
Abstract
Background:
Bone is an important target of thyroid hormones (THs), which require transport into target cells to exert their actions. Recently, the TH-specific monocarboxylate transporter 8 (Mct8) was reported as a regulator of bone mass in male mice. However, its global deletion leads to high 3,3′,5-L-triiodothyronine (T3) serum concentrations that may mask direct effects of Mct8-deficiency on bone. In this study, we assessed the bone cell intrinsic function of Mct8 ex vivo and in vivo using conditional Mct8-knockout lines specifically targeting osteoclast and osteoblast progenitors, as well as mature osteoblasts and osteocytes.
Materials and Methods:
Twelve-week-old male mice with a global Mct8-deficiency or a conditional Mct8-knockout in osteoclast precursors, osteoprogenitors, or mature osteoblasts/osteocytes were analyzed regarding their bone microarchitecture, turnover, and strength. Furthermore, ex vivo studies were conducted to investigate the role of Mct8 in bone cell differentiation and functionality, as well as TH uptake.
Results:
Global Mct8-knockout mice demonstrated 1.7-fold higher T3 serum concentrations and trabecular bone loss (−28%) likely due to an increased bone turnover as shown by increased osteoblast (+45%) and osteoclast numbers (+41%). However, cortical bone mineral density was increased. Ex vivo cultures of bone marrow-derived osteoblasts and osteoclasts revealed highest expression of Mct8 in mature bone cells. In addition, Mct8-deficiency resulted in a lower mRNA expression of osteoblast and osteoclast differentiation markers, as well as a reduced mineralization capacity and osteoclast numbers, respectively, indicating a bone cell intrinsic role of Mct8. In fact, conditional Mct8-knockout and inhibition of Mct8 in osteoblasts led to an attenuated T3 uptake ex vivo. In vivo, osteoprogenitor-specific Mct8-knockout enhanced trabecular bone volume (+16%) with osteoblast numbers being increased 3.7 fold. Interestingly, Mct8-deficiency in osteoprogenitors and late osteoblasts/osteocytes both resulted in cortical bone loss. Finally, Mct8-deletion in osteoclast progenitors increased trabecular bone volume (+20%) due to reduced osteoclast numbers (−32%), whereas osteoblast numbers were enhanced (+25%).
Conclusions:
This study confirms that high systemic T3 in global Mct8-knockout mice masks the direct effect of Mct8. Moreover, it identifies Mct8 as a critical regulator of trabecular vs. cortical bone by regulating T3 uptake and highlights its cell intrinsic role in osteoclast and osteoblast progenitors.
Introduction
Bone homeostasis requires a balance between bone formation and bone resorption. Bone-forming osteoblasts and bone-resorbing osteoclasts need to act in concert and are influenced by stimulating or repressing cytokines and endocrine factors (1,2). Thyroid hormones (THs) are indispensable regulators of bone homeostasis and bone maintenance in adults (3 –5). The synthesis of the prohormone L-thyroxine (T4) and its receptor active form 3,3′,5-L-triiodothyronine (T3) is tightly controlled by the hypothalamic-pituitary-thyroid axis. Clinical studies revealed that untreated hyperthyroidism can lead to secondary osteoporosis accompanied by an increased fracture risk as a result of an accelerated bone turnover favoring bone resorption (6 –8). On the contrary, lack of TH decelerates bone turnover, and thus, hypothyroid patients display increased secondary mineralization (7,9).
Despite the well-known effects of TH on bone turnover, their role in bone cells remains ill defined. In murine osteoblasts, T3 enhances differentiation and stimulates the synthesis of bone matrix (10 –16). The effects of TH on osteoblasts are mediated mainly through modulation of genomic signaling by the nuclear thyroid hormone receptors (TR), TRα1 and TRβ1 (17 –20). Furthermore, iodothyronine deiodinases (DIO) are able to activate or inactivate TH intracellularly by monodeiodination. Locally, DIO2 catalyzes T4 to T3 conversion, while DIO3 can inactivate T3 into 3,3′-T2 and T4 into reverse T3, respectively. Thus, both enzymes contribute to TH homeostasis in osteoblasts depending on the developmental stage at the cellular, as well as the systemic level (4,21,22).
In murine osteocytes, however, which represent terminally differentiated cells of the osteoblast lineage and act as mechanosensors in bone, expression of TRs and deiodinases, as well as TH-mediated effects, has not been investigated yet (3). In osteoclasts, Thra1 and Thrb1 expression, as well as DIO3 activity, has been described indicating that osteoclasts are responsive to TH (19 –21). Osteoclastogenesis is enhanced by T3, yet the underlying mechanisms are not well understood and might be indirectly mediated by cells of the osteoblast lineage (3,23 –27).
Besides nuclear receptor availability and a balanced DIO2:DIO3 ratio, TH actions rely on their import through TH transporters (28). L-Type amino acid transporters (Lat)1 and Lat2 guide the uptake of large neutral, branched-chain and aromatic amino acids, but show only a low specificity for TH (21,29,30). Monocarboxylate transporters (Mcts) export and import monocarboxylate substrates such as pyruvate, lactate, ketone bodies, carnitine, and aromatic amino acids (31,32). Among the 14 members of the Mct family, Mct8 is rather unique as it only accepts TH as substrates (33 –35).
The Mct8 encoding gene SLC16A2 is located on the X chromosome in humans and mice. Patients with inactivating mutations in SLC16A2 develop profound intellectual and physical disabilities, also known as the Allan-Herndon-Dudley (AHD) syndrome. Apart from their mental and motor disabilities, affected males display highly elevated serum T3 concentrations, but rather low concentrations of T4. As a consequence, Mct8-dependent organs such as the brain are TH deficient, whereas tissues expressing additional TH transporters like the kidney, liver, or skeletal muscle are exposed to high serum T3 levels and are therefore in a thyrotoxic state (36 –38). Mct8 expression was previously reported in bone cells of mice (21,30), suggesting a role of this transporter in bone physiology.
In fact, a recent study of Mct8-knockout mice that closely replicates the abnormal serum TH profile seen in human patients revealed a decrease in trabecular bone mass, but the underlying pathogenic mechanisms remain elusive (39). To distinguish between direct effects of Mct8-deficiency in bone cells and indirect effects caused by the abnormal serum TH profile, we generated conditional Mct8 mutant animals that lack Mct8 specifically in osteoclasts, osteoblasts, and osteocytes. Our study shows that Mct8 plays a cell-intrinsic role in osteoblasts and osteoclasts and that it may differentially affect trabecular versus cortical bone turnover.
Materials and Methods
Animal models
Mct8-knockout (KO) mice were obtained from Deltagen (Palo Alto, CA) and have been described previously (40). Wild-type littermates were used as controls. All mice were on a C57BL/6 background. Floxed Mct8 mice (Slc16a2 tm1a(KOMP)Wtsi, internal abbreviation Mct8 fl) (41) were obtained from the KOMP repository (Oakland, CA) and carry loxP sites flanking exon 3 of the Mct8 encoding gene Slc16a2 (internal abbreviation Mct8). For generating osteoblast-specific Mct8-deficient animals, Mct8 fl mice were crossed with doxycycline-repressible Osx:Cre mice (42). Breeding pairs of the generated Mct8 fl;Osx:Cre mice received 10 mg/mL doxycycline dissolved in 3% sucrose solution in their drinking water ad libitum to inhibit Cre-activity during embryonic development. In addition, Mct8 fl;Osx:Cre offspring received doxycycline drinking water up to 5 weeks after weaning. Similarly, Mct8 fl;Dmp1:Cre and Mct8 fl;LysM:Cre were generated to inactive Mct8 specifically in osteocytes/late osteoblasts and osteoclast progenitors by crossing Mct8 fl mice with Dmp1:Cre mice (43) and LysM:Cre mice (44), respectively. Respective Cre-negative littermates were used as controls. Efficient Mct8 deletion and recombination in Cre-targeted cells and organs were confirmed by polymerase chain reaction (PCR) analysis (40 –42). All experiments were performed using 12-weeks-old male mice. Animals were maintained in groups up to four animals in a 12-h light/12-h dark cycle at room temperature in filter-top cages with cardboard houses for enrichment purposes and had ad libitum access to their respective drinking water and standard chow diet. Animal procedures were approved by the Institutional Animal Care Committee of the Technische Universität Dresden and the Landesdirektion Dresden and were performed according to the ARRIVE guidelines.
