
Abstract
Background:
Glycine is a classical neurotransmitter that has role in both inhibitory and excitatory synapses. To understand whether glycinergic inputs are involved in the regulation of the hypophysiotropic thyrotropin-releasing hormone (TRH) neurons, the central controllers of the hypothalamic-pituitary-thyroid axis, the glycinergic innervation of the TRH neurons was studied in the hypothalamic paraventricular nucleus (PVN).
Methods:
Double-labeling immunocytochemistry and patch-clamp electrophysiology were used to determine the role of glycinergic neurons in the regulation of TRH neurons in the PVN. Anterograde and retrograde tracing methods were used to determine the sources of the glycinergic input of TRH neurons.
Results:
Glycine transporter-2 (GLYT2), a marker of glycinergic neurons, containing axons were found to establish symmetric type of synapses on TRH neurons in the PVN. Furthermore, glycine receptor immunoreactivity was observed in these TRH neurons. The raphe magnus (RMg) and the ventrolateral periaqueductal gray (VLPAG) were found to be the exclusive sources of the glycinergic innervation of the TRH neurons within the PVN. Patch-clamp electrophysiology using sections of TRH-IRES-tdTomato mice showed that glycine hyperpolarized the TRH neurons and completely blocked the firing of these neurons. Glycine also markedly hyperpolarized the TRH neurons in the presence of tetrodotoxin demonstrating the direct effect of glycine. In more than 60% of the TRH neurons, spontaneous inhibitory postsynaptic currents (sIPSCs) were observed, even after the pharmacological inhibition of glutamatergic and GABAergic neuronal transmission. The glycine antagonist, strychnine, almost completely abolished these sIPSCs, demonstrating the inhibitory nature of the glycinergic input of TRH neurons.
Conclusions:
These data demonstrate that TRH neurons in the PVN receive glycinergic inputs from the RMg and the VLPAG. The symmetric type of synaptic connection and the results of the electrophysiological experiments demonstrate the inhibitory nature of these inputs.
Introduction
Glycine is a classical neurotransmitter that has an important role in both inhibitory and excitatory synapses (1). Glycinergic neurons are primarily located in the brain stem and spinal cord (2), however, glycinergic axons are also observed in the hypothalamus (3). Two glycine-transporter subtypes (GlyT) were identified in the central nervous system: glycine transporter-1 (GlyT1), mainly located in glial cells and nerve terminals of excitatory neurons, and glycine transporter-2 (GlyT2), present exclusively in glycinergic neurons (4,5).
Glycine is released from inhibitory, presynaptic terminals and binds to strychnine-sensitive glycine receptors (GlyRs) that are located in the postsynaptic membrane of target cells (1). Activation of GlyRs leads to influx of chloride ions into the cytoplasm and thereby inhibits the postsynaptic neuron (6). In addition, glycine is also important for excitatory glutamatergic neurotransmission because it serves as an essential coagonist of glutamate at N-methyl-aspartate receptors (7).
The hypothalamic paraventricular nucleus (PVN), where hypophysiotropic thyrotropin-releasing hormone (TRH)-synthesizing neurons, the main central regulators of the hypothalamic-pituitary-thyroid (HPT) axis reside (8), is densely innervated by glycinergic axons (3). Within the PVN, glycine has been shown to elicit large, inward currents (9). Furthermore, evoked glycinergic currents can be observed in parvocellular PVN neurons in the presence of the blockers of glutamate and GABA receptors (9), suggesting that ascending glycinergic pathways regulate parvocellular neurons in the PVN.
Based on these data, we hypothesized that glycinergic brain stem neurons are involved in the regulation of the TRH neurons in the PVN, including the hypophysiotropic TRH neurons. To test this hypothesis, we performed neuroanatomical and electrophysiological experiments. Since TRH neurons are dispersed in the PVN, identification of these cells was difficult in electrophysiological studies. In addition, in morphological studies, the cell bodies of TRH neurons could only be identified after inhibition of axonal transport by colchicine treatment. Therefore, to facilitate both the electrophysiological and morphological studies, we generated a TRH-IRES-tdTomato knockin mouse line in which the TRH neurons can be easily identified based on the presence of the red fluorescent protein in their perikarya.
Materials and Methods
Animals
The experiments were carried out in adult, male, CD1 mice (n = 8), GlyT2::GFP mice (n = 9) (3), TRH-IRES-tdTomato mice (n = 30), double transgenic mice heterozygous for TRH-IRES-tdTomato and GlyT2::Cre (10) (TRH-IRES-tdTomato//GlyT2::Cre; n = 10) weighing 30–40 g, housed under standard environmental conditions (light between 06:00 and 18:00 hours, temperature 22°C±1°C, rat chow and water ad libitum). The in vitro patch-clamp electrophysiology studies were performed on mice between P40 and P60 days of age. All experimental protocols were reviewed and approved by the Animal Welfare Committee at the Institute of Experimental Medicine of the Hungarian Academy of Sciences.
Generation of the TRH-IRES-tdTomato mouse line
The TRH-IRES-tdTomato mouse line was generated using CRISPR/Cas9 technology (11). The details of the methods used for the generation and characterization of the mouse line are described in the Supplementary Materials and Methods.
Generation of a sheep antiserum against tdTomato
Antiserum against tdTomato was generated in sheep using recombinant His-tagged tdTomato as antigen. The generation of the antiserum is described in detail in the Supplementary Materials and Methods.
Animal preparation for examination of the glycinergic input to TRH neurons in the PVN
CD1 mice (n = 8) were deeply anesthetized with ketamine/xylazine (ketamine 50 mg/kg, xylazine 10 mg/kg body weight, intraperitoneal [ip]) and the animals perfused transcardially with 10 mL 0.01 M phosphate-buffered saline (PBS), pH 7.4, followed sequentially by 50 mL of 2% paraformaldehyde (PFA)/4% acrolein in 0.1 M phosphate buffer (PB), pH 7.4, and then by 20 mL of 2% PFA in the same buffer. The brains were rapidly removed and stored in 4% PFA for two hours for light microscopy (n = 4) at room temperature (RT), or for 24 hours for electron microscopy (n = 4) at 4°C. The sectioning and section preparation for light and electron microscopic studies are described in the Supplementary Materials and Methods.
Double-labeling immunofluorescence for examination of the glycinergic input of TRH neurons in the PVN
Pretreated sections of CD1 mice were incubated in a mixture of rabbit anti-GlyT2 IgG (12,13) and sheep anti-TRH IgG (1:4000) (12) in PBS containing 2% normal horse serum (NHS) and 0.2% sodium azide (antiserum diluent) for two days at 4°C. After rinses in PBS, the sections were incubated in a mixture of Alexa 555-conjugated donkey anti-rabbit IgG (1:500; Jackson ImmunoResearch Labs, West Grove, PA) and Alexa 488-conjugated donkey anti-sheep IgG (1:500; Jackson ImmunoResearch Labs) in antiserum diluent. After rinses in PBS, the sections were mounted onto glass slides and coverslipped with the Vectashield mounting medium (Vector Laboratories, Inc., Burlingame, CA).
