
Abstract
Background:
Pathological conditions of the thyroid hormone (TH) system are routinely diagnosed by using serum concentrations of thyrotropin (TSH), which is sufficient in most cases. However, in certain conditions, such as resistance to TH due to mutations in THRB (RTHb) or TSH-releasing pituitary adenoma (TSHoma), TSH may be insufficient for a correct diagnosis, even in combination with serum TH concentrations. Likewise, under TH replacement therapy, these parameters can be misleading and do not always allow optimal treatment. Hence, additional biomarkers to assess challenging clinical conditions would be highly beneficial.
Methods:
Data from untargeted multi-omics analyses of plasma samples from experimental thyrotoxicosis in human and mouse were exploited to identify proteins that might represent possible biomarkers of TH function. Subsequent mouse studies were used to identify the tissue of origin and the involvement of the two different TH receptors (TR). For in-depth characterization of the underlying cellular mechanisms, primary mouse cells were used.
Results:
The analysis of the plasma proteome data sets revealed 16 plasma proteins that were concordantly differentially abundant under thyroxine treatment compared with euthyroid controls across the two species. These originated predominantly from liver, spleen, and bone. Independent studies in a clinical cohort and different mouse models identified CD5L as the most robust putative biomarker under different serum TH states and treatment periods. In vitro studies revealed that CD5L originates from proinflammatory M1 macrophages, which are similar to liver-residing Kupffer cells, and is regulated by an indirect mechanism requiring the secretion of a yet unknown factor from hepatocytes. In agreement with the role of TRα1 in immune cells and the TRβ-dependent hepatocyte-derived signaling, the in vivo regulation of Cd5l expression depended on both TR isoforms.
Conclusion:
Our results identify several novel targets of TH action in serum, with CD5L as the most robust marker. Although further studies will be needed to validate the specificity of these targets, CD5L seems to be a promising candidate to assess TH action in hepatocyte-macrophage crosstalk.
Introduction
Thyroid hormones (THs) are essential regulators for development and homeostasis, hence their physiological ranges have to be tightly controlled. THs occur in the biologically active form 3,3′,5-triiodothyronine (T3) and the less biologically active transport form L-thyroxine (T4). Their secretion by the thyroid gland is stimulated by the hypophyseal thyrotropin (TSH), which, in turn, is stimulated by the hypothalamic thyrotropin releasing hormone (TRH). Both TSH and TRH are negatively regulated by THs, constituting a self-adjusting negative feedback loop termed the hypothalamic-pituitary-thyroid axis (HPT axis) (1,2). Within this axis, TSH and THs show a log-linear relationship (3), implicating high TSH variation already at minor TH changes (4). Therefore, TSH measurement is the standard method to evaluate TH status in patients.
Although TSH concentrations are commonly used to diagnose most thyroid disorders, lab-specific differences and interferences in TSH measurement may occasionally cause misleading results. Consequently, TSH measurements are often combined with TH determinations (free T4 [fT4] and/or free T3 [fT3]). Although this approach sufficiently allows to diagnose the majority of thyroid-related problems, certain conditions still require additional biomarkers for correct diagnosis (5). This is exemplified for the differential diagnosis between carriers of mutations in TH receptor (TR) β (resistance to TH, RTHβ) and patients with TSH-secreting pituitary adenomas, both of which present with elevated TSH concentrations in the presence of high circulating T3 and/or T4 levels (6).
Monotherapy with levothyroxine (LT4), the treatment of choice for the substitution of primary hypothyroidism, represents another example as a subnormal fT3 status is still observed in 15% and an abnormal fT3/fT4 ratio in >30% of all cases despite TSH and fT4 levels being carefully titrated into their physiological ranges. This indicates an incomplete substitution not captured by the conventional diagnostic approach (7) and has resulted in a highly controversial discussion, whether L-triiodothyronine (LT3) should be added during the supplementation therapy, although a clear beneficial effect has yet to be demonstrated and a valid method to assess tissue-specific effects is missing (8 –11).
A major factor that determines the individual and unique condition of each affected patient is the tissue-specific distribution of TH transporters (MCT8, MCT10, OATP1c1, or LAT1/2) (12), activating/deactivating deiodinases 1, 2, or 3 (DIO1, DIO2, or DIO3) (13), or TRs isoforms α1 and β (TRα1 or TRβ) (14,15), which may vary from case to case due to subtle changes in gene expression, such as single-nucleotide polymorphisms (16,17) or different epigenetic control (18 –20), within the HPT axis. In more extreme conditions, inactivating mutations can occur in these essential components of TH signaling, which usually result in various forms of TH resistance (RTH) (21 –23).
