
Abstract
Background:
Despite active studies of the clinical importance of BRAFV600E, suitable research models to investigate the role of this mutation in the etiopathogenesis of human thyroid cancers are limited. Thus, we generated cell lines by transducing the simian virus (SV)-40 immortalized human thyroid cell line Nthy-ori 3-1 (Nthy) with lentiviral vectors expressing either BRAFWT (Nthy/WT) or BRAFV600E . Nthy/WT and Nthy/V600E cells were then xenografted into mice to evaluate the carcinogenic role of BRAFV600E.
Methods:
Each cell line was subcutaneously injected into NOD.Cg-Prkdcscid Il2rgtm1Wjl/SzJ mice, and a pathological analysis was performed. The effects of the mutation were further verified by using a BRAFV600E-selective inhibitor (PLX-4032, vemurafenib). The transcriptome was analyzed by RNA sequencing and compared with data from The Cancer Cell Line Encyclopedia and Gene Expression Omnibus.
Results:
While Nthy/WT was not tumorigenic in vivo, Nthy/V600E formed tumors reaching 2784.343 mm3 in 4 weeks, on average. A pathological analysis indicated that Nthy/V600E tumors were dedifferentiated thyroid cancer. We found metastases in the lung, liver, and relevant lymph nodes. A transcriptomic analysis revealed 5512 differentially expressed genes (DEGs) between the mutant and wild-type cell lines, and more DEGs were shared with anaplastic thyroid cancer than with papillary thyroid cancer. BRAFV600E activated the cell cycle mainly by regulating G1/S phases. PLX-4032 treatment significantly inhibited tumor growth and metastasis.
Conclusions:
Our data show that BRAFV600E plays a pivotal role in the carcinogenic transformation of an SV40-transfected immortalized normal human thyroid cell line. This xenograft model is expected to contribute to studies of the etiopathogenesis and treatment of highly malignant thyroid cancers.
Introduction
According to the National Cancer Institute (Bethesda, MD) and Cancer Research UK (London, UK), the incidence of thyroid carcinomas has increased over the past 10 years (1,2). A particularly dramatic increase has been reported in South Korea, provoking controversy regarding over-diagnosis (3). However, thyroid cancer is still highly prevalent, with the third highest incidence among all cancers in 2016 despite this controversy (National Cancer Information Center, Korea) (4).
Thyroid cancers are mainly classified based on histological phenotypes. However, recent studies based on next-generation sequencing have provided a basis for the establishment of molecular classification for new diagnostic and therapeutic approaches (5 –7). In thyroid cancers, major driver mutations, such as BRAFV600E and RAS mutations, are closely associated with specific histological types, clinical outcomes, and patterns of gene expression, that is, BRAFV600E -like, RAS-like, and Non-BRAF Non-RAS subtypes (5,6).
Although most thyroid cancers are indolent, anaplastic thyroid carcinoma (ATC) has a very poor prognosis, including a high rate of distant metastasis and frequent recurrence, resulting in almost 100% disease-related mortality (8). ATC is diagnosed based on clinical and pathologic features (8). According to The Papanicolaou Society of Cytopathology Consensus Recommendations, ATC grows rapidly, and several metastases are often found at initial diagnosis (8). Cytologic features include highly malignant cells, with a combination of spindle, giant, and squamous cells with high-grade nuclear features, such as marked pleomorphism, dark clumped chromatin, macronucleoli, and atypical mitoses. Thyroglobulin (Tg) and thyroid transcription factor 1 (TTF-1) are rarely expressed, while paired box gene 8 (PAX8) expression is usually maintained (9). Even though the Food and Drug Administration (FDA, Silver Spring, MD) recently approved a combined therapy using dabrafenib and trametinib for BRAFV600E-positive ATC, it is still not possible to completely and successfully treat patients with ATC (10).
For translational research, genetically engineered mouse models are useful for understanding the molecular mechanisms by which genetic alterations contribute to disease phenotypes, but these models cannot fully represent the characteristics of human thyroid cancer owing to differences in genomic background between species (11,12). In addition, xenograft models using human thyroid cancer cell lines have limitations for use in precision medicine owing to the complex mutational profile (13 –15). It is, therefore, necessary to establish new animal models to understand the role of individual genetic alterations in human thyroid carcinomas and to develop precision medicine approaches that consider mutational status.
In addition to driver mutations such as BRAFV600E and RAS mutations, genomic alterations in TP53, the TERT promoter, and PI3K-AKT pathways induce the dedifferentiation of well-differentiated thyroid cancers (DTCs), resulting in progression to poorly differentiated thyroid cancer (PDTC) and ATC (7,12,16,17). A recent analysis of the molecular background of 779 advanced thyroid carcinomas has shown that loss-of-function mutations in tumor suppressor genes, such as TP53 and RB1, are closely associated with ATC (7).
BRAFV600E , which is the most common mutation in thyroid cancers, is found in about 45.7% of all cases and in 48.5–80.8% of papillary thyroid cancers (PTCs) (18 –20). To study the role of BRAFV600E in thyroid cancers, our group previously developed the two cell lines Nthy/BRAFWT (Nthy/WT) and Nthy/BRAFV600E (Nthy/V600E), which have the same genetic background and differ only by the mutation status of BRAF, generated by transducing an immortalized human thyroid cell line Nthy-ori 3-1 (Nthy) with vectors expressing either BRAFWT or BRAFV600E (21). The expression of BRAFV600E induces epithelial–mesenchymal transition (EMT), including changes in both cell morphology and gene expression, resulting in the enrichment of cancer-related pathways and increased migration, invasion, and anchorage-independent growth (21). Because Nthy is the only available normal human thyroid epithelial cell line immortalized by simian virus (SV)40 transfection, we aimed at establishing a mouse xenograft model by using Nthy/V600E to investigate whether in vivo tumorigenesis of an immortalized human thyroid cell line can be induced by the BRAFV600E mutation and to establish a new in vivo research model for BRAFV600E -mutated thyroid carcinomas (22). Moreover, we also aimed at seeing whether BRAFV600E transduction of Nthy would induce PTC as Charles et al. showed thyroid-specific expression of BRAFV600E elicits murine PTC (11). SV40 is one of the oncogenic viruses expressing SV40 large T-antigen (SV40 T-ag), which is an oncoprotein binding to the DNA-binding site of p53 and the LXCXE-binding site of pRB, resulting in inactivation of p53 and pRB (23 –25). While SV40 has been widely used to immortalize cells, it has also been reported that SV40 is associated with poor prognosis in breast, prostate, and lung cancers. Therefore, we investigated the pathologic features of xenografts to see how SV40 affected Nthy/WT and Nthy/V600E.
