
Abstract
Background:
Several clinical and experimental studies have implicated thyroid hormones in cancer progression. Cancer-relevant effects, including stimulation of tumor growth and new blood vessel formation by angiogenesis, are thought to be mediated by a nonclassical signaling pathway initiated through integrin αvβ3 expressed on cancer cells and proliferating endothelium. In an earlier study, we established mesenchymal stem cells (MSCs), important contributors to the fibrovascular network of tumors, as new thyroid hormone-dependent targets. Here, we evaluated the effects of the thyroid hormones triiodothyronine (T3) and thyroxine (T4) versus Tetrac, an integrin-specific inhibitor of thyroid hormone action, on MSCs in tumor angiogenesis.
Methods:
Modulation of the expression and secretion of angiogenesis-relevant factors by thyroid hormones in primary human MSCs and their effect on endothelial cell tube formation were tested in vitro. We further engineered MSCs to express the sodium iodide symporter (NIS) reporter gene under control of a hypoxia-responsive promoter and the vascular endothelial growth factor (VEGF) promoter to test effects on these pathways in vitro and, for VEGF, in vivo in an orthotopic hepatocellular carcinoma (HCC) xenograft mouse model by positron emission tomography imaging.
Results:
T3 and T4 increased the expression of pro-angiogenic genes in MSCs and NIS-mediated radioiodide uptake in both NIS reporter MSC lines in the presence of HCC cell-conditioned medium. Supernatant from thyroid hormone-treated MSCs significantly enhanced endothelial cell tube formation. Tetrac and/or inhibitors of signaling pathways downstream of the integrin reversed all these effects. Tumoral radioiodide uptake in vivo demonstrated successful recruitment of MSCs to tumors and VEGF promoter-driven NIS expression. Hyperthyroid mice showed an increased radioiodide uptake compared with euthyroid mice, while tracer uptake was markedly reduced in hypothyroid and Tetrac-treated mice.
Conclusions:
Our data suggest that thyroid hormones influence angiogenic signaling in MSCs via integrin αvβ3 and further substantiate the anti-angiogenic activity of Tetrac in the tumor microenvironment.
Introduction
Angiogenesis, the formation of new blood vessels from the pre-existing vasculature, is a fundamental process during embryonic development, wound healing, and tissue repair. Physiologically, angiogenesis is a highly regulated process controlled by the balance between pro- and anti-angiogenic regulatory molecules (1,2). The angiogenic switch occurs when this equilibrium is changed, leading to endothelial cell migration and proliferation from existing vessels, vascular sprouting, and three-dimensional vessel formation, followed by maturation and stabilization of newly formed vessels through their coverage by pericytes and smooth muscle cells (1).
Although the endothelium remains mostly quiescent in adulthood, endothelial cells retain their ability to rapidly proliferate in response to specific stimuli such as hypoxia or inflammation—stimuli that are typically found in the tumor microenvironment (3). As a tumor grows, so does its demand for oxygen and nutrients, requiring an expansion of the vascular network to meet this need. Hypoxia, a common feature of solid tumors, leads to increased activity of hypoxia-inducible transcription factors (HIFs) that in turn initiate the expression of target genes regulating angiogenesis (4). Thus, pathologically activated angiogenesis contributes to cancer progression.
Numerous angiogenic factors have been discovered in the secretome of mesenchymal stem cells (MSCs). First described by Friedenstein et al. in the 1960s, these multipotent nonhematopoietic stromal-like cells migrate to sites of tissue injury and inflammation, where they contribute to recovery of tissue integrity (5 –7). Governed by highly similar migratory signals, MSCs also home to the growing tumor stroma, where they differentiate into stroma-associated cells, including cancer-associated fibroblast- and pericyte-like cells (8 –12).
In the context of angiogenesis, MSCs influence endothelial cell proliferation, migration, and tube formation through their secretion of pro-angiogenic chemokines and growth factors. They are further involved in blood vessel maturation in their function as pericytes that help stabilize newly formed vessels (2). Furthermore, the secretion of angiogenic factors by MSCs was reported to be upregulated under hypoxic conditions (2).
In an earlier study, we observed that the thyroid hormones triiodothyronine (T3) and thyroxine (T4) stimulate the differentiation of MSCs toward a cancer-associated fibroblast-/pericyte-like phenotype (9). This included their upregulation of the pro-angiogenic endothelial growth factor genes vascular endothelial growth factor (VEGF) and basic fibroblast growth factor 2 (FGF2), in addition to the pericyte markers α-smooth muscle actin and desmin, as well as less specific factors that have been reported to be pro-angiogenic, such as hepatocyte growth factor, epidermal growth factor (EGF), transforming growth factor β1, and interleukin 6 (9). The upregulation of these factors by thyroid hormones in MSCs was shown to be initiated at the cell surface through integrin αvβ3 (9).
Discovered as a thyroid hormone receptor (TR) in 2005, integrin αvβ3 is present in the plasma membrane of proliferating endothelial cells and cancer cells, and also MSCs (9,13 –16). The role of integrin αvβ3 in cancer-related angiogenesis has been known since the 1990s (17). Subsequently, over the past two decades, iodothyronines have emerged as nonclassical pro-angiogenic modulators mediated through αvβ3 and have been shown to induce neovascularization in a variety of settings, such as the heart, ischemic striated muscle, and tumor beds (18).
The complex pro-angiogenic actions of T4 and T3 include the stimulation of endothelial cell proliferation, migration, and vascular tube formation. This involves the transcription and/or secretion of, among others, FGF2, VEGF, and HIF-1α, in addition to the modification of vascular growth factor receptor function via cross talk between the integrin and co-clustered growth factor receptors (16,18 –24).
The αvβ3 integrin contains two distinct binding sites for thyroid hormones that are localized near the Arg-Gly-Asp (RGD) recognition site that is essential for extracellular matrix binding (25). Upon binding of T3 to the S1 site, the PI3K/Akt pathway is activated, resulting in downstream effects that may include the transcription of HIF-1α (1,13,26). The second site, S2, binds both T4 and T3 and the hormone signal is transduced via the ERK1/2 (MAPK) pathway that leads to enhanced cell proliferation and the transcription of pro-angiogenic modulators, including FGF2 and VEGF (1,27).
Of note is that in contrast to classical thyroid hormone signaling via nuclear TRs, T4 is not merely a pro-hormone for T3, but actually functions as an active hormone at the integrin. 3,3′,5,5′-Tetraiodothyroacetic acid (Tetrac), a deaminated T4 derivative, specifically inhibits thyroid hormone action at both binding sites on the integrin (1,13,26).
