
Abstract
Background:
Although most differentiated thyroid carcinomas (DTCs) have indolent behavior, DTCs with distant metastasis have a poor prognosis. However, there are no validated markers that predict the risk of distant metastasis and the prognosis of DTC. We aimed to develop a genetic classifier for predicting the outcomes of DTC patients with distant metastases.
Methods:
Targeted deep sequencing of 157 cancer-related genes was performed for 61 DTCs with distant metastases. A candidate mutation was validated with independent thyroid cancer samples using digital polymerase chain reaction.
Results:
The most frequently mutated gene in the 61 DTCs was BRAF (n = 31, 51%), followed by TERT promoter (n = 28, 46%), NRAS (n = 13, 11%), PLEKHS1 promoter (n = 6, 10%), and STK11 (n = 6, 10%) mutations. PLEKHS1 promoter mutations were more common in the radioactive iodine (RAI)-refractory cases (p = 0.003). Losses of 9q and 11q were associated with RAI-refractory disease (p = 0.002) and cancer-specific mortality (p = 0.028), respectively. In multivariate analysis, bone metastasis (adjusted odds ratio [aOR] = 15.17, 95% confidence interval [CI 3.38–68.06], p < 0.001) and at least one mutation in the TERT promoter, the PLEKHS1 promoter, or TP53 (aOR = 7.64 [CI 1.78–32.76], p = 0.006) remained significant factors associated with RAI-refractoriness. In independently collected papillary thyroid carcinomas without initial distant metastasis (n = 75), a PLEKHS1 promoter mutation was only found in one case that developed distant metastasis during the follow-up period. We developed a genetic classifier consisting of BRAF, RAS, the TERT promoter, the PLEKHS1 promoter, and TP53 for categorizing the prognosis of patients with DTC with distant metastasis. In the poor-prognosis group, 61% of the patients were RAI-refractory and death occurred in 21% during the follow-up. In the intermediate-prognosis group, 29% were RAI-refractory, but no death occurred. In the good-prognosis group, all patients were RAI-responsive and no death occurred.
Conclusions:
Mutations in the PLEKHS1 promoter are a novel genetic marker of aggressive DTC. Our genetic classifier can be useful for predicting RAI-refractory disease and poor prognosis in DTC patients with distant metastases.
Introduction
Differentiated thyroid carcinoma (DTC) arises from follicular cells and constitutes >95% of all thyroid cancers. DTC is histologically classified as papillary thyroid carcinoma (PTC), follicular thyroid carcinoma (FTC), and poorly differentiated thyroid carcinoma (PDTC). Although the majority of DTCs show indolent behavior compared with most other cancers, 1–9% of DTC patients initially present with locally advanced diseases with distant metastasis and 7–23% develop distant metastasis after initial treatment (1).
Distant metastasis is the main cause of thyroid cancer-related deaths. Disease-specific mortality rates at 5 and 10 years for patients with distant metastases were reported to be 65% and 75%, respectively (2,3). The known risk factors associated with distant metastasis of DTC are lateral lymph node metastasis, histological type (tall cell, columnar cell, hobnail, solid, and diffuse sclerosing variants of PTC, angioinvasive FTC, widely invasive FTC, and PDTC), TERT promoter mutations, and multiple concurrent mutations (4). However, these pathological and genetic factors cannot accurately predict the risk of distant metastasis and the prognosis of individual DTC patients.
Among driver gene alterations in DTC, the two most common events are somatic mutations of the BRAF and RAS genes (5). The progression of DTC to a more aggressive form is associated with the accumulation of mutations in cancer-related genes, including TP53, PIK3CA, AKT1, TERT, histone methyltransferase genes, mismatch repair genes, and the chromatin remodeling complex (5 –7). Mutations in the promoter regions can regulate the transcriptional activity of the genes through altering or disrupting the binding of transcription factors to cis-acting DNA sequences (8,9). For example, TERT promoter mutations increase the promoter activity and transcription of the TERT gene, which activates telomerase (10). TERT promoter mutations occur in 10–20% of DTCs and are associated with distant metastasis, disease recurrence, and increased mortality (11 –16).
Recently, whole-genome sequencing revealed new recurrent mutations in the transcription regulatory regions of some cancer-related genes, such as PLEKHS1, WDR74, and SDHD, which seem to have a potential regulatory impact on the development of cancer (9). They can be used as markers for predicting clinical outcomes including the therapeutic responses of DTC patients. Despite advances in the understanding of the molecular pathogenesis of thyroid cancer, genetic testing results can only partially predict the clinical outcomes of DTC patients. Hence, the identification of genetic markers predicting the prognosis of patients with thyroid cancer is still needed.
In this study, by using targeted deep sequencing designed to detect both coding and noncoding DNA alterations in 157 cancer-related genes, we aimed to identify recurrent somatic mutations and copy number alterations (CNAs) in DTC. We also attempted to develop a genetic classifier for predicting clinical outcomes including the therapeutic responses of DTC patients with distant metastases.
Materials and Methods
Patients and tissue samples
Sixty-one DTC patients treated at Seoul St. Mary's Hospital between 2006 and 2018 were included in the study (48 PTCs, 9 FTCs, and 4 PDTCs). Of the 61 DTC patients, 20 had distant metastases at initial diagnosis and 41 developed distant metastases during follow-up after thyroid surgery. Distant metastases were identified by the pathological examination of surgical specimens (n = 32) and radiological imaging (n = 29), including whole-body scans, computed tomography, magnetic resonance imaging, and positron emission tomography when the pathological diagnosis of metastatic disease was not available.
All pathology slides were reviewed and classified by an endocrine pathologist (C.K.J.) following the diagnostic criteria of the 2017 World Health Organization (17). PTC was classified as a follicular variant if it exclusively consisted of a follicular growth pattern without any true papillae; a classic variant with tall cell features if it consisted of <30% tall cells; and a tall cell variant if it consisted of 30% or more tall cells.
A total of 67 tumor tissues of primary thyroid tumors (n = 46) and distant metastatic tumors (n = 21) from the 61 patients were used for molecular testing. Of them, matched pairs of primary and metastatic tumors were available for six patients. For 15 DTC patients, distant metastatic tumor tissues were used because the primary tumor tissues were absent or inadequate for molecular testing. To validate the clinical utility of metastasis-associated genes, we additionally enrolled 75 PTC patients without distant metastases at the time of initial thyroid surgery.
