GLIS3 gene have been typically associated with neonatal diabetes and congenital hypothyroidism (CH) in a syndrome called NDH. NDH patients present developmental abnormalities including endocrine pancreas defects and a spectrum of thyroid abnormalities, mainly including thyroid dysgenesis (TD). The mouse models revealed a key role of Glis3 in pancreatic islets but not in early thyroid development, as Glis3 was described to retain a role in regulating thyroid hormone synthesis downstream the thyrotropin (TSH)/TSHR signaling pathway and in postnatal follicle proliferation. Hence, in this study, we have been taking advantage of the zebrafish model to gain insights on the Glis3 activity during thyroid organogenesis.
Methods:
Transient glis3-knockdown zebrafish embryos (called glis3 morphants) were generated by the microinjection of specific glis3 morpholinos at one- to two-cell stage to analyze the thyroid phenotype in vivo. Several additional analyses (in situ hybridization, immunohistochemistry, and pharmacological treatments) were performed for further molecular characterization.
Results:
The analysis of thyroid embryonic development revealed that Glis3 is involved in early steps of thyroid specification. glis3 morphants exhibited a reduced expression of the early transcription factors nkx2.4 and pax2a at the thyroid primordium level, which is not caused by changes in proliferation or apoptosis of the pharyngeal endoderm. As a result, the differentiated thyroid tissue in morphants appeared reduced in size with decreased expression of tg and slc5a5, a low number of thyroxine (T4)-producing follicles, associated with an elevation of tshba (homologous of the human TSHβ), thus resembling the clinical and biochemical manifestations of patients with TD. Interestingly, glis3 morphants have pancreatic β-cell defects, but not liver defects. In vitro and in vivo data also demonstrated that Glis3 is an effector of the Sonic Hedgehog (SHH) pathway. Molecular and pharmacological inhibition of SHH reproduced the thyroid defects observed in glis3 morphant.
Conclusions:
Our results demonstrate that glis3, within the SHH pathway, appears to determine the number of endodermal cells committed to a thyroid fate. This is the first evidence of the involvement of Glis3 in TD, thereby expanding the understanding of the genetic basis of thyroid development and CH.
Introduction
Several key steps of thyroid development have been understood owing to the seminal work of several groups that identified the set of thyroid transcription factors that define the thyroid precursor cells as well as differentiated thyrocytes, but the mechanism underlying the specification of the thyroid anlage from the foregut endoderm is currently unknown (1 –3).
The expression of NKX2–1, PAX8, FOXE1, and HHEX is known to be required for adequate thyroid development and function in several models, including mice and zebrafish, and mutations affecting these genes have been associated with congenital hypothyroidism (CH), one of the most common congenital endocrine diseases and the most prevalent preventable form of mental retardation (1 –6).
Recent findings indicate that the transcription factor GLI-Similar protein 3 (GLIS3) is a candidate gene for CH, although its role in thyroid pathophysiology remains largely unexplored. Homozygous and compound heterozygous mutations in GLIS3 have been associated with a rare syndrome, referred to as NDH, characterized by neonatal diabetes (ND) and CH, and several additional features, including renal, hepatic, and skeletal abnormalities (6 –8). All NDH cases have low thyroxine (T4) and high thyrotropin (TSH) levels; at the level of the thyroid, they are associated with a broad spectrum of clinical manifestations ranging from dysgenetic defects (from athyreosis to hypoplasia) to functional defects with an in situ thyroid gland. Moreover, we identified a number of heterozygous GLIS3 missense substitutions in a large cohort of patients with variable forms of CH (9). Interestingly, all the GLIS3 variants are associated with other rare mutations in genes involved in CH pathology, thus supporting the concept of a frequent oligogenic origin of CH (5,6,9).
GLIS3 is a component of Krüppel-like zinc-finger nuclear transcriptional regulators, which share a highly conservation of five C2-H2-type zinc-finger motifs (10). GLIS3 plays critical roles in development and function of several organs, controlling gene transcription through the binding with GLIS3-binding sites (G3BS) present in the regulatory regions of target genes. In humans, GLIS3 is expressed in a tissue-specific manner, with abundant expression in thyroid, pancreas, and kidney. The GLIS3 gene generates several transcripts, some of which might have variable functional activities (10 –12).
Up to now, the functional role of Glis3 has been extensively studied in mice. These monogenic diabetes models revealed that Glis3 is a fundamental player for endocrine pancreas differentiation from the endoderm, the definition of the β-cell mass, and that sustained postnatal expression is required for adequate insulin secretion and glucose metabolism (10,13 –16). In this context, several findings support the concept that Glis3 is one of the transcription factors belonging to the Sonic Hedgehog (SHH) signaling pathway. In particular, the binding with the Suppressor of Fused (SUFU), a negative regulator of SHH, accounts for the stability and transactivation activities of GLIS3 (13 –20).
As far as the thyroid gland is concerned, evidence from Glis3 knockout (KO) mice indicated that Glis3 is essential for thyroid hormone (TH) biosynthesis and postnatal thyroid gland proliferation, acting downstream of the TSH/TSHR system (10,21). Although the size of the thyroid follicles is smaller in Glis3-KO pups in the first week after birth, the thyroid morphology and serum TH levels are similar compared with wild-type mice, suggesting that the development of CH in this particular Glis3-KO mouse is related to functional rather than developmental defects. These findings are in contrast to those observed in the NDH patients, in which thyroid dysgenesis (TD) is a common feature (7 –9,22).
In this study, we took advantage of the zebrafish model to characterize the role of Glis3 during early thyroid development. Zebrafish has emerged as a suitable model system in thyroid research, thanks to the high conservation of the molecular mechanisms involved in thyroid organogenesis, TH transport and metabolism, as well as TH action in target tissues (23 –27). We previously described that the glis3 transcript is detectable early during zebrafish embryonic development, and its signal depicts the foregut endoderm pouches, the tissue that will give rise to endocrine organs (e.g., thyroid and pancreas) (28). In the present work, we describe the effects of glis3 knockdown (KD) in zebrafish. The multiple genetic and pharmacological manipulations demonstrate that glis3, within the SHH pathway, is a critical factor for the correct specification of thyroid primordium, likely regulating the number of endodermal cells committed to thyroid fate.