Double-labeling immuno-electron microscopy for examination of the glycinergic input of TRH neurons in the PVN
Pretreated sections were placed into a mixture of rabbit anti-GlyT2 IgG (1 μg/mL) and sheep anti-TRH IgG (1:1000) for four days at 4°C. After rinsing in PBS and in a mixture of 0.1% cold water fish gelatin (Aurion, Wageningen, Netherlands) and 1% bovine serum albumin (BSA; Sigma-Aldrich, St. Louis, MO) in PBS, the sections were incubated in a cocktail of donkey anti-sheep IgG conjugated with 0.8 nm colloidal gold (1:100; Electron Microscopy Sciences, Fort Washington, PA) and biotinylated donkey anti-rabbit IgG (1:500; Jackson ImmunoResearch Labs) in the same cold water fish gelatin- and BSA-containing solution for 20 hours at 4°C. After washing in PBS, the sections were fixed in 1.25% glutaraldehyde (Electron Microscopy Sciences) in 0.1 M PB for 10 minutes. The gold particles were silver intensified with the Aurion R-Gent SE-LM Kit (Aurion, Wageningen, Netherlands) (14). After rinsing in 0.2 M sodium citrate, pH 7.5, the sections were immersed in ABC Elite Complex (1:1000; Vector Laboratories, Inc.) diluted in 0.05 M TRIS buffer for one hour at RT. The GlyT2 immunoreactivity was detected with NiDAB developer (0.025% DAB/0.0036% H2O2 in 0.05 M Tris buffer, pH 7.6). The sections were osmicated and then treated with 2% uranyl acetate in 70% ethanol for 30 minutes. Following dehydration in an ascending series of ethanol and acetonitrile, the sections were flat embedded in Durcupan ACM epoxy resin (Fluka; Sigma-Aldrich) on liquid release agent (Electron Microscopy Sciences)-coated slides, and polymerized at 56°C for two days. Ultrathin 50–60 nm sections were cut with a Leica Ultracut UCT ultramicrotome (Leica Microsystems, Wetzlar, Germany), collected onto Formvar-coated, single-slot grids, and examined with a JEOL electron microscope (JEOL, Tokyo, Japan).
Detection of the glycine receptor content of TRH neurons in the PVN
Sections of TRH-IRES-tdTomato mice (n = 5) were prepared for immunocytochemistry as described in the Supplementary Materials and Methods.
Series of sections from each brain were then subjected to pepsin pretreatment to facilitate antigen exposure (13). The sections were incubated in 1 mg/mL pepsin (Dako Agilent, Santa Clara, CA) for seven minutes at 37°C. After washing in PBS, the sections were treated with 0.5% Triton X-100/0.5% H2O2 in PBS for 15 minutes, immersed in 2% NHS in PBS for 20 minutes, and then incubated in a mixture of guinea pig-GlyRa IgG (12,13) and rabbit-RFP antiserum (1:3000; Rockland Immunochemicals, Inc., Limerick, PA) in antiserum diluent for two days at 4°C. After rinses in PBS, the sections were incubated in biotinylated donkey anti-guinea pig IgG (1:500; Jackson ImmunoResearch Labs) for two hours and in ABC Elite Complex (1:1000; Vector Laboratories, Inc.) in 0.05 M TRIS buffer for an hour. The immunoreaction was amplified with biotinylated tyramide using the TSA amplification kit (Perkin Elmer Life and Analytical Sciences, Waltham, MA). After further washes, the sections were incubated in a mixture of fluorescein DTAF-conjugated streptavidin (1:300; Jackson ImmunoResearch Labs) and Alexa 555-conjucated donkey anti-rabbit IgG (1:500; Jackson ImmunoResearch Labs), mounted onto glass slides, and coverslipped with the Vectashield mounting medium (Vector Laboratories, Inc.).
Retrograde tract tracing and tissue preparation for the identification of the sources of the glycinergic innervation of the PVN
The retrograde tracer, cholera toxin β subunit (CTB; List Biological Labs, Campbell, CA), was injected by iontophoresis into the PVN of GlyT2/GFP mice (n = 9). The animals were anesthetized ip with ketamine/xylazine (ketamine: 50 mg/kg; xylazine: 10 mg/kg body weight) and their head positioned in a stereotaxic apparatus with the Bregma and Lambda in the horizontal plane. Through a burr hole in the skull, a glass micropipette (17.5–20 μm outer tip diameter) filled with 0.5% CTB in 0.01 M PB at pH 8.0 was lowered into the brain at stereotaxic coordinates corresponding to the PVN (anterior–posterior, −0.9 mm from the Bregma; lateral, −0.15 mm; and dorsoventral, −4.8 mm from the surface of the skull), based on the atlas of Paxinos and Watson (15). CTB was deposited over three minutes of positive current (4 μA, pulsed on/off at seven-second intervals) using a constant-current source (Stoelting, Wood Dale, IL). The animals were anesthetized 7–10 days after tracer deposition and perfused with 10 mL PBS, pH 7.4, followed by 50 mL of 4% PFA in 0.1 M PB, pH 7.4. The brains were rapidly removed, postfixed in 4% PFA for two hours at RT, cryoprotected in 30% sucrose in PBS overnight at RT, then frozen using powdered dry ice, and stored at −20°C until use. Twenty-five-micrometer-thick coronal sections were cut on a freezing microtome into one-in-four series of sections.
Immunohistochemical identification of the CTB injection sites
Series of sections were pretreated as described above and then incubated in goat anti-CTB serum (1:10,000; List Biological Labs) for two days. Following washes in PBS, the sections were immersed in biotinylated donkey anti-sheep IgG (1:500; Jackson ImmunoResearch Labs) for two hours and in ABC (1:1000, ABC Elite; Vector Laboratories, Inc.) diluted in 0.05 M Tris buffer for one hour. Following rinses in PBS, peroxidase activity was visualized with NiDAB developer. The sections were mounted on gelatin-coated slides, air dried, counterstained with 1% cresyl-violet, and coverslipped with DPX mounting medium (Sigma-Aldrich).
Double-labeling immunofluorescence for identification of the sources of the glycinergic input of the PVN
One of the four series of sections was pretreated as described above and incubated in a mixture of goat anti-CTB serum (1:5000; List Biological Labs) and rabbit anti-GFP serum (1:10,000; Life technologies) for two days at 4° C. Then, the sections were immersed in biotinylated donkey anti-sheep IgG (1:500; Jackson ImmunoResearch Labs) for two hours, followed by ABC (1:1000, ABC Elite; Vector Laboratories, Inc.) for two hours. The immunoreaction product was amplified with the TSA amplification kit for 10 minutes. The signals were visualized by incubation in a mixture of Alexa 488-conjugated donkey anti-rabbit IgG (1:500; Jackson ImmunoResearch Labs) and Alexa 555-conjugated streptavidin (1:500; Jackson ImmunoResearch Labs) for two hours, mounted onto glass slides, and coverslipped with Vectashield mounting medium (Vector Laboratories, Inc.).