Cancer staging and treatment strategies were decided by the multidisciplinary institutional Thyroid Cancer Tumor Board, which consists of endocrinologists, thyroid surgeons, radiologists, nuclear medicine radiologists, radiation oncologists, medical oncologists, and pathologists.
The study was approved by the Institutional Review Board of Seoul St. Mary's Hospital of the Catholic University of Korea (KC16SISI0709). Informed consent was obtained from each patient.
Evaluation of response to radioiodine therapy
All 61 DTC patients with distant metastases underwent radioactive iodine (RAI) therapy. The response to RAI ablation was evaluated by whole-body 131I scans; the measurement of serum thyroglobulin levels; and radiologic imaging including computerized tomography scan, magnetic resonance imaging, and positron emission tomography. Therapeutic outcomes to RAI were classified as RAI-responsive and RAI-refractory. RAI-refractory metastatic disease was defined as any of the following (18): (i) at least one distant metastasis without radioiodine uptake, (ii) continued growth of disease despite RAI treatment and confirmed uptake, (iii) distant disease growing over a 1-year period after RAI, and (iv) persistent disease after a cumulative dose of RAI ≥600 mCi.
DNA isolation
Genomic DNA was isolated from 10 μm thick formalin-fixed paraffin-embedded tissue sections using the RecoverAll™ Total Nucleic Acid Isolation Kit (Life Technologies, Carlsbad, CA) according to the manufacturer's instructions. Hematoxylin- and eosin-stained slides were used to select areas of the tumor tissue. Unstained tissue sections matched with the selected areas were manually dissected using a sterile scalpel blade under microscopic guidance.
The quality and quantity of the extracted genomic DNA were initially analyzed by an ND-1000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA), and the DNA concentration was determined using the Qubit™ dsDNA HS Assay Kit on a Qubit 2.0 Fluorometer (Thermo Fisher Scientific).
Targeted deep sequencing
Targeted deep sequencing was performed with genomic DNA obtained from 67 tumor tissues (46 primary thyroid lesions and 21 metastatic lesions) including 6 matched primary and metastatic tissue pairs using the OncoChase cancer panel (ConnectaGen, Seoul, Korea) according to the manufacturer's instructions. The OncoChase cancer panel targets 157 cancer-related genes, and detailed information is available in Supplementary Table S1. Acquisition and processing of the sequencing data were performed as previously described (19). In brief, 101 paired-end sequence reads were generated by using the Illumina HiSeq 2000 platform. A Burrows–Wheeler aligner was used to align the sequencing reads to the human reference genome (UCSC hg19). The aligned sequencing reads were evaluated using Qualimap (20). To understand the mutational characteristics between primary thyroid and metastatic tumors, we analyzed the six matched pairs of primary and metastatic tumors.
Identification of somatic variants and driver mutations
Single nucleotide variants (SNVs) and indels were identified using the MuTect (21) and SomaticIndelDetector (22), respectively. The ANNOVAR package (23) was used to select somatic variants located in the exonic sequences and to predict their functional consequences. For reliable and robust calling, the following variants were eliminated (24): (i) read depths <20 in the tumor and (ii) polymorphisms referenced in either the 1000 Genomes Project or the Exome Aggregation Consortium with a minor allele frequency of 0.1% or more in East Asians. Variants of >0.1% in minor allele frequency in our in-house normal database (38 whole-genome and 2283 whole-exome sequencing data from Koreans) were also filtered out. The remaining variants were considered as candidate somatic variants. Among the filtered variants, the mutations overlapped with the COSMIC database (>30 times) were manually rescued.
DNA copy number analysis
DNA copy number profiling was performed using the targeted deep sequencing data. The multiscale reference module and the SNPRank Segmentation statistical algorithm in NEXUS software v10 (Biodiscovery, El Segundo, CA) were used to define the CNAs in each sample (25). Segments were classified as gains and losses when the log2 ratio was greater than 0.18 and less than −0.18, respectively. All the identified CNA events were manually curated in terms of depth ratio.
Validation and replication of targeted deep sequencing results
We validated the mutations detected by targeted deep sequencing based on digital polymerase chain reaction (PCR). The primers and probes used for the validation are available in Supplementary Table S2. Digital PCR was performed using the TaqMan Genotyping assay and a QuantStudio 3D digital PCR system (Life Technologies) as described elsewhere (26). Briefly, 10 ng of genomic DNA was amplified with the TaqMan genotyping probe. In each assay, the reference and mutant alleles were labeled with reporter dyes 2′-chloro-7′phenyl-1,4-dichloro-6-carboxyfluorescein and 6-carboxyfluorescein, respectively. After scanning the nanofluidic chip, the raw data were subsequently analyzed using the Rare Mutation module of the QuantStudio 3D AnalysisSuite Cloud Software.
Statistical analyses
The molecular analyses were performed blinded to clinicopathologic data, and conversely, the clinicopathologic utility of the molecular markers was determined blinded to the molecular results. The relationships between the clinicopathologic features and the mutational profiles were analyzed using parametric (chi-squared) and nonparametric (Fisher's exact) tests. Logistic regression was performed to assess the associations of the clinicopathologic variables and mutations with the RAI treatment response. Disease-specific survival curves were plotted using the Kaplan–Meier method, and statistical differences between the curves were tested by the log-rank test. Disease-specific survival was defined as the time interval between the initial diagnosis of distant metastasis and death from DTC or the last follow-up. All statistical analyses were performed using Prism (version 6.05; GraphPad Software, La Jolla, CA) and SPSS (version 21.0; IBM Corp, Armonk, NY). p-Values of <0.05 were considered statistically significant.