Materials and Methods
Ethical statement
Current Italian national rules: no approval needs to be given for research on zebrafish embryos. Fish were maintained and raised according to EU regulations on laboratory animals (Directive 2010/63/EU).
Zebrafish line and maintenance
Wild-type adults (AB strain) were obtained from the Wilson laboratory, University College of London, United Kingdom, and maintained in a flow-through system in charcoal-filtered tap water at a constant temperature (28°C ± 1°C), with a photoperiod of 14:10 (light:dark). Zebrafish embryos, obtained from natural spawning, were raised and maintained according to established techniques (29,30) and staged according to morphological criteria (31). Beginning from 24 hours postfertilization (hpf), the embryos were cultured in fish water containing 0.002% of 1-phenyl-2-thiourea (PTU; SIGMA) to prevent pigmentation and 0.01% methylene blue to prevent fungal growth. Such a low dose of PTU (0.002%) does not affect TH synthesis, and the T4 concentrations are similar in treated and untreated fish (32). The wild-type (WT) AB strain zebrafish line was used.
Probes preparation and whole-mount in situ hybridization
For glis3 (ENSDART00000142833.2) antisense digoxigenin (DIG) probe synthesis, a 1024 bp template was generated by reverse-transcription (RT)-polymerase chain reaction (PCR) on total RNA extracted from pools of embryos at 26–48 hpf. The primers used for probe amplification are the following: FW: 5′ TGGGAAAGGCTGTAACCTGA 3′ and REV: 5′ GGACACCTCAAACTGAAGCG 3′ (28,33). The PCR products were cloned into the pCRII-TOPO vector (Invitrogen, Carlsbad, CA), and the cDNA-containing plasmids were enzymatically linearized and in vitro transcribed to antisense DIG riboprobes using T7 or SP6 with RNA polymerase (Roche).
The riboprobes against nkx2.4, pax2a, tg, slc5a5 (33), tshba (34), shha (35), foxa2 (36), isl1 (37), foxa3 (38), ptf1a (39), ins2 (40) mRNAs were used to evaluate thyroid, endoderm, liver, and pancreas development and function. Whole-mount in situ hybridization (WISH) experiments were performed according to Thisse and Thisse (41). Depending on the experiment, the DIG probes were detected using anti-DIG-alkaline phosphatase (AP) and nitro-blue tetrazolium/5-bromo-4-chloro-3'-indolyphosphate (NBT/BCIP) or Fast Blue. For the analysis of thyroid volume quantification (NBT/BCIP), the embryos were flat mounted, and the number of positive cells was calculated by dividing the area of positive staining by the average area of a single cell using the ImageJ software (
Loss-of-function analysis
The embryos were injected with a specific Morpholino (MO), a specific antisense oligonucleotide to transiently knockdown the gene function.
The specific glis3 morpholino (GLIS3MO_SPL: 5′ TTCTTGTTTTTACCTTTCATACCGC 3′) was designed by Gene Tools (LLC, Philomath, OR) to recognize the splice donor site of the exon 2–intron 2 boundary of the pre-mRNA. The negative controls were generated by the injection of standard control morpholino (std-CtrlMO: 5′ CCTCTTACCTCAGTTACAATTTATA 3′) that targets the human β-globin gene. Morpholinos were dissolved in Danieau's buffer (58 mM NaCl; 0.7 mM KCl; 0.4 mM MgSO4 · H2O; 0.6 mM Ca(NO3)2; 5 mM HEPES, pH 7.2). Embryos were microinjected at the one- to two-cell stage and Rodamine dextran (Molecular Probes) was usually coinjected as a tracer.
The GLIS3MO_SPL was validated by RT and quantitative real-time (qRT)-PCR. Total RNA was extracted from injected and control embryos at 1 and 2 days post-fertilization (dpf) using TRIzol™ (Thermo Fisher Scientific). The reaction was carried out following the protocol of GoScript™ Reverse Transcription System (Promega). The cDNAs were then subjected to PCR amplification using GoTaq polymerase (Promega) following the manufacturer's instructions. qRT-PCR was performed by ABI PRISM™ 7900HT Fast Real-Time PCR System using SYBR Green Master Mix (Invitrogen). A second glis3 splicing morpholino (43) (GLIS3_MO2: 5′ ACCTGCTGCAAGAGATCAGTTAAAA 3′) has been used to confirm the results obtained with the GLIS3MO_SPL. The shha_MO (44) (5′ CAGCACTCTCGTCAAAAGCCGCATT 3′) was used to investigate the relationship between glis3 and Shh.
Rescue experiments
Full-length zebrafish wild-type glis3 cDNA was cloned into the pcs2+ vector (Invitrogen) using the following primers: glis3CDS_FW: 5′ ATGGACATGAATGGGAAAG 3′ and glis3CDS_REV: 5′ TCAGCCTTCAGTGAACACACA 3′.
The mutated form of glis3 mRNA was generated introducing a point mutation GGA>TGA at position 1223 bp that causes the formation of a premature stop codon upstream of the zinc-finger functional domain. Both constructs (WT and mutant [MUT]) were linearized and in vitro transcribed using the mMessage mMachine Sp6 kit (Thermo Fisher Scientific) following the manufacturer's instructions. The GFP mRNA was used as control for glis3 mRNAs microinjection, which has no targets in zebrafish.
Immunofluorescence
The qualitative analysis of TH production was performed by immunofluorescence according to the standard procedures using a rabbit anti-T4 BSA serum (1:1000; ICN Biochemicals) and the AlexaFluor 555 anti-rabbit IgG as secondary antibody (1:500, Life Technologies) (23,36).
To test proliferation and apoptosis, after WISH with nkx2.4-DIG riboprobes and Fast Blue staining, embryos were stripped and rinsed in PBS. Anti-phospho-histone H3 (PH3) and anti-active caspase-3 (AC-3) antibodies (1:250; Cell Signaling) were used as primary antibodies, followed by incubation with an antirabbit IgG secondary antibody/AlexaFluor 488 (1:500; Life Technologies) (36).