Materials and Methods
Cell culture
Both Nthy/WT and Nthy/V600E were grown in RPMI 1640 supplemented with 10% fetal bovine serum (Biowest), 2 mM GlutaMAX™ (Gibco), and 100 U/mL penicillin-streptomycin (Gibco) in a humidified atmosphere of 5% carbon dioxide (CO2) at 37°C. Eighty percent-confluent cells were used for xenografts. RNA for RNA-sequencing was extracted after a 24-hour incubation in 100%-confluent condition, keeping the media fresh. Cell stocks with a same passage were used for each set of experiments. A Short Tandem Repeat (STR) analysis was performed by the Korean Cell Line Bank (Seoul, South Korea) by using AmpFLSTR™ identifiler™ PCR Amplification Kit (Applied Biosystems) and validated that both cell lines have the same STR profile as Nthy. The absence of mycoplasma in the cell lines was demonstrated by the Korean Cell Line Bank using e-Myco™ plus Mycoplasma PCR Detection Kit.
Animal experiments
All studies were performed under a protocol approved by an Institutional Animal Care and Use Committee at Seoul National University (SNU-171221-3-2), and mice were cared for and maintained in accordance with the recommendations of the Biomedical Center for Animal Resource Development at Seoul National University College of Medicine. Each cell line was harvested by using TrypLE™ Express (Gibco), re-suspended at 1E7 cells/mL in cold phosphate buffered saline (PBS), and kept at 4°C. The re-suspended cell suspension (100 μL) was subcutaneously injected into the right flank of 6–8 week-old male NSG mice (NOD.Cg-PrkdcscidIl2rgtm1Wjl/SzJl, The Jackson Laboratory). Tumors were measured with a caliper, and the volume of the tumors was calculated by width2 × length/2 (mm3). All injections and tumor measurements were performed under isoflurane anesthesia to minimize stress. At the end-point of experiments, mice were euthanized by using a CO2 chamber, and an autopsy was conducted to collect the primary tumors and any distinguishable metastatic tumors in the lung, liver, lymph nodes (LNs), and other subcutaneous sites. The collected tumors were fixed in 10% neutral buffered formalin solution (Sigma) for 12–24 hours and paraffin embedded. A part of the collected tumors was frozen in a deep freezer.
For drug treatment, Nthy/V600E-xenografted mice were incubated for 14 days without any treatment and treated with either 10 mg/kg of PLX-4032 (vemurafenib; STEMCELL technologies) or vehicle intraperitoneally on a daily basis from days 14 to 28. PLX-4032 was solubilized in 6% dimethyl sulfoxide, 10% Tween 80 (Sigma), and 84% PBS (v/v) at 3 mg/mL, and it was freshly prepared every day before treatment.
Histology and immunohistochemistry
For histological analysis, the formalin-fixed paraffin-embedded (FFPE) tissues were sectioned at a thickness of about 4–5 μm, stained with hematoxylin and eosin (H&E) by using Autostainer XL (Leica), and immunohistochemistry (IHC) analysis was performed with commercially available antibodies against BRAFV600E (clone VE1, ready-to-use, mouse monoclonal; Ventana), PAX8 (clone MRQ-50, 1:100, mouse monoclonal; Cell Marque), Ki-67 (clone MIB-1, 1:100, mouse monoclonal; DAKO), p53 (clone DO-7, mouse monoclonal; DAKO), TTF-1 (clone SP141, ready-to-use, rabbit monoclonal; Ventana), and Tg (A 0251, 1:30,000, rabbit polyclonal; DAKO) by the department of pathology at Seoul National University Hospital. The results of H&E and IHC analysis were examined by a specialized pathologist at Seoul National University Hospital.
RNA sequencing
Three biological replicates were prepared for each cell line, which were cultured for 4–5 days while keeping media fresh to place them under a contact inhibition. RNA was extracted by using RNeasy Mini Kit (QIAGEN) and assessed for quantity and quality by using 2100 Bioanalyzer (Agilent Technologies). The sequencing libraries were prepared by using TruSeq Stranded mRNA LT Sample Prep Kit (Illumina) following the manufacturer's recommendation and sequenced on a NovaSeq 6000 platform (Illumina) by Macrogen (South Korea). The sequenced paired-end reads were aligned to UCSC hg19 by using HISAT2 (26), and StringTie (27) was used for transcript assembly.
To confirm the presence of SV40 T-ag expression, the sequenced reads were also aligned to SV40gp6 sequence (NC_001669.1).
Comparative analysis of transcriptome
To compare transcriptome of our cell lines with other established cell lines and human specimens, we utilized The Cancer Cell Line Encyclopedia (CCLE) database (Broad Institute, Cambridge, MA) and Gene Expression Omnibus database (GSE33630).
In the CCLE database, we used reads per kilobase of transcript per million mapped reads (RPKM) data of 9 human thyroid cancer cell lines: 4 ATC (8305c, 8505c, BHT101, CAL62), 1 PDTC (BCPAP), 4 FTC (TT2609C02, FTC238, FTC133, ML1). In GSE33630, we downloaded complementary DNA microarray data of human frozen specimens: 11 of ATC, 49 of PTC, and 45 of normal thyroid tissues.
Mean fragments per kilobase of transcript per million mapped reads (FPKM) of our cell lines was analyzed together with RPKM of the nine cell lines from CCLE to show gene expression associated with thyroid differentiation and cancer prognosis. For heatmap display, gene expression data were illustrated based on row z-score of median-centered log2 RPKM (FPKM), and a thyroid differentiation score (TDS) was calculated with the mean value of the row z-scores across 14 thyroid metabolism and function genes excluding PAX8, which is known to be usually conserved even in ATC.
Differentially expressed genes (DEGs) between Nthy/WT and Nthy/V600E were compared with those of ATC-normal tissues and PTC-normal tissues to confirm that Nthy/V600E corresponds more closely to ATC than well-DTCs. DESeq2 package was used to select the DEGs meeting the following criteria: twofold change or fourfold change with adjusted p-value (Q-value) <0.01.