The first descriptions of genetic causes for RTH were mutations in the T3-binding domain of TRβ, causing RTHβ with its typical clinical symptoms such as palpable goiter, palpitation, resting tachycardia, reduced bone mineral density, and higher resting energy expenditure, together with increased fT3 and fT4 and inadequately normal to high TSH level (24,25). Resistance to TH caused by a mutation in THRA encoding TRα1 (RTHα) has been discovered only recently, and it leads to severe phenotypes including epiphyseal dysgenesis, increased mineral bone density, delayed and reduced mental development, bradycardia, and reduced resting energy expenditure, with normal or high-normal TSH, low or low-normal total T4 (tT4), and high or high-normal total T3 (tT3), resulting in a low T4/T3 ratio (21,22,26).
In both forms of RTH, serum TH and TSH levels do not reflect tissue-specific TH actions, as the unequal distribution of TRs leads to a tissue-specific resistance to the circulating levels of TH. TRα1 is the abundant receptor in heart, intestine, bone, brain, and innate immune cells, whereas TRβ is predominately expressed in liver, kidney, retina, inner ear, hypothalamus and pituitary (15). These examples illustrate that additional biomarkers on tissue-specific TH action would be highly desirable for clinical diagnosis. Consequently, several attempts have been made in the past decades to establish such biomarkers, including total cholesterol (27), low-density lipoproteins (27), osteocalcin (28,29), creatine kinase (30), ferritin (30), testosterone-binding globulin (25), tissue plasminogen activator (31), angiotensin-converting enzyme (32), and glucose-6-phosphate dehydrogenase (33). However, none of these markers was recommended for clinical use in recent guidelines because of their low sensitivity and/or specificity or lack of availability for routine testing (3).
Many of these markers are predominantly liver derived such as lipid parameters or sex hormone binding globin (SHBG), which has been suggested for the differential diagnosis of inappropriately high TSH (34). As SHBG is co-regulated by estrogens, androgens, insulin, and prolactin, it lacks specificity (and sensitivity) (8,11,35). Because the liver is predominantly controlled by TRβ-specific markers, it has been tested to differentiate RTHβ from TSH-secreting adenomas and RTHβ. It has been shown that measurement of serum Se and Cu could be helpful (36,37). The ratio of these easily measurable trace elements that are not subject to sample degradation have been proposed for the differential diagnosis of RTHβ in mice and humans, but these interesting findings need to be further confirmed in larger clinical trials.
In summary, new TH-dependent biomarkers are urgently needed to provide more detailed information about TH actions within specific target tissues such as the liver. For this study, we screened for new common biomarkers in mouse and human liver by using a translational proteomics-based approach. In an attempt to better characterize the diagnostic potential and limitations, we subsequently evaluated the underlying molecular mechanism of TH action for the most promising candidate, CD5L, and tested its relation to TH status in clinical samples.
Materials and Methods
Animal husbandry and treatment
Animals were housed in groups under constant 12 hours/12 hours light/dark cycle at ambient temperature (22°C ± 1°C) and had ad libitum access to food and water. TRβ−/− and TRα1+/m mutant mice (38,39) were bred in the animal core facility Lübeck and compared with TRβ+/− or TRα1+/+ littermates. Wild-type C57BL/6NCrl animals were purchased from Charles River Laboratories GmbH (Erkrath, Germany) and rendered hypothyroid with methimazole (0.1% m/v) (M8506; Sigma-Aldrich Chemie GmbH, Taufkirchen, Germany) and sodium perchlorate (0.2% m/v) (106564; Merck KGaA, Darmstadt, Germany) for three weeks in the drinking water. Mild or severe thyrotoxicosis was induced by adding either T4 (T2376; Sigma-Aldrich Chemie GmbH) at 1 mg/L or T3 (T6397; Sigma-Aldrich Chemie GmbH) at 0.5 mg/L to the drinking water, respectively. To maintain regular drinking behavior, 0.1% m/v bovine serum albumin (01400.1; BIOMOL GmbH, Hamburg, Germany) was added into the drinking water of the thyrotoxic and the untreated control groups, and 40 pills/L of the nondigestive sweetener, consisting of 33.8% sodium cyclamate (E952) and 3.8% sodium saccharin (E954) (Suketter; Cederroth International, Väsby, Sweden), was added into the drinking water of the hypothyroid group. All animal procedures were approved by the Schleswig-Holstein Ministerium für Energiewende, Landwirtschaft, Umwelt, Natur und Digitalisierung, Kiel.
Analysis of omics datasets
Plasma proteome data of a previously conducted human longitudinal thyrotoxicosis study were further analyzed (40,41). Briefly, 16 healthy male volunteers were experimentally rendered hyperthyroid by a daily oral administration of 250 μg LT4 for eight weeks. Plasma samples were drawn before, during, at the end of an eight-week T4 administration, and after a subsequent eight-week recovery period. Plasma proteins were analyzed by using an untargeted LC-MS/MS approach after depletion of six abundant plasma proteins with an MARS-6 column.