Immunocytochemistry
The entire CDS of the zebrafish sufu and glis3 were amplified and cloned into the pEGFP-C3 and pcDNA4-Myc/His, respectively. HeLa cells were transfected with 1 μg of pEGFP_sufu or pcDNA4-Myc/His-glis3 plasmids (0.5 μg each in co-transfection assay). After 24 hours, the transfected cells were fixed with 4% paraformaldehyde (Sigma–Aldrich) and permeabilized with 0.1% Triton X-100 (Sigma–Aldrich). For the detection of the glis3 construct, the cells were incubated overnight with α-c-Myc primary antibody (1:100; Life Technologies) and then detected with secondary antibody Alexa Fluor 555 goat anti-mouse IgG (1:500; Life Technologies). The fixed cells were mounted with Slow-Fade Gold antifade reagent with DAPI (Life Technologies) and acquired using the Nikon Eclipse Ti confocal microscope equipped with a 60 × oil immersion objective.
Co-immunoprecipitation
HeLa cells were transfected as previously described, and after 24 hours, the total protein content was extracted using Cell Lysis Buffer (20 mM Tris–HCl, pH 7.5, 150 mM NaCl, 1 mM EDTA, 1 mM EGTA, 1% Triton X-100, protease inhibitor cocktail [Roche]) and sonication. Protein lysates were quantified by BCA protein assay (Thermo Fisher Scientific), and 300 μg of protein sample was incubated for 45 minutes at room temperature with 450 μg of Dynabeads™ Protein G (Invitrogen) conjugated with 1 μg of anti-c-Myc Monoclonal Antibody (Invitrogen) or 1 μg of normal mouse IgG (Santa Cruz Biotechnology). Magnetic beads were washed four times in PBS with 0.02% Tween-20 and resuspended in 20 μL 2 × sample loading buffer (50 mM Tris–HCL, pH 6.8, 2% SDS, 0.1% bromophenol blue, 10% glycerol) and 200 mM DTT (Sigma–Aldrich). Immunocomplexes and input fractions were examined by Western blot using 4–12% NuPAGE Bis-Tris pre-cast polyacrylamide gels (Thermo Fisher Scientific). Immunoblots were performed with mouse c-Myc antibodies (1:500; Life Technologies) or mouse GFP antibody (1:1000; Origene) diluted in 5% milk in TBS with 0.1% Tween-20 (Sigma–Aldrich).
Cyclopamine treatment
For the inhibition of the Shh pathways, from 50% epiboly stage to 26 or 48 hpf, the embryos were treated with 10 μM cyclopamine (Sigma–Aldrich) harvested in fish water (45,46); 1% DMSO (dimethyl sulfoxide) was used as a control vehicle.
Statistical analyses
All data are shown as mean ± standard deviation or percentages. The t-student test was used for the analysis of differences between groups. The p < 0.05 was considered statistically significant. All the analyses were conducted with the software package GraphPad Prism 4.0 (GraphPad, San Diego, CA).
Results
glis3 knockdown affects the specification of thyroid primordium
Knockdown experiments were performed by the microinjection of a specific glis3 morpholino (glis3MO_SPL) into zebrafish embryos (called morphants). Following the setup of glis3 gene knockdown and embryonic phenotype analysis (see the Molecular and Morphological Validation of glis3MO_SPL section in the Supplementary Data), we performed WISH to test whether glis3 plays a role in zebrafish thyroid organogenesis.
At 1 dpf, the thyroid primordium was defective with a marked reduction of nkx2.4 and pax2a expression in the glis3 morphants (Fig. 1A–C’, E–G’). The nkx2.4 was markedly reduced in the thyroid bud compared with controls in 65.6% of embryos injected with 0.3 pmol/embryo of glis3MO_SPL and undetectable in almost all the embryos injected with 0.5 pmol/embryo of glis3MO_SPL (Fig. 1D). Consistently, a reduced to absent pax2a expression was seen in 17.6% to 33.9%, respectively, of the embryos injected with 0.3 pmol/embryo MO, and the expression of pax2a was reduced to absent in the 31.2% and 68.8% of morphants injected with 0.5 pmol/embryo (Fig. 1E–H). When compared with the size in control embryos, the volume of the thyroid primordium was reduced by 50–95% in morphants (Fig. 1 Graphs D’, H’).