Results
Patient characteristics
The general characteristics of the patients are shown in Table 1. Among the 61 DTC patients with distant metastases, 48 had PTCs (21 classic PTCs, 11 tall cell variants, 5 PTCs with tall cell features, 3 infiltrative follicular variants, 2 columnar cell variants, 2 diffuse sclerosing variants, 2 invasive encapsulated follicular variants, 1 solid variant, and 1 hobnail variant). Nine patients had FTCs (eight encapsulated angioinvasive FTCs with extensive vascular invasion [n = 6] and limited vascular invasion [n = 2], one minimally invasive FTC) and four had PDTCs. The lungs were the most common site of distant metastasis, followed by the bones. Sixteen patients (26%) had both lung and bone metastases. Twenty-four patients had aggressive histology (11 tall cell variant PTCs, 2 columnar cell variant PTCs, 1 hobnail variant PTC, 6 encapsulated angioinvasive FTCs with extensive vascular invasion, and 4 PDTCs). The median cumulative RAI dose was 300 mCi (range, 100–900 mCi). Of the 61 patients, 25 (41%) were RAI-refractory. After the median follow-up of 4.0 years (range, 0.6–11.9 years), 7 patients died from tumor progression and 54 patients survived or were lost to follow-up.
Clinicopathologic and Molecular Features of Differentiated Thyroid Cancer Presenting with Distant Metastasis
FTC, follicular thyroid carcinoma; PDTC, poorly differentiated thyroid carcinoma; PTC, papillary thyroid carcinoma; RAI, radioactive iodine.
Targeted deep sequencing of metastatic thyroid cancer genomes
To analyze the mutational profiles of the 61 DTCs with distant metastasis, we sequenced 46 primary thyroid and 15 metastatic tumors from the 61 patients using targeted deep sequencing of 157 cancer-related genes. The mean sequencing depth was 477.1 × (range, 104.0 × –1272.9 × ) across the entire genome (Supplementary Table S3). A total of 203 somatic mutations (201 SNVs and 2 indels) were identified, and the majority of the metastatic DTCs (58/61, 95%) had nonsilent mutations in at least one target gene (Supplementary Table S4).
The most frequent variants in the 61 DTCs with distant metastasis were identified in the BRAF (n = 31, 51%), TERT promoter (n = 28, 46%), NRAS (n = 13, 11%), PLEKHS1 promoter (n = 6, 10%), and STK11 (n = 6, 10%) genes (Fig. 1A).
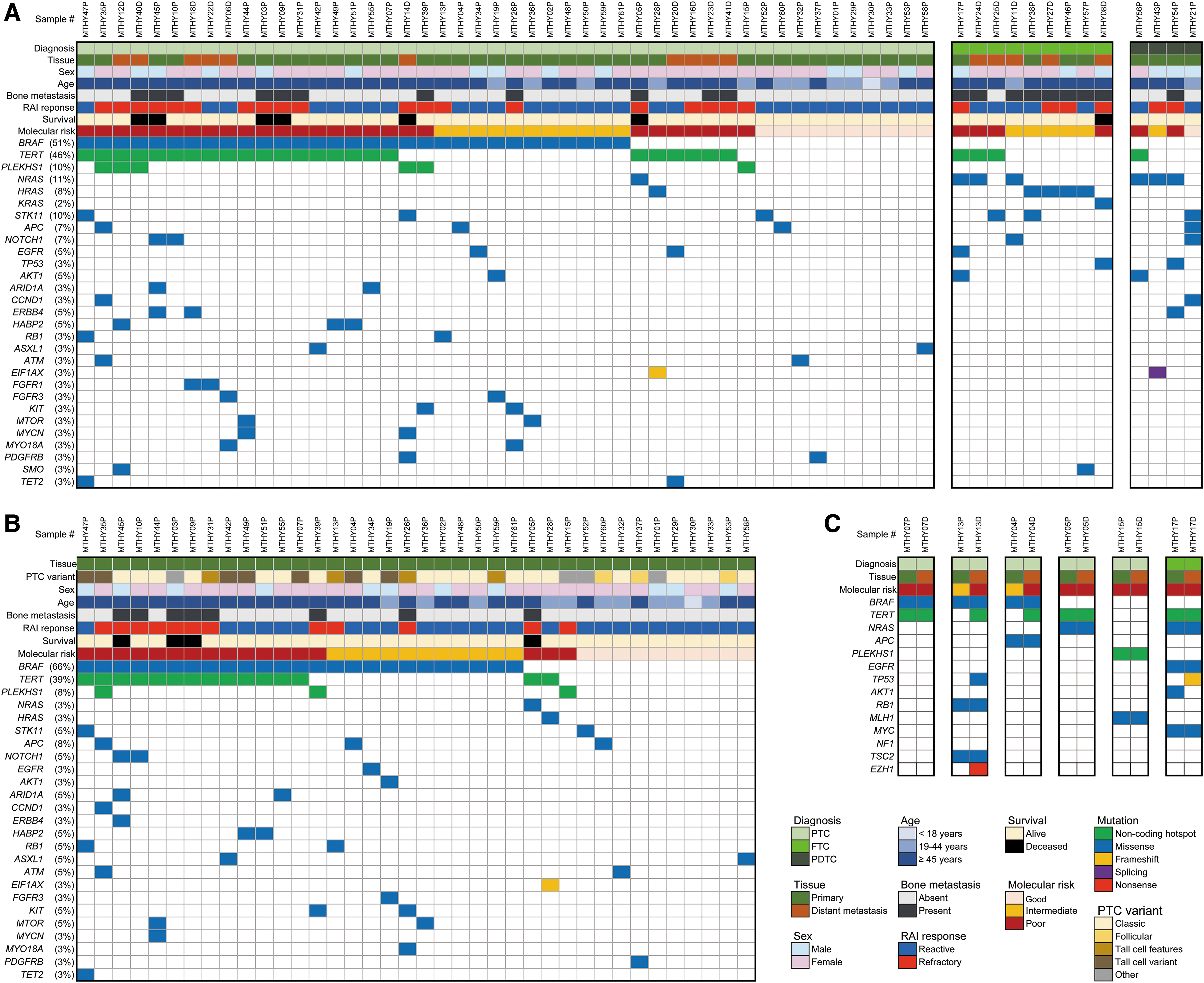
Frequent somatic mutations and CNAs in DTC with distant metastasis. Clinicopathologic features and molecular profiling of all 61 patients performed on 46 primary and 15 distant metastatic tissues (
TERT promoter mutations were found in 24 (50%) of 48 PTCs, 3 (33%) of 9 FTCs, and 1 (25%) of 4 PDTCs. Among the TERT promoter mutations, the most common type was C228T (n = 26, 43%), followed by C250T (n = 2, 3%), both of which are already known as noncoding promoter hot spot sites (14,16) (Supplementary Fig. S1A).