Virus injection and tissue preparation for identification of the sources of the glycinergic innervation of the TRH neurons in the PVN
As our retrograde tract-tracing experiment showed that the ventrolateral periaqueductal gray (VLPAG) and the raphe magnus (RMg) are the sources of the glycinergic input of the PVN, DIO-hChR2(H134R)-eYFP adeno-associated virus (AAV) was injected to the VLPAG (n = 5) or RMg (n = 5) in TRH-IRES-tdTomato//GlyT2-Cre mice. The Cre expression in the GLYT2 cells allowed the glycinergic neuron-specific expression of the hChR2(H134R)-eYFP fusion protein, while the tdTomato expression of TRH neurons facilitated the detection of TRH neurons. The virus injection and tissue preparation were performed as described in the Supplementary Materials and Methods.
Localization of virus injection sites
Series of sections containing the VLPAG or RMg were mounted onto glass slides and coverslipped with DAPI containing Vectashield mounting medium (Vector Laboratories, Inc.). The injection sites were detected based on the fluorescence of yellow fluorescent protein (YFP).
Tissue preparation for examination of the innervation of TRH neurons by glycinergic neurons of the VLPAG and RMg
Sections of the PVN pretreated as described above were incubated in a mixture of primary antisera: rabbit anti-GFP (1:10,000; Thermo Fisher, Waltham, MA) and sheep anti-tdTomato (1:80,000, generated in our laboratory) for 48 hours. After washing in PBS, sections were incubated in a cocktail of Alexa 488-conjucated donkey anti-rabbit IgG (1:250; Invitrogen, Carlsbad, CA) and Alexa 555-conjugated donkey anti-sheep IgG (1:500; Invitrogen) for two hours, mounted onto glass slides, and coverslipped with Vectashield mounting medium.
Image analysis of light microscopic preparations
The image analysis is described in the Supplementary Materials and Methods.
In vitro patch-clamp electrophysiology
Data acquisition and analysis
The slice preparation for electrophysiological studies and the used chemicals are described in the Supplementary Materials and Methods.
The slices were transferred to a submersion-type recording chamber containing arteficial cerebrospinal fluid (aCSF) at 32–33°C and were perfused with aCSF at a rate of ∼3 mL/min. TRH neurons were identified by the red fluorescence of the tdTomato under short epifluorescent illumination using an FN1 microscope (Nikon, Tokyo, Japan) equipped with 40 × water-immersion objective with additional zoom (up to 2 × ) and Zyla CCD camera (ANDOR). Afterward, the selected cell was studied under infrared differential interference contrast illumination. The patch pipettes (6–7 MΩ) were pulled from borosilicate capillaries (outer diameter = 1.5 mm thin wall; Garner Co, Maharashtra, India.) with a P-1000 horizontal puller (Sutter Instrument Co., Novato, CA). The intracellular pipette solution used for current-clamp (I = 0) electrophysiological recordings contained (in mM) 110 K-gluconate, 4 NaCl, 20 HEPES, 0.1 EGTA, 10 phosphocreatine di(tris) salt, 2 ATP, and 0.3 GTP (pH 7.25; 280–300 mOsm/L). For spontaneous inhibitory postsynaptic current (sIPSC) recording, the intracellular solution had the following composition: (in mM) 130 CsCl, 8 NaCl, 0.1 CaCl2, 0.1 EGTA, 10 HEPES, 4 Mg-ATP, 0.3 Na2-GTP, and 5 lidocaine N-ethyl bromide (pH 7.25; 280–300 mOsm/L). Recordings were performed with a MultiClamp 700B patch-clamp amplifier, Digidata-1440A data acquisition system, and pCLAMP 10.4 software (Molecular Devices). The headstage of the amplifier was fitted onto a Luigs & Neumann SM7 micromanipulator system. Whole-cell current-clamp recordings were filtered at 10 kHz using the built-in Bessel filter of the amplifier and digitized at 10 kHz. Slow and fast capacitive components were automatically compensated for. The stability of the patch was checked by repetitively monitoring the access resistance during the experiment, and TRH neurons in which the series resistance changed >25% were excluded from the statistics. Liquid junction potential was 14.4 mV and not compensated.
Whole-cell patch-clamp recording
After establishing a stable, whole-cell, patch-clamp recording, a control period was recorded for two to three minutes that was followed by a drug treatment phase for three to four minutes. First, the effect of glycine (0.5 mM) was measured on the firing frequency of TRH neurons in the presence or absence of the GlyR inhibitor, strychnine (125 μM), in current-clamp mode (IC). Then, the voltage-dependent sodium channel inhibitor, tetrodotoxin (TTX; 1.2 mM), was added to the aCSF to prevent the potential indirect effects of glycine treatment, and the effect of glycine was also examined in the presence or absence of strychnine. The washout of the drugs restored the spiking frequency or the membrane potential of TRH neurons in all cases.
For glycinergic sIPSC recordings, cells were voltage clamped using a whole-cell clamp configuration at a holding potential of −63 mV, the average resting membrane potential of the TRH neurons. A control value was recorded for two to three minutes in the presence of gabazine (3 μM) and kynurenic acid sodium salt (KYNA) (1 mM) to inhibit the effects of the GABAergic or glutamatergic inputs, followed by application of strychnine in the presence of gabazine and KYNA for three to four minutes.
Statistical analysis of the data of electrophysiological recordings
Changes of spiking frequency, membrane potential, and sIPSC recording were analyzed with Clampfit module of the pCLAMP 10.4 software (Molecular Devices, San José, CA) and OriginPro 2015. Statistical analysis was performed using one-way analysis of variance followed by the Bonferroni post hoc test to determine differences between treatment groups in the current-clamp experiments, and the paired t-test was carried out in the voltage-clamp experiment. The number of studied cells is described in the results of each experiment. All data are reported as mean ± standard error of mean. A p-value <0.05 was considered significant in all cases.
Results
Colocalization of tdTomato immunoreactivity and proTRH mRNA in the PVN of TRH-IRES-tdTomato mice
Drying results in fading of tdTomato's fluorescence, and therefore, its detection was performed by immunofluorescence in TRH-IRES-tdTomato mice. The distribution of tdTomato immunofluorescence (Fig. 1A) was found to be identical with the known distribution of TRH in the hypothalamus. In situ hybridization combined with immunofluorescence demonstrated a complete overlap between tdTomato-immunofluorescence and the in situ hybridization signal detecting the proTRH mRNA in the PVN. This demonstrates the selectivity and specificity of tdTomato expression in the generated TRH-IRES-tdTomato mice (Fig. 1B, C).
Characterization of the TRH-IRES-tdTomato mice. Low-magnification image (
TRH neurons receive GLYT2-IR innervation in the PVN
GlyT2-IR axons were observed in close proximity to TRH-IR neurons in all subdivisions of the PVN where TRH neurons are located. By double-labeling immunofluorescence, large GLYT2-IR varicosities were observed on the surface of TRH-IR neurons (Fig. 2). Quantification of the interaction of the two systems showed that GlyT2-IR axon varicosities established contacts with 53% ± 2% of TRH neurons. An average of 1.9 ± 0.1 GlyT2-IR contacts were found on the surface of the innervated TRH neurons. At the ultrastructural level, synaptic associations were observed between NiDAB-labeled, GlyT2-IR varicosities and TRH neurons labeled with silver-intensified colloidal gold particles (Fig. 3). Both axosomatic and axodendritic synapses were found between the two systems. All of the observed synaptic associations were of the symmetric type, indicating the inhibitory nature of these inputs.