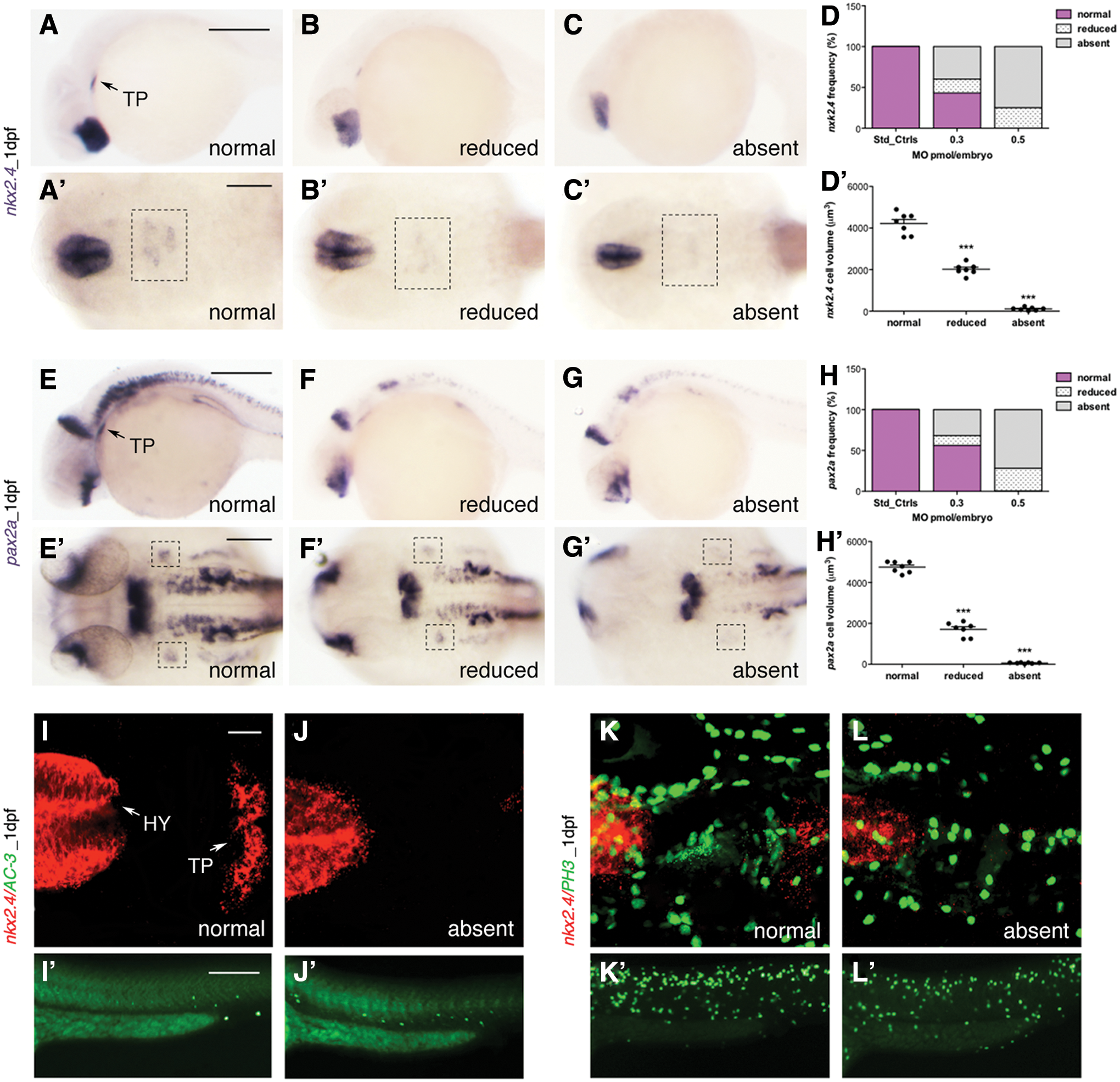
Analysis of the developing TP.
The mechanisms underlying the altered thyroid phenotype in the glis3-knockdown embryos were investigated by activated caspase-3 (AC-3) or phospho-histone H3 (PH3) immune staining as markers of apoptosis or proliferation, respectively (Fig. 1I–L). At 1 dpf, no significant differences in AC-3 (Fig 1I, J) or PH3 (Fig. 1K, L) staining were seen by confocal microscopy in the area of nkx2.4 expression in all morphants. These findings indicate that the reduced expression of the early thyroid markers is not associated with a decreased proliferation nor an increased apoptosis of the endoderm precursors committed to thyroid fate. In aggregate, these findings point to the potential role of glis3 in regulating the amount of endoderm cells that will be addressed to thyroid precursors.
glis3 morphants have TD and primary hypothyroidism
At 2 dpf, the precursors of thyroid primordium differentiate into thyroid follicles, expressing the functional markers tg and slc5a5, and proliferation along the ventral aorta by 3 dpf (26). In agreement with the findings reported above, the injection of 0.3 or 0.5 pmol/embryo MO caused a marked and dose-dependent reduction of all the examined markers. The tg transcript was reduced in 51.7% and 74.4% of morphants, respectively (Fig. 2A–C). Similarly, the slc5a5 transcript was reduced, frequently below the limit of detection, in 74% or 88% of morphants (Fig. 2D–G). At 3 dpf, the thyroid gland elongates along the midline forming the classical V-shape (Fig. 2H): the staining of tg continued to be reduced in 32% or 50% of the morphants (Fig. 2I, K). Interestingly, a significant portion of morphants exhibited a normal expression of tg, but their thyroid follicles were disorganized and ectopically displaced along the ventral aorta (Fig. 2J, K).

Analysis of development and function of thyroid gland.
The quantification of tg+ cell number confirmed the distribution of the thyroid phenotype of glis3 morphants at 2 and 3 dpf (Fig. 2L, M).
A double fluorescent WISH of early and late thyroid markers was also performed to understand the variable degree of impairment detected in the in glis3 morphants up to 60 hpf (see the Double Fluorescent Analysis of the Early and Late Thyroid Markers section in the Supplementary Data). Most of the embryos presented a significant reduction of both early transcripts (nkx2.4 and pax2a), but pax2a was generally diminished to a larger extent. However, when the expression was completely absent, no thyroid bud was present and all markers were affected.
Immune staining with an anti-T4 antibody was performed to assess the status of thyroid function at 5 dpf by comparing the control and morphants injected with 0.5 pmol/embryo. By counting individual T4-positive follicles, we observed that the glis3 morphants exhibited a significantly (p < 0.01) reduced number of follicles (n: 3 ± 1 vs. 5 ± 1) (Fig. 2N–P). Consistently, 85% of the glis3 morphants presented a significant increase (p < 0.001) in the number of the tshba (homologous of human TSHβ)-positive pituitary cells at 5 dpf (n: 13 ± 2 vs. 4 ± 1) (Fig. 2Q–S).
The low T4 production and the high tshba expression observed in glis3-knockdown larvae well recapitulate the clinical findings described in patients with TD. The thyroid phenotype in glis3-deficient embryos was faithfully confirmed by using a second glis3 splicing morpholino that had been previously validated (43) (see the Molecular and Morphological Analysis of the Alternative glis3MO2 section in the Supplementary Data).
The glis3 gene dosage regulates the number of the differentiated thyroid follicles
To confirm the specific thyroid phenotype resulting from glis3 morpholino activity, we performed mRNA rescue experiments co-injecting the glis3MO_SPL with the wild-type or mutated zebrafish glis3 transcripts. The mutant glis3 mRNA was generated introducing a point mutation GGA>TGA at position 1223, which causes the formation of a premature stop codon upstream of the ZF domain, mimicking natural human GLIS3 mutations (7).