Statistical analysis
All statistical analyses were performed by using GraphPad PRISM version 8.0.0 (GraphPad Software, San Diego, CA). Data are presented as mean ± standard deviation (SD). The independent t-test, paired t-test, or Mann–Whitney test (n < 10) were used for comparison of continuous variables. Statistical significance was defined as two-sided p-values <0.05.
Results
BRAFV600E transduction initiates the carcinogenesis of an SV40-immortalized normal thyroid cell line
To investigate the role of BRAFV600E in the carcinogenesis of this immortalized human thyroid cell line, we xenografted both Nthy/WT and Nthy/V600E into NSG mice. Nthy/V600E formed a distinguishable tumor in all xenografted mice (11 out of 11) in a week. Mice showed rapid tumor growth (2784.343 mm3, on average, n = 11, SD = 922.463 mm3, 4 weeks) and dramatic weight loss (29.49 g on day 24 to 27.79 g on day 28, n = 11, p = 0.0038, two-tailed paired t-test), reaching a humane end-point in 4 weeks (Fig. 1A, B, D). The end-point tumor weight was 1.941 g (n = 11, SD = 0.569 g) (Fig. 1C). Nthy/WT, however, did not result in tumor formation or measurable changes, even after 6 weeks of observation, that is, 2 weeks after euthanizing Nthy/V600E-xenografted mice (n = 11). Based on an autopsy, a small mass of Nthy/WT was found under the skin at the injection site, covered by a fat layer (Fig. 1A). The mean volume of the masses was 4.892 mm3 (n = 10, SD = 2.035 mm3, 6 weeks), and the weight was unmeasurable (marked as 0.01 g) (Fig. 1B, C). These results suggest that BRAFV600E plays a crucial role in the carcinogenic transformation of this immortalized human thyroid cell line.
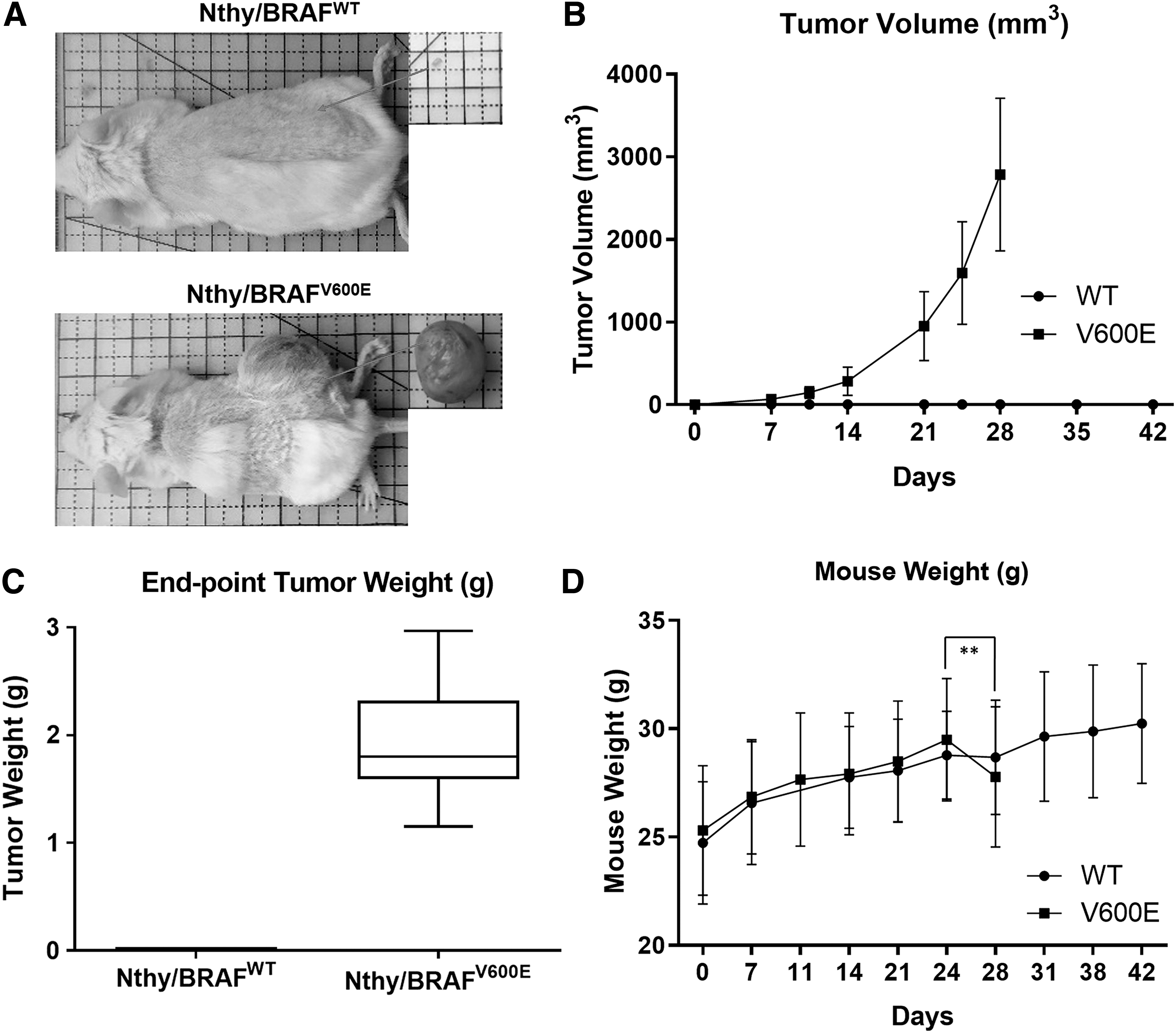
In vivo tumorigenesis of an SV40-immortalized normal human thyroid epithelial cell line initiated by BRAFV600E
transduction. (
BRAFV600E transduction induces dedifferentiated thyroid carcinogenesis of an SV40-immortalized normal thyroid cell line
We confirmed that both Nthy/WT and Nthy/V600E are expressing SV40gp6 (NC_001669.1) by using RNA-sequencing data, which encodes SV40 T-ag-inactivating p53 and pRb (data not included) (28).