Serum analysis
Animals were euthanized using CO2, and blood was collected through heart puncture. Subsequently, serum was obtained by centrifugation at 400 g for 15 minutes. Concentrations of type 1 procollagen aminoterminal-propeptide (P1NP) and C-terminal telopeptide (CTX) were measured using enzyme-linked immunosorbent assays (ELISAs) according to the manufacturers' protocol (IDS, Frankfurt/Main, Germany). Total T4 and T3 serum concentrations were quantified using radioimmunoassays (RIA-4524 and RIA-4525, respectively; DRG Instruments, Marburg, Germany) as previously described (45,46). The lower limit of analysis was 10 and 0.22 nM for T4 and T3, respectively. Intra-assay coefficients of variations were below 7.5%.
Analysis of femur length, bone mass, and microarchitecture
Length of the femur was measured using a caliper. For bone mass and microarchitecture assessment, the distal femur and fourth lumbar vertebra (L4) were analyzed post mortem using μCT (vivaCT 40, Scanco Medical, Brüttisellen, Switzerland) with an X-ray energy of 70 kVp and isotropic voxel size of 10.5 μm (114 mA, 200 ms integration time). All trabecular (Tb) and cortical (Ct) bone parameters are based on calculations, including 100 scan slices using standard protocols from Scanco Medical. Parameters include bone volume per total volume (BV/TV), trabecular number (Tb.N), trabecular separation (Tb.Sp), and trabecular thickness (
Bone histology and histomorphometry
Dynamic bone histomorphometry was conducted as described previously (48). Briefly, mice were i.p. injected with calcein (20 mg/kg; Sigma, Germany) 5 and 2 days before sacrifice. The third lumbar vertebrae and tibias were fixated in 4% phosphate buffered saline (PBS)-buffered paraformaldehyde for 48 hours and dehydrated through ascending ethanol series. Subsequently, bones were embedded in methyl methacrylate (Technovit 9100; Heraeus Kulzer, Hanau, Germany) and cut into 7 μm sections for calcein label quantification. Using fluorescence microscopy and the Osteomeasure software (OsteoMetrics, Atlanta, GA), mineralized surface/bone surface, the mineral apposition rate (MAR), and the bone formation rate/bone surface were assessed following international established protocols (49). Furthermore, Technovit sections were stained with von Kossa/van Gieson for visualization of unmineralized bone matrix (osteoid). Using Osteomeasure software, the osteoid width (O.Wi) and mineralized surface/osteoid surface (MS/OS) were quantified and used for calculation of the maturation lag time (MLT = O.Wi/(MAR*MS/OS)).
Tartrate-resistant acid phosphatase (TRAP) staining was performed on 2 μm thick paraffin sections of the fourth lumbar vertebrae and femora, which were beforehand fixed in 4% PBS-buffered paraformaldehyde, decalcified in Osteosoft (Merck, Germany) for 7 days, and dehydrated using ascending ethanol series.
Biomechanical testing
Femurs were rehydrated in PBS for 24 hours before 3-point bending test (Zwick Roell, Ulm, Germany). Analysis started after reaching a preload of 1 N and continued with a load rate of 0.05 mm/s until failure. Results were evaluated using testXpert II—V3.7 software. Energy to failure was determined based on raw load displacement curves.
Cell culture and functionality tests of primary murine osteoblasts and osteoclasts
Hind legs of Mct8-knockout and wild-type mice were harvested, and mesenchymal stromal cells (MSCs) were acquired by flushing the bone marrow. Cells were cultured until 80% confluence in growth medium consisting of Dulbecco's Modified Eagle's Medium (ThermoFisher Scientific, Schwerte, Germany) (DMEM), 10% fetal calf serum (FCS; from Biochrom, Berlin, Germany), 1% penicillin/streptomycin (P/S; from Invitrogen, Carlsbad, CA) in a humidified atmosphere of 95% air and 5% CO2 at 37°C, and then differentiated into osteoblasts by adding 100 μM ascorbate phosphate and 5 mM β-glycerol phosphate (both from Sigma-Aldrich, St. Louis, MO) to the growth medium. To evaluate mineralization ability, primary osteoblasts were differentiated over 10 and 21 days. Subsequently, cells were fixated in 10% paraformaldehyde for 15 minutes and stained with 1% Alizarin red S solution (pH 5.5; Sigma-Aldrich, St. Louis) for 30 minutes at room temperature. Excessive staining was removed by repeated washing steps using distilled water. Cetylpyridinium chloride (Sigma-Aldrich, St. Louis) was used to dissolve and quantify the amount of incorporated calcium by its photometric measurement at a wavelength of 540 nm.
Alkaline phosphatase (ALP) activity was assessed as a marker of osteoblastic differentiation. Therefore, Mct8 fl;Osx:Cre mice-derived osteoblasts were treated over 48 hours with 1 μM T3. Cells were washed twice in PBS and then scraped in ALP lysis buffer (10 mM Tris-HCl pH 8.0, 1 mM MgCl2, and 0.5% Triton X-100) for isolation of total proteins. Subsequently, cell lysates were processed through a 24-gauge needle and centrifuged at 25,000 g for 30 minutes at 4°C. Ten microliters of diluted supernatants (1:5 in distilled water) were incubated with 90 μL ALP substrate buffer (100 mM diethanolamine, 150 mM NaCl, 2 mM MgCl2, and 2.5 g/mL p-nitrophenyl phosphate) for 30 minutes at 37°C. Color change was observed due to hydrolysis of p-nitrophenyl phosphate by enzymatic activity of ALP. Absorbance was measured at 410 nm using FLUOstar Omega (BMG Labtech, Offenburg, Germany). Results were normalized to the total protein content that was quantified using Pierce™ BCA Protein Assay Kit (Thermo Fisher Scientific, Schwerte, Germany) according to the manufacturer's protocol.
To obtain osteoclasts, bone marrow cells were grown in Alpha-Medium (Biochrom GmbH, Berlin, Germany) supplemented with 10% FCS, 1% P/S, and 25 ng/mL macrophage colony-stimulating factor (from R&D Systems, Minneapolis) for 48 hours, and then further treated with 50 ng/mL receptor activator of nuclear factor kappa-B ligand (RANKL; from R&D Systems) for up to 7 days. The number of TRAP-positive osteoclasts was quantified by TRAP staining at days 5 and 7 after starting RANKL treatment. Therefore, cells were first fixated for 10 minutes in acetone/citrate buffer containing 37% paraformaldehyde at room temperature. After two washing steps with tap water, staining was performed over 20 minutes in the dark using TRAP-Kit (from Sigma-Aldrich, St. Louis). TRAP-positive cells with three or more nuclei were counted as osteoclasts.
RNA from osteoblasts and osteoclasts was collected after 24 hours of starvation in the respective medium with only 1% FCS at different indicated time points.