The co-injection with the glis3MO_SPL and the glis3_WT mRNA was able to rescue the loss of expression of nkx2.4 and tg (Fig. 3D, D’) induced in the vast majority of the embryos injected with glis3MO_SPL alone (Fig. 3C, C’). Interestingly, the co-injection of the glis3MO_SPL and the glis3_MUT mRNA (Fig. 3E, E’) failed to rescue the thyroid defects of glis3 morphants and even worsened the phenotype. The thyroid was not affected after the injection of the same doses (100 pg/embryo) of WT or MUT glis3 mRNAs (Fig. 3B, B’).
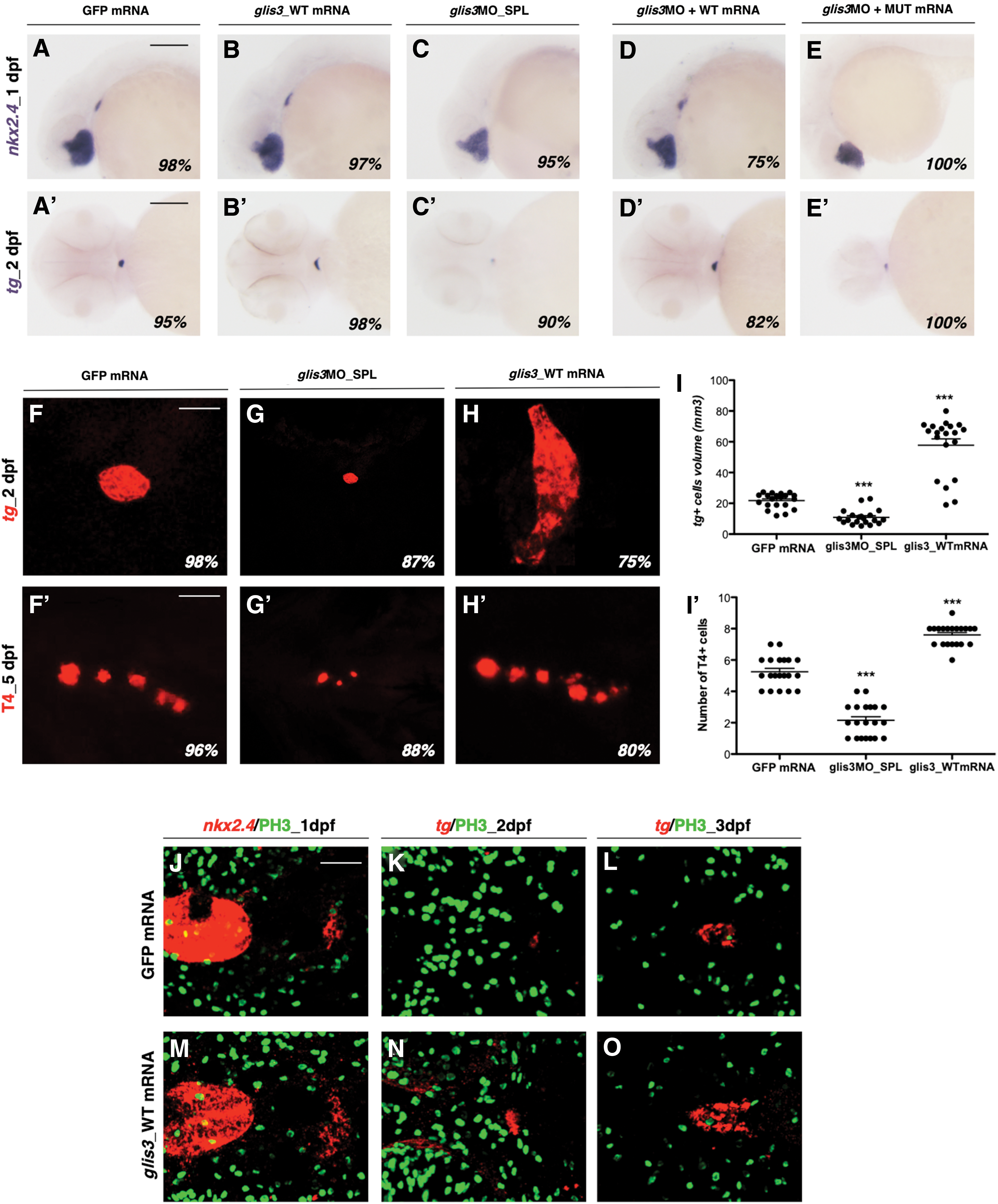
Rescue experiment.
To further define whether glis3 gene dosage contributes to the onset of thyroid defects, we performed overexpression experiments microinjecting growing doses of glis3_WT mRNA. When compared with controls at 2 dpf (Fig. 3F), the injection of 250 pg/embryo of the glis3_WT mRNA was associated with an upregulation of tg expression in 75% of embryos, and an enlargement of the thyroid anlage (+130% than controls) (Fig. 3H, I). Confirming the previous results, the thyroid was smaller in 87% of glis3 morphants compared with controls (Fig. 3G, I), and the number of T4-producing follicles was reduced by 50% (Fig. 3F’, G’). In contrast, the overexpression of glis3 led to a significant increase of the T4 production (Fig. 3H’, I’).
To test whether the increased size of the thyroid gland in the glis3-overexpressing embryos was caused by an increased proliferation in the foregut endoderm or an enlarged number of endodermal cells committed to the thyroid fate, we performed proliferation assays at different time points. Since no differences in phospho-histone H3 levels were detected in the pharyngeal floor in early and late developmental stages after the injection of 250 pg/embryo glis3_WT mRNA (Fig. 3J–O), these data also support the concept that the amount of glis3 activity is critical for the determination of thyroid primordium size.
Glis3 regulates the development of the endoderm-derived endocrine organs
To exclude a global impairment of the pharyngeal endoderm in the glis3 morphants, we analyzed the expression of foxa2, an early endoderm-specific transcription factor. When thyroid bud becomes evident (22 hpf), the endoderm pouches appeared unaffected in glis3 morphants (Fig. 4A, B).

Analysis of pharyngeal endoderm and endoderm-derived organs in glis3 morphants.