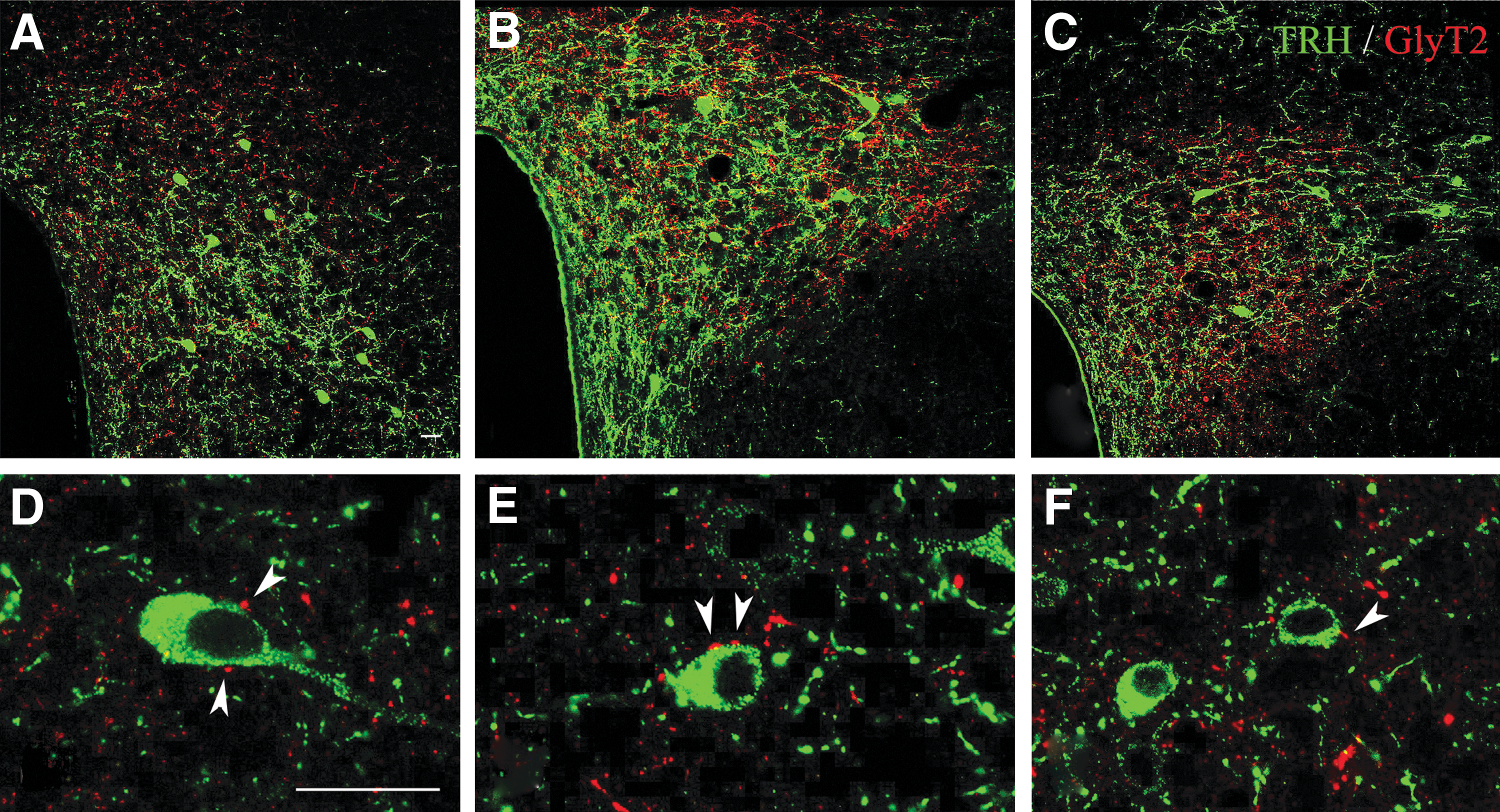
Relationship of GLYT2-IR axons and TRH-IR neurons in the PVN of CD1 mice. GlyT2-IR (red) axons were observed in close proximity of the TRH-IR (green) neurons in all subdivisions of the PVN. (
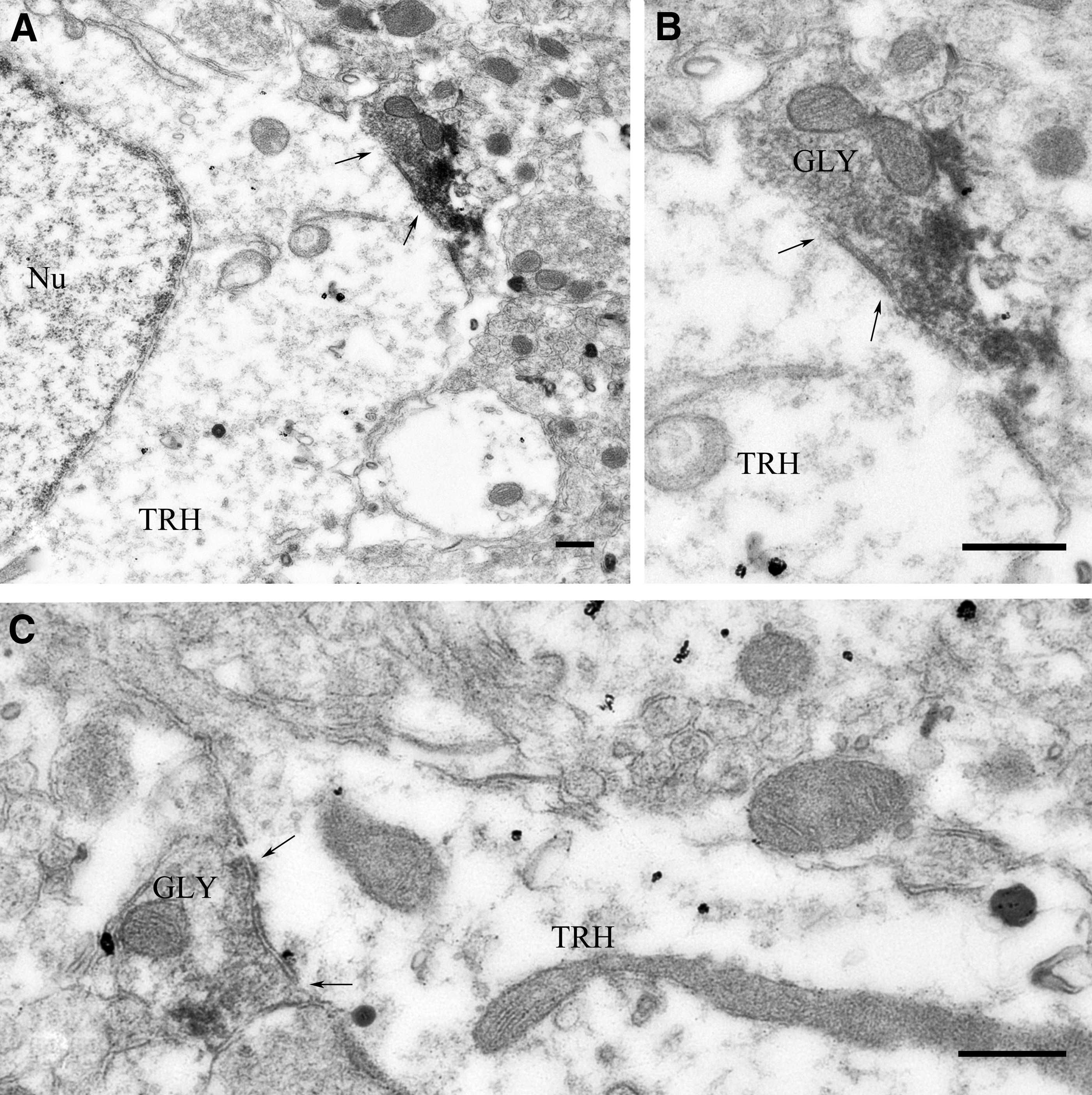
GLYT2-IR varicosities establish symmetric-type synapses on TRH neurons in the PVN of CD1 mice. At the ultrastructural level, synaptic associations were observed between NiDAB-labeled GlyT2-IR varicosities and TRH neurons labeled with silver-intensified colloidal gold particles. Both axosomatic (
TRH neurons express GlyRs in the PVN
To understand whether glycine released from axon terminals can influence TRH neurons in the PVN, the GlyR content of TRH neurons was studied in TRH-IRES-tdTomato mice. Punctuate GLYR-immunoreactivity was observed in all TRH neurons in all subdivisions of the PVN (Fig. 4).