The BRAF V600E mutation was the predominant BRAF mutation (Supplementary Fig. S1B) and identified only in PTCs (n = 31, 65% of PTCs) (Fig. 1A).
PLEKHS1 promoter mutations were identified only in PTCs (Fig. 1A). Of 48 PTCs, the 6 (13%) with PLEKHS1 promoter mutations consisted of 3 tall cell, 1 classic, 1 diffuse sclerosing, and 1 hobnail variant. All six PLEKHS1 promoter mutations were located in known hot spot sites (C593T, n = 4 and G590A, n = 2) (9) (Supplementary Fig. S1C).
All RAS mutations were detected at hot spot codon 61, one NRAS p.Q61R and one HRAS p.Q61R in two PTCs; three NRAS p.Q61R, four HRAS p.Q61R, and one KRAS p.Q61R in eight FTCs; and two NRAS p.Q61R and one NRAS p.Q61K in three PDTCs (Fig. 1A).
All six STK11 mutations were p.F354L, seen in three PTCs, two FTCs, and one PDTC (Fig. 1A).
Besides these seven genes, mutations in well-known cancer-related genes were also detected in the metastatic DTCs: TP53 (n = 2; p.M237I, p.S240G), AKT1 (n = 3; p.E17K), EIF1AX (n = 2; p.A113fs, c.338–1G>A), PIK3CA (n = 1; p.E545K and p.E726K double mutation), and TSHR (n = 1; p.A623V) (Fig. 1A, Supplementary Table S4).
Among the six patients with PLEKHS1 promoter mutations, three had coexisting BRAF V600E and TERT promoter mutations, one had coexisting BRAF V600E and PIK3CA mutations, one had a BRAF V600E mutation, and one had an MLH1 p.V704L mutation (Supplementary Table S4). Of three cases with AKT1, two had a coexisting mutation of the TERT promoter and one had a BRAF V600E mutation (Fig. 1A). Of two cases with EIF1AX mutations, one had coexisting HRAS and TERT promoter mutations and the other had a coexisting NRAS mutation (Fig. 1A). One case with a TSHR mutation harbored a coexisting mutation of the TERT promoter (Supplementary Table S4).
Molecular profiling of primary PTC tissues was available for 38 patients (Fig. 1B). The most frequently mutated genes were BRAF V600E (n = 25, 66%), TERT promoter (n = 15, 39%), and PLEKHS1 promoter (n = 3, 8%).
Comparison of mutational characteristics between primary tumor and distant metastasis
We analyzed the matched pairs of primary thyroid and distant metastatic tumors in six cases (five PTCs and one FTC) (Fig. 1C). Of 14 mutant genes identified in 6 primary thyroid tumors, 10, including PLEKHS1, were found in both the primary and the corresponding metastatic tumors. In two cases (MTHY04 and MTHY13), mutations in the TERT promoter and TP53 were not identified in primary tumors but were present in their matched metastatic tumors. In one case (MTHY17P), an AKT1 mutation was found in the primary tumor but not in its metastatic site. An EZH1 mutation was only found in one metastatic tumor (MTHY13).
PLEKHS1 promoter mutations associated with distant metastasis
The majority of the PLEKHS1 promoter mutations (5/6) occurred at low (range, 2.6–6.8%) variant allele frequencies (VAFs). The mean VAF of the PLEKHS1 promoter mutations (median of 4.0%) was significantly lower than the other mutations detected in >10% of the metastatic DTCs (Supplementary Fig. S2). To validate the PLEKHS1 promoter mutations identified by targeted deep sequencing, we performed digital PCR and demonstrated similar VAFs for all mutations (Supplementary Table S5). Next, we performed digital PCR for the PLEKHS1 promoter mutations in the primary tumors of independent PTC patients without distant metastasis (n = 75) at the time of initial surgery to verify whether the PLEKHS1 promoter mutation was a metastasis-associated event. A PLEKHS1 promoter mutation was found in only one case (1/75, 1.3%), in a patient whose tumor showed aggressive histology (hobnail variant), extrathyroidal extension, and central lymph node metastasis. When we checked the medical history of the mutation-positive patient in the four years since the initial surgery, a chest computed tomography scan showed multiple bilateral metastatic nodules in the lungs three years after thyroidectomy, further supporting that PLEKHS1 promoter mutations are very rare in PTC without distant metastasis. Except for this single case, no distant metastases were observed in the other 74 cases.
CNAs in metastatic thyroid cancer genomes
We analyzed the CNA profiles of the 51 metastatic thyroid cancers using the targeted deep sequencing data (Fig. 2). Ten cases were excluded due to high background noise levels. Among the 106 CNAs identified (Supplementary Table S6), 22 CNAs appeared recurrently (>2 cases) (Supplementary Table S7). The most highly recurrent CNA (n = 18, 35%) was a 22q11.1–q13.33 deletion, a 35 Mb-sized region that encompasses the NF2, EP300, and CHEK2 genes (Supplementary Fig. S3). Fourteen (34%) of 41 PTCs, 3 (43%) of 7 FTCs, and 1 (33%) of 3 PDTCs harbored this deletion. A recurrent copy number loss of 9p24.3–p11.2, 9q12–q34.3, and 16q11.2–q24.3, and a gain of 1q12–q44 were detected only in the PTCs and their frequencies were 15% (n = 6), 22% (n = 9), 12% (n = 5), and 20% (n = 8) of 41 PTCs, respectively.