The plasma proteomes of animals from a previously performed mouse study (42), in which one group of C57BL/6 wild-type mice was rendered hyperthyroid for two weeks (thyrotoxicosis group), one group was rendered hyperthyroid for two weeks followed by two weeks of recovery (recovery group), and one control group remained untreated for four weeks (control group), were analyzed by applying a similar untargeted proteomics workflow as for the human samples but substituting the MARS-6 by an MARS-3 column.
For target gene identification, four filter steps were used: (1) exclusion of proteins with FDR-adjusted p-values >0.05, (2) comparison of recovery group or control group with the thyrotoxicosis group and exclusion of proteins with a fold change < −/+1.5, (3) exclusion of all proteins that are not present in both comparisons, and (4) exclusion of all discordantly altered proteins between mice and humans. The final list after all filtering steps can be found in Table 1. Microarray-based analysis of the hepatic transcriptome was performed as previously described (43). For identifying target gene expressing tissues, the databases UniProt (44) and Expression Atlas (EMBL-EBI, Hinxton, United Kingdom) were used. For creating the heatmap and performing a hierarchical clustering, the R package “complexHeatmap” was used.
Plasma Proteins with Significantly Altered Abundance Between T4 and Control Groups and T4 and Recovery Groups, But No Difference Between Control and Recovery Groups in Human and Mouse Proteome
The fold change represents the abundance alterations of proteins between T4 and control groups.
T4, thyroxine.
ELISA
Serum concentrations of tT4 (EIA-1781; DRG Diagnostics GmbH, Marburg, Germany) and tT3 (DNOV053; NovaTec Immundiagnostica GmbH, Dietzenbach, Germany) were measured with commercially available immunoassays in accordance to the manufacturer's instructions.
Western Blot analysis
A set of serum samples with relatively elevated or suppressed TSH values, respectively, were selected from a cross-sectional study of thyroid patients (45). Serum aliquots (0.4 μL per sample) were separated by SDS-PAGE in a 10% SDS gel at 80 V. After blotting onto a nitrocellulose membrane (Protan™; Merck, Darmstadt, Germany), total protein was quantified by Ponceau S staining. CD5L was detected by rabbit antibodies against the N-terminus of human CD5L (AHP1169; Bio-Rad Laboratories GmbH, Feldkirchen, Germany) in combination with goat anti-rabbit-immunoglobulin-HRP antibody (P0448; Agilent Dako, Santa Clara, CA). Relative CD5L expression was quantified by using ImageJ 1.48v (National Institutes of Health).
Separation of macrophages and parenchymal cells from mouse livers
Mouse livers were perfused and digested as previously described to allow for the isolation and separation of parenchymal and nonparenchymal cells (46). Parenchymal cells (mainly hepatocytes) were pelleted by centrifugation at 35 g for 3 minutes. To obtain liver macrophages, supernatants were centrifuged at 650 g for 7 minutes; pellets were subsequently resuspended in Dulbecco's modified Eagle's medium (DMEM) and subjected to selective plating by using uncoated tissue culture flasks. After two to three hours, flasks were vigorously washed to remove nonadherent cells. Adherent cells were collected as the liver macrophage fraction.
RNA isolation, cDNA generation, and gene expression analysis
Soft tissue in general
Twenty to thirty milligrams of frozen tissue was transferred into 1 mL QIAzol lysis reagent (79306; Qiagen GmbH, Hilden, Germany) and homogenized in a potter-type tissue grinder. Subsequent isolation steps were in accordance with the manufacturer's instructions. RNA samples were further purified by using NucleoSpin RNA Clean-up Kit (740948.250; Macherey-Nagel GmbH & Co. KG, Düren, Germany) in accordance with the manufacturer's instructions.
Bones
Surrounding soft tissue and bone marrow was removed, and bone was cut into three to five pieces before deep freezing in liquid nitrogen. After acclimatization in the vapor phase above a liquid nitrogen reservoir, 1 mL QIAzol was added and the tissue was homogenized with a hard tissue lyser (TH-02; Omni International, Kennesaw, GA). RNA isolation and purification were performed as described for soft tissue.
Primary cell culture
Cells for RNA isolation were seeded in six-well plates, washed with HBSS buffer (14175095; ThermoFisher Scientific, Waltham, MA), and lysed with 500 μL QIAzol lysis reagent. Further RNA isolation and purification was performed with the Direct-zol RNA kit (R2052; Zymo Research Europe GmbH, Freiburg, Germany) in accordance with the manufacturer's instructions. For cDNA generation, 500 ng total RNA was reverse transcribed by using RevertAid First Strand cDNA Synthesis kit (K1622; ThermoFisher Scientific) following the manufacturer's instructions.