Presence of glycine receptor immunoreactivity in the TRH neurons in the PVN of TRH-IRES-tdTomato mice. Punctuate glycine receptor-immunoreactivity (green) is present in all TRH neurons (red) studied. Scale bar = 5 μm.
The VLPAG and RMg are the sources of the glycinergic input of TRH neurons in the PVN
As a first step, the retrograde tracer CTB was injected into the PVN of GlyT2::GFP mice to investigate the origins of the glycinergic input of the PVN. CTB injection sites localized well within the borders of the PVN (Fig. 5) were used for the colocalization of CTB-immunoreactivity and GFP, the latter labeling glycinergic neurons. Double-labeled PVN projecting glycinergic neurons were observed in only two regions of the brain stem: in the RMg and VLPAG (Fig. 5).

Origin of the glycinergic input of the PVN of GLYT2::GFP mice (
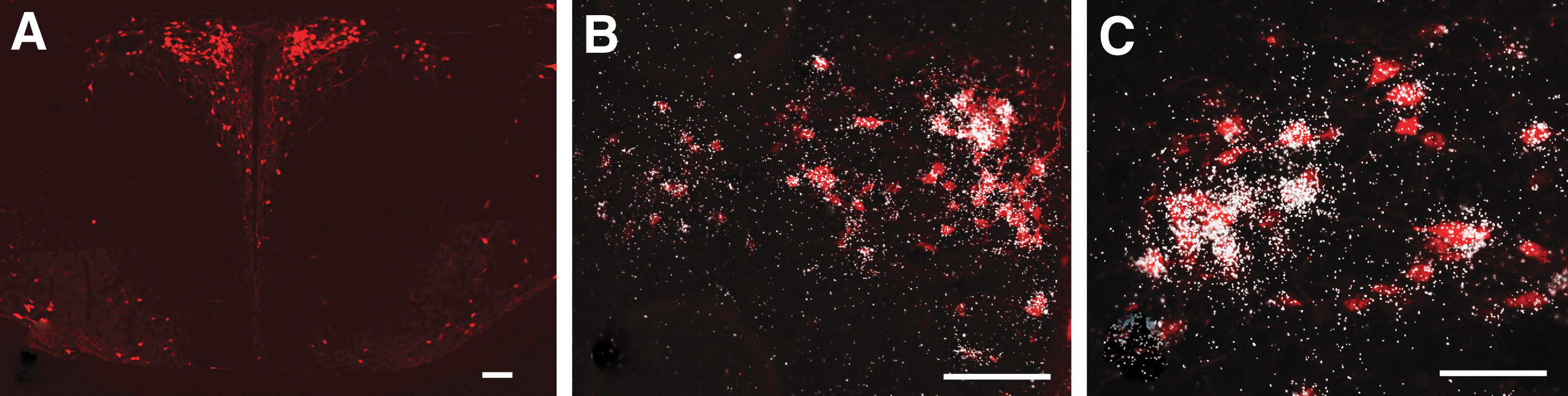
To determine whether the TRH neurons of the PVN receive glycinergic input from both of these brain stem nuclei, the hChR2(H134R)-eYFP fusion protein was expressed specifically in the glycinergic neurons of the RMg or the VLPAG of TRH-IRES-tdTomato//GlyT2::Cre mice using AAV-mediated gene transfer. The AAV injection site was centered within the RMg in four mice and in the VLPAG in six mice (Fig. 6A, J). Axons of glycinergic neurons of RMg origin innervated symmetrically both parts of the PVN. While these axons densely innervated all parvocellular subdivisions of the PVN (Fig. 6B–D), the magnocellular division of the PVN received less dense innervation. Glycinergic axons of VLPAG origin innervated primarily the ipsilateral side of the PVN, but sparse glycinergic axons were also observed on the contralateral side. Higher magnification images (Fig. 6E–I, N–Q) demonstrated that glycinergic axons originating from both the RMg and VLPAG contacted TRH neurons in all parvocellular subdivisions of the PVN, demonstrating that the glycinergic input of TRH neurons originates from both the RMg and the VLPAG.