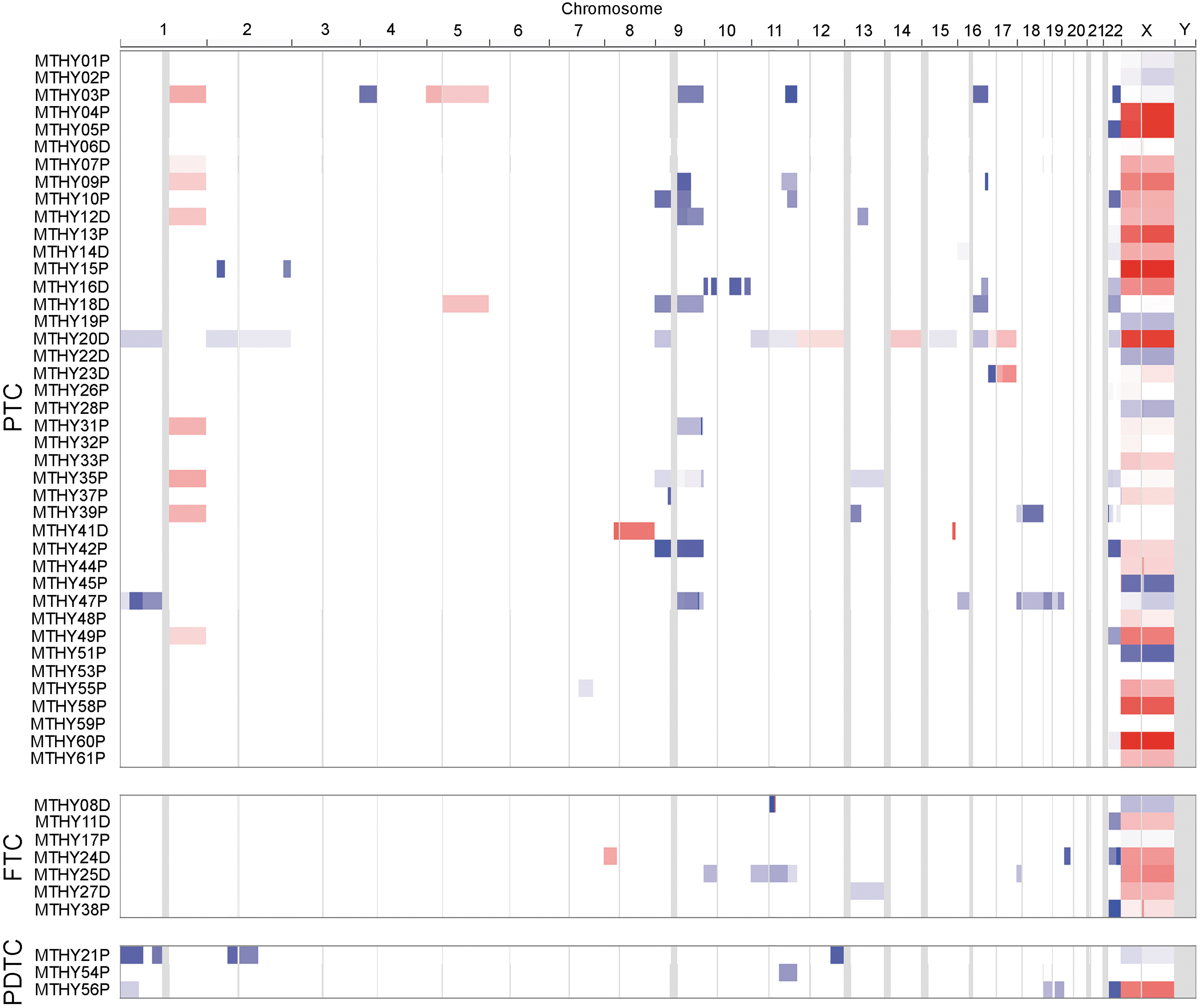
CNAs in 51 DTCs with distant metastasis stratified by cancer types.
Clinicopathologic significance of recurrent mutations and CNAs
As shown in Table 1, BRAF V600E (p = 0.031) and TERT promoter (p = 0.002) mutations were found more frequently in patients older than 45 years. RAS mutations were found more frequently in patients with bone metastases (p < 0.001). PLEKHS1 promoter mutations were more common in RAI-refractory cases (p = 0.003).
Regarding the CNAs, a loss of 11q was significantly associated with bone metastasis (p = 0.034) and cancer-specific death (p = 0.028). BRAF V600E mutations were significantly more common in the cases with 1q gains (p = 0.005) and 9q losses (p = 0.002). A loss of 9q was also significantly more common in the RAI-refractory cases (p = 0.002) (Supplementary Table S8).
Factors related to RAI refractivity
In univariate analyses, clinical factors significantly associated with RAI refractivity were age ≥45 years (p = 0.021) and bone metastasis (p < 0.001) (Table 2). To improve the predictive ability of the RAI responses of patients with DTCs, we added combinations of the recurrent mutations as independent variables. The existence of any mutations in the TERT promoter, the PLEKHS1 promoter, or TP53 was significantly associated with RAI refractivity (p = 0.001). Sex and aggressive histology were not significantly associated with RAI refractivity.
Univariate and Multivariate Logistic Regression Analyses for Risk Factors of Therapeutic Response to Radioactive Iodine Ablation in the Differentiated Thyroid Cancer Presenting with Distant Metastasis
Variables showing a tendency of association with recurrence (p < 0.25) in the univariate analysis were included in the multivariate model.
CI, 95% confidence interval; OR, odds ratio.
In multivariate logistic regression, bone metastasis (adjusted odds ratio [aOR] = 15.17, 95% confidence interval [CI 3.38–68.06], p < 0.001) and the existence of any mutations in the TERT promoter, or the PLEKHS1 promoter, or TP53 (aOR = 7.64 [CI 1.78–32.76], p = 0.006) remained significant (Table 2).
Cancer-specific survival of patients with metastatic DTC
RAI-refractory patients showed significantly worse survival (p = 0.005). Bone metastasis (p = 0.009), mutations in at least one of the three genes (TERT promoter, PLEKHS1 promoter, and TP53) (p = 0.023), and the 11q loss CNAs (p < 0.001) were significantly associated with worse cancer-specific survival (Fig. 3A–C). The coexistence of BRAF V600E or RAS mutations with one of the three high-risk genes (TERT promoter, PLEKHS1 promoter, and TP53) had no additional impact on the metastatic DTC survival rates (data not shown). Age of ≥45 years at the diagnosis of distant metastasis (p = 0.090) also showed worse cancer-specific survival, but it was not statistically significant. There was no relationship between aggressive histology and cancer-specific survival (data not shown).