Gene expression was determined by using FastStart Universal SYBR Green Master (04913914001; Roche Diagnostics GmbH, Mannheim, Germany) and Mastercycler RealPlex2 (Eppendorf Vertrieb Deutschland GmbH, Wesseling-Berzdorf, Germany). Efficiency and specificity of real-time quantitative PCR (RT-qPCR) runs were determined by standard and melting curves in each run, and gene expression was normalized to housekeeping genes as indicated. Primer sequences can be found in Supplementary Table S1.
Hepatic total RNA used for transcriptome analysis
RNA from liver tissue that was subsequently used for the array-based transcriptome analyses was prepared and quality controlled as previously described (43).
Primary cell and cell line culture
All cells and cell lines were incubated at 37°C in a humidified 5% CO2-containing atmosphere.
Osteoblasts
Mesenchymal stem cells were isolated from femur and tibia of wild type, TRβ−/−, or TRα1+/m mice and seeded in six-well plates at a density of 1 × 106 cells/cm2 in DMEM (41965-039; ThermoFisher Scientific) +20% fetal bovine serum (FBS) (10270106; ThermoFisher Scientific) +1% Penicillin/Streptomycin (P/S) (15140122; Gibco by Life Technologies, Ltd., Paisley, United Kingdom) for five days. Calcification of differentiated osteoblasts was induced by reducing medium to 10% FBS and adding 1 μM of glycerophosphate (G9422; Sigma-Aldrich Chemie GmbH, Munich, Germany) and 100 nM ascorbic acid phosphate (A8960; Sigma-Aldrich Chemie GmbH). Mature osteoblasts were cultured for 14 days, and medium was changed every second day.
Osteoclasts
Bone marrow cells were isolated and seeded at a density of 1 × 106 cells/cm2 in α-minimum essential medium (F-0915; Biochrom GmbH, Berlin, Germany) with 10% FBS, 1% P/S, 1 mM
Bone marrow-derived macrophages
Stem cells from femur and tibia of wild type, TRβ−/−, or TRα1+/m mice were flushed out and seeded at a density of 1 × 105 cells/cm2 in Roswell Park Memorial Institute (RPMI) 1640 medium (A10491-01; ThermoFisher Scientific) with 1% P/S and 20 ng/mL mouse M-CSF. At day 5, M-CSF was removed from medium and replaced with 100 ng/mL lipopolysaccharides (LPS) Escherichia coli O111:B4 (L2630; Sigma-Aldrich Chemie GmbH) and 20 ng/mL interferon γ (IFNγ) (130-105-774; Miltenyi Biotec GmbH, Bergisch Gladbach, Germany) for 24 hours.
Primary hepatocytes
Mouse livers were perfused with 20 mL of Earle's balanced salt solution (14155048; ThermoFisher Scientific) with 500 μM EGTA (E3889; Sigma-Aldrich Chemie GmbH) followed by 20 mL of Hank's balanced salt solution (L2035; Biochrom GmbH) with 0.3 mg/mL collagenase type 1 (LS004196; Worthington Biochemical Corp., Lakewood, NJ). Liver cells were shaken out of the liver capsule, washed, and finally plated at a density of 2 × 104 cells/cm2 in six-well plates in DMEM containing 10% FBS, 1% P/S, 1%
LX-2 cell line
Cells were grown in DMEM containing 2% FBS and 1% P/S and were split every 2–3 days or when they reached 80% confluency.
Stimulation procedure
FBS for the stimulation procedure was stripped with Amberlite IRA-400 chloride form (247669; Sigma-Aldrich Chemie GmbH) for 24 hours, to deplete THs, as described elsewhere (47). Cells were conditioned 12 hours before stimulation in cell-type specific medium with 1% FBS, washed with HBSS, and finally stimulated for 24 hours in cell-type specific medium with 1% TH-depleted FBS or 1% native FBS for control groups. This medium, which was conditioned by either LX-2 or primary hepatocytes, was subsequently transferred onto conditioned bone marrow-derived macrophages (BMDMs).
Statistical analysis
For all statistical analyses, Graphpad Prism 6 (Graph Pad Software, San Diego, CA) was used. Statistical significance of differences between variables was determined by an unpaired two-tailed Student's t-test and Welch's correction and was considered significant at p < 0.05 (*), p < 0.01 (**), p < 0.001 (***), and p < 0.0001 (****). Values are illustrated as mean ± SEM. Pearson correlation coefficient was determined for calculating variable dependence, and a two-tailed p-value was used to ascertain statistical significance.