To confirm Mct8-deletion in osteoclast progenitors, bone marrow-derived osteoclast precursors were sorted through magnetic-activated cell separation. To that end, bone marrow of Mct8 fl;LysM:Cre mice was harvested, and Cd11b-positive macrophages were isolated using Dynabeads™ Biotin Binder and CD11b Monoclonal Antibody (M1/70), eBioscience™ (both Invitrogen, Darmstadt, Germany) according to the manufacturer's protocol. Subsequently, sorted cells were differentiated into osteoclasts over 7 days, and RNA was isolated. Mct8-knockdown in osteocytes of Mct8 fl;Dmp1:Cre mice was assessed from bone marrow-derived MSCs that were differentiated over 28 days into osteocytes, as well as in cortical bone tissue that mainly contains osteocytes.
TH uptake assay
The protocol of the nonradioactive TH uptake assay is based on the Sandell–Kolthoff reactions and was modified after the protocol from Jayarama-Naidu et al. (50). Primary murine MSCs derived from wild-type and Mct8 fl;Osx:Cre mice were seeded into 96-well plates, grown until 80% confluence, and differentiated over 7 days. In addition, RAW264.7 cells (kindly provided by Dr. Ute Hempel, Dresden, Germany) and MLO-Y4 cells (kindly provided by Dr. Lynda Bonewald, Indianapolis) were used to assess TH uptake into osteoclasts and osteocytes, respectively. Cells were washed twice in PBS and treated with indicated concentrations of T3 or T4 and specific Mct8-blocker silychristin (51) (Sigma-Aldrich, Munich, Germany) both diluted in uptake buffer (50 mM HEPES; pH 7.4 supplemented with 125 mM sodium chloride, 1.3 mM calcium chloride, 1.2 mM magnesium chloride, 5 mM potassium chloride, and 5.6 mM D-glucose) over 15 minutes at 37°C. Subsequently, wells were washed twice with 0.1% BSA/PBS solution and once with ice-cold distilled water. To release the iodide from the cells, an oxidative digest was performed by adding 50 μL ammonium persulfate solution (0.68 M) to each well. Then, plates were tightly sealed with self-adhesive foil and incubated for 1 hour at 90°C. After that, plates were quickly centrifuged at 2000 g, and 45 μL of the supernatant was transferred into a new 96-well plate. Forty-five microliters ceric solution (25 mM ammonium cerium (IV) sulfate hydrate and 0.5 M sulfuric acid) was added into each well. To start the iodide-catalyzed destaining reaction, 45 μL arsenic solution (25 mM sodium meta-arsenite, 0.5 M sulfuric acid, and 0.2 M sodium chloride) was applied into each well. Intermediately, absorption was measured at a wavelength of 415 nm using FLUOstar Omega. Measurement was repeated after 30 minutes. Relative iodine content was calculated as the difference of absorption at the starting point (1 minute) and after 30 minutes. The cellular iodine content reflects the relative uptake of iodinated test compounds such as T3 or T4.
mRNA isolation, reverse transcription-PCR, and quantitative real-time PCR
Total RNA from primary cells was extracted with the High Pure RNA Isolation Kit (Roche, Mannheim, Germany) according to the manufacturer's protocol and quantified using the NanoDrop spectrophotometer (Peqlab, Erlangen, Germany).
To validate the osteocyte-specific Mct8-knockdown, long bones of Mct8 fl;Dmp1:Cre mice were flushed and cortical bone that is rich of osteocytes was harvested for total RNA isolation using TRIzol reagent (Invitrogen, Darmstadt, Germany) following the manufacturer's instructions. Five hundred nanograms of RNA were reverse transcribed using Superscript II (Invitrogen, Darmstadt, Germany) followed by SYBR Green-based quantitative real-time PCR according to established protocols (ABI7500 Fast; Applied Biosystems, Carlsbad, CA). The primer sequences were: β-actin sense: GATCTGGCACCACACCTTCT, β-actin antisense: GGGGTGTTGAAGGTCTCAAA;
Alp sense: CTACTTGTGTGGCGTGAAGG, Alp antisense: CTGGTGGCATCTCGTTATCC; Ctsk sense: AAGTGGTTCAGAAGATGACGGGAC, Ctsk antisense: TCTTCAGAGTCAATGCCTCCGTTC; Gapdh sense: AAGGTCATCCCAGAGCTGAA, Gapdh antisense: CTGCTTCACCACCTTCTTGA; Mct8 (Slc16a2) sense: CCCTGGACTTAAGAAGATATACTTGCA, Mct8 (Slc16a2) antisense: CCCGAAGTCCCGGCATA; Nfatc1 sense: GTTCCTTCAGCCAATCATCC, Nfatc1 antisense: GGAGGTGATCTCGATTCTCG; Ocn sense: GCGCTCTGTCTCTCTGACCT, Ocn antisense: ACCTTATTGCCCTCCTGCTT; Runx2 sense: AAATGCCTCCGCTGTTATGAA, Runx2 antisense: GCTCCGGCCCACAAATCT; and Trap sense: ACTTGCGACCATTGTTAGCC, Trap antisense: AGAGGGATCCATGAAGTTGC.
PCR conditions were: 50°C for 5 minutes and 95°C for 10 minutes followed by 40 cycles with 95°C for 15 seconds and 60°C for 1 minute. Melting curves were assessed by the following scheme: 95°C for 15 seconds, 60°C for 1 minute, and 95°C for 30 seconds. Results were calculated based on the ΔΔCT method and are shown in x-fold increase normalized to β-actin and Gapdh mRNA levels, respectively.
Statistical analysis
Results are shown as mean ± standard deviation. Statistical analysis was performed using Student's t-test. Group analyses were performed using one-way analysis of variance (ANOVA) by GraphPad Prism 7.0 based on Bonferroni tests (GraphPad, La Jolla, CA). Values of p < 0.05 were considered statistically significant.
Results
Global Mct8-deficiency leads to trabecular bone loss due to enhanced bone resorption and decelerated bone mineralization
Given the crucial role of TH in bone metabolism, we analyzed the bones of 12-week-old male mice with a global Mct8-knockout and compared them to littermate wild-type controls to obtain information about the number and functionality of bone cells. No significant differences regarding body weight and femur length were detected between knockout and wild-type animals (Table 1). In line with other studies (40,52), Mct8-deficient mice had 1.7-fold higher T3 serum concentrations, while T4 concentrations were decreased by 56.7% (Table 2). μCT analysis revealed trabecular bone loss at the spine (−27.7% BV/TV) of Mct8-knockout mice with significantly reduced Tb.N (−15.4%) and thickness (−8.8%), while Tb.Sp was increased by 8.8% (Fig. 1A–D). Cortical bone volume fraction, bone mineral density, and thickness were increased in Mct8-deficient mice (Fig. 1E–G). Furthermore, there was a trend toward higher bone strength in femora from those mice as indicated by a higher energy to failure, although this did not reach statistical significance (Fig. 1H). At the histological level, the number of osteoclasts was increased (+41.0%), and there was a trend toward higher serum concentration of the bone resorption marker CTX indicating elevated bone resorption in knockout mice (Fig. 1I, J). In addition, bone formation was augmented with enhanced osteoblast numbers (+44.6%) (Fig. 1K) and nonsignificant increases in the bone formation rate (data not shown) and serum concentration of the bone formation marker P1NP (Fig. 1L). Finally, Mct8-knockout mice had an increased O.Wi (+25.4%) and a prolonged MLT (+72.3%) suggesting delayed mineralization of osteoid by Mct8-deficient osteoblasts (Fig. 1M–O). Similar trends regarding trabecular bone mass, microarchitecture, and histomorphometry were seen at the femur (Supplementary Fig. S1A–H). Thus, these data indicate that trabecular bone loss of global Mct8-knockout mice may be due to accelerated bone turnover with predominant bone resorption and delayed bone mineralization.