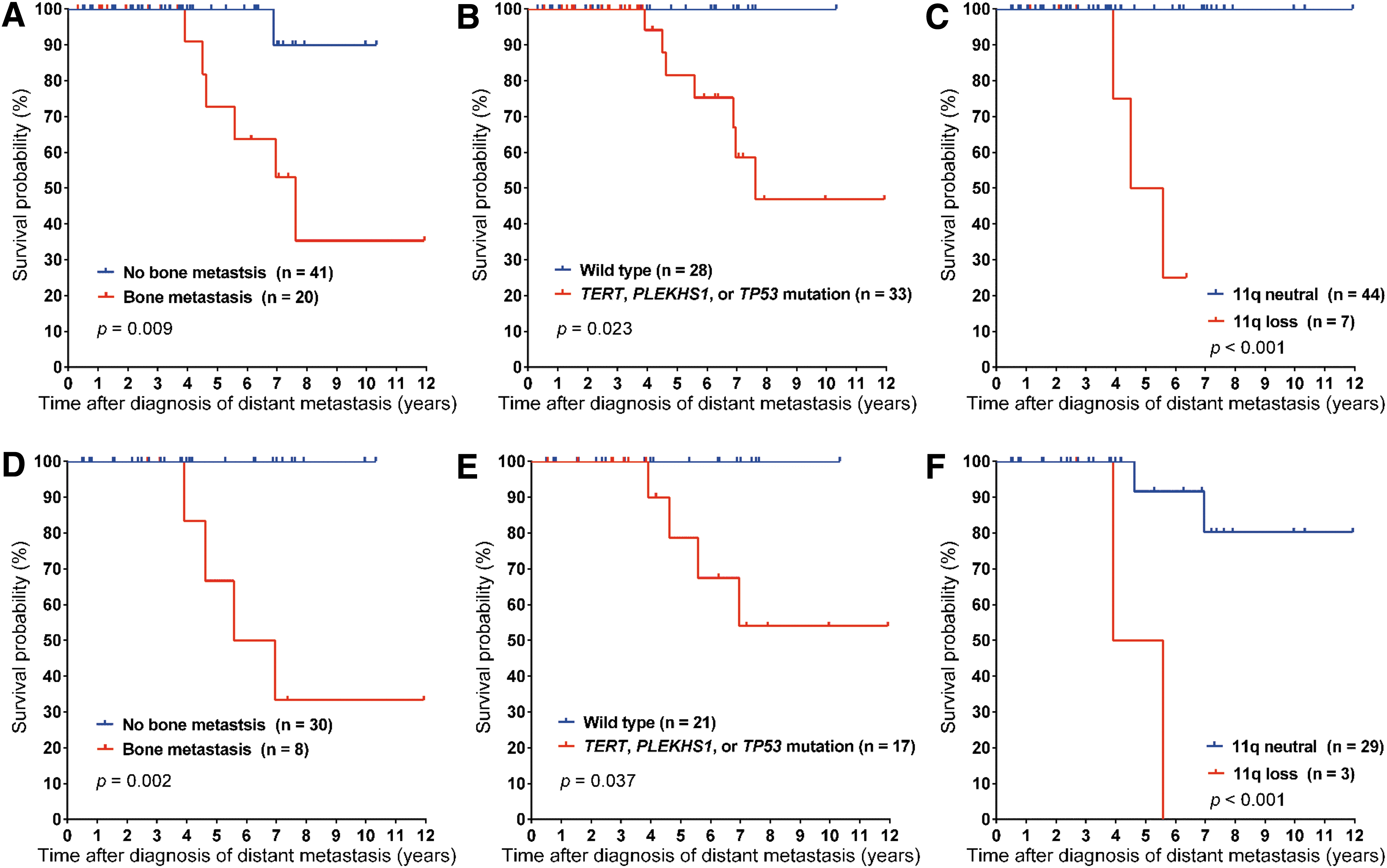
Kaplan–Meier curves of cancer-specific survival by bone metastasis (
Cancer-specific survival of patients with metastatic PTC
We further analyzed the impact of the prognostic factors described above on cancer-specific survival in a subgroup of 38 primary PTC tissues. Bone metastasis (p = 0.002), mutations in at least one of the three high-risk genes (p = 0.037), and the 11q loss CNA (p < 0.001) were significantly associated with worse cancer-specific survival (Fig. 3D–F).
Genetic classifier and risk stratification of metastatic DTC
Using the recurrently mutated genes, we aimed at developing a genetic classifier for predicting therapeutic response and prognosis based on the following principles: (i) selecting mutations found in >3.0% including mutations supported by literature evidence, such as BRAF V600E and RAS for basic molecular classification (5), and TERT promoter and TP53 mutations for predicting poor prognosis (6,14,16); (ii) including mutations statistically associated with adverse outcomes (distant metastasis, RAI refractoriness, and disease-specific survival) in our study. According to these principles, seven genes (BRAF, three RAS isoforms, TERT promoter, PLEKHS1 promoter, and TP53) were selected. A loss of 11q was not selected because CNA data were missing in 10 cases.
Analyzing the relationships between clinical outcomes and the mutational profiles of these genes allowed us to develop a new genetic classifier that could categorize DTC patients with distant metastases into three prognostic groups (Fig. 4): Poor-prognosis group (54%, 33/61 DTCs): the patients harbored at least one mutation in the TERT promoter, the PLEKHS1 promoter, or TP53 regardless of the mutational status of BRAF and RAS. In this group, the rate of RAI-refractory tumors was 61% (20/33), and all seven deaths occurred in this group (21%, 7/33) during the follow-up period. The 5-, 7-, and 10-year cancer-specific survival rates in this group were 82%, 59%, and 47%, respectively. Intermediate-prognosis group (28%, 17/61 DTCs): the patients harbored at least one mutation in either BRAF or RAS and no mutations in the TERT promoter, the PLEKHS1 promoter, or TP53. In this group, the rate of RAI-refractory tumors was 29% (5/17), but no deaths occurred. Good-prognosis group (18%, 11/61 DTCs): the patients harbored no mutations in the seven included genes. In this group, the rate of RAI-refractory tumors was 0% and no death occurred.