We performed H&E staining of FFPE samples to determine the histologic features of Nthy/V600E and Nthy/WT xenografts. Nthy/V600E showed a highly malignant cell morphology with marked pleomorphic- and macro-nuclei, frequent mitosis, and intranuclear inclusions (Fig. 2A). Nthy/WT showed monomorphic nuclei and a relatively high fraction of murine stromal cells (Fig. 2A).
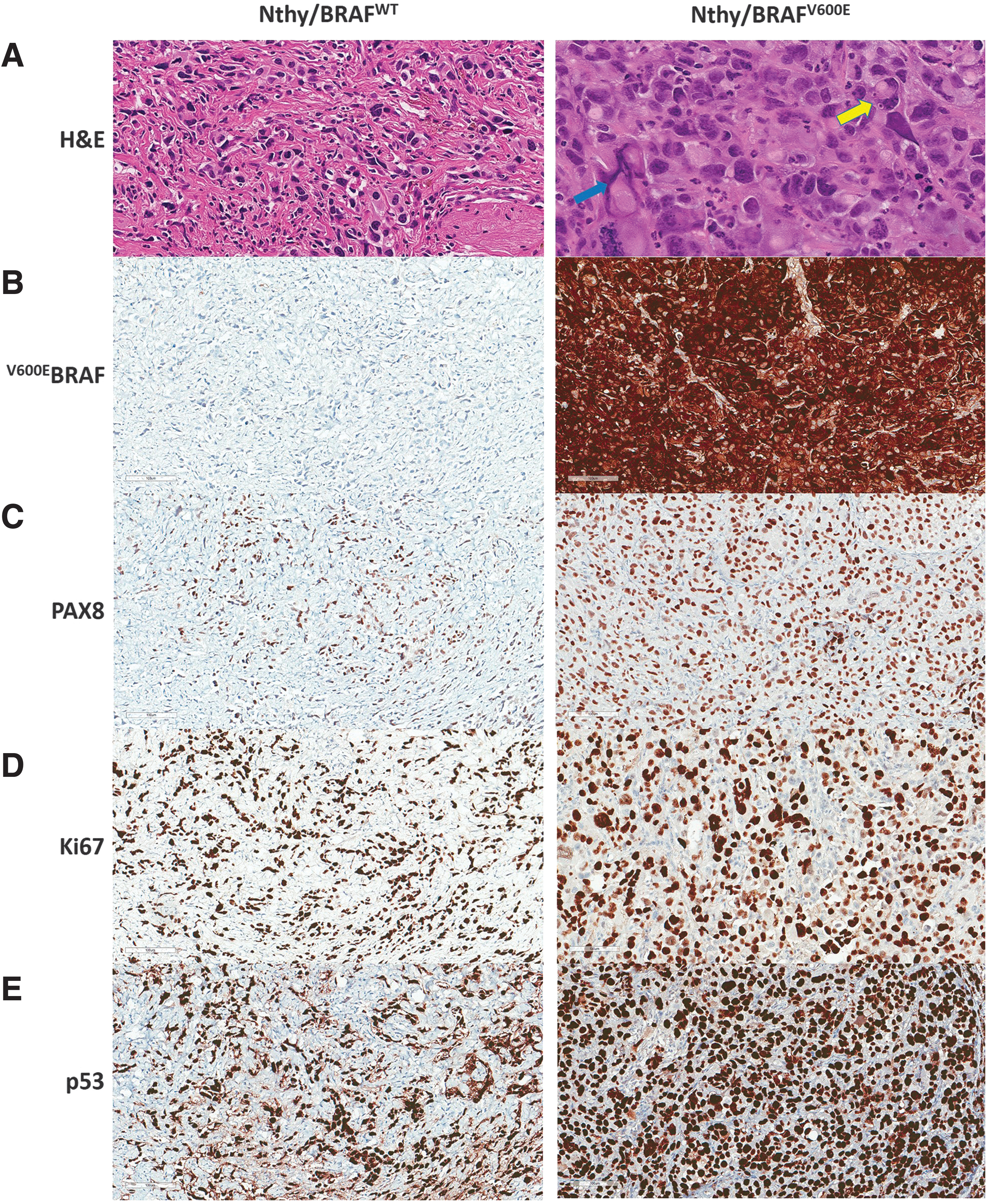
Histological and immunohistochemical phenotypes of SV40-immortalized thyroid epithelial cells depending on BRAFV600E status. (
Cytoplasmic expression of BRAFV600E in Nthy/V600E was confirmed by IHC using an anti-BRAFV600E (VE1) antibody (Fig. 2B). Tg and TTF-1 were negative in both Nthy/WT and Nthy/V600E (data not included), but PAX8 was positive in both, confirming that the fraction of Nthy/WT cells was low in the Nthy/WT xenograft tissues (Fig. 2C). p53 expression was very high in both Nthy/WT and Nthy/V600E, probably due to structural stabilization by SV40 T-ag (Fig. 2E).
Ki67 expression was analyzed to evaluate in vivo proliferation. In accordance with the in vivo tumor growth rate, Nthy/V600E showed high expression of Ki67 (Fig. 2D). However, even though Nthy/WT did not demonstrate proliferation in vivo, it also showed high Ki67 expression (Fig. 2D). A pathologic review of Nthy/V600E tumors by a specialized pathologist indicated dedifferentiated thyroid carcinoma (dDTC), not PTC. This means that SV40 in this immortalized human thyroid cell line might contribute to the dedifferentiation of BRAFV600E -induced thyroid cancer.
BRAFV600E expression leads to metastasis of SV40-immortalized thyroid cells
The presence of BRAFV600E in PTC can be associated with poor prognosis; accordingly, we investigated metastasis in organs and anatomically relevant LNs based on the primary injection site (29). After humane euthanization, we performed an autopsy to find visible metastatic tumors. In all Nthy/V600E-xenografted mice, we found metastatic LNs in both (or either) the sciatic LN (SaLN) and (or) the subiliac LN (SiLN), with a few exceptions in which a primary tumor was big enough to invade the LN region (data not included). We also found metastasis in the proper axillary LN (PALN) and (or) the accessory axillary LN (AALN) in 72.7% (8/11) of cases but not in the left axillary LNs (Fig. 3A, B). However, we could not find any evidence of metastasis to LNs in Nthy/WT-xenografted mice, and this result was confirmed by H&E staining (Fig. 3A).