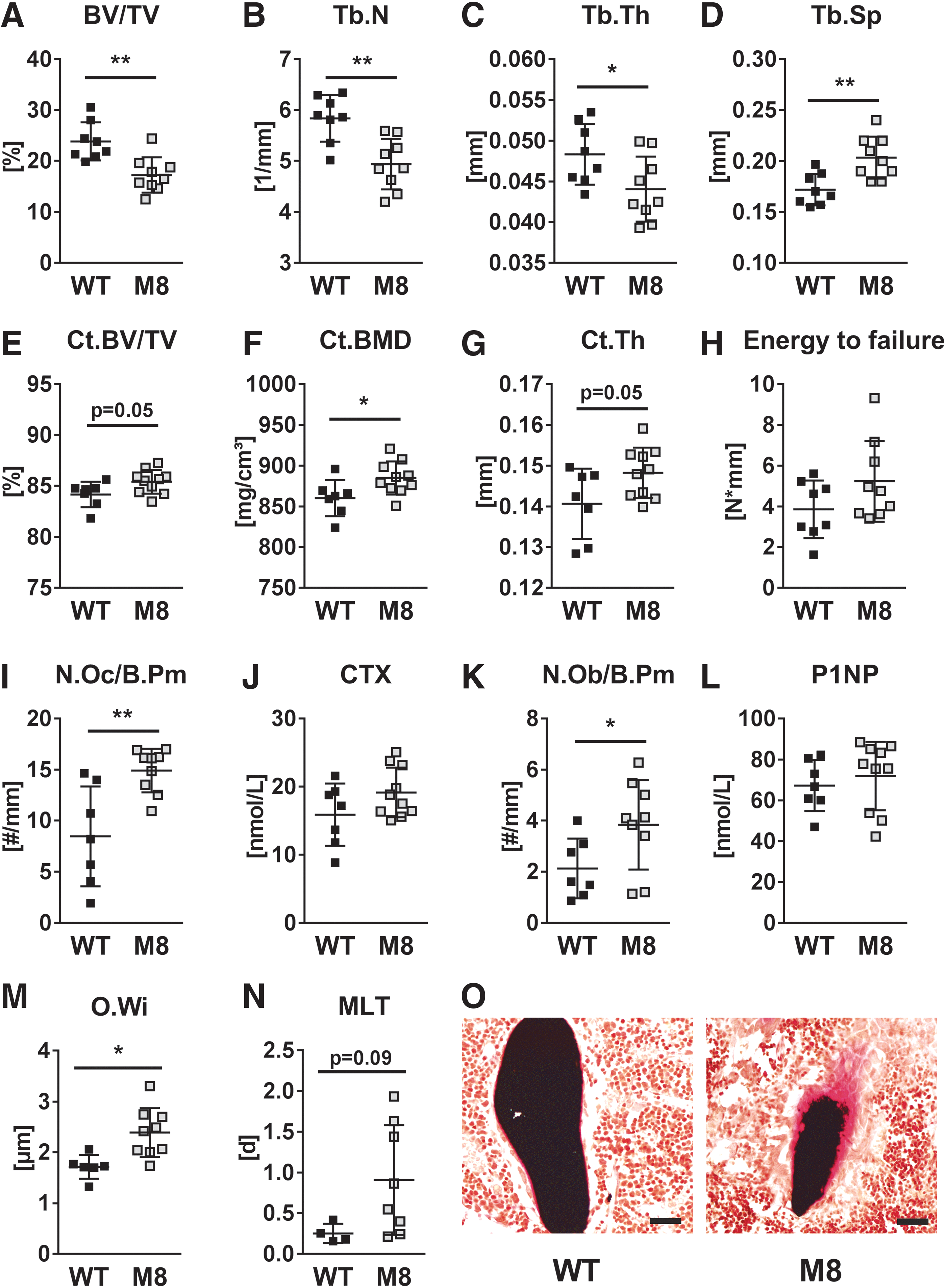
Global deletion of Mct8 results in trabecular bone loss, but increased cortical bone. Trabecular bone compartment of L4 vertebra and femora of 12-week-old male Mct8 global knockout mice (M8) vs. WT controls was analyzed by μCT. (
Body Weight and Femoral Length of 12-Week-Old Mct8-Knockout, Mct8 fl;LysM:Cre Mice, Mct8 fl;Osx:Cre, and Mct8 fl;Dmp1:Cre Mice
Data are represented as the mean ± SD. Statistical analysis was performed by the Student's t-test.
KO, knockout; Mct8, monocarboxylate transporter 8; SD, standard deviation.
3,3′,5-L-Triiodothyronine and L-Thyroxine Serum Concentrations of 12-Week-Old Mct8-Knockout, Mct8 fl;LysM:Cre Mice, Mct8 fl;Osx:Cre, and Mct8 fl;Dmp1:Cre Mice
Data are represented as the mean ± SD. Statistical analysis was performed by the Student's t-test.
Mct8-deficiency impairs differentiation and functionality of bone cells in vitro
To test whether the enhanced bone turnover in Mct8-deficient mice is a consequence of high serum T3 concentrations or whether Mct8-deficiency directly compromises bone homeostasis, we analyzed Mct8-knockout osteoblasts and osteoclasts ex vivo under euthyroid conditions. First, we evaluated the expression of Mct8 in primary murine wild-type bone cells during the course of their differentiation as indicated by a rising expression of osteoblast marker osteocalcin (Ocn) and osteoclastic marker cathepsin K (Ctsk), respectively. In osteoblasts, Mct8 expression increased from day 0 up to its maximum at day 21 (6.6-fold, d0 vs. d21) (Fig. 2A). Osteoclasts expressed the highest levels of Mct8 mRNA at day 5 after starting RANKL-induced differentiation (d0 vs. d5; 40-fold increase) (Fig. 2B). Thus, Mct8 is expressed during osteoblast and osteoclast differentiation and shows the highest expression in mature bone cells.

mRNA expression of Mct8 increases in murine bone marrow-derived osteoblasts and osteoclasts during differentiation. Using real-time PCR analysis, mRNA expression of (
Subsequently, we tested whether a loss of Mct8 expression affects osteoblast and osteoclast differentiation, as well as functionality. Beforehand, we confirmed the absence of Mct8 mRNA in osteoblasts and osteoclasts derived from Mct8 global knockout mice (Fig. 3A, G). Mct8-deficient osteoblasts showed a reduced expression of differentiation marker genes Alp, Ocn and Runt-related transcription factor 2 (Runx2) analyzed at day 7 of differentiation (−58.4%/−89.2%/−61.1%) (Fig. 3B–D). In addition, their mineralization capacity was impaired at day 10 and 21 (Fig. 3E, F). Similarly, Mct8-deficiency resulted in a reduced gene expression of osteoclast differentiation markers (Ctsk, nuclear factor of activated T cells, cytoplasmic 1, and Trap) (Fig. 3H–J) and an impaired generation of mature osteoclasts (Fig. 3K, L). Thus, these data suggest a cell-intrinsic role of Mct8 in bone cells.