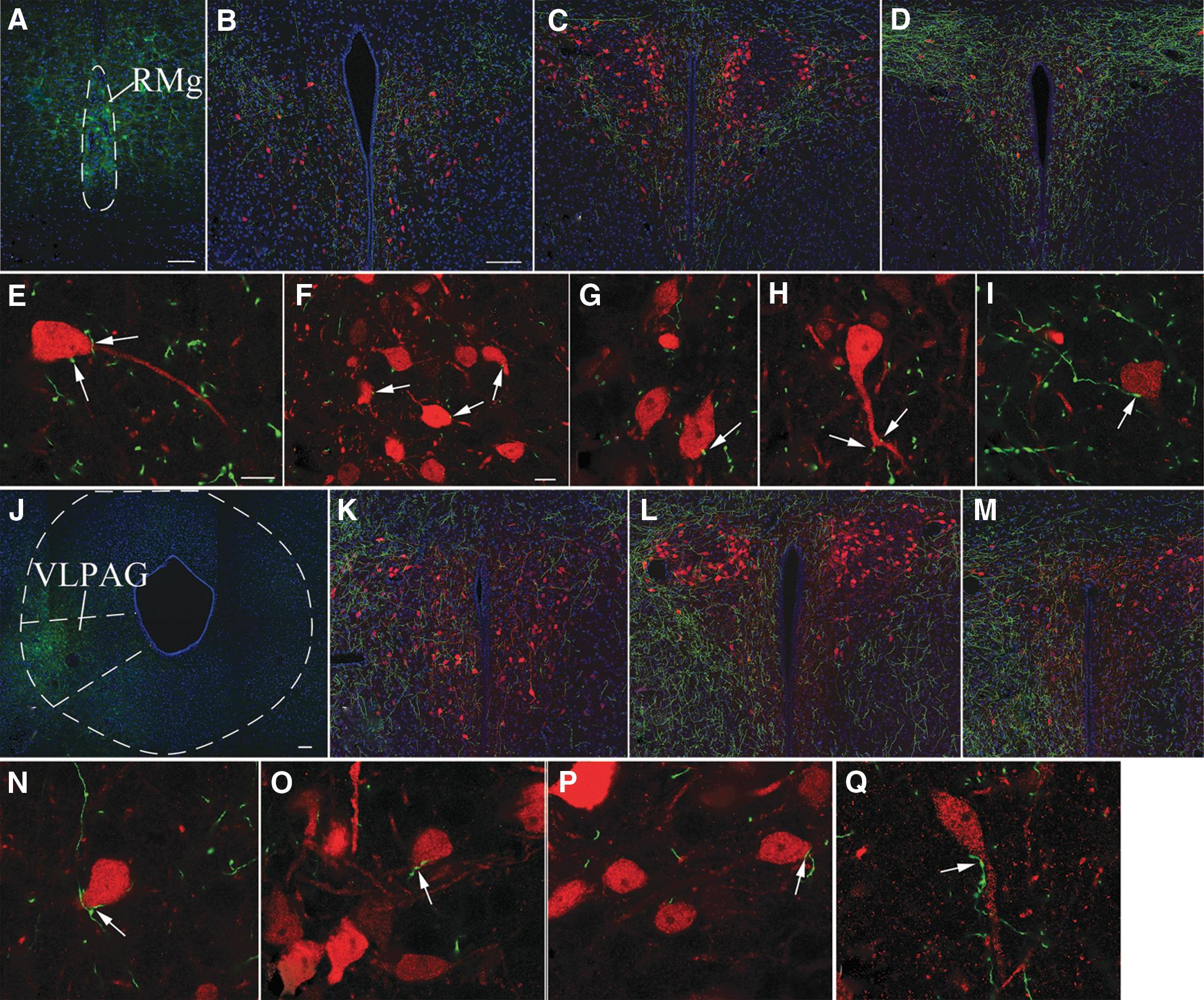
Involvement of the glycinergic neurons of the RMg and VLPAG in the innervation of the TRH neurons in the PVN of TRH-IRES-tdTomato//GlyT2::Cre mice. A representative image (
Glycine inhibits the firing of TRH neurons in the PVN
To understand the role of glycine in the regulation of TRH neurons, the effects of this amino acid transmitter were studied on the membrane potential and firing of TRH neurons in the midlevel of the PVN of TRH-IRES-tdTomato mice, where the hypophysiotropic TRH neurons are enriched (16). Application of glycine markedly decreased the membrane potential (−14.23 ± 2.55 mV; n = 6, p < 0.001; Fig. 7) and completely blocked the firing (control: 3.15 ± 0.52 Hz vs. glycine: 0.08 ± 0.08 Hz, n = 6, p < 0.001; Fig. 7) of TRH neurons. Coapplication of strychnine completely prevented the glycine-induced changes of the membrane potential (0.16 ± 1.65 mV, p = 1 vs. control and p < 0.001 vs. glycine, n = 6; Fig. 7) and the firing rate (3.54 ± 0.84 Hz, p = 1 vs. control and p < 0.001 vs. glycine, n = 6; Fig. 7) of TRH neurons.
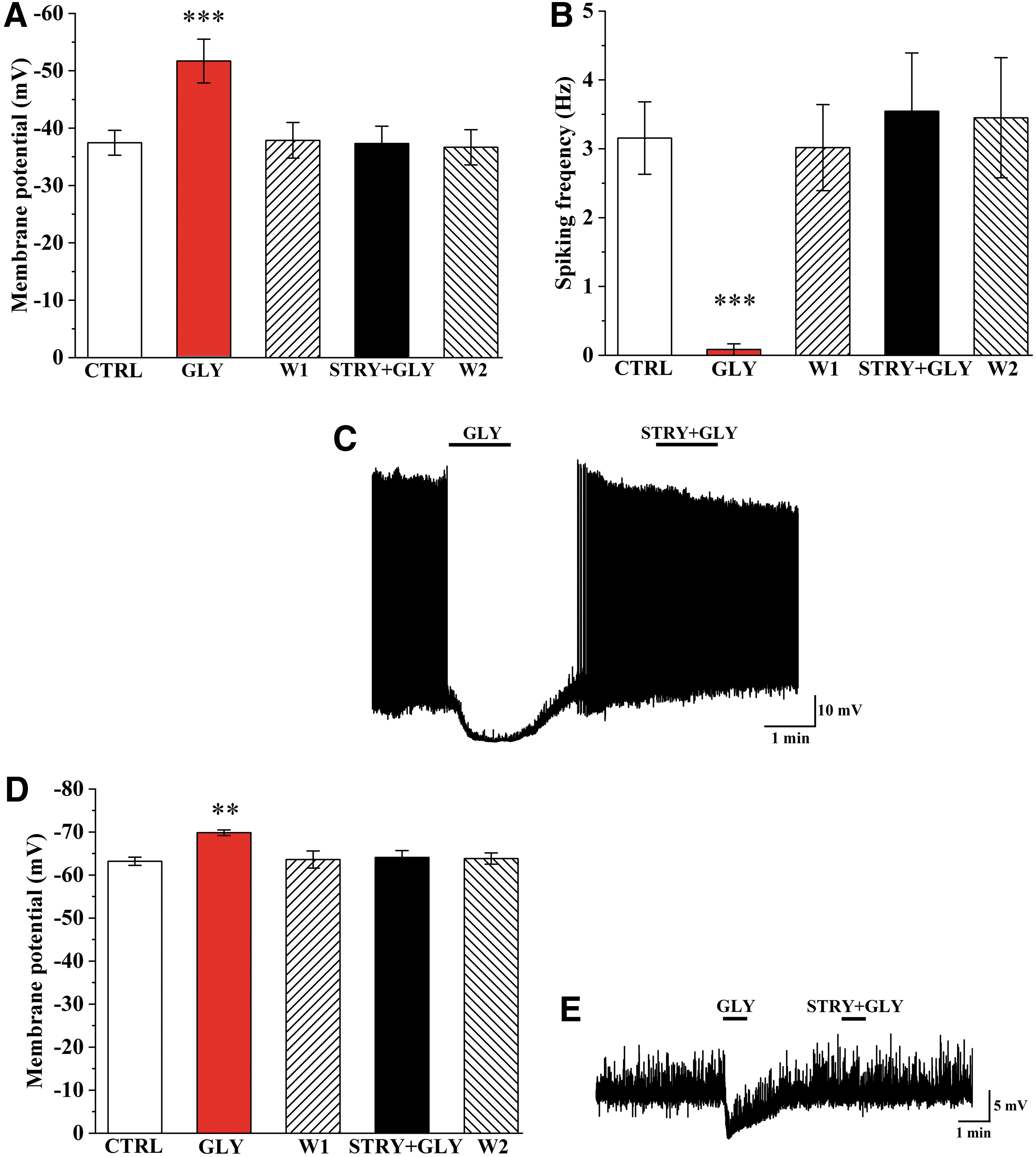
Effect of GLY on the membrane potential and the spiking frequency of TRH neurons. Bar graphs show changes of membrane potential (
To understand whether the inhibitory effect of glycine is exerted directly on the TRH neurons, the effect of glycine was also studied in the absence of neuronal inputs in TTX (1.2 mM)-treated slides. Application of glycine caused an ∼6 mV hyperpolarization of TRH neurons (control: −63.20 ± 0.94 mV vs. Gly: −69.84 ± 0.65 mV, n = 5, p = 0.0011; Fig. 7D, E) even in the presence of TTX. Application of strychnine abolished this effect of glycine (−64.11 ± 1.58 mV, n = 5; Fig. 7D, E).