Metastatic characteristics of SV40-immortalized thyroid epithelial cells are induced by BRAFV600E (
In the Nthy/V600E group, metastasis was found not only in distinguishable metastatic LNs but also in suspicious and indistinguishable right axillary LNs (based on size), as determined by an H&E analysis (Fig. 3A). Metastatic LNs were found in 72.7% (8/11) cases in the right axillary LN(s) and showed highly pleomorphic nuclei, frequent mitosis, and intranuclear inclusion (Fig. 3A). In an H&E analysis of suspicious and indistinguishable LNs, the right axillary LN(s) were dotted with or overwhelmed by metastasis of Nthy/V600E, depending on the size; similar results were not obtained for the left axillary LN(s) (data not included). Moreover, we found distant metastases in the lung (66%, 2/3) and the liver (33%, 1/3) in an additional experiment after 6 weeks of incubation (Fig. 3C, D). Based on these data, we conclude that BRAFV600E transduction enables SV40-immortalized thyroid cells to metastasize to both relevant LNs and distant organs.
Nthy/V600E shows similar transcriptomic patterns with ATC
To determine whether the transcriptomic features of the transformed cell lines are similar to ATC, we compared the transcriptomes of the cell lines with those of other thyroid cancer cell lines and human specimens available in CCLE and GSE33630.
In a DEG analysis of Nthy/V600E over Nthy/WT, we detected 5512 total twofold DEGs (1473 upregulated and 4039 downregulated genes, adjusted p-value <0.01) and 3526 fourfold DEGs (627 upregulated and 2899 downregulated genes, adjusted p-value <0.01).
When we compared the DEGs with those between ATC and normal tissues in GSE33630, 305 shared DEGs were upregulated by at least twofold (56 genes, fourfold) and 557 genes were downregulated by at least twofold (145 genes, fourfold) (Fig. 4A, B). In a gene ontology analysis, the shared upregulated DEGs (fourfold) were involved in cell migration, proinflammatory signals, immune cell chemotaxis, and extracellular structure organization (Supplementary Fig. S1; Supplementary Table S1). The shared downregulated DEGs (fourfold) were associated with the following terms: development and morphogenesis of gland, regulation of osteoblast differentiation, detoxification, inorganic anion transmembrane transport, thyroid hormone generation and metabolic process, hydrogen peroxide metabolic and catabolic process, and apoptotic process (Supplementary Fig. S2; Supplementary Table S2).
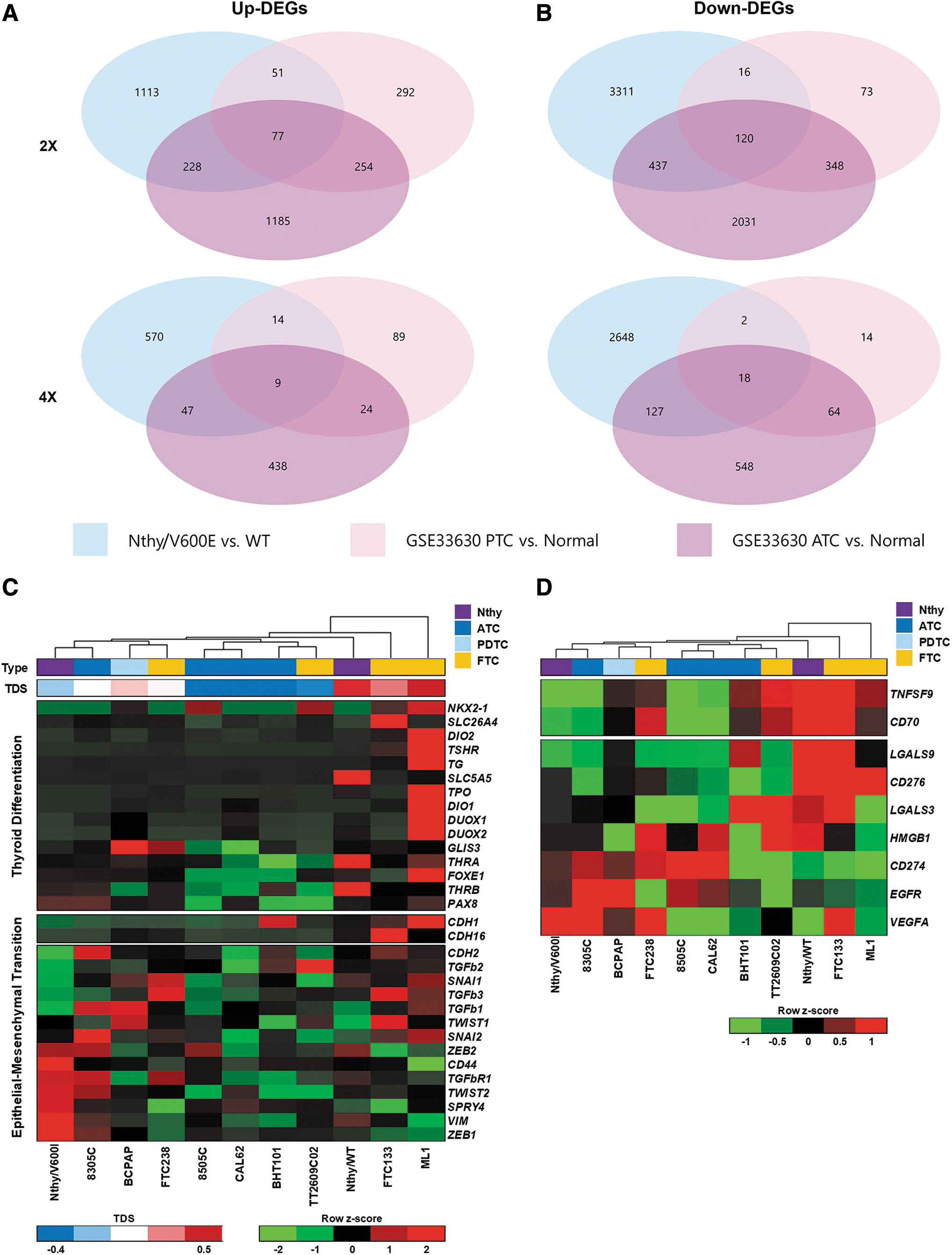
Transcriptomic pattern of Nthy/V600E is closer to profiles seen in anaplastic thyroid cancer. (
Shared DEGs between Nthy/V600E-Nthy/WT and PTC-normal tissues pairs included 128 upregulated genes over twofold (23, fourfold) and 136 downregulated genes over twofold (20, fourfold), far fewer than those shared with ATC-normal tissues (Fig. 4A, B). The shared upregulated DEGs were related to extracellular structure organization, endoderm formation and development, cell adhesion to matrix and substrate, negative regulation of MAPK signaling, and immune cell chemotaxis, while the shared downregulated DEGs were involved in thyroid hormone generation and metabolic processes (Supplementary Figs. S3 and S4; Supplementary Tables S3 and S4).