Based on the observation that MSCs, in their role as progenitor cells of the tumor's fibrovascular network, are sensitive to thyroid hormones and the accumulating evidence for the pro-angiogenic action of thyroid hormones, the aim of the current study was to evaluate the potential effects of T3 and T4 versus Tetrac on MSCs in the context of tumor angiogenesis.
Materials and Methods
Cell culture
Primary human CD34-negative MSCs extracted from bone marrow (apceth GmbH, Munich, Germany) were cultured until passage 8 in Dulbecco's modified Eagle's medium (DMEM; Sigma–Aldrich, St. Louis, MO) containing 10% (v/v) fetal bovine serum (FBS) (FBS Superior; Biochrom/Merck Millipore, Berlin, Germany) and 100 U/mL penicillin/100 μg/mL streptomycin (Sigma–Aldrich). For reporter gene transfection, the SV40-immortalized human bone marrow-derived MSC line L87 was used (28,29). These cells are easily engineered and expandable, show similar differentiation capacity, as well as homing and activation characteristics as primary MSCs (30,31). L87 was cultured in RPMI (Sigma–Aldrich) supplemented with 10% (v/v) FBS and 100 U/mL penicillin/100 μg/mL streptomycin.
Primary human umbilical vein endothelial cells (HUVECs; provitro AG, Berlin, Germany) were used until passage 8 and cultured in endothelial cell proliferation medium (provitro AG).
The human hepatocellular carcinoma (HCC) cell line HuH7 (JCRB Cell Bank, Osaka, Japan) was cultured in DMEM containing 10% (v/v) FBS and 100 U/mL penicillin/100 μg/mL streptomycin and passaged up to 10 times.
All cell lines were maintained at 37°C in a humidified 5% (v/v) CO2 atmosphere.
Stable transfection of MSCs
For reporter gene assays, L87 MSCs transfected with the human sodium iodide symporter (NIS) gene under control of either a promoter containing hypoxia response elements [HRE-NIS-MSCs; established in Müller et al. (31)] or the VEGF promoter (VEGF-NIS-MSCs) were used. VEGF-NIS-MSCs were established by stable transfection of wild-type L87 MSCs with the expression vector
Cells were transfected with 2 μg
Hormone treatment in vitro
The preparation of and treatment with HuH7-conditioned medium and T3 or T4 with or without Tetrac was performed as described previously (9). For in vitro experiments, both T3 and T4 were used at physiological and supraphysiological concentrations, that is, 1 or 10 nM T3 and 100 or 1000 nM T4, respectively. The reference ranges for total hormone concentrations in healthy euthyroid humans are 64–154 nM for T4 and 1.1–2.9 nM for T3 (34). Tetrac was applied at an end concentration of 100 nM. As FBS, per se, contains thyroid hormones, charcoal-stripped serum was used for stimulation experiments (T3 concentration <0.2 pg/mL; T4 concentration 0.2–0.6 ng/dL).
Quantitative real-time polymerase chain reaction
RNA extraction, reverse transcription, and quantitative real-time polymerase chain reaction (qPCR) were performed as described previously (9). Primer sequences are listed in Table 1. Relative expression levels were calculated from ΔΔCt values normalized to the geometric means of internal β-actin (ACTB) and 18S rRNA (R18S). Samples were run in duplicate for each of four independent experiments.
Quantitative Real-Time Polymerase Chain Reaction Primers
Enzyme-linked immunosorbent assay
Supernatant from MSCs after 24 and 48 hours of stimulation was centrifuged and stored at −80°C until assayed for angiogenin (ANG), angiopoietin 1 (ANGPT1), angiopoietin 2 (ANGPT2), interleukin 8 (CXCL8), insulin-like growth factor 1 (IGF1), and placental growth factor (PGF) using the respective DuoSet ELISA (enzyme-linked immunosorbent assay) kit (R&D Systems, Abingdon, United Kingdom). Where necessary, supernatants were diluted as follows to be within the linear range of the assay: ANG 24 hours 1:20, 48 hours 1:80; ANGPT1 48 hours 1:4; CXCL8 24 hours 1:20, 48 hours 1:60; IGF1 24 and 48 hours 1:10; PGF 24 and 48 hours 1:2. Samples were analyzed in duplicate in four independent experiments.
Radioiodide uptake in vitro
NIS-mediated uptake of 125I (PerkinElmer, Waltham, MA) by HRE-NIS-MSCs, VEGF-NIS-MSCs, and wild-type MSCs was measured according to Spitzweg and colleagues (35). To test inducibility of the hypoxia-responsive and the VEGF promoter, cells were stimulated with 300 μM of the hypoxia-simulating agent cobalt chloride (CoCl2; Sigma–Aldrich) or with 20% FBS, respectively, for 24 hours.
Thyroid hormone treatment was performed as described above. HRE-NIS-MSCs were additionally stimulated with 300 μM CoCl2 during thyroid hormone treatment. The signaling pathway inhibitors LY 294002 (InSolution LY 294002; Calbiochem/Merck Millipore, Darmstadt, Germany) and RAF265 (Novartis, Basel, Switzerland), both stored as 10 mM stock solutions at −20°C in DMSO, were used at 10 and 1 μM end concentrations, respectively, and were added to cells simultaneously with thyroid hormones.
For each iodide uptake assay, two to four technical replicates were performed in three independent experiments each. Radioiodide uptake was normalized to cell viability determined by MTS assay (Promega, Mannheim, Germany) as described previously (36) using the absorbance at 620 nm relative to control cells.
Tube formation assay
HUVECs were seeded at a density of 7.5 × 103 cells/well in a 96-well μ-Plate Angiogenesis (ibidi, Martinsried, Germany) on growth factor-reduced Matrigel (Becton Dickinson Biosciences, Heidelberg, Germany). Five to six wells were plated for each condition in each of three or four independent experiments. Plates were incubated for 12 hours at 37°C before photomicroscopy. Images were acquired on a Leica DM IL microscope (Leica Microsystems, Wetzlar, Germany) equipped with a Jenoptik ProgRes CCD camera (Jenoptik, Jena, Germany) at 5 × magnification. Tube formation was quantified in an automated manner with the ImageJ (National Institutes of Health, Bethesda, MD) plug-in Angiogenesis Analyzer (37). Evaluated parameters were the mean number of meshes, junctions, and the mean total tube length per well.
Animals
All experiments were conducted with approval from the regional governmental commission for animals (Regierung von Oberbayern, Munich, Germany). Four- to five-week-old male CD1 nu/nu mice (Charles River, Sulzfeld, Germany) were allowed to acclimatize for one week under specific pathogen-free conditions with access to standard nude mouse diet (ssniff, Soest, Germany) and water ad libitum. HuH7 tumor cells (1 × 106) were implanted into the livers of mice after laparotomy as described previously (31).