Results
Target gene identification by an untargeted omics approach
For the identification of new TH-dependent serum biomarkers, a comparative plasma proteome analysis was performed by using data from a previously conducted experimental human and mouse thyrotoxicosis study (Fig. 1A) (40,41). We decided to use this two-species approach for several reasons. First, we aimed at eliminating any genetic effects in the human sample by confirming the effects in genetically homogenous wild-type animals. Second, we reasoned that any regulation that could be detected consistently in both mice and humans would be more robust, and third, this approach would allow us to use organs or cells from animal models lacking TR for in-depth studies.

Identification of biomarker candidates in previously conducted thyrotoxicosis studies in human and mice. (
After filtering for statistically significant differences in protein levels between the euthyroid (control or recovery) groups and the thyrotoxic T4-treated groups, we identified 32 common plasma proteins, with 16 of these concordantly affected across species (Fig. 1B+C and Table 1). A subsequent comparison with the respective liver transcriptomes revealed that three of these candidates, CD5L, IL1AP, and TTR, could be of hepatic origin.
Induction of acute thyrotoxicosis in wild-type mice
To confirm whether the genes encoding these 16 proteins are, indeed, differentially expressed in hypothyroidism and thyrotoxicosis, we conducted an additional independent animal study, in which wild-type mice were treated with either a mixture of methimazole and perchlorate to induce hypothyroidism and T4 or T3 to induce states of mild or severe thyrotoxicosis in the drinking water, respectively (Fig. 2A). The efficiency of this treatment was confirmed with serum tT4 and tT3 ELISA (Fig. 2B, C), confirming the expected hypo- or hyperthyroidism. We then determined gene expression of all 16 candidate genes in liver, spleen, and bone, as these tissues were identified by database searches as being potentially relevant sources for the circulating proteins. Altogether 14 genes were found to be expressed in the liver; the mRNA levels of Angt, Apob, Camp, Fbln1, Il1ap, and Ttr were not affected by TH status. Statistically significant changes in MMI/ClO4-treated mice were found for the transcript levels of Apoa1, Apoc3, Fetua, Lum, and Pgrp2; while in the moderately thyrotoxic T4-treated group those of Apoc3, Apoc4, and Cd5l were significantly altered (Fig. 2D). In addition, expression of Apoc4, Cd5l, Lum, and Vcam1 was significantly increased in highly thyrotoxic T3-treated animals (Fig. 2D). The known hepatic TH target genes Spot14 (48) and Dio1 (49) were included as positive controls and showed the expected positive regulation in the hyperthyroid groups. In spleen, the expression of 6 of our 16 candidate genes was changed with Cd5l and Vcam1, showing decreased transcript levels in the MMI/ClO4 group, but no changes in hyperthyroid groups (Fig. 2E). In bone, transcripts of six of the candidate genes were detected, but only Cd5l revealed altered gene expression in T3-treated animals compared with controls (Fig. 2F). To test which of the hepatic expressed genes display a good correlation with serum tT3 or tT4 levels, we plotted the mRNA levels and serum tT3 or tT4 levels of the individual animals. The transcript levels of Apoc3, Apoc4, Cd5l, Fetua, Lum, and Vcam1 correlated significantly with serum tT3 with the exception of Vcam1, and also with serum tT4 levels (Supplementary Fig. S1A–L), suggesting that these genes constitute valuable novel readouts for hepatic TH action in animal models, in addition to well-known targets such as Dio1 and Spot14.