Furthermore, we investigated whether other endoderm-derived organs, such as liver and pancreas, show developmental defects in glis3 morphants. Liver development was unaltered in 100% of the glis3 morphants, as shown in the WISH analysis of the expression of the liver-specific genes isl1 and foxa3 (Fig. 4C–F). In contrast, the glis3 morphants showed endocrine pancreas defects (Fig. 4G–I). In 52% of morphants, we observed an impaired ins2 expression at 2 dpf (Fig. 4H), and in the remaining 48%, the ins2 expression was disorganized, that is, the insulin2-positive cells failed to converge on the midline to form a single islet (Fig. 4I). The WISH of the acinal specific marker ptf1a also confirmed that the pancreatic defects of morphants were restricted to the endocrine compartment (see the Analysis of the Pancreatic Acini Development in the glis3 Knockdown Embryos section in the Supplementary Data).
These results suggest that glis3 action is required for the correct development of the thyroid and the endocrine pancreas, but its downregulation does affect the formation of pharyngeal endoderm pouches and liver development.
Glis3 acts as an effector of the SHH pathway
Preliminary in vitro assays confirmed that sufu and glis3 interact even in zebrafish, since they colocalized and co-immunoprecipitated in a protein complex (see the Glis3 and sufu Zebrafish Proteins Interact In Vitro section in the Supplementary Data).
To study the relationship between glis3 and Shh, the expression of shha was analyzed in glis3 morphants by WISH. When compared with controls at 2 dpf (Fig. 5A, B), the 100% of glis3 morphants injected with 0.5 pmol/embryo presented a significant reduction of shha expression in the pharyngeal endoderm (Fig. 5C, D).
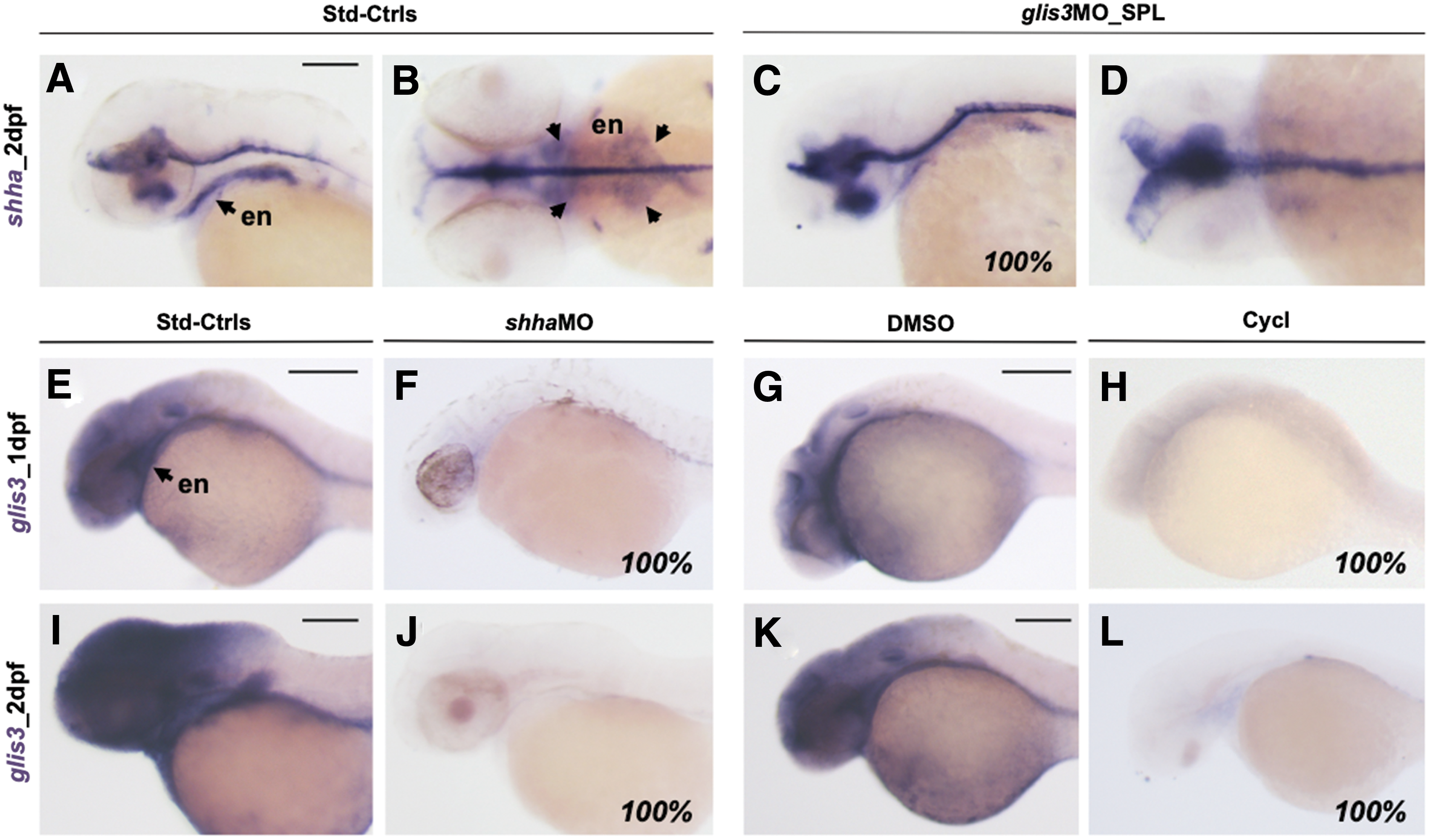
Analysis shha expression in glis3 morphants.
The expression of glis3 was previously shown to be expressed early in the pharyngeal endoderm and absent in the differentiated thyroid (28). Here, we show that the downregulation of the shh pathway by injection of shha_MO (1.25 pmol/embryo) (44) or treatment with cyclopamine (10 μM) (45,46) is associated with the lack of glis3 expression in both brain and pharyngeal endoderm (Fig. 5E–L).