Hormone treatment in vivo
Starting from one week after tumor cell injection, mice received drinking water supplemented with 0.02% (w/v) 2-mercapto-1-methylimidazole (MMI; Sigma–Aldrich) and 1% (w/v) sodium perchlorate (Sigma–Aldrich) to induce hypothyroidism and generate the same baseline thyroid hormone levels for all groups (9,38). 0.3% (w/v) Saccharin (Sigma–Aldrich) was added as a sweetener to mask the bitter taste of MMI/perchlorate.
Two weeks later, mice were randomly assigned to different treatment groups and received the following intraperitoneal (i.p.) injections daily: the hyperthyroid group (n = 6) received 100 ng/g body weight T4 (Sigma–Aldrich), the euthyroid group 20 ng/g body weight T4 with (n = 7) or without (n = 6) 10 μg/g body weight Tetrac (Sigma–Aldrich), while hypothyroid mice (n = 8) received saline only.
Three days before imaging (see “124I positron emission tomography” section), perchlorate was removed from the drinking water and only MMI treatment continued to prevent interference with radioiodide uptake. Thyroid hormone status was regularly monitored in serum samples by ELISA (DRG Diagnostics, Marburg, Germany).
124I positron emission tomography
Three weeks later, that is, six weeks after tumor cell inoculation, 5 × 105 VEGF-NIS-MSCs or wild-type MSCs (n = 2), were injected systemically in 500 μL phosphate-buffered saline via the tail vein. Seventy-two hours later, 10 MBq 124I (PerkinElmer or DSD Pharma, Purkersdorf, Austria) was applied i.p. and radioiodide distribution assessed using an Inveon P120 microPET (Siemens Healthcare, Erlangen, Germany). In three mice, the competitive NIS inhibitor sodium perchlorate (2 mg/mouse; Sigma–Aldrich) was injected i.p. 30 minutes before radioiodide administration to verify that tumoral radioiodide accumulation was NIS-mediated.
Positron emission tomography (PET) images were reconstructed with the software Inveon Acquisition Workplace (Siemens) and analyzed with Inveon Research Workplace (Siemens). Three-dimensional regions of interest were defined and quantified as mean percent of the injected dose per milliliter of tumor (% ID/mL tumor).
After PET imaging, mice were sacrificed and tumors embedded in Tissue-Tek O.C.T. compound (Sakura Finetek, Alphen aan den Rijn, The Netherlands), snap-frozen on dry ice, and stored at −80°C until further processing (see “Ex vivo immunofluorescence staining” and “Ex vivo analysis of NIS mRNA expression” sections).
Ex vivo immunofluorescence staining
Immunofluorescence staining for human NIS and mouse CD31 on frozen tissue sections was performed as described previously (31,36) using the mouse monoclonal anti-NIS antibody clone FP5A (Merck Millipore, Berlin, Germany) or the rat monoclonal anti-mouse CD31 antibody clone MEC 13.3 and a Cy3-conjugated donkey anti-mouse secondary antibody (Jackson ImmunoResearch, Cambridgeshire, United Kingdom) or a Cy3-conjugated donkey anti-rat secondary antibody (Jackson ImmunoResearch), respectively. Nuclei were counterstained with 5 μg/mL bisbenzimide Hoechst 33258 (Sigma–Aldrich).
Stained sections were mounted with Dako Fluorescence Mounting Medium (Dako, Carpinteria, CA) and imaged at 20 × (NIS) or 10 × (CD31) magnification on a Leica DMI6000 B fluorescence microscope equipped with a Leica DFC365 FX CCD camera and the software Leica Application Suite X (Leica Microsystems). A minimum of five visual fields was recorded per tumor for three to four mice per treatment group. The number of NIS-positive cells was counted manually, while the CD31-positive area per visual field was quantified using ImageJ, blinded to group assignment.
Ex vivo analysis of NIS mRNA expression
To normalize NIS expression to MSC recruitment, total RNA was isolated from tumors (euthyroid n = 5; hyperthyroid n = 6; hypothyroid n = 5; euthyroid+Tetrac n = 6) and qPCR was performed as described above using primers specific for NIS, SV40, which was used for MSC immortalization, and the blasticidin resistance gene (bsd), which is encoded by the NIS expression plasmid and constitutively expressed (for primer sequences, see Table 1). β-Actin and 18S rRNA served as internal controls. Results were calculated from ΔΔCt values for NIS divided by the mean of the respective ΔΔCt values for SV40 and bsd as a measure of the amount of MSCs present.
Statistics
All data are expressed as mean ± SEM (standard error of the mean) or mean fold change ± SEM. Statistical significance was tested by one-way analysis of variance followed by Tukey's post hoc test. p-Values <0.05 were considered statistically significant (*p < 0.05; **p < 0.01; ***p < 0.001).
Results
Thyroid hormones increase the expression of angiogenesis modulators in MSCs in vitro
Initially, we examined the expression of angiogenesis relevant factors in MSCs. To this end, we treated primary human bone marrow-derived MSCs with physiological or supraphysiological concentrations of T3 or T4 and HCC cell-conditioned medium to simulate the tumor environment in vitro. mRNA (Fig. 1A) and protein concentrations (Fig. 1B, C) for genes associated with angiogenesis were analyzed by qPCR or ELISA, respectively.