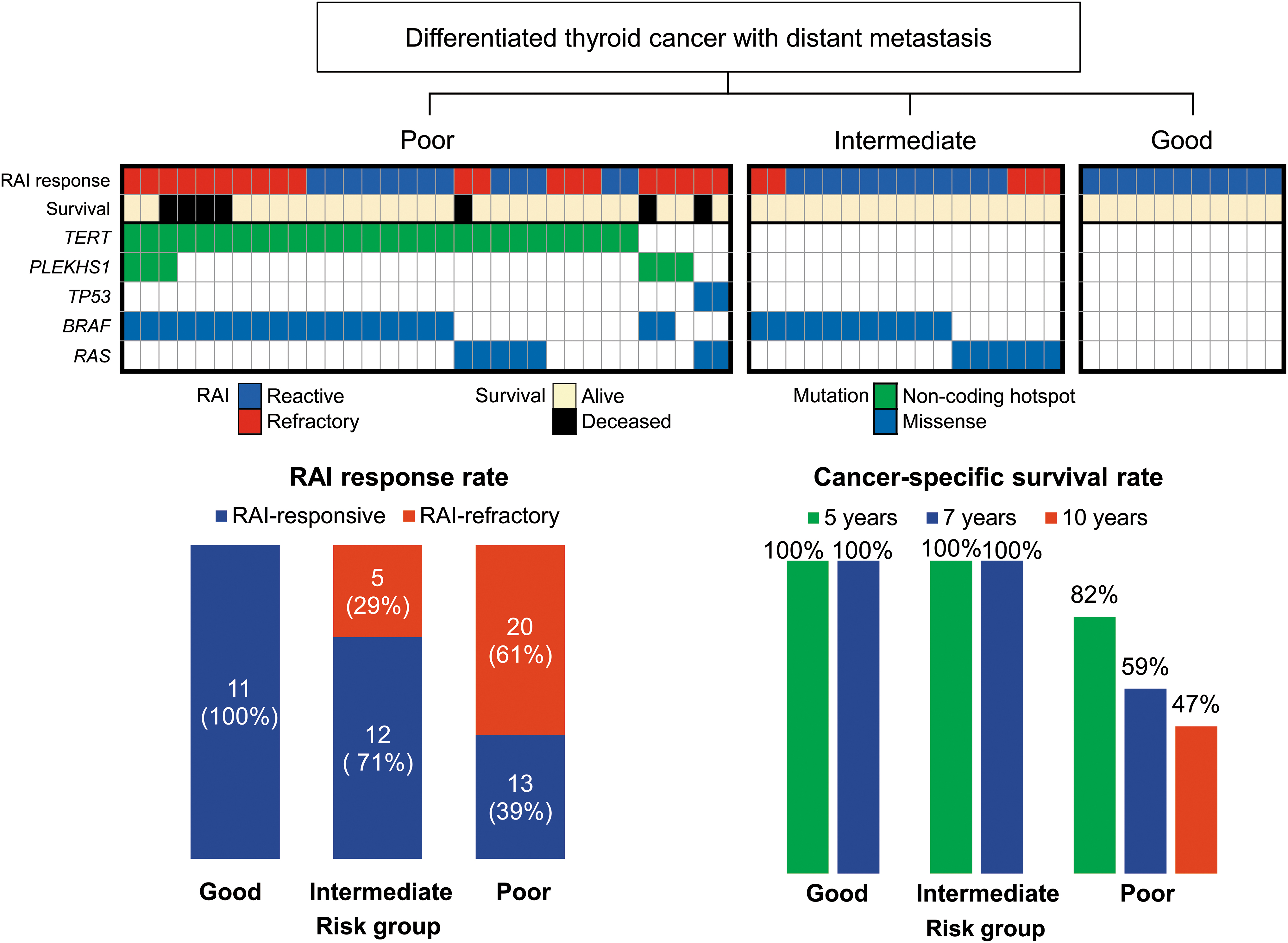
Risk stratification for RAI response and cancer-specific survival in patients with metastatic DTC according to the mutational profiles of BRAF, three RAS isoforms, the TERT promoter, the PLEKHS1 promoter, and TP53 genes. Patients in the poor-prognosis group had at least one mutation in the TERT promoter, the PLEKHS1 promoter, or TP53 genes. Intermediate-prognosis patients had BRAF or RAS mutations and no mutations in the TERT promoter, the PLEKHS1 promoter, and TP53. Good-prognosis patients had no mutations in these genes. RAS mutations included NRAS (n = 7), HRAS (n = 5), and KRAS (n = 1).
Discussion
DTC patients who develop distant metastasis are known to have poorer survival. More than half of these patients are refractory to RAI, which contributes to even worse prognosis (18). It is important to identify the patients at high risk of distant metastasis early in the treatment process. Metastatic DTCs frequently have TERT promoter mutations, but their existence alone is insufficient to predict the risk of distant metastasis because more than half of the metastatic DTC patients do not have this mutation (13,14). Through this study, we suggested that PLEKHS1 promoter mutations, which occurred recurrently (10%), were a potential prognostic factor for DTC metastasis. PLEKHS1 promoter mutations were found in 9% (3/33) of metastatic DTCs without TERT promoter mutations. We developed a genetic classifier based on the mutational profiles of BRAF, the TERT promoter, the PLEKHS1 promoter, three RAS isoforms, and TP53, which could classify metastatic DTC patients into three prognostic groups in terms of RAI treatment response and survival.
The PLEKHS1 gene contains a pleckstrin homology domain that may play a role in intracellular signaling (9). Two hot spot mutations in the PLEKHS1 promoter are located at 115, 511, 590 bp (C590T) and 115, 511, 593 bp (C593T) on chromosome 10, which have been reported in some solid cancers including bladder, breast, and lung cancers (9,27). However, the clinical implications of PLEKHS1 promoter mutations in human cancers have not been well studied. The analysis of PLEKHS1 expression from The Cancer Genome Atlas (TCGA) and other public sources showed that individuals with promoter hot spot mutations had lower expression than those without the mutations in bladder cancer and all tumor types (9). However, the role of PLEKHS1 in tumorigenesis is largely unknown. In the present study, the majority (5/6) of the PLEKHS1 promoter mutations showed low VAFs (range, 2.6–6.8%) suggesting that acquisition of the PLEKHS1 promoter mutation might be a later event during tumor progression. To the best of our knowledge, this is the first report that PLEKHS1 promoter mutations may be associated with distant metastasis and RAI refractoriness in DTC patients.