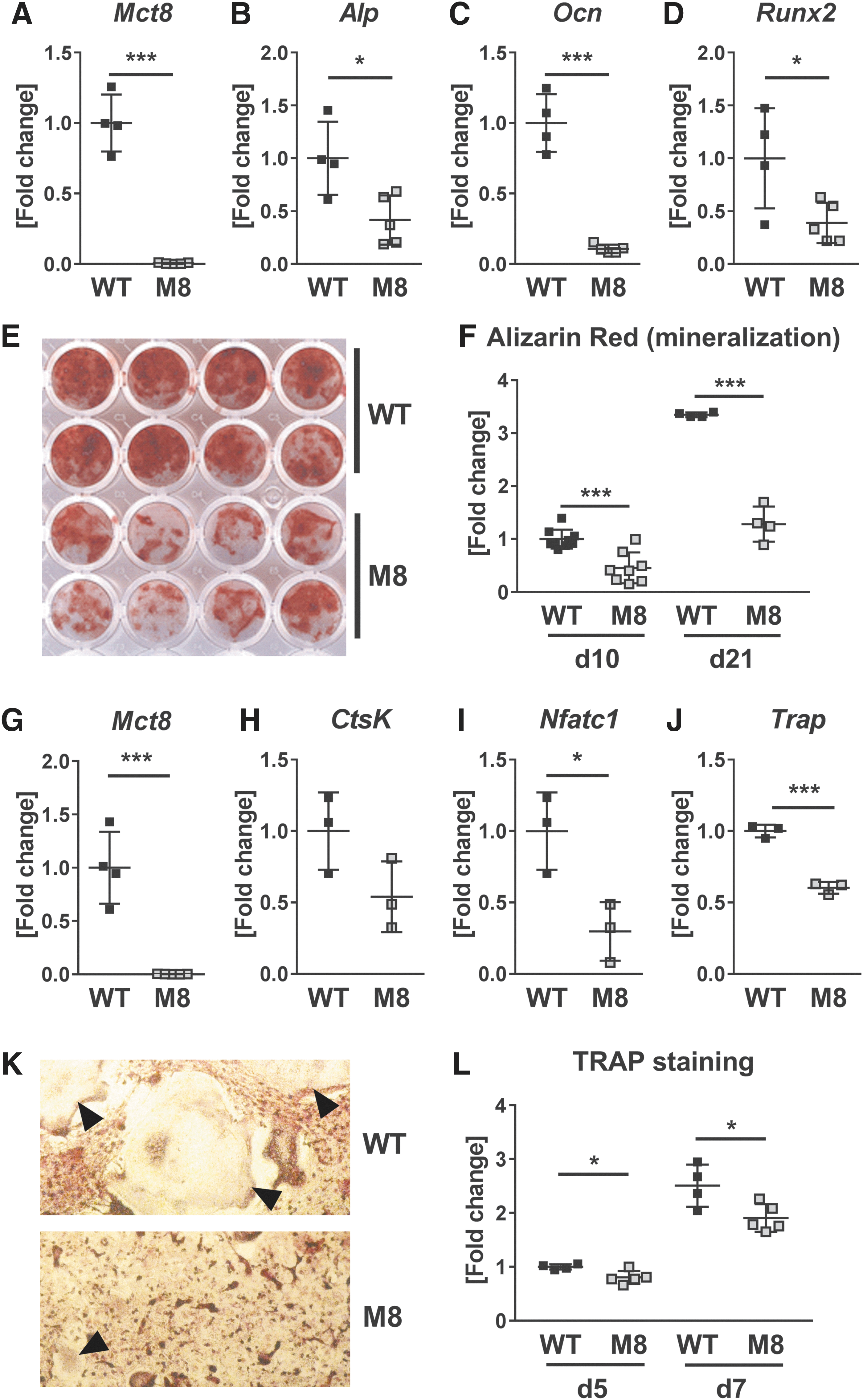
Differentiation and performance of primary murine osteoblasts and osteoclasts decrease with Mct8-deletion. (
Conditional Mct8-deletion in osteoclast progenitors results in trabecular bone gain
As Mct8 is highly induced during osteoclast differentiation and its deficiency results in impaired osteoclastogenesis, we crossed Mct8-floxed mice with LysM:Cre mice to inactivate Mct8 in osteoclast progenitors and examine its role in osteoclasts. Knockdown of Mct8 was confirmed in bone-marrow derived osteoclasts ex vivo (−74%) (Supplementary Fig. S2A). Cre-positive mice and Cre-negative control littermates displayed similar body weight, femur length, and TH serum concentrations (Tables 1 and 2). Reduced Mct8 expression in osteoclast progenitors of Cre-positive mice led to increased trabecular bone volume at the spine, while cortical parameters and bone strength were not changed (Fig. 4A–H). Trabecular bone volume was increased by 19.5% (Fig. 4A) and
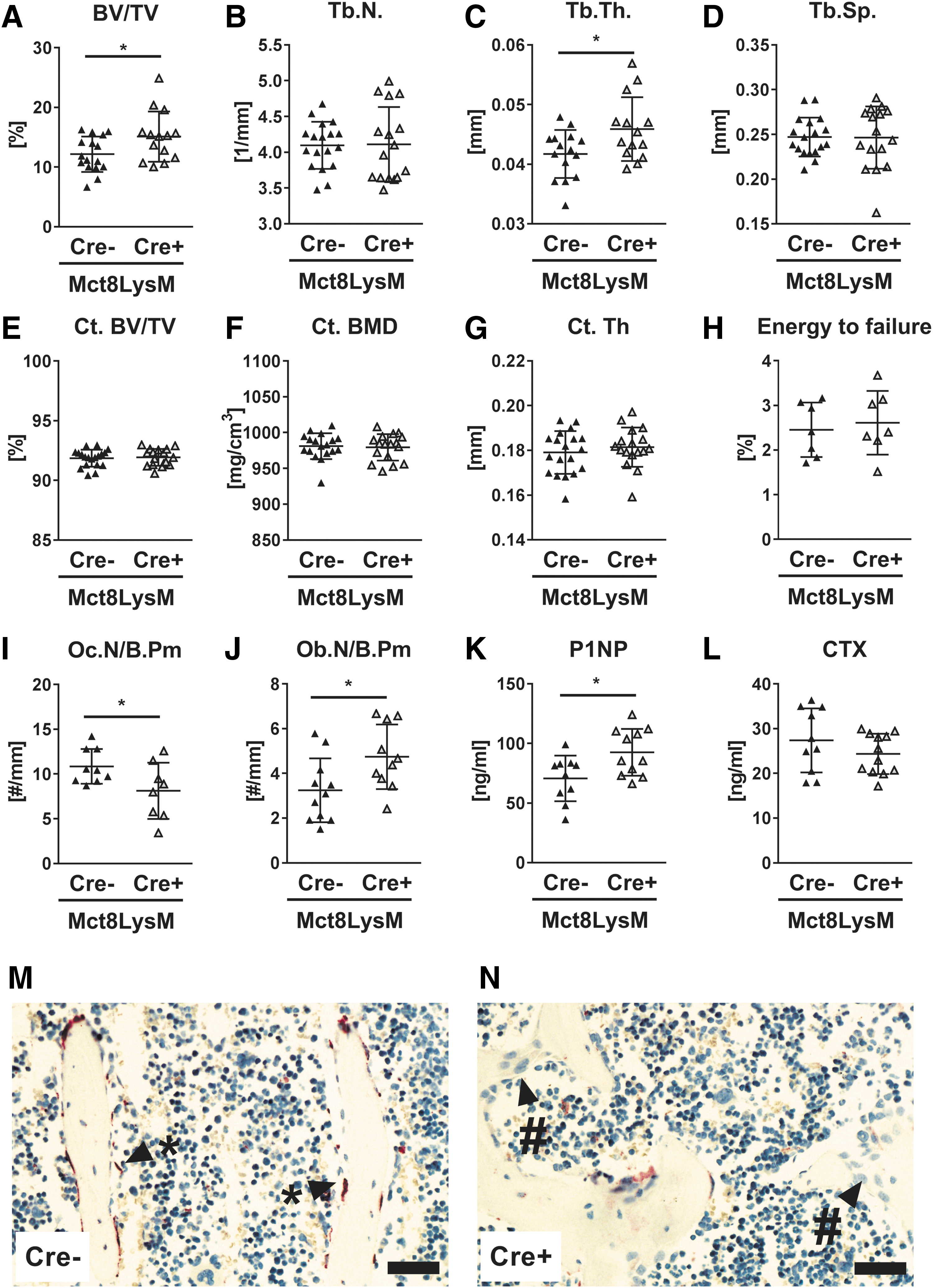
Osteoclast progenitor-specific Mct8-deletion enhances trabecular bone volume. Trabecular bone compartment of L4 vertebra and femora of 12-week-old male Mct8 fl;LysM:Cre mice were analyzed by μCT. Mct8 fl;LysM:Cre-positive mice (Cre+) were compared to littermate controls (Cre−). (
Conditional Mct8-deletion in osteoblast progenitors increases trabecular bone mass while reducing cortical bone
As ex vivo data suggested a cell intrinsic function of Mct8 in bone, we aimed to investigate the role of Mct8 in different osteoblast maturation stages in vivo. Therefore we generated Mct8 fl;Osx:Cre and Mct8 fl;Dmp1:Cre mice that target osteoprogenitors and late osteoblasts/osteocytes, respectively. Cre-positive mice and Cre-negative littermate controls did not differ in body weight, femur length, and TH serum concentrations (Tables 1 and 2). At the age of 12 weeks, male Mct8 fl;Osx:Cre-positive mice displayed a higher trabecular bone volume at the lumbar vertebra (+16.2%) and more trabeculae (+9.1%), whereas Tb.Sp was reduced (−8.2%) (Fig. 5A–D). Knockdown of Mct8 in osteoprogenitor cells decreased cortical bone mineral density and cortical bone volume, while thickness was not altered (Fig. 5E–G). Nonetheless, Cre-positive mice depicted enhanced bone strength in the femur in comparison to their wild-type littermate controls (+41.5%) (Fig. 5H).