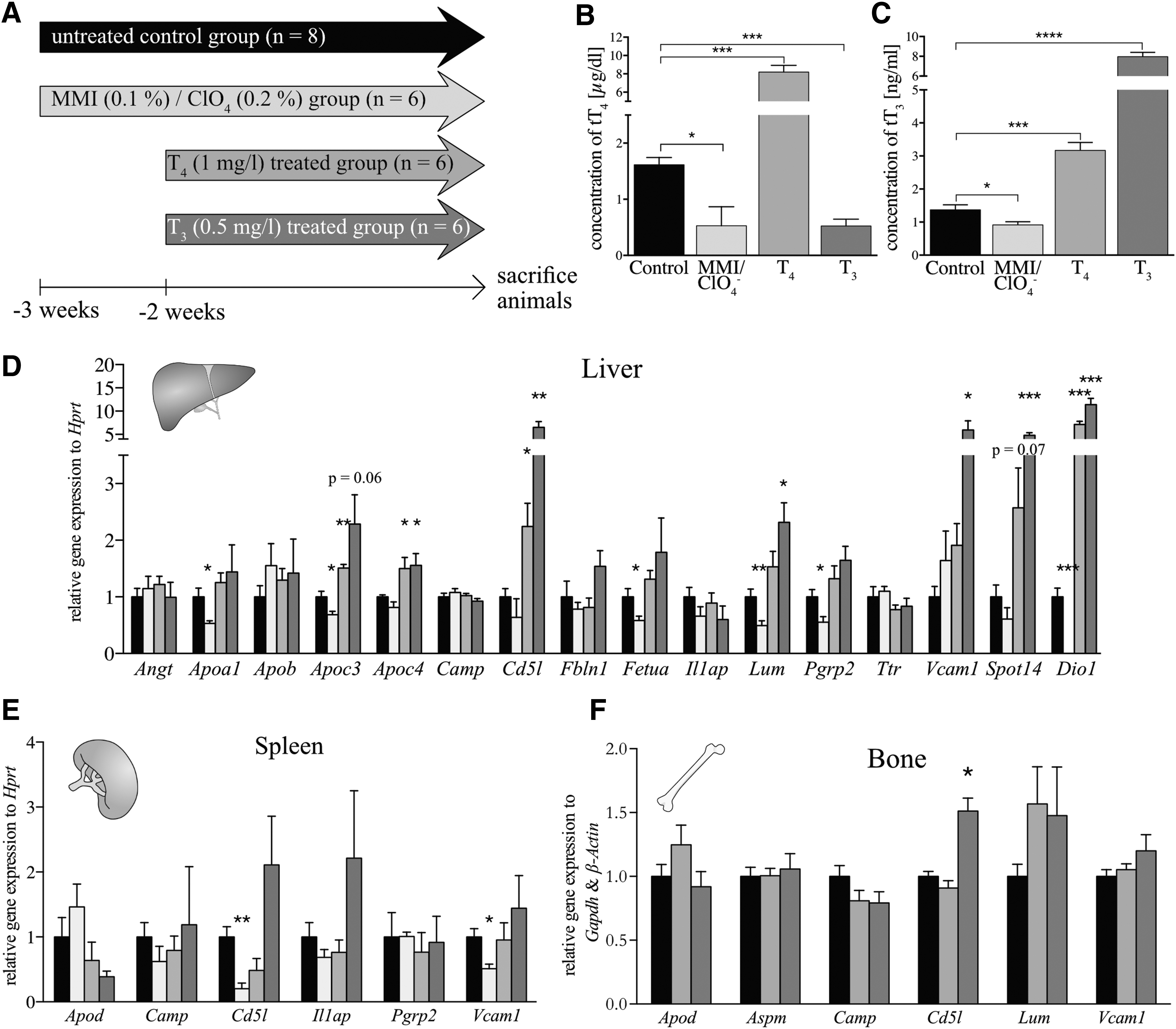
Validation of biomarker candidates under thyrostatic or thyrotoxic conditions. (
Short-term effects of THs on potential target genes
To evaluate whether the candidate genes rapidly respond to TH, which would point to a direct rather than indirect target of TH action, we conducted a short-term thyrotoxicosis study. To this end, mice were rendered hyperthyroid with T4 (1 mg/mL) for 4 days or with T3 (0.5 mg/mL) for 24 hours in drinking water (Fig. 3A). The resulting serum tT4 and tT3 concentrations of the T4-treated group (Fig. 3B+C) already reflected a similar degree of hyperthyroidism and hyper-T3-emia as after the two-week treatment (Fig. 2B+C). Surprisingly, in T3-treated animals, serum tT3 concentrations were even more increased (Fig. 3C) as compared with the two-week treatment (Fig. 2C). Subsequent RT-qPCR analysis of hepatic gene expression (Fig. 3D) revealed decreased mRNA amounts of Angt as well as increased transcript levels of Cd5l and Fbln1 on T4 treatment, with a similar trend for Cd5l (p = 0.09) and Fbln1 (p = 0.06) in the T3-treated mice. The positive controls Spot14 and Dio1 showed a fast and direct response as expected. Most interestingly, Cd5l expression was strongly and significantly correlated with serum tT4 (p = 0.0143) and tT3 (p = 0.0133) concentrations (Supplementary Fig. S2A+B), suggesting that this gene could, indeed, represent a suitable biomarker for hepatic TH action. In bone, Aspm, Cd5l, and Lum expression was reduced in the T3 group compared with the T4 and control groups (Fig. 3E).