We focused on the expression of genes associated with poor clinical outcomes in the studied cell lines, such as genes related to thyroid differentiation, EMT, and cancer prognosis (e.g., immune checkpoint genes, EGFR, and VEGF-A). The thyroid differentiation-related gene expression pattern for Nthy/V600E was similar to those of ATC cell lines, and the TDS for Nthy/V600E was lower than that for 8305c, an ATC cell line carrying pathogenic mutations in TP53 and TERT promoters as well as in BRAF (Fig. 4C). However, Nthy/WT showed a higher TDS than that of Nthy/V600E, which is similar to FTC133, but a lower score than that of ML1 (Fig. 4C). Although Nthy/WT is not fully differentiated, it expressed the highest levels of SLC5A5 (NIS) and thyroid hormone receptor beta among the cell lines and showed similar or higher expression levels of all of the target genes, except PAX8, than those of Nthy/V600E (Fig. 4C).
Moreover, Nthy/V600E showed similar patterns of EMT-related gene expression with those of 8305c (Fig. 4C). It expressed lower levels of epithelial genes, such as CDH1 and CDH16, and higher levels of mesenchymal genes, such as ZEB1/2, CD44, TGFbR1, SPRY4, VIM, TWIST1/2, and SNAI2, even though several mesenchymal genes (e.g., SNAI1, CDH2, and TGFb family genes) were expressed at lower levels than those in Nthy/WT (Fig. 4C).
We selected nine genes associated with cancer prognosis for further analysis (excluding genes with zero or near-zero values in all cell lines); these protein-coding genes have gained attention owing to their implications for cancer immunotherapies, for example, immune-stimulatory genes, including TNFSF9 (4-1BBL via 4-1BB) and CD70 (CD70 via CD27), immune-inhibitory genes including LGALS3 and 9 (Galectin3 and 9 via TIM-3 and LAG-3), CD276 (B7-H3 via CD28 family), and HMGB1 (HMGB1 via TIM-3), and growth factor pathway genes, such as EGFR (EGFR) and VEGFA (VEGF-A). Interestingly, the gene expression pattern in Nthy/V600E was similar to those of 8305c and 8505c, while Nthy/WT showed essentially the opposite pattern, similar to the expression of genes related to thyroid differentiation and EMT (Fig. 4D). These findings confirm that BRAFV600E in conjunction with SV40 induces a highly malignant and aggressive form of thyroid cancer showing ATC-like transcriptomic features.
BRAFV600E activates G1/S phase of SV40-transfected immortalized thyroid cells
To investigate the mechanism by which BRAFV600E enables SV40-transfected immortalized human thyroid cell line to form tumors in vivo, we analyzed the expression of cell cycle-related genes in Nthy/WT and Nthy/V600E cultured until reaching confluence on a culture dish (Fig. 5A) (30 –36).

Regulation of cell cycle-associated gene expression by BRAFV600E in SV40-transfected immortalized human thyroid cells. (
The inhibition of p53 and pRb by oncogenic viruses, such as human papillomavirus (HPV), can result in the overexpression of p16INK4A and p21Cip1/WAF1 and the downregulation of cyclin D1; Nthy/WT showed similar expression patterns for these genes, which were distinctly different from those of other cell lines (Supplementary Fig. S5A–C) (37,38). However, BRAFV600E transduction dramatically altered the overall expression levels of cell cycle-related genes (Fig. 5B). All of the major components that are important in G1 phase progression, such as CDK4, CDK6, CCND1, and CCNE1 (but not CDK2), were increased. CDKN1A and CDKN2A, which encode the key G1/S phase arrest proteins p21 and p16, were dramatically downregulated (Fig. 5A, B). G2 phase was relatively activated in Nthy/WT showing upregulated CDC2, CCNB1, and CDC25 family genes with a slight decrease in WEE1 (Fig. 5A, B). These findings imply that G1/S phases regulators such as CDKN2A and CDKN1A are strongly associated with the underlying mechanism of BRAFV600E-induced carcinogenesis.
Treatment with the BRAFV600E-selective inhibitor PLX-4032 inhibits the growth and metastasis of Nthy/V600E
To confirm that in vivo tumorigenesis and metastasis in Nthy/V600E is induced by BRAFV600E, we intraperitoneally injected mice with 10 mg/kg PLX-4032, which is a BRAFV600E-selective inhibitor with FDA approval for BRAFV600E-positive late-stage melanoma, daily from days 14 to 28 (Fig. 6A).