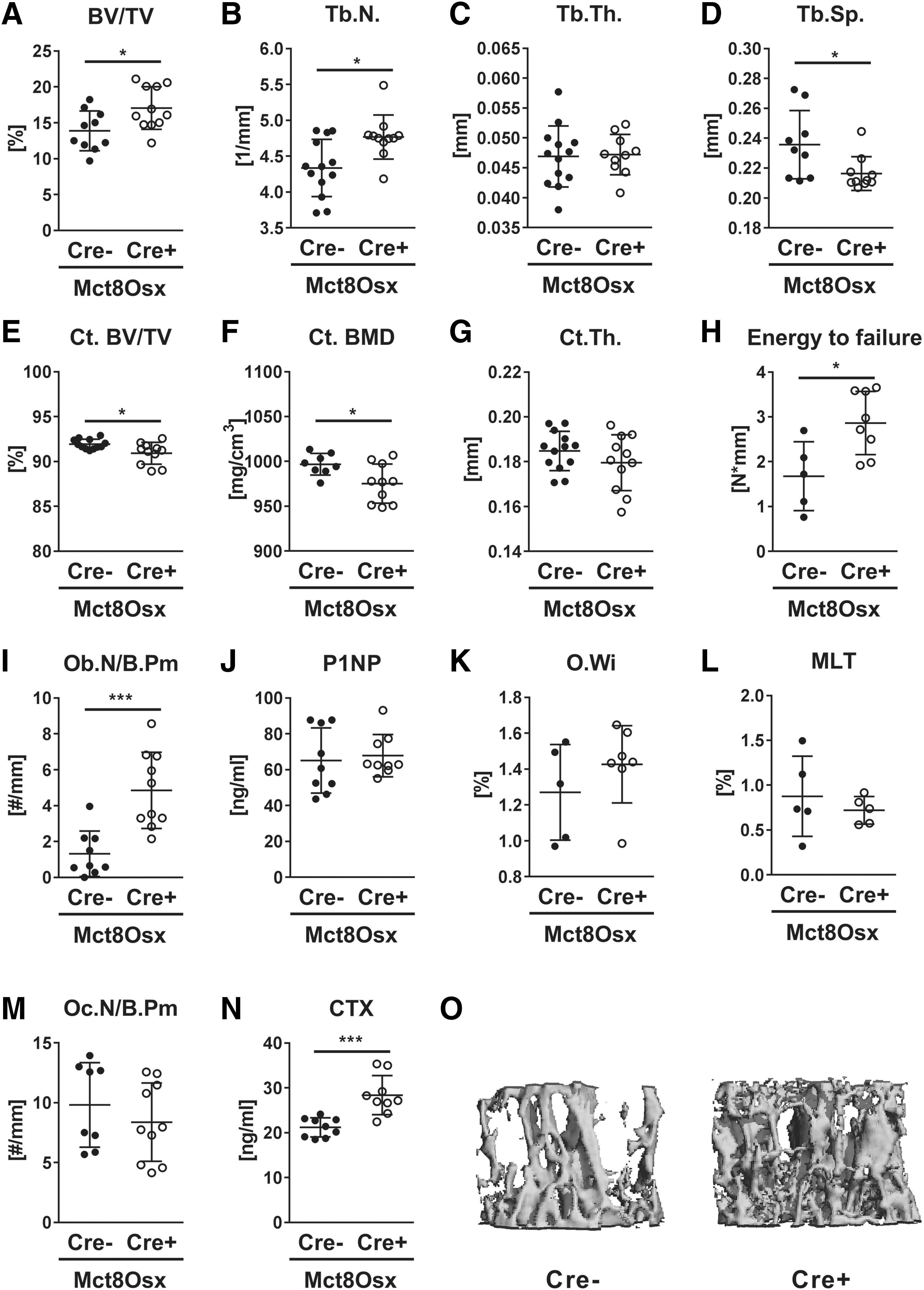
Lack of Mct8 in osteoprogenitors leads to trabecular bone gain, but cortical bone loss. Trabecular bone compartment of L4 vertebra and femora of 12-week-old male Mct8 fl;Osx:Cre mice were analyzed by μCT. Mct8 fl;Osx:Cre-positive mice (Cre+) were compared to littermate controls (Cre−). (
Histological analysis revealed a 3.7-fold increase in osteoblast numbers of Cre-positive animals, while P1NP serum concentration, as well as bone mineralization parameters, remained unchanged (Fig. 5I–L). Osteoclast numbers were not affected, but the CTX serum concentration was 1.3-fold increased (Fig. 5M, N). Representative images of bone microarchitecture are shown in Figure 5O. Changes in bone microarchitecture, histology, and histomorphometry at the femur were similar to those seen at the spine (Supplementary Fig. S4A–H).
As trabecular bone gain and cortical bone loss were observed concomitantly in Mct8 fl;Osx:Cre-positive mice targeting osteoprogenitors, we further specified the targeted cell population and exclusively deleted Mct8 in late osteoblasts and osteocytes. Mct8-knockdown in Mct8 fl;Dmp1:Cre-positive mice was confirmed in cultured and cortical bone-derived osteocytes (Supplementary Fig. S2B, C). Analysis of the bone phenotype of Mct8 fl;Dmp1:Cre-positive mice versus littermate controls indicated a trend toward increased trabecular bone volume, while the cortical bone volume of mice was significantly decreased (Supplementary Fig. S5A–G). Bone strength (Supplementary Fig. S5H) and bone turnover parameters (data not shown) did not significantly differ between knockout and control mice.