A subset of well-differentiated thyroid cancer may become RAI-refractory at metastatic sites and undergo dedifferentiation to PDTC and anaplastic thyroid cancer. These tumors frequently harbor mutations in the TERT promoter and various genes including members of the Wingless/int-1 signaling pathway, the phosphatidylinositol 3-kinase (PI3K)/protein kinase B (AKT) pathway, the switch/sucrose nonfermentable chromatin remodeling complex, histone methyltransferases, DNA mismatch repair, and tumor suppressors (6). Among mutations affecting the PI3K/AKT pathway, PIK3CA and AKT1 mutations are associated with tumor progression and distant metastasis of DTC (28 –30). When we analyzed the mutational profiles in the PTDC (n = 84) and anaplastic thyroid cancers (n = 33) in patients from Memorial Sloan Kettering Cancer Center (MSKCC) using the cBioPortal for Cancer Genomics, seven of eight cancers with PIK3CA mutations had concurrent TERT promoter mutations (6). The mutation of other genes mostly coexisted with TERT promoter and/or TP53 mutations (Supplementary Fig. S4). In another study performed at the MD Anderson Cancer Center, PIK3CA mutations were found in 6 of 190 advanced DTC cases with distant metastases or persistent/recurrent local disease (30). Of six tumors with PIK3CA mutations, one had a coexisting TP53 mutation. In our study, 33 (54%) of 61 cases had at least 1 mutation either in the TERT promoter, the PLEKHS1 promoter or the TP53 gene. A PIK3CA mutation was found in one case with a concurrent PLEKHS1 promoter mutation (Supplementary Table S4). Other uncommon mutations related to advanced thyroid cancers mostly coexisted with mutations of these three genes (Supplementary Table S4). Therefore, we thought that a genetic classifier based on the TERT promoter, the PLEKHS1 promoter, or TP53 mutations could improve the clinical utility of other recurrent genetic alterations including PIK3CA and AKT1 mutations, in predicting the treatment outcomes of DTC patients.
A big challenge in managing metastatic DTC is to find a balance between active treatment and active surveillance. The prognosis of metastatic DTC is usually favorable. Even when DTC patients are refractory to RAI treatment, some of them show good long-term survival without additional therapy (4). In the case of RAI-refractory metastatic tumors, additional therapy should be tailored to patients based on the extent of the metastatic diseases, symptoms, comorbidities, and the rate of progression (4). In the current American Thyroid Association management guidelines for DTC, molecular testing is not routinely recommended for prognostic purposes in patients with RAI-refractory or metastatic DTC due to lack of robust evidence (4). In this study, we developed a genetic classifier that could stratify metastatic DTC patients into three prognostic groups (good, intermediate, and poor). Patients in the good-prognosis group had stable disease, whereas more than half of the poor-prognosis group patients died of progressive diseases within 10 years. Therefore, this genetic classifier can be a clinically useful tool to help predict patients who are likely to become RAI-refractory and have a poor prognosis during follow-up. This system may also help guide physicians to make strategic decisions on therapeutic goals and other treatment options besides RAI for patients with metastatic DTC.
Germline mutations of the tumor suppressor serine-threonine kinase 11 (STK11) gene were initially identified in Peutz–Jeghers syndrome (31). Somatic mutations of the STK11 gene have also been found in many solid tumors, including thyroid (6), lung (32), pancreatic and biliary (33), liver (34), and colon cancers (35). In a previous study from MSKCC, STK11 mutations were found in 1% of the PDTCs and 6% the patients with anaplastic thyroid cancer (6). In our study, STK11 mutations were found in 10% (6/61) of all DTCs. However, we could not rule out the possibility of germline variants when considering its minor allele frequency in the East Asian population (3–5%) and high VAF, ranging from 39.9% to 85.1%, in our study (Supplementary Fig. S2).
The analysis of somatic CNAs from TCGA showed that a 1q gain was frequently found in tall cell variant PTC and BRAF V600E mutations and was associated with more aggressive forms of PTC (5). The loss of 22q was enriched in the follicular variant of PTC (5). In the MSKCC study, 1q gains and 22q losses were found in 26% and 14% of 57 patients with fatal DTCs (35 PDTCs, 18 PTCs, and 4 Hürthle cell carcinomas), respectively (36). In 18 PTC cases, 1q gains and 22q losses were identified in 5 (28%) and 2 (11%) tumors, respectively (36). In our study, a 1q gain was found in 20% (8/41) of the PTCs and enriched for BRAF V600E mutations but was not related to the tall cell variant. The loss of 22q was found in 34% (14/41) of the PTCs and 43% (3/7) of the FTCs and was not associated with a follicular growth pattern. Differences among the studies, including ours, could be caused by the limited number of cases and differences in study designs. However, the high frequency of 1q gains and 22q losses in our study was largely consistent with previous TCGA and MSKCC studies (36), suggesting that 1q gains and 22q losses (22q11.1–q13.33) may play roles in conferring aggressiveness in DTC.
A limitation of our study is the small sample size, which may preclude us from deriving conclusive insights into the mutational profiles and their clinical significance. Of the 61 DTC patients, only 46 primary thyroid tumor tissues were available for molecular analysis. In the analysis of primary and paired-metastatic tumor tissues, mutations in high-risk genes TERT and TP53 occurred more frequently in metastatic tumors. The mutational changes were used by our genetic classifier system to reclassify the initial risk as poor risk. All patients classified into the poor-risk group using our molecular algorithm for the metastatic tumor tissue were refractory to RAI therapy. Therefore, the molecular classifier may be better at predicting patient outcomes and prognosis from the analysis of tumor tissues obtained from metastatic tumor sites rather than primary tumor tissue. Despite this limitation, we reconfirmed the prognostic implications of mutations in the TERT promoter and TP53 in metastatic DTC and demonstrate that PLEKHS1 promoter mutations may be associated with distant metastasis of PTC. The genetic classifier system presented here can add useful information for predicting the risk of mortality or unfavorable outcomes in DTC patients.
Footnotes
Author Disclosure Statement
No potential conflicts of interest were disclosed.
Funding Information
This study was supported by grants from the Korean Health Technology R&D Project (HI16C2013) of the Ministry of Health and Welfare and the National Research Foundation of Korea (2017R1D1A1B03029597, 2017R1E1A1A01074913, 2019R1C1C1004909, and 2019R1A5A2027588).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
Supplementary Table S5
Supplementary Table S6
Supplementary Table S7
Supplementary Table S8