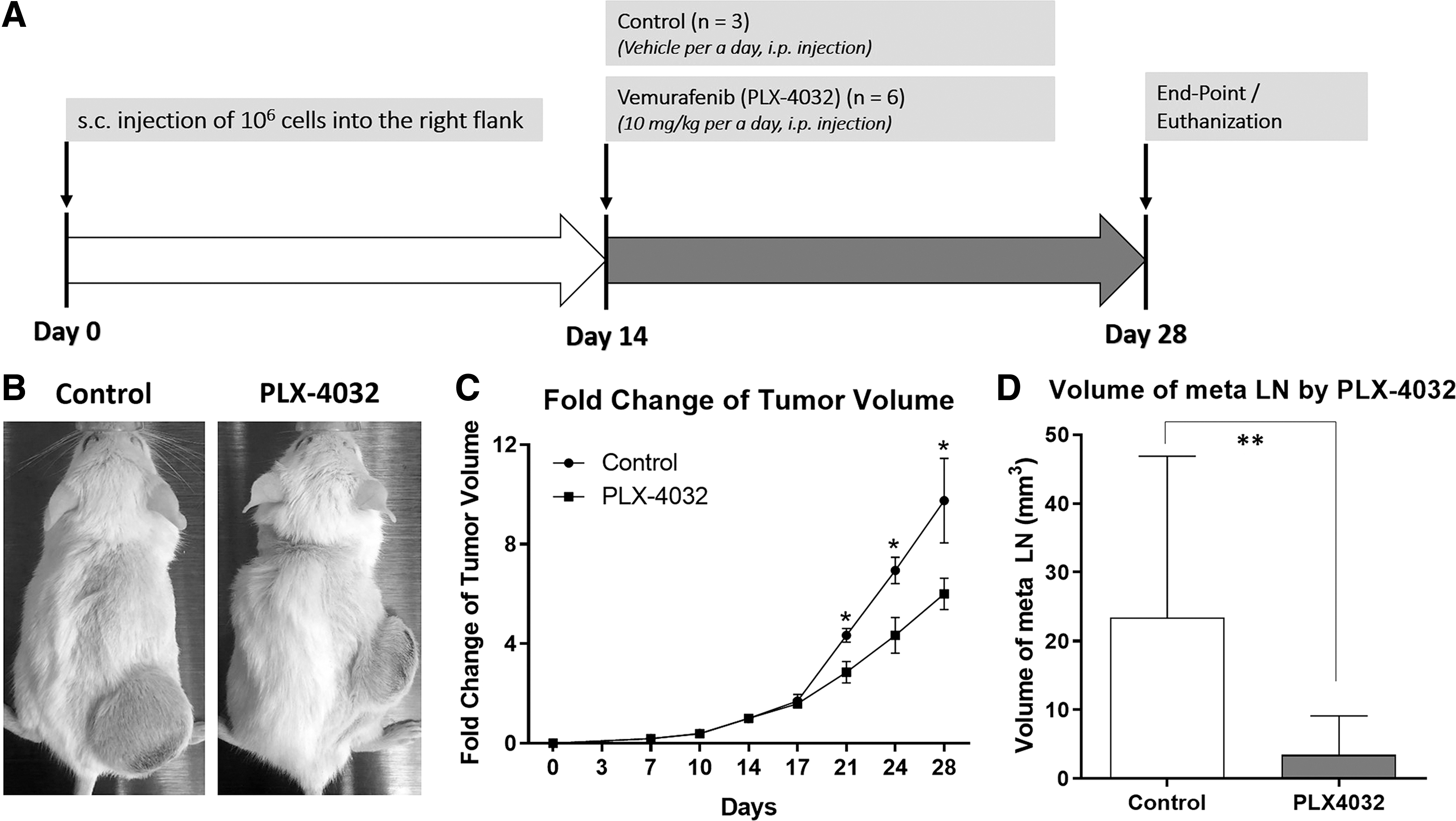
Inhibition of tumor growth and metastasis of Nthy/V600E by the BRAFV600E-selective inhibitor Vemurafenib (PLX-4032). (
Compared with the vehicle-treated group (n = 3), the mean volume at the end-point was not significantly different from that of the 11 mice in the nontreated group, and the growth rate of the PLX4032-treated group (n = 6) was significantly lower (p < 0.05) (Fig. 6B, C). However, in a pathologic review by a specialized pathologist, no regions of apoptosis in response to PLX-4032 treatment could be found on H&E histology (data not included).
Treatment with PLX-4032 significantly reduced the volume of metastatic LNs (Fig. 6D). Even though evidence for metastasis was found regardless of PLX-4032 treatment, possibly due to the two-week incubation period before treatment, the volume of metastatic LNs was significantly smaller in the PLX4032-treated group (n = 5, 3.43 mm3) than in the nontreated and vehicle-treated group (n = 11, 23.38 mm3) (p = 0.0087, Mann–Whitney test) (Fig. 6D). These results support the pivotal role of BRAFV600E in thyroid carcinogenesis as well as metastatic potential.
Discussion
We successfully established a new xenograft model for BRAFV600E-positive human dDTC by transducing an SV40-transfected immortalized human thyroid cell line with BRAFV600E . This model provides evidence that BRAFV600E is sufficient to induce the carcinogenic transformation of this immortalized human thyroid cell line. It was previously reported that Nthy is not tumorigenic in immunodeficient nude mice; consistent with this, Nthy/WT was not tumorigenic, even in NSG mice, indicating that SV40 transfection of normal human thyroid cells was not able to induce in vivo tumorigenesis (22). However, BRAFV600E transduction enabled Nthy to form a tumor and metastasize to both relevant LNs and distant sites in vivo. The xenograft tumors of Nthy/V600E had a pathologic phenotype consistent with dDTC, with little or no expression of thyroid differentiation markers, such as Tg and TTF-1. In addition, Nthy/V600E was more similar at the transcriptome level to ATC than to PTC when we analyzed the expression of 40 genes associated with cancer prognosis together with public transcriptome data for thyroid cancer tissues and cell lines.
As a human dDTC xenograft model for preclinical studies, our model has several distinct benefits. First, our model could be easily adapted to evaluate the subcutaneous microenvironment of mice without any supportive materials like Matrigel by active vascularization. Second, Nthy/V600E tumors grew rapidly, reaching a mean tumor volume of about 2800 mm3 in a month. When we xenografted the human ATC cell line 8505c in the same conditions, Nthy/V600E grew much faster (data not included). Finally, the transformed cells metastasized to both relevant LNs and distant organs in six weeks. After the inoculation of Nthy/V600E, metastasis was initially found in the right SaLN and SiLN, followed by the PALN and AALN. Four weeks were sufficient for Nthy/V600E to metastasize to the PALN and AALN from a primary tumor. Metastasis was only found in the LNs on the right side of mice. From an anatomical perspective, the primary injection site, the right flank, is between the SaLN and the SiLN (39). In the lymphatic drainage system in this region, the lymph flows from the SaLN to the PALN through the AALN and from the SiLN directly to the PALN (40,41). This would explain the occurrence of metastatic LNs in the right axillary LNs but not on the left side. Distant metastasis could be found after six weeks in xenograft models. They were first detected in the lung, followed by the liver, as observed during an autopsy at eight weeks. Considering that metastasis and rapid growth are the most important determinants of cancer-mediated mortality, these characteristics provide a model for preclinical tests of anticancer therapies.