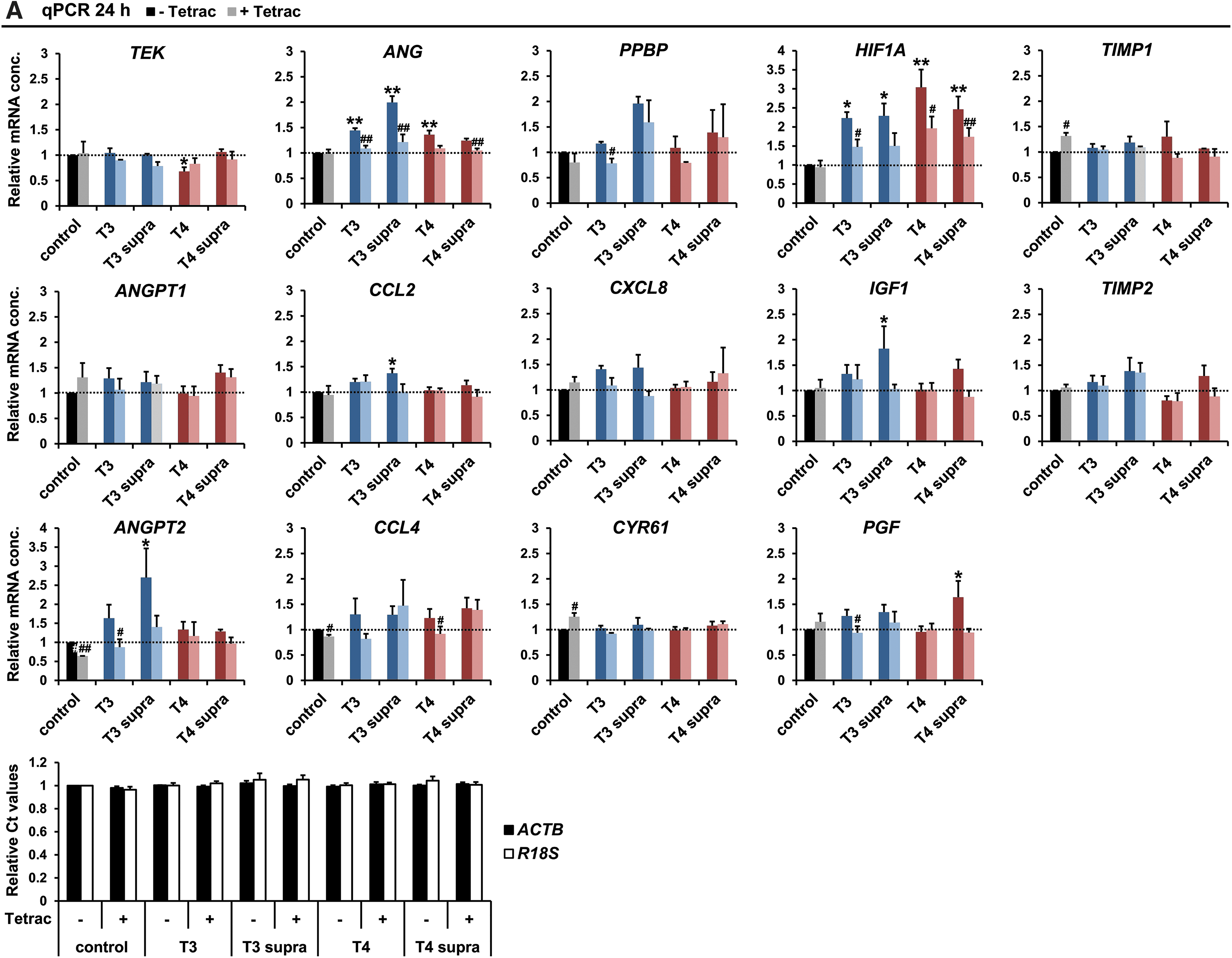
Expression of pro-angiogenic genes. Primary human bone marrow-derived MSCs were treated with hepatocellular carcinoma cell-conditioned medium and either physiological (1 nM, “T3”) and supraphysiological (10 nM, “T3 supra”) concentrations of T3 or physiological (100 nM “T4”) and supraphysiological (1000 nM “T4 supra”) concentrations of T4 with and without Tetrac (100 nM). (
For genes of the angiopoietin/TEK tyrosine kinase (TIE2) axis that control vessel maturation, expression levels of the receptor itself were reduced under stimulation with 100 nM T4, while no significant changes for its ligand ANGPT1 were detected at the mRNA level. ANGPT1 contributes to the maintenance of the resting endothelial state. In contrast, mRNA concentrations of ANGPT2, the naturally occurring antagonist of ANGPT1 that enhances vascular permeability and destabilizes the endothelium, were dose dependently increased upon T3 stimulation compared with cells treated with tumor cell-conditioned medium alone, an effect that was significant at 10 nM T3 and was blocked by Tetrac.
Pro-angiogenic ANG and the growth factor IGF1 showed a similar expression pattern. HIF-1A mRNA expression was significantly upregulated by both physiological and supraphysiological concentrations of both T3 and T4, and T4 effects were reduced by Tetrac. The chemokines CCL4, pro-platelet basic protein (PPBP/CXCL7), and CXCL8 showed only slight nonsignificant upregulation under T3 stimulation that was decreased by Tetrac, while CCL2 expression was significantly increased under stimulation with 10 nM T3. No T4 effects were observed for these chemokines. PGF showed only weak nonsignificant stimulation under T3 treatment but was significantly increased under stimulation with 1000 nM T4 at the mRNA level.
The extracellular matrix-associated angiogenic modulator cysteine-rich 61 (CYR61), as well as tissue inhibitors of metalloproteinases 1 and 2 (TIMP1/2), showed no significant changes in expression levels under thyroid hormone stimulation. CYR61 and TIMP1 mRNAs were upregulated when cells were treated with HCC cell-conditioned medium and Tetrac compared with cells treated with HCC cell-conditioned medium alone (Fig. 1A). The reference genes ACTB and R18S used for normalization were not impacted by the different treatment conditions (Fig. 1A, bottom panel).