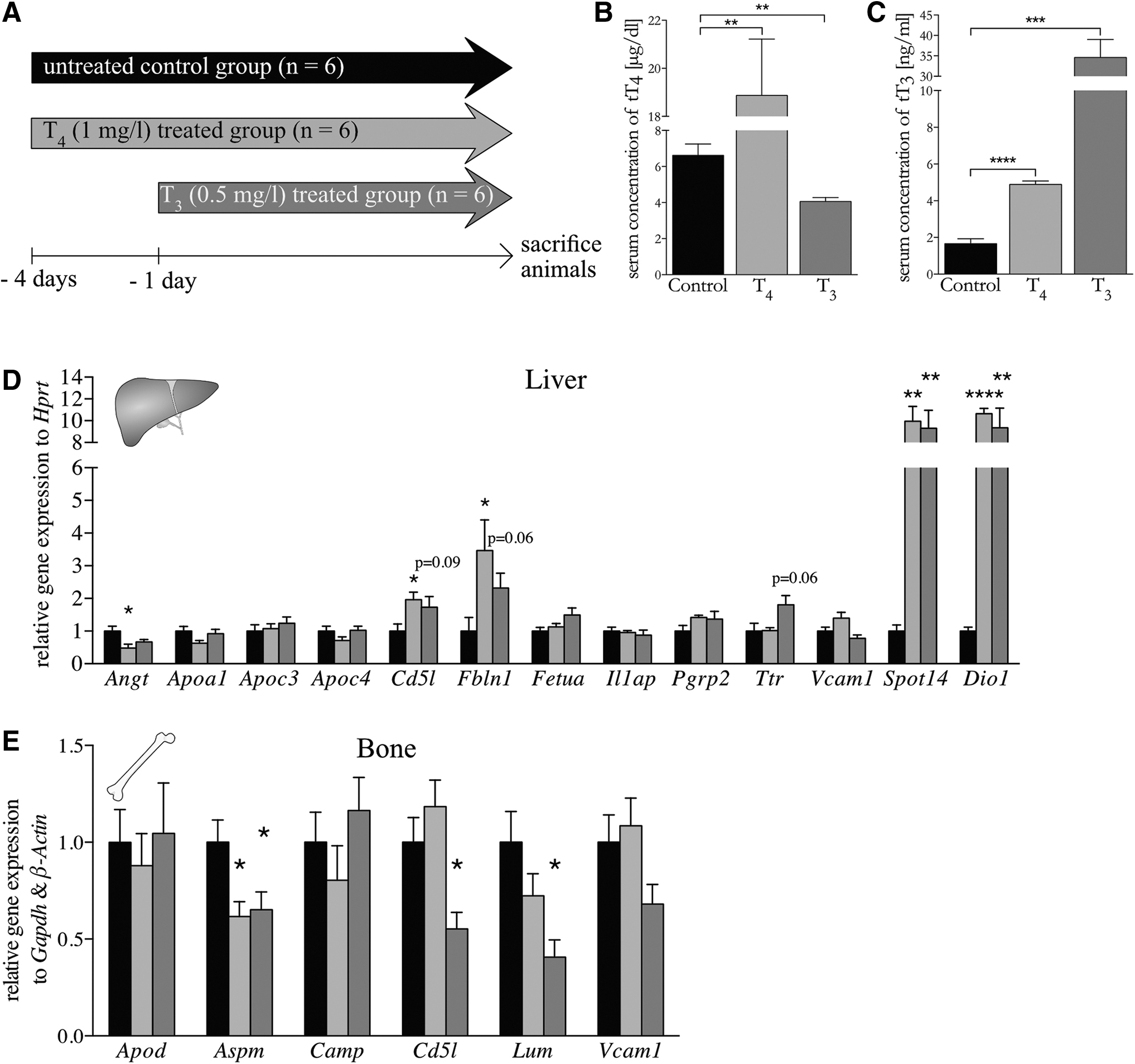
Validation of target gene expression under mild or severe thyrotoxic conditions for a short treatment period. (
Circulating CD5L abundance in hypo- and hyperthyroid subjects
The product of the most promising target gene CD5L was tested for serum abundance in hypo- and hyperthyroid subjects selected from a cross-sectional study of thyroid patients by using a commercially available polyclonal antibody. The relative CD5L abundance was significantly different between these two groups (Fig. 4A). In addition, basal TSH, fT4, and fT3 were measured in the serum of these patients and correlated with relative CD5L abundance. Relative CD5L abundance showed a significant inverse correlation with basal TSH (Fig. 4B), a correlation that did not reach statistical significance with fT4 (Fig. 4C), and a strong significant positive correlation with fT3 concentrations (Fig. 4D).