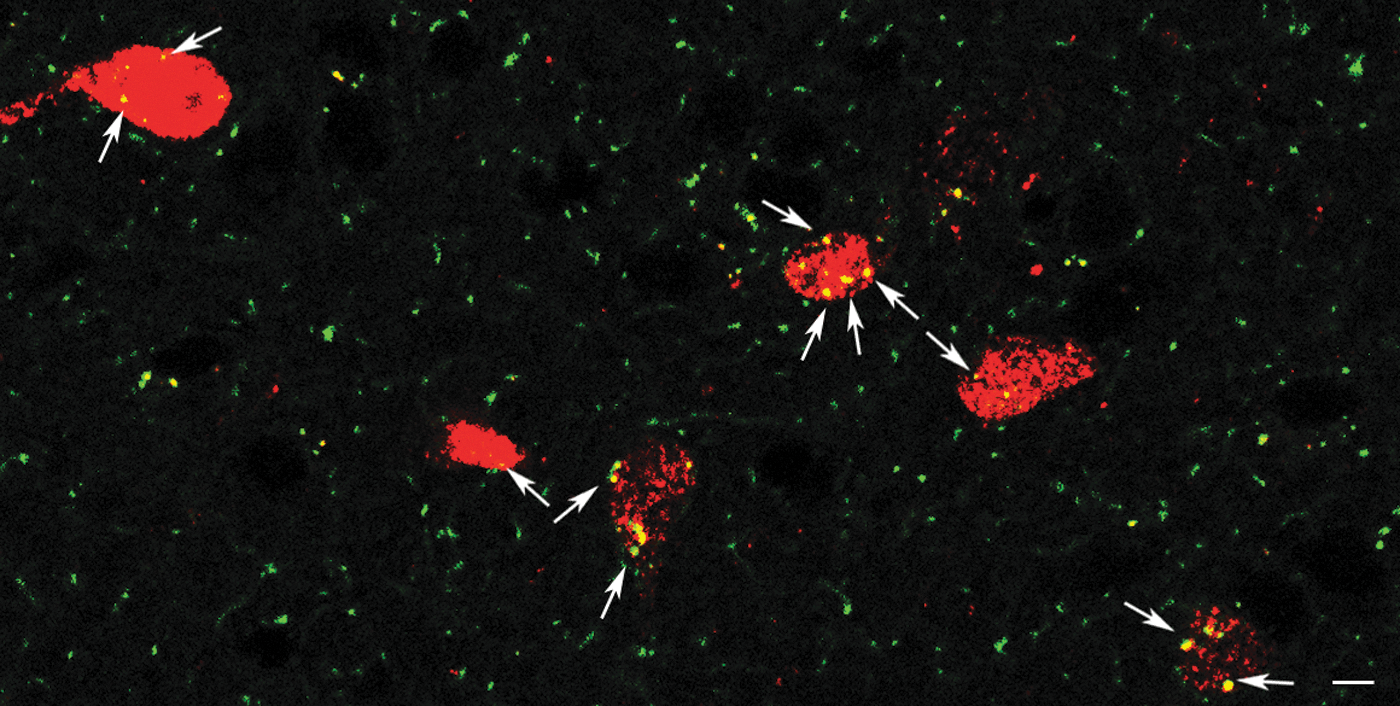
TRH neurons receive spontaneous inputs from glycinergic terminals
To demonstrate the involvement of the glycinergic inputs in the regulation of TRH neurons of the PVN, the spontaneous glycinergic currents of these cells were studied in TRH-IRES-tdTomato mice. Glycinergic sIPSCs were isolated by simultaneous inhibition of the glutamatergic and GABAergic inputs with a mixture of KYNA and gabazine. Despite the presence of these inhibitors, sIPSCs were observed in 62.5% of TRH neurons. The frequency of these sIPSCs was 0.824 ± 0.19 Hz (Fig. 8). Strychnine markedly decreased the frequency of these sIPSCs (0.12 ± 0.04 Hz, n = 5, p = 0.038; Fig. 8), suggesting that glycinergic synaptic inputs inhibit a population of TRH neurons. In 37.5% of the studied TRH neurons (n = 3), the frequency of sIPSCs was only 0.080 ± 0.014 Hz after inhibition of GABAergic and glutamatergic current cells with vs. without glycinergic sIPSC: p = 0.028). Strychnine had no effect on the sIPSC frequency of these cells (0.080 ± 0.045 Hz; p = 0.963).