Our results support several previous findings regarding the initiation and progression of thyroid cancers. Charles et al. proved that BRAFV600E alone can induce murine thyroid carcinogenesis, and our results confirm the pivotal role of BRAFV600E (11). Further, the findings presented here emphasize the importance of p53 and pRB in the progression to highly malignant and aggressive cancer subtypes, such as ATC. According to McFadden et al., the deletion of p53 induces the anaplastic transformation of BRAFV600E-initiated murine PTC, while BRAFV600E induces PTCs (12). Pozdeyev et al. have shown that mutations in TP53 and RB1 are significantly associated with ATC (7). Considering that SV40 inactivates p53 and pRB, it is not surprising that Nthy/V600E, which is a normal thyroid epithelial cell line expressing BRAFV600E and SV40, formed dDTC, rather than PTC, in vivo.
Previous studies have revealed that SV40 is associated with prognosis in thyroid cancers as well as other cancers. Deeb et al. (42) reported that SV40 is strongly associated with poor prognosis in aggressive human breast, prostate, and lung carcinomas, and a viral transcriptome study by Wang et al. (43) revealed that SV40-positive low-grade glioma have a significantly shorter survival than that of SV40-negative tumors. Surprisingly, the presence of SV40 even in thyroid cancer tissues has been reported by Vivaldi et al. (44); SV40 T-ag messenger RNA was detected at a similar frequency in both PTC and ATC (69% and 73%, respectively), but the SV40 T-ag protein was found more frequently in ATC than in PTC (33% of PTC and 100% of ATC) (44). Taking our results together with previous findings, SV40 has the potential to be utilized as a prognostic marker and therapeutic target for BRAFV600E-mutated thyroid cancers. Further research is required to utilize the detection of SV40 T-ag in combination with BRAFV600E for clinical purposes.
Transcriptome analyses of our models could provide insight into BRAFV600E-induced carcinogenesis. Although Nthy is an immortalized normal thyroid cell line and is not well differentiated due to SV40 transfection, it was not tumorigenic in immunodeficient NSG mice and showed highly upregulated CDKN1A and CDKN2A expression (which encode the tumor suppressors p21 and p16, respectively) compared with levels in other cancer cell lines, including Nthy/V600E (Fig. 1A and 4C; Supplementary Fig. S5A, B). Interestingly, similar results have been found in HPV-infected tonsillar squamous cell carcinoma (TSCC) (38). Like SV40, HPV is a well-known oncogenic virus that causes cancer or precancerous lesions in multiple regions by inhibiting p53 and pRb via HPV E6 and E7 oncoproteins (45). According to Hafkamp et al., TSCC shows significantly higher expression of p21 and p16 in HPV-infected cases and p21 overexpression is strongly associated with a good prognosis (38). Considering these results, p21 and p16 are likely to be closely associated with the cellular defense mechanism against oncogenic viruses that inhibit p53 and pRb. However, BRAFV600E transduction enabled Nthy/WT to form a tumor in vivo and dramatically decreased CDKN1A and CDKN2A expression to similar levels in other cancer cell lines (Supplementary Fig. S5A, B). p21 is rarely detected in ATC but not in well-DTCs, and BRAFV600E inhibition upregulates CDKN1A in BRAFV600E-mutated thyroid cancer cell lines (46 –48). Taken together, CDKN1A and CDKN2A are likely key molecules in BRAFV600E-induced carcinogenesis.
While we characterized a new xenograft model of human dDTC, two major limitations remain. First, the current paradigm in cancer treatment is mainly based on host immunity. Vanden et al. have developed an immunocompetent murine ATC model, with applications to the preclinical evaluation of onco-immunotherapies for ATC (49). Our model is not suitable for immune-oncology studies, but the use of humanized mice might overcome this limitation (50,51). Second, using this model, we could not fully demonstrate that BRAFV600E alone can induce carcinogenesis in normal human thyroid cells. Since we have not shown that BRAFV600E is solely able to transform normal human thyrocytes without other pathogenic alterations such as SV40 transfection, we need to confirm that SV40-inhibited Nthy/V600E would be tumorigenic in vivo and form human PTC, not dDTC.
Despite these limitations, our xenograft model is a promising tool for drug discovery, considering the aggressive behavior in vivo and mutational simplicity. The rapid growth and active metastasis without Matrigel supplementation in the ectopic xenograft model are beneficial for effective drug screening. Moreover, as Landa et al. recently characterized the genomic alterations and transcriptome of 60 authenticated thyroid cell lines for more rational mechanism-based research, the well-defined in vitro, in vivo, genomic, and transcriptomic features of our model are expected to facilitate well-designed pre-clinical studies (15). Among currently existing human thyroid cancer cell lines, Nthy/V600E is possibly the only one that shows anaplastic characteristics without any well-studied genomic alterations related to anaplastic transformation of thyroid cancers such as TP53 and TERT promoter mutations; it might, therefore, be a useful tool to study the molecular pathogenesis of thyroid cancers by introducing further modifications (15). For example, genome editing using the CRISPR-Cas9 system is now feasible for knock-in and knock-out studies of prognosis-related gene alterations and the SV40 genome to predict clinical outcomes and develop individualized therapeutic strategies for patients with thyroid cancer based on mutation status. Indeed, in genetic profiles of 196 ATCs screened by Pozdeyev et al. in 2019, some of the BRAFV600E-positive and -negative ATCs showed a relatively low mutation burden and did not harbor any common genomic alterations found in others (7). Considering those cases, more studies are required for presurgical diagnosis and precision medicine of thyroid cancers, and this model might be a well suited to characterize them.
In conclusion, our data show that BRAFV600E plays a crucial role in the induction of carcinogenesis in a immortalized SV40-transfected human thyroid cell line by activating ATC-like cell signaling pathways. Although p53 and pRb inhibition by SV40 T-ag might also contribute to carcinogenesis in this model, BRAFV600E is clearly essential for in vivo tumorigenesis by activating G1 phase and promoting progression to S phase. Therefore, we expect this mouse xenograft model of human dDTC mediated by BRAFV600E and SV40 to contribute to further research on the etiopathogenesis of thyroid carcinomas and the development of targeted therapies for patients with BRAF-mutated thyroid cancer.
Footnotes
Author Disclosure Statement
All authors certify that they have no competing financial interests pertaining to any of the data or statements given in this article.
Funding Information
This work was supported by the National Research Foundation of Korea (NRF) grants funded by the Korea government (MSIP) (Grant Nos. 2018R1C1B5045216 and 2017R1D1A1B03030073) and the Seoul National University Hospital (SNUH) Research Fund (Grant No. 03-2017-0190).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4