Taken together, Mct8 appears to be a negative regulator of osteoblasts, as well as trabecular bone mass, and may distinctly regulate trabecular versus cortical bone mass. Furthermore, Mct8-deficiency appears to affect early osteoblast stages more strongly as the bone phenotype of Mct8 fl;Osx:Cre-positive mice targeting osteoprogenitors was more severe compared with Mct8 fl;Dmp1:Cre-positive mice targeting late osteoblasts and osteocytes.
Mct8-inhibition compromises T3 uptake of murine bone cells
To ascertain whether Mct8 is involved in TH uptake in osteoblasts, we used the Mct8-specific inhibitor silychristin to block TH import (51). Silychristin significantly inhibited the uptake of T3 concentrations <5 μM (Fig. 6A), while T4 import remained unaffected (Fig. 6B). Furthermore, we tested Mct8-deficient osteoblasts derived from Mct8 fl;Osx:Cre-positive mice that exhibit a reduced Mct8 expression by −60% (Fig. 6C) and demonstrated an attenuated T3 uptake at T3 concentration <5 μM compared to respective controls (Fig. 6F). Again, T4 import was not changed with Mct8-deficiency (Fig. 6G). In line with osteoblasts generated from global Mct8-knockout mice, ALP activity and mineralization capacity of conditional Mct8-knockout osteoblasts were decreased (Fig. 6D, E).
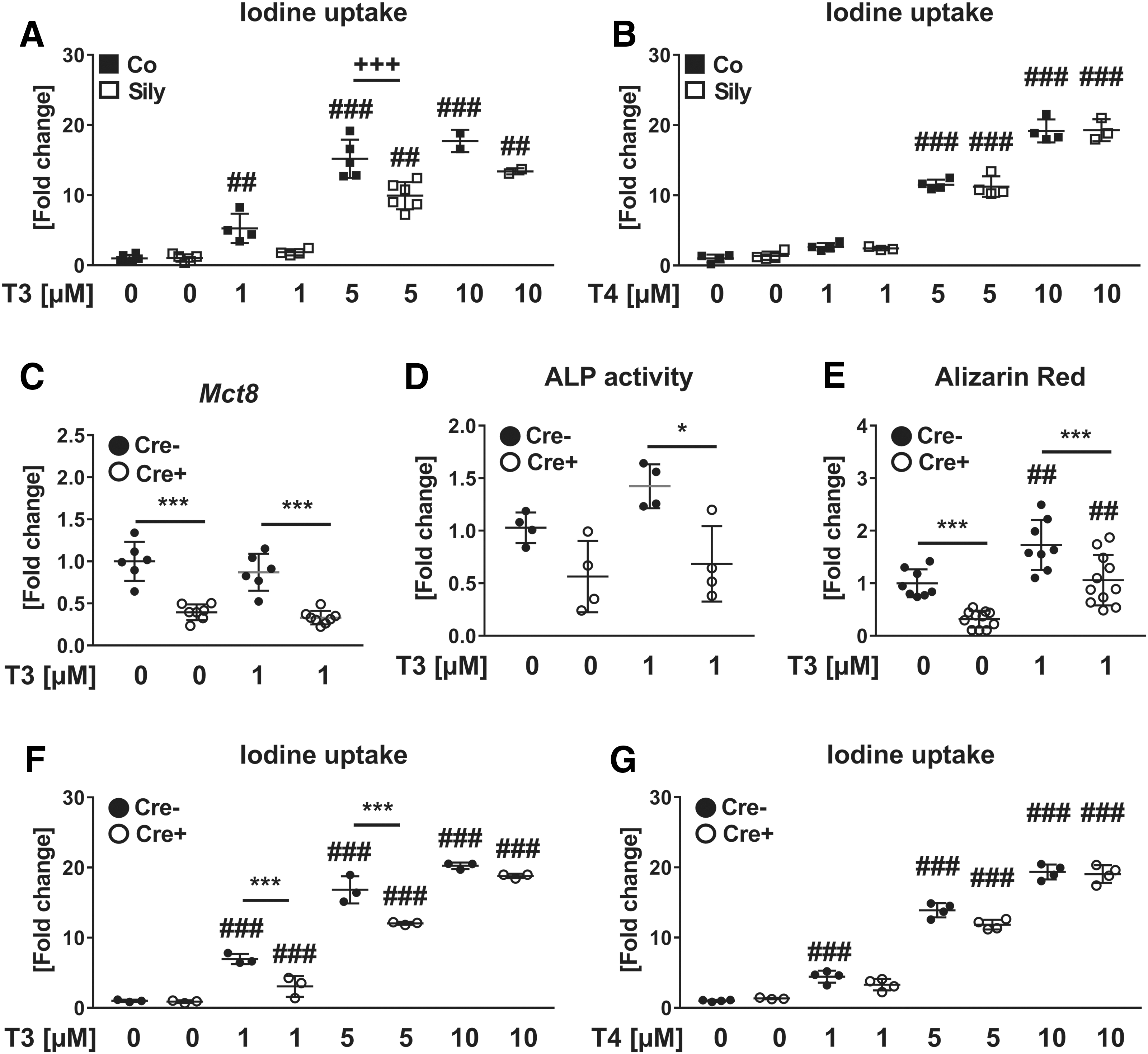
Silychristin-mediated Mct8-blockade and conditional Mct8-knockout restrict T3, but not T4 uptake in murine osteoblasts. (
Furthermore, we analyzed the role of Mct8 in TH transport into osteoclasts and osteocytes using established cell lines. Pharmacological Mct8 blockade led to a significantly reduced T3 uptake in osteoclast-like RAW264.7 and osteocyte-like MLO-Y4 cells when treated with 1 and 5 μM T3, respectively (Supplementary Fig. S6A, B). In line with our findings in osteoblasts, T4 import was not altered by silychristin in MLO-Y4 cells (Supplementary Fig. S6C). Interestingly, RAW264.7 cells, which were treated with 1 μM T4, displayed an even higher intracellular T4 concentration due to Mct8 blockade, suggesting that alternative transporters might compensate impaired Mct8 function or that silychristin might inhibit T4 efflux (Supplementary Fig. S6D).
Discussion
THs are essential regulators of bone homeostasis. It is well established that T3 improves osteoblast differentiation and activity, thereby enhancing bone formation (10 –14). Mice with exogenously induced hyperthyroidism have a high bone turnover state that in turn causes bone loss due to predominant bone resorption (45,46). Thus, osteoclasts are targeted by TH (3,23 –27). To exert their effects, TH requires transport proteins that mediate their transmembrane passage. Mct8 is a highly specific and efficient TH transporter that was reported to be present in osteoblasts and osteoclasts (21,30,33,34). Indeed, Mct8 global knockout mice were recently shown to have low trabecular bone mass (39). However, it remained unknown whether the increased bone turnover merely reflects the high T3 serum concentrations (40,52) or whether the absence of Mct8 in bone cells directly affects bone homeostasis. To address this question, we took advantage of conditional Mct8-knockout mice and eliminated Mct8 specifically either in osteoblast or osteoclast precursors.
Analysis of the bone phenotype of global Mct8-knockout mice revealed low bone mass in the trabecular bone compartment thereby confirming previous findings (39). However, in contrast to Leitch et al. (39), who found no differences in cortical bone parameters, our data show an increased cortical bone mass and a trend toward increased bone strength. The reason for this discrepancy is unclear but may be explained by the different ages of the animals at the time point of investigation. Nonetheless, as cortical bone volume and bone strength are reduced in mice with exogenously induced hyperthyroidism (45,46), Mct8 loss might oppose T3 excess-induced cortical bone changes suggesting a compartment-specific function of Mct8.