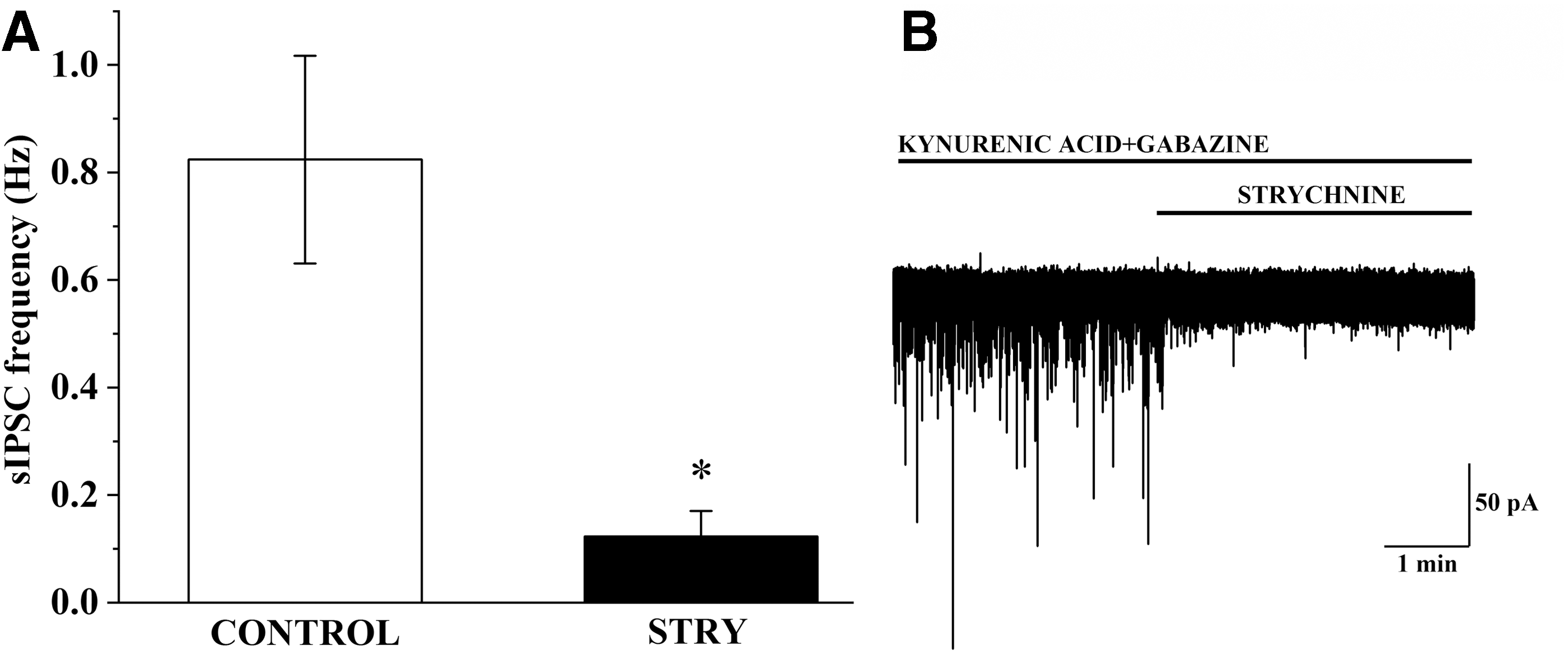
Examination of glycinergic sIPSCs on TRH neurons. Bar graph (
Discussion
An important function of the HPT axis is to maintain stable thyroid hormone levels in the circulation under normal conditions and thus to provide a continuous supply of thyroid hormones for tissues (8). This function is primarily controlled by the negative feedback effect of thyroid hormones on hypophysiotropic TRH neurons and pituitary thyrotrophs (8). Under certain conditions, such as fasting, infection, or cold exposure, however, the activity of the HPT axis is altered by neuronal inputs or by the activity of tanycytes (8). Peptidergic inputs originating from the arcuate nucleus are known to mediate the effects of fasting and leptin treatment on the hypophysiotropic TRH neurons (8), while adrenergic innervation stimulates the hypophysiotropic TRH neurons during cold exposure (17). Our laboratories described that the TRH neurons of the PVN also receive inputs from neurons utilizing classical neurotransmitters such as GABA and glutamate (18,19). Since glycinergic axons are also present in the PVN (3), we investigated whether TRH neurons are also controlled by glycine to regulate the thyroid axis.
To facilitate these studies, we generated a novel knockin mouse model expressing tdTomato specifically in TRH-producing cells using the CRISPR/Cas9 technology. By combined immunocytochemistry and in situ hybridization, we show that there is a complete overlap of tdTomato protein and the proTRH mRNA in the PVN of these mice, indicating that the TRH-IRES-tdTomato mouse line allows specific identification of the TRH neurons. This new mouse line also enabled us to study the electrophysiology of TRH neurons and identify the TRH neurons in morphological studies without the use of colchicine, an axonal transport inhibitor.
The data presented here demonstrate that glycinergic neurons innervate approximately half of the TRH neurons in the PVN, establishing symmetric-type synapsis, suggesting the inhibitory nature of these connections (20,21). In addition, we show that the vast majority of TRH neurons express GlyRs. In agreement with these morphological findings, exogenous glycine inhibits all TRH neurons studied in the PVN. Namely, glycine treatment markedly hyperpolarized TRH neurons and almost completely abolished the firing of these cells.
While approximately half of the TRH neurons received glycinergic innervation, almost all TRH neurons in the PVN contained GlyRs and responded to glycine treatment. This discrepancy raised the question whether the sensitivity of the immunocytochemical method was insufficient to detect all of the glycinergic input to TRH neurons, or whether the GlyRs of these neurons have other roles in addition to the detection of synaptically released glycine. To address this question, we studied the glycinergic sIPSCs of TRH neurons.
After inhibition of glutamatergic and GABAergic currents, sIPSCs were still observed in ∼60% of TRH neurons in the PVN. Administration of strychnine almost completely blocked the sIPSCs, demonstrating that endogenous glycine released from neuronal terminals exerts an inhibitory effect on TRH neurons in the PVN. However, glycinergic sIPSCs were not observed in ∼40% of TRH neurons in the PVN. These data indicate that extrasynaptic GlyRs are also involved in the regulation of TRH neurons.
In addition to glycine, taurine is also a known ligand of GlyRs (22). In the supraoptic nucleus (SON), Deleuze et al. (22) showed that the neurons express GlyRs, but do not receive functional glycinergic synapses. Furthermore, they observed that the GlyR expressing cells are contacted by taurine containing astrocytes (19,22). Based on these observations, they suggested that the ligand of GlyR in the SON is taurine, released from astrocytes (22). Thus, it seems feasible that TRH neurons in the PVN may also be regulated by both glycine released from neuronal inputs and by taurine secreted by astrocytes and will require further investigation.
Although glycinergic neurons are present in most parts of the brain stem, the origin of the glycinergic input to TRH neurons in the PVN is restricted to two brain stem nuclei, the VLPAG and RMg. The role of these nuclei in the regulation of the TRH neurons and the HPT axis is currently unknown. Published data, however, indicate that some of the conditions known to inhibit the HPT axis, such as the administration of bacterial lipopolysaccharide (LPS), a model of infection or different stressors (8), induce neuronal activation in the VLPAG and RMg (10,22 –27). Thus, further studies are needed to determine whether projections from glycinergic neurons in these two brain stem regions to TRH neurons in the PVN have a role in the mediation of stress and LPS-induced inhibition of the hypophysiotropic TRH neurons and the HPT axis.
Glycinergic inputs of TRH neurons are found not only in the midlevel of the PVN, where the hypophysiotropic TRH neurons are concentrated (16), but also in other regions of the PVN, such as in the anterior parvocellular subdivision, where the TRH neurons have no hypophysiotropic function (16). Therefore, our data indicate that glycine regulates both hypophysiotropic and nonhypophysiotropic populations of the TRH neurons in the PVN. Currently, very little information is available about the function of nonhypophysiotropic TRH neurons of the PVN. The TRH neurons residing in the anterior parvocellular subdivision of the PVN were, however, shown to project to energy homeostasis-related areas, such as the arcuate and ventromedial nuclei (23), and thus, it is feasible that the glycinergic neurons regulate the energy homeostasis via the nonhypophysiotropic TRH neurons.
In summary, these data demonstrate that TRH neurons in the PVN receive a functional, glycinergic input from the VLPAG and RMg that exert an inhibitory effect on the TRH neurons, indicating that the glycinergic system may have an important role in the central regulation of the HPT axis. However, as not all TRH neurons in the PVN are innervated by glycinergic inputs yet responsive to glycine in vitro, we hypothesize that glial release of glycine or taurine, or yet another GlyR agonist, may also influence TRH neurons via glial/neuronal interactions.
Footnotes
Acknowledgments
The authors are very grateful to Dr. Hanns Ulrich Zeilhofer (University of Zurich, Zurich, Switzerland) for the kind gift of GLYT2::GFP and GLYT2::Cre mice.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by Grants from the Hungarian Science Foundation (OTKA K124767), the Hungarian National Brain Research Program (2017-1.2.1-NKP-2017-00002) and EU H2020 THYRAGE no. 666869.
Supplementary Material
Supplementary Data