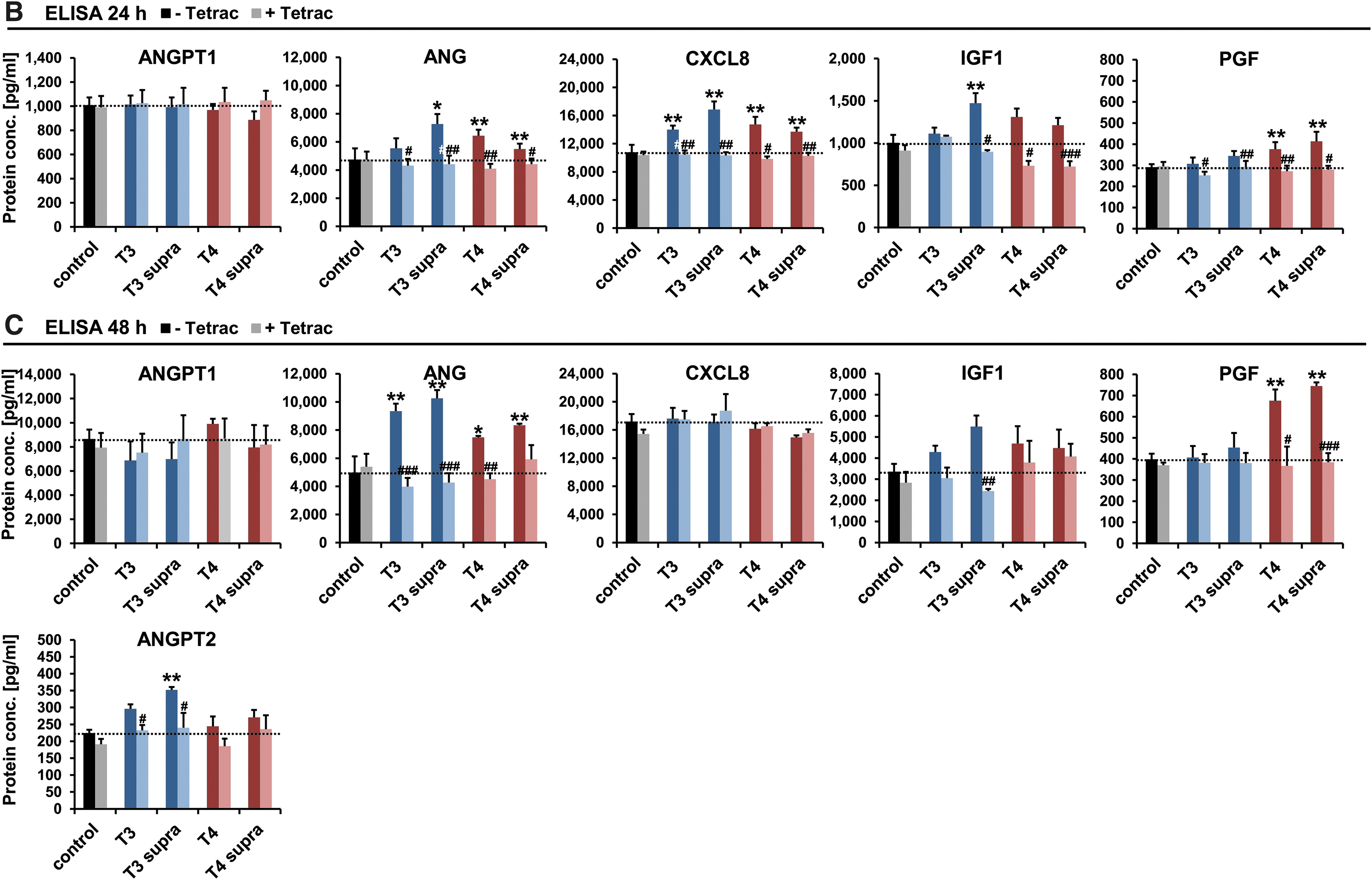
For a subset of these genes, the concentrations of secreted proteins in MSC supernatants after 24 hours (Fig. 1B) and 48 hours (Fig. 1C) of stimulation were validated by ELISA. Effects seen at the mRNA level were confirmed at the protein level for ANGPT1, ANG, and IGF1 by ELISA (Fig. 1B, C). ANGPT2 expression was increased significantly and dose dependently under T3 stimulation in a Tetrac-dependent manner after 48 hours, reflecting the effects seen at the mRNA level (Fig. 1C). At 24 hours, ANGPT2 levels were below the detection limit of the ELISA for all treatment conditions. CXCL8 showed a significant increase in secreted protein at 24 hours that was completely absent after 48 hours. PGF expression, by contrast, was induced by T4 after 24 hours, an effect that was even stronger after 48 hours. This effect was blocked by additional treatment with Tetrac.
Supernatant from thyroid hormone-treated MSCs stimulates endothelial cell tube formation
To examine the paracrine effects of angiogenic factors secreted by MSCs on endothelial cells, HUVECs were seeded on growth factor-reduced Matrigel and subjected to conditioned medium from stimulated MSCs. Tube formation was analyzed and quantified in an automated manner on microscopy images after 12 hours (Fig. 2A).
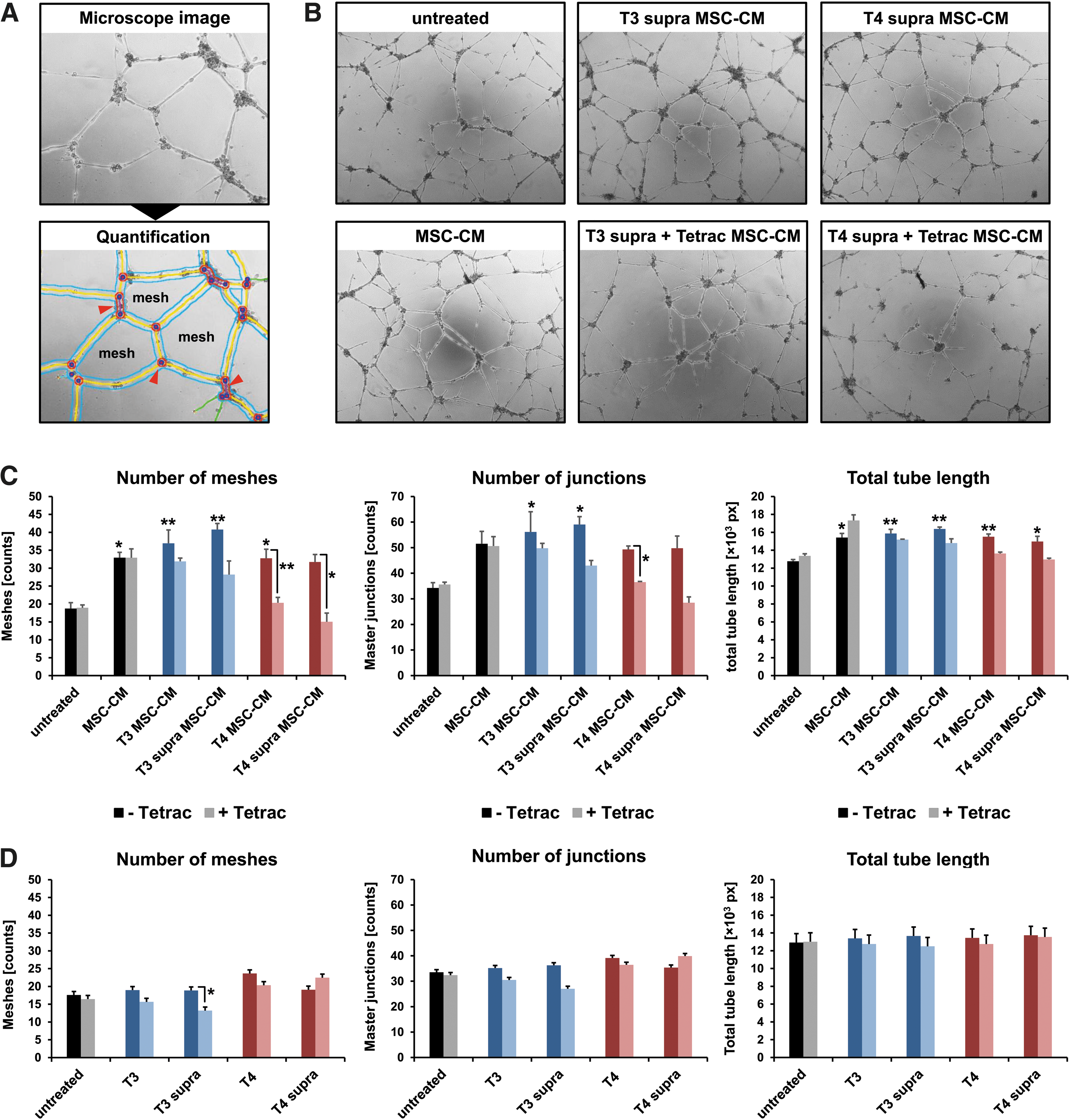
Tube formation assay. (
Compared with untreated HUVECs, supernatant from MSCs enhanced tube formation, as evidenced by more intricate tubular networks with a larger number of meshes and junctions as well as a higher total tube length after 12 hours (Fig. 2B, C). This effect was further enhanced when MSCs had been additionally treated with thyroid hormones (Fig. 2B, C). While Tetrac had no effect on basal HUVEC tube formation, or MSC supernatant-stimulated tube formation, supernatant from MSCs stimulated with Tetrac in addition to thyroid hormone reduced tube formation to, or even below, basal levels (Fig. 2B, C).