To test whether Shh signaling plays a role in thyroid development, the expression of endodermal and thyroid markers was analyzed by WISH in the cyclopamine-treated embryos. In agreement with the key role of the shh pathway during endoderm formation and development, foxa2 was compromised in the cyclopamine-treated embryos (Fig. 6A, B). As a consequence of the pharmacological inhibition of the shh pathway, thyroid development was severely compromised. In fact, at 1 dpf, nkx2.4 and pax2a signals were reduced/absent at the level of thyroid primordium in the 100% of the cyclopamine-treated embryos (Fig. 6C–F), and at 2 and 3 dpf, tg expression was reduced in 68% or absent in 32% of treated embryos (Fig. 6G–L).
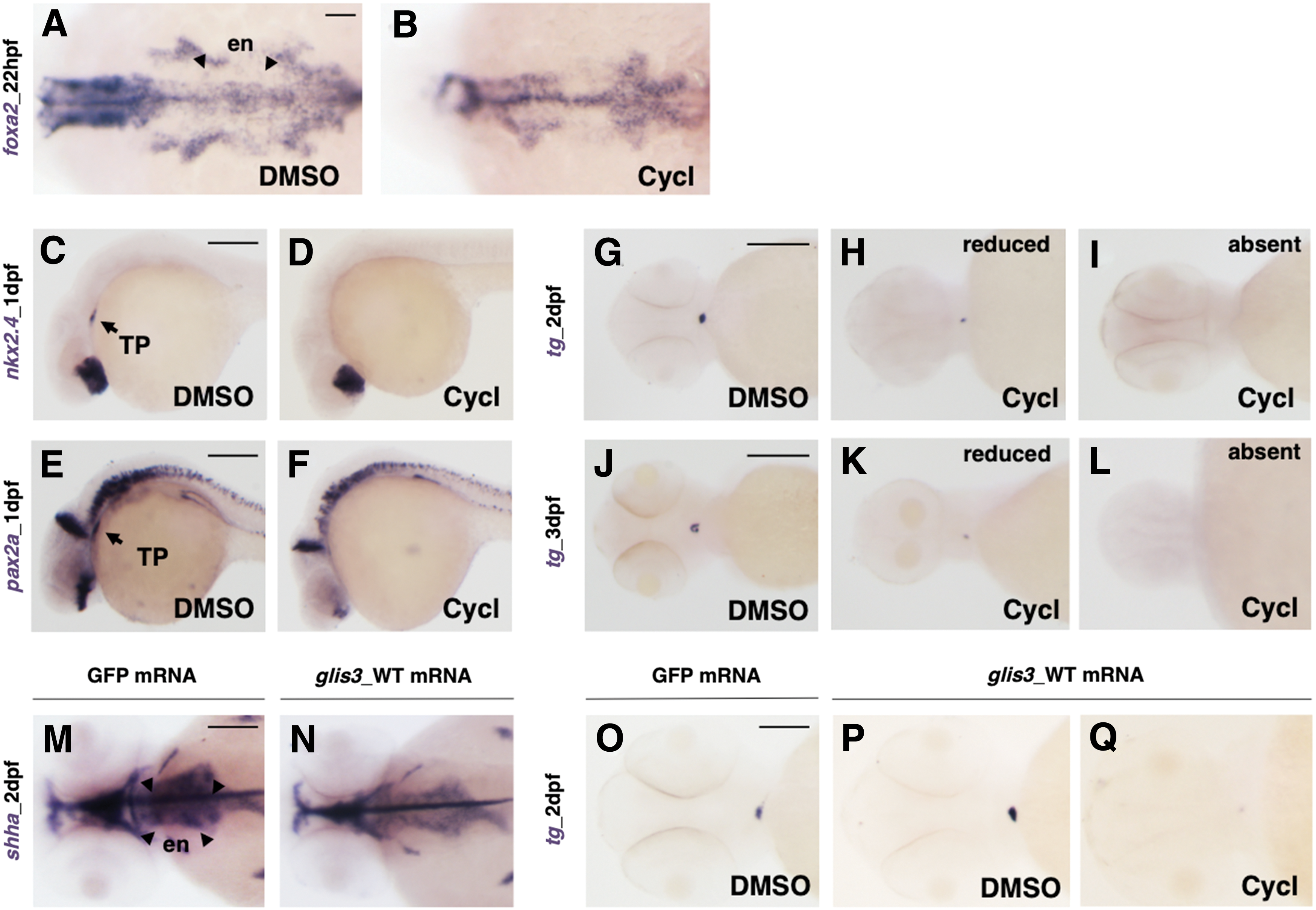
Analysis of thyroid gland development in cyclopamine-treated embryos.
The overexpression of glis3 after the injection of WT glis3 mRNA (250 pg/embryo) did not alter shha transcript levels at 2 dpf (Fig. 6M, N) but enhanced the number of endodermal precursor cells toward the thyroid fate, thus resulting in an excess of tg-expressing cells (Fig. 3). Surprisingly, cyclopamine treatment is able to completely counteract the effects of glis3 overexpression and thus negatively affects thyroid development (Fig. 6O–Q). Accordingly, glis3 requires shh signaling for its activity and is likely to be a main downstream effector of this pathway in thyroid specification.
Discussion
Here, we demonstrate that the downregulation of glis3 in zebrafish embryos leads to developmental defects in the thyroid, replicating the TD reported in several patients with GLIS3 mutations. The step-by-step analysis of thyroid organogenesis reveals that glis3 knockdown interferes with the specification of thyroid precursors from the pharyngeal endoderm, as shown by the absent or reduced expression of the early transcription factors nkx2.4 and pax2a at 1 dpf. Interestingly, at early stages of development, the glis3 knockdown appears to affect pax2a to an extent larger than nkx2.4. However, at later stages (2–3 dpf), glis3 morphants show a reduced or absent expression of tg and slc5a5, two of the key genes involved in thyroid hormone synthesis. As a consequence of these developmental defects, the glis3 morphant larvae present thyroid hypoplasia, a reduced number of T4-producing follicles, and high levels of thsba at 5 dpf, thus recapitulating the primary hypothyroidism observed in NDH patients.