In the past years, cortical bone has received particular attention in bone research and is considered an important determinant of overall bone strength (53). Different molecules such as periostin (54) and Wnt16 (55) have emerged as regulators of cortical bone mass as systemic knockouts in mice resulted in decreased cortical bone, whereas trabecular bone was not changed. Further evidence for compartment-specific effects of Mct8 is provided by the osteoprogenitor-specific and osteocyte-specific knockout mice, which also revealed lower cortical bone volume (Table 3). However, more studies are needed to reliably appoint Mct8 as a potential positive regulator of cortical bone.
Overview of Skeletal Phenotypes of Global and Conditional Mct8-Knockout Models
Arrows indicate increases (↑), decreases (↓), or no changes (↔) between mutant animals and respective wild-type/Cre-negative controls.
CTX, C-terminal telopeptide; P1NP, type 1 procollagen aminoterminal-propeptide; TH, thyroid hormone.
In this study, we report a high bone turnover state in Mct8-knockout mice by identifying enhanced osteoclast and osteoblast parameters. These results are in line with our previous findings showing an accelerated bone metabolism in mice treated with TH (45,46), indicating that the phenotype of Mct8-knockout mice may depend on high systemic T3 concentrations. Besides bone, other studies have demonstrated hypermetabolism of Mct8-deficient mice mediated by organs that do not mainly depend on Mct8 for TH uptake such as skeletal muscle, kidney, and liver (41,56,57). These systemic changes might also contribute to skeletal abnormalities in Mct8-knockout mice as bone homeostasis is maintained by a tight interplay of multiple endocrine, paracrine, and autocrine factors.
Overall, the increase in bone turnover was less pronounced in Mct8-knockout mice compared to TH-treated mice (45,46). This difference might be explained by a smaller rise in systemic T3 serum levels in Mct8-knockout mice (1.5-fold) compared to TH-treated animals [3-fold (45,46)] or may be caused by the opposite alterations in serum T4 concentrations [low in Mct8-knockout mice vs. elevated in TH-treated mice (45,46)]. Taken together, our data demonstrate that systemic Mct8-deletion results in a hyperthyroidism-like high bone turnover state and delayed mineralization with subsequent trabecular bone loss. These findings also indicate that TH must enter osteoblasts, osteoclasts, and osteocytes through another TH transporter that can compensate for the absence of Mct8 as also suggested by Leitch et al. (39). This transporter yet remains to be identified. As patients with Mct8-deficiency are often not able to sit or stand independently (36 –38), and bone structure is determined by its mechanical use, bone parameters of these patients cannot be reliably correlated to Mct8 loss only.
To distinguish between effects mediated by high systemic T3 versus effects due to the lack of Mct8 in bone, we differentiated bone cells from Mct8-knockout mice ex vivo. Both, osteoblasts and osteoclasts showed highest expression of Mct8 in a more mature state suggesting an important role in terminal differentiation steps. In line with this, analysis of the late differentiation marker Ocn and mineralization at day 21 revealed lower levels in Mct8-deficient cells compared to the early differentiation marker Runx2 and mineralization that were assessed at day 10. In fact, inhibition of Mct8 in osteoblasts, osteoclast-like RAW264.7, and osteocyte-like MLO-Y4 cells resulted in impaired T3 uptake, while T4 uptake was only altered in osteoclasts. Nonetheless, Mct8-deficient osteoblasts responded to T3 treatment, suggesting that another transporter must compensate for Mct8 under high T3 conditions. Besides osteoblasts, the number of osteoclasts and expression of differentiation markers were also reduced in Mct8-deficiency. However, mixed cultures of osteoclasts with bone marrow stromal cells do not allow a definite clarification whether Mct8-knockout affects osteoclasts directly or indirectly by impaired stimulation through paracrine signals emanating from surrounding Mct8-deficient cells of the osteoblast lineage. Overall our ex vivo data indicate a cell intrinsic role of Mct8 in bone cells.
To substantiate our ex vivo findings, we conditionally knocked out Mct8 in osteoclast or osteoblast progenitors, as well as late osteoblasts/osteocytes. Importantly, TH status, body weight, and femur length of 12-week-old knockout versus control mice did not differ indicating that conditional knockout in all three cell types did not affect circulating TH levels and skeletal growth. However, our results show that both knockout lines targeting bone progenitors show an increased trabecular bone volume supporting a cell intrinsic role of Mct8 in adult bone metabolism (Table 3).
Increased trabecular bone mass in those two Mct8 mutant lines was likely caused by different cellular mechanisms. Knockout of Mct8 in osteoclast precursors reduced their differentiation capacity, similar to the ex vivo studies, and enhanced osteoblast numbers, indicating cross talk between Mct8-deficient osteoclasts and wild-type osteoblasts. Several pathways may mediate such interactions, such as the semaphorin-plexin pathway (58), or also the sphingosine-1-phosphate pathway (59,60). Further studies are necessary to investigate the underlying mechanisms. While osteoclast-specific knockout resulted in increased bone formation parameters, Mct8 appears to act in a cell intrinsic manner in osteoblasts.
Osteoblast progenitor-specific knockout mice displayed high osteoblast numbers indicating that Mct8 negatively regulates osteoblasts and bone formation, which is in contrast to our ex vivo studies. Possibly, responsiveness to T3 and T4 in vivo and in vitro and/or different knockdown efficiencies may account for the discrepancy as osteoblasts from global knockouts showed a downregulation of Mct8 expression by 100%, whereas Mct8 expression was only reduced by 60% in osteoblast-specific conditional Mct8-knockout mice (Fig. 6C). In addition, it should be noted that the Osx-Cre may target additional cell populations such as hypertrophic chondrocytes, stromal cells, adipocytes, and perivascular cells of bone marrow (61), which may contribute to differential effects. Moreover, off-target Dmp1-Cre activity was reported in skeletal muscles and certain brain areas (62).
In conclusion, our study demonstrates an essential cell intrinsic role of Mct8 in osteoblasts and osteoclasts and identifies Mct8 as a negative regulator of trabecular bone mass.
Footnotes
Acknowledgments
F.L. and M.R. designed the experiments. F.L., E.T., and M.R. performed all mouse and in vitro experiments and analyzed the data. E.R. and J.K. determined serum TH concentrations. F.L., E.T., E.R., J.K., L.C.H., H.H., and M.R. contributed to data discussion and interpretation. F.L. and M.R. drafted the article. All authors critically read, revised, and approved the final version of the article. The authors thank Ina Gloe, Tina Dybek, Maria Heier, and Eva Schubert for excellent technical assistance. In addition, the authors thank Jens Mittag, Kornelia Johann, and Sebastian Müller (Universität zu Lübeck, Lübeck, Germany) for their collaboration.
Author Disclosure Statement
M.R. reports honoraria for lectures from Amgen. L.C.H. reports honoraria for lectures and educational material from Alexion, Amgen, Merck, Radius, Roche, Shire, and UCB to his institution and himself. J.K. reported honoraria for lectures from Ipsen, Novo-Nordisk, and Sanofi/Genzyme to his institution and himself. Elena Tsourdi reports honoraria for lectures from Amgen and educational grants from UCB. The remaining authors have nothing to disclose. F.L., E.R., and H.H. have no competing financial interests.
Funding Information
This work was supported by the Deutsche Forschungsgemeinschaft Schwerpunktprogramm 1629 ThyroidTransAct (to L.C.H., J.K, H.H. (HE3418/7–2), and M.R.).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6