To estimate potential effects of residual thyroid hormones in the MSC supernatant on HUVEC tube formation, HUVECs were treated with thyroid hormones alone in a further tube formation experiment. No significant effects were observed at the thyroid hormone concentrations used, although the data do imply a slight inhibiting effect of Tetrac on HUVEC tube formation under parallel T3 stimulation (Fig. 2D).
Thyroid hormone affects VEGF signaling in MSCs in vitro
As the hypoxia response/VEGF axis critically mediates angiogenesis, we established two reporter gene systems to monitor the involvement of HIF-1 and VEGF signaling in thyroid hormone-mediated angiogenic effects of MSCs in more detail. We engineered MSCs to express the reporter gene NIS under the control of the hypoxia-responsive promoter (HRE-NIS-MSCs) or the VEGF promoter (VEGF-NIS-MSCs). We could then use radioiodide uptake by NIS in transfected cells as readout of active hypoxia or VEGF signaling.
To induce promoter activity, HRE-NIS-MSCs were stimulated with the hypoxia-simulating agent CoCl2 and VEGF-NIS-MSCs with FBS. In a functional assay, both HRE-NIS-MSCs and VEGF-NIS-MSCs showed inducible radioiodide uptake activity that was blocked upon treatment with the competitive NIS inhibitor perchlorate (Fig. 3A). While untreated HRE-NIS-MSCs and wild-type MSCs showed no 125I uptake above background levels, unstimulated VEGF-NIS-MSCs showed some basal activity (Fig. 3A).

Hypoxia and VEGF signaling in vitro. Two different reporter constructs were established by placing the NIS gene under control of a hypoxia-responsive promoter (HRE-NIS, left panel) or the VEGF promoter (VEGF-NIS, right panel). (
We then stimulated HRE-NIS-MSCs with HCC cell-conditioned medium and thyroid hormones with or without Tetrac in the presence of CoCl2 to simulate hypoxia and thus stabilize newly expressed HIF-1α. Compared with HRE-NIS-MSCs treated with HCC cell-conditioned medium only, additional stimulation with T3 or T4 led to a slight nonsignificant increase in iodide uptake activity (Fig. 3B, left panel). VEGF-NIS-MSCs showed a stronger more robust response to thyroid hormone stimulation, as evidenced by a significant Tetrac-dependent increase of 1.50- and 1.53-fold for 1 and 10 nM T3 and 1.57- and 1.73-fold for 100 and 1000 nM T4, respectively (Fig. 3B, right panel). Tetrac reduced 125I uptake induced by thyroid hormones in both cell lines, although only reaching statistical significance for VEGF-NIS-MSCs.
To further verify integrin αvβ3 involvement, we applied inhibitors of signaling pathways downstream of the integrin in addition to the integrin-specific inhibitor Tetrac. The PI3K inhibitor LY 294002 was used to probe for T3 signaling via the S1 binding site in the thyroid hormone binding domain of integrin αvβ3 and the ERK1/2 (MAPK) pathway inhibitor RAF265 for T3 and T4 signaling via the S2 binding site (Fig. 3C). In both HRE-NIS-MSCs and VEGF-NIS-MSCs, the T3 effects were blocked upon Tetrac, LY 294002, and RAF265 treatment, while T4 effects were only blocked by Tetrac and RAF265 and not by LY 294002 (Fig. 3D). Note that in this set of experiments, T3 and T4 effects did reach statistical significance in HRE-NIS-MSCs.
Thyroid hormone stimulates VEGF signaling in MSCs in the liver tumors in vivo
Based on the stronger thyroid hormone-dependent effects seen in vitro using the VEGF-NIS-MSCs, compared with the HRE-NIS-MSCs, the former were further evaluated in vivo by 124I PET in an orthotopic HCC xenograft nude mouse model (Fig. 4A). Mice were subjected to different thyroid hormone treatments (euthyroid, hyperthyroid, hypothyroid, and euthyroid+Tetrac; Fig. 4B). Seventy-two hours after systemic injection of VEGF-NIS-MSCs, tumoral 124I accumulation and thus functional NIS expression were assessed by three-dimensional high-resolution small animal PET (Fig. 4B–D).

VEGF signaling in vivo. (
Radioiodide signals in the liver region demonstrated successful tumoral recruitment of VEGF-NIS-MSCs after systemic application, followed by VEGF promoter-driven NIS expression in tumors (Fig. 4C). Compared with euthyroid mice, the tumors of hyperthyroid mice accumulated markedly more radioiodide one hour after 124I injection, while hypothyroid and Tetrac-treated mice accumulated less radioiodide in their liver tumors (Fig. 4C, D).
Tracer uptake was also observed in mouse tissues that endogenously express NIS (thyroid, stomach, nasal mucosa, and salivary glands) as well as in the urinary bladder, due to renal radioiodide elimination (Fig. 4C). The competitive NIS inhibitor perchlorate inhibited both tumoral NIS expression as well as endogenous expression, thus verifying that tracer uptake in tumors was indeed NIS-mediated (Fig. 4C). No 124I uptake above background was observed in tumors of mice injected with wild-type MSCs (Fig. 4C).
Immunofluorescence staining for human NIS in tumors reflected PET imaging results with a significantly higher number of NIS-positive cells in hyperthyroid mice and significantly less in hypothyroid and Tetrac-treated animals compared with euthyroid mice (Fig. 4E, F). Normalization to MSC recruitment confirmed the observed effects on NIS expression levels (Fig. 4G).
Tumor growth was not affected by thyroid hormone status or Tetrac treatment in this model (data not shown). Thyroid hormone concentrations in serum at sacrifice reflected the different thyroid states of the treatment groups (Fig. 4H). The ELISA used could not differentiate between T4 and Tetrac, leading to false high serum T4 values in Tetrac-treated mice (data not shown). Tetrac-treated animals received the same body weight-adjusted dose of T4 as the euthyroid group, which leads to highly reproducible serum T4 levels in the range of untreated naturally euthyroid mice.