Importantly, because no variations in proliferation or apoptosis were detected under glis3 knockdown in the region including the thyroid bud, our findings support the concept that glis3 action is fundamental for the quantitative regulation of the amount of endodermal precursors committed to a thyroid fate. Consistently, the recovery of the thyroid phenotype in the rescue experiment, as well as the opposite variation in the number of thyroid cells and T4 production obtained through overexpression of the glis3 mRNA, confirms that the gene dosage of glis3 is critical for correct development and size definition of the zebrafish thyroid. These effects appear independent of the tsh/tshr signal since these transcripts can be detected in zebrafish from 40 to 42 hpf (24,26), and tshba was found to be high in glis3 morphants. Previous work on zebrafish shh mutants reported an impaired pituitary gland and absence of tshba-positive cells (47 –49). This discrepant effect on pituitary markers may perhaps be explained by the involvement of different downstream effectors of the Shh pathway in pituitary and thyroid development: Glis3 may have a more relevant role in thyroid (and endocrine pancreas) development, whereas other Gli-like factors may exert a main developmental role in the pituitary.
Interestingly, the pharyngeal endoderm appeared unaffected in glis3 morphants (Fig. 4), thus excluding that the observed thyroid defects are a consequence of aberrant endoderm formation. Accordingly, we investigated whether other organs that arise from the endoderm (e.g., liver, exocrine and endocrine pancreas) display some developmental defects. Only the endocrine pancreas development appears to be affected by glis3 knockdown, as glis3 morphants display a reduced differentiation of the ins2-positive pancreatic β-cells that frequently fail to migrate and localize in a disorganized shape. This phenotype displays similarities with the previously reported alterations of endocrine pancreas differentiation in Glis3-deficient conditions (10,18,21,50).
The decrease of the functional thyroid markers during glis3 knockdown at 2–3 dpf could be the consequence of direct or indirect effects. Indeed, PAX8 and NKX2.1 were shown to regulate the transcription of thyroidal genes involved in thyroid hormone synthesis through the direct binding of their promoter regions (1,2,21); however, CHIP-sequencing of mice thyroid tissues identified Pax8, Pds, and Slc5a5 as direct transcriptional targets of Glis3, thus pointing to a direct regulatory role of Glis3 on thyroid function. In apparent contrast with our findings, transcriptome analysis showed that the expression of Pax8 and Nkx2.1 was unchanged in Glis3KO mice 1 month after birth (21). This discrepancy might be explained by the existence of compensatory mechanisms (e.g., other Gli-like activities) that are able to rescue, at least partially, Glis3 deficiency in the murine thyroid. In partial agreement with our findings, Glis3KO pups tend to have smaller thyroid glands than wild-type littermates (21). In aggregate, these findings support the concept of a dual role for glis3 on thyroid gland activity by promoting its commitment from the pharyngeal endoderm and, later on, the function/proliferation of thyroid cells. Indeed, Glis3 was previously reported to have a dual role also in pancreatic function: (a) by regulating the number of endocrine progenitors that will differentiate into β-cells during embryonic growth and (b) by postnatally mediating insulin secretion and β-cell proliferation (13 –19,51).
The post-translational stability and transcriptional activity of GLIS3 were reported to be regulated by the binding with the SUFU, a negative regulator of the SHH signaling (20,52,53). In the absence of SHH, SUFU would anchor GLIS3 in the cytoplasm preventing its interaction with the ubiquitin ligase CUL3 and ITCH, thus protecting GLIS3 from proteasome degradation (20,54). However, the GLIS3-SUFU complex was also reported to translocate into the nucleus, where it recruits and interacts with co-repressors, suppressing the transcription of some target genes (e.g., Ins2) (13,17,20,55). Through in vitro experiments, we then found that the zebrafish proteins glis3 and sufu co-localized in the nuclei of transfected cells and precipitate together in a co-IP assay, thus confirming that the interplay between glis3 and shh signaling occurs also in zebrafish. Subsequently, we observed that the shha signal is significantly reduced in the pharyngeal endoderm of glis3 morphants, and coherently, the expression of glis3 is abolished in shh-deficient embryos (shha morphants and cyclopamine-treated embryos). Altogether these data point to a relevant developmental role of the shha-glis3 pathway in the pharyngeal endoderm and suggest that the shh activity would induce the expression of glis3 in the zebrafish endoderm.
Moreover, embryos treated with the shha antagonist developed a thyroid phenotype characterized by reduced/absent expression of early and late markers, a phenotype similar to that described in glis3 morphants. Interestingly, cyclopamine treatment also prevents the tg upregulation induced by glis3 overexpression. Therefore, glis3 appears to require an active Shh signal for its function, likely acting as a downstream effector of the Shh pathway during zebrafish thyroid development.
In conclusion, we propose that glis3 acts as an effector of the Shh pathway with a critical role during embryonic thyroid development. In particular, our data provide significant insights into the signaling pathways involved in the induction of thyroid precursors from the undifferentiated endoderm. However, additional experiments are needed to characterize the molecular events linking the shh-glis3 pathway and the expression and action of early thyroid transcription factors. Our findings may provide an explanation for the frequent developmental thyroid defects in patients with GLIS3 mutations. Finally, zebrafish may represent an alternative in vivo model suitable for high-throughput screening of drugs able to counteract GLIS3 deficiency (50).
Footnotes
Acknowledgments
The authors wish to thank Dr. Annamaria Maraschi (Milan) for the supply of pEGFP-C3 and pcDNA4-Myc/His constructs and for help with the setting of the experimental protocols for in vitro studies.
Authors' Contributions
G.R. designed and performed the in vivo experiments and wrote the draft of the article. F.M. supervised and contributed the zebrafish studies and the article preparation. P.D. performed the in vitro experiments. L.P. conceived the study, obtained research funds, and revised and finalized the article. All authors approved the final version of the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work has been partially supported by the Ricerca Corrente funds of Istituto Auxologico Italiano, Milan, Italy (code: 05C102_2011), and by the PhD and Post-Doc programs of the University of Milan, Italy.
Supplementary Material
Supplementary Data
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6