To further assess the effects on angiogenesis, ex vivo CD31 straining on tumor sections was performed. The blood vessel densities of the different treatment groups showed a trend reflecting the effects on angiogenesis seen in vitro and in vivo with a significantly decreased tumor vascularization in hypothyroid mice compared with euthyroid mice, and a significantly increased vascularization in hyperthyroid mice compared with hypothyroid and Tetrac-treated mice (Fig. 5A, B).
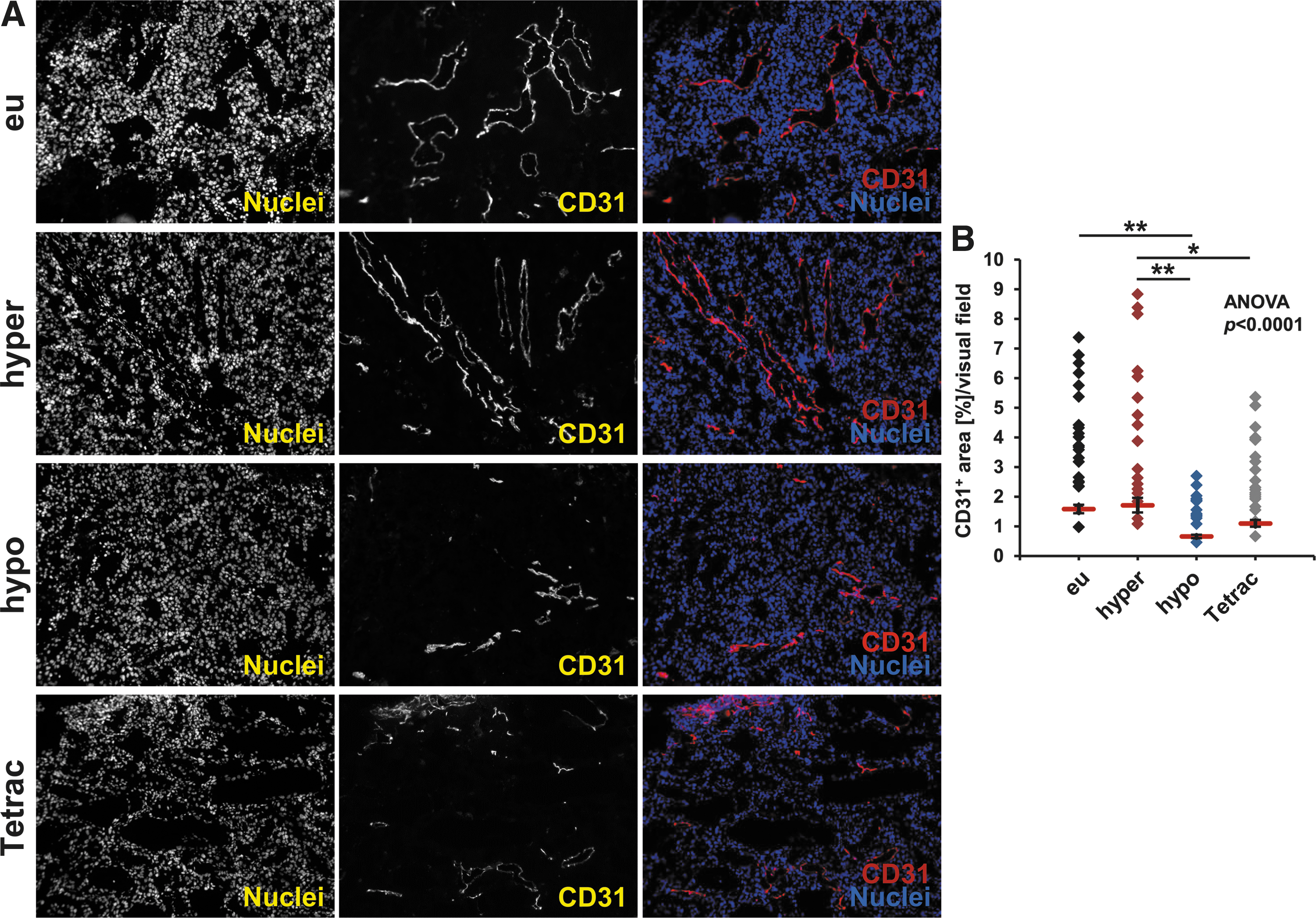
Ex vivo CD31 staining. (
Discussion
Angiogenesis is pivotal to tumor growth and progression. Therefore, understanding the molecular regulation of cellular events involved in angiogenesis has immense clinical implications. MSCs have been shown to secrete pro-angiogenic factors in the tumor microenvironment upon their homing to the tumor stroma and their differentiation into cancer-associated fibroblast- and pericyte-like cells (9,11). It is assumed that the angiogenic effect of MSCs is predominantly caused by their secretion of pro-angiogenic factors (2).
In a previous study, we observed that the thyroid hormones T3 and T4 stimulate the differentiation of MSCs toward a tumor-promoting, pro-angiogenic phenotype via a cell surface receptor on integrin αvβ3 (9). The thyroid hormone metabolite Tetrac, a specific inhibitor of thyroid hormone action at the integrin site, reversed the effects of agonist thyroid hormone.
Indeed, thyroid hormone has been shown to be pro-angiogenic via integrin αvβ3 expressed on tumor cells and endothelial cells in numerous studies (16,20,24). Our data suggest another layer to the story, in that nonhematopoietic cells of the tumor stroma, namely MSCs, can be driven toward a more pro-angiogenic phenotype by thyroid hormones via their signaling through αvβ3 integrin. Hence, we decided to take a closer look at thyroid hormone regulation of angiogenic effects of MSCs in the current study.
Analyzing the response of a broader range of angiogenic factors found in the MSC secretome to thyroid hormone versus Tetrac treatment, we here show that thyroid hormones, especially T3, stimulate pro-angiogenic gene expression and protein secretion in MSCs at physiologically relevant hormone concentrations.
Interestingly, HIF-1α expression was significantly induced by both T3 and T4. The transcription factor HIF-1 is a key modulator of a cell's adaptation to hypoxia and an important angiogenesis mediator. HIF-1 forms a heterodimer composed of the constitutively expressed HIF-1β and the oxygen-sensitive HIF-1α subunit that is tightly regulated and degraded under normoxia (39). Under hypoxia, HIF-1α accumulates leading to transcriptional activation of genes containing hypoxia response elements, such as VEGF (4). The HIF-1α-related pro-angiogenic activity of thyroid hormones is especially interesting in the context of solid tumors that are typically associated with local hypoxia (18).
T3 has been reported to induce HIF-1α expression by a PI3K-dependent mechanism in human skin fibroblasts, an effect that has been proposed to be mediated by cytoplasmically located TRβ (40,41). Furthermore, T3, and not T4, has been shown to induce HIF-1α expression in human glioma cells, again via PI3K (42). Whether PI3K-dependent induction of HIF-1α expression is initiated at the cell surface through integrin αvβ3 or at a cytoplasmically located TR, or both, has, however, not yet been conclusively shown (16,26,41,42).
T4, however, has been shown to induce internalization of integrin αvβ3 in human nonsmall cell lung cancer and ovarian carcinoma cells, resulting in a complex formed between the αv monomer and ERK1/2 (MAPK), leading to nuclear import (43). Tetrac inhibited internalization of integrin αvβ3. In the nucleus, αv acts as a transcriptional co-activator that helps drive the transcription of HIF-1α (18,44,45).
The pro-angiogenic effect of thyroid hormones on MSCs was further confirmed in an in vitro endothelial tube formation assay demonstrating significantly enhanced tube formation under treatment with supernatants from T3-stimulated MSCs. The increased expression and secretion of pro-angiogenic factors by MSCs under thyroid hormone stimulation detected by qPCR and ELISA directly stimulated HUVEC tube formation. The tube formation data also reflect the observation that T3 generally seems to have stronger effects in our system. Many of the effects on gene expression could be directly blocked by additional treatment with Tetrac. The inhibitory effects of Tetrac on angiogenesis were strikingly apparent in the tube formation assay. Thus, we conclude that the observed effects are largely initiated through integrin αvβ3.
Interestingly, we observed no Tetrac effects on basal tube formation, or on the increased tube formation triggered by MSC-conditioned medium. A possible explanation for the strong Tetrac effects only under parallel treatment with thyroid hormone may be due to integrin activation and/or clustering in the cell membrane of MSCs as a result of agonist thyroid hormone stimulation. Tetrac would then be able to exert its inhibitory effects only once this activation has occurred. In this context, Tetrac was shown to prevent radiation-induced activation of the β3 monomer in vitro, while it did not affect the basal nonirradiated activation state (46).
Furthermore, integrin cluster formation and resulting activation is not only known to be essential for extracellular matrix adhesion but has also been proposed to influence integrin-related signaling (47). Indeed, when the binding site for RGD, which provokes integrin clustering, is blocked, thyroid hormone action through integrin is inhibited (23). Whether thyroid hormone influences integrin cluster formation has not been thoroughly investigated to date, although T4 was reported to regulate integrin clustering in astrocytes (48,49).
The observation that co-clustering and resulting cross talk between integrin αvβ3 and angiogenesis-relevant growth factor receptors for VEGF, FGF2, EGF, platelet-derived growth factor, and IGF1 occurs, adds a further dimension to the complex regulation of angiogenesis by thyroid hormones (18,50). Tetrac has been proposed to interfere with this cross talk on the surface of endothelial cells, and similar effects in MSCs are conceivable (18).
HIF-1α and VEGF are two master regulators of angiogenesis with central importance in angiogenic signaling, the expression of which in MSCs has been shown to be regulated by thyroid hormones and Tetrac in the current study (HIF-1α), and in a previous study (VEGF) (9). We therefore established hypoxia-responsive and VEGF-promoter reporter constructs. As reporter gene we used the transmembrane protein NIS that transports iodide into thyroid follicular cells and thus provides the molecular basis for the diagnostic and therapeutic application of radioiodide in thyroid cancer patients (51).
In our previous work, we have gathered extensive experience in targeting NIS to nonthyroidal tumors using genetically engineered MSCs as gene transfer vehicles based on their inherent tumor homing capacity (29,31,33,52 –55). One of the many advantages of NIS is that it allows detailed, noninvasive in vivo tracking of MSCs, as well as quantifiable functional NIS expression, by 123I scintigraphy and 124I PET. Based on these studies, we employed this system to study thyroid hormone action on MSC biology within the tumor microenvironment.
In vitro we observed a thyroid hormone-dependent upregulation of hypoxia-responsive and VEGF promoter activity that was already evident at physiological hormone concentrations and could be blocked by Tetrac, as well as inhibitors of signaling pathways downstream of the hormone binding site at the integrin site. These observations further support the involvement of integrin αvβ3.
Based on the more promising in vitro results using the VEGF-NIS reporter construct and the central role of VEGF in regulating practically all relevant aspects of tumor angiogenesis, we applied VEGF-NIS-MSCs in vivo in an orthotopic HCC xenograft mouse model. We then measured 124I uptake by PET in mice with different thyroid hormone states.
Tumoral radioiodide uptake demonstrated successful recruitment of VEGF-NIS-MSCs after systemic application, followed by VEGF promoter-driven NIS expression. In hyperthyroid animals, a strongly enhanced radioiodide signal was detected compared with euthyroid mice, while hypothyroidism or treatment with Tetrac markedly reduced the signal. Ex vivo analysis of NIS-specific immunoreactivity and NIS mRNA expression normalized to MSC recruitment are consistent with the PET imaging results. These data confirm the in vitro data suggesting thyroid hormone-mediated stimulation of VEGF that is inhibited by Tetrac.
In addition, tumor vascularization reflect the pro-angiogenic effects of thyroid hormones seen in vitro and in vivo as evidenced by a lower blood vessel density in hypothyroid and Tetrac-treated mice compared with euthyroid and hyperthyroid mice.
Our data imply that T3 and T4 influence angiogenic signaling via integrin αvβ3 in the tumor microenvironment not only in tumor cells and endothelial cells but also in MSCs, further establishing MSCs as thyroid hormone-dependent targets through signaling via integrin αvβ3. Stimulation of cancer-related angiogenesis is obviously undesirable in cancer patients and thyroid hormones have been proposed to limit anti-angiogenic therapy (20,56).
Growing experimental evidence, supported by clinical data and population-based studies, suggests that thyroid hormones are involved in cancer development, progression, and metastasis, with established effects on cancer cell proliferation and cancer cell defense pathways, in addition to angiogenesis (16,44,57). Against this background, understanding the complex mechanisms underlying pro-angiogenic effects of thyroid hormones is of paramount importance, not least due to the promising anti-angiogenic activity of Tetrac.
Footnotes
Acknowledgments
We are grateful to Dr. Barbara von Ungern-Sternberg and Rosel Oos (Department of Nuclear Medicine, LMU Munich, Munich, Germany) for their assistance with animal care and PET imaging studies.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by grants from the Deutsche Forschungsgemeinschaft within the Priority Programme SPP1629 to C.S. and P.J.N. (SP 581/6-1, SP 581/6-2, NE 648/5-2), and within the Collaborative Research Center SFB 824 to C.S. (project C8) as well as a grant from the Wilhelm-Sander-Stiftung to C.S. (2014.129.1).