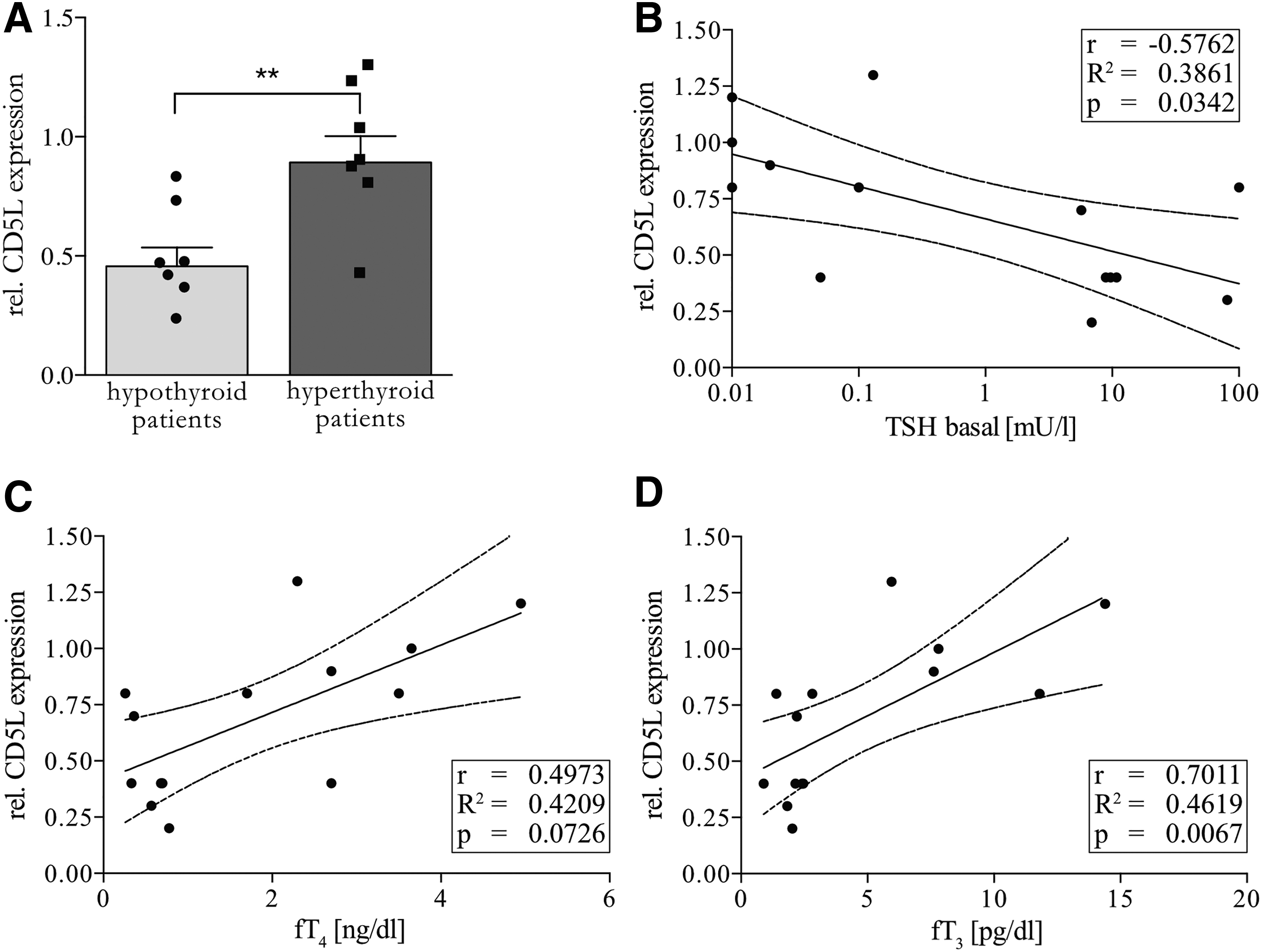
Relative CD5L abundance in serum of patients with acute hypo- or hyperthyroidism by Western blot. (
Target gene expression in TR mutants
To better characterize the underlying molecular mechanism and to identify TR isoforms involved in the regulation of these genes, TRα1+/m and TRβ−/− mutant mice with littermate controls were studied (38,39). TRα1+/m mice, which have normal levels of serum TH, displayed an increased hepatic expression of Angt, Apoa1, Fetua, Pgrp2, and Dio1 compared with wild-type littermates, while the endogenously hyperthyroid TRβ−/− mice had an increased expression of Camp and a decreased expression of Dio1, compared with heterozygous littermates (Fig. 5A). Remarkably, expression of Cd5l was not upregulated despite the elevated TH levels in TRβ−/− mice, suggesting that its induction by TH depends on this receptor isoform. In spleen, expression of Camp was increased and Apod expression was decreased in TRα1+/m (Fig. 5B). No changes in target gene expression in TRβ−/− mice were detectable, although there was a tendency of increased transcript levels of Apoa1 and Apoc3 (Fig. 5B). In bones of TRα1+/m mice, we observed an increased expression of Apod, Aspm, and Camp, while Lum mRNA levels were reduced compared with wild-type mice (Fig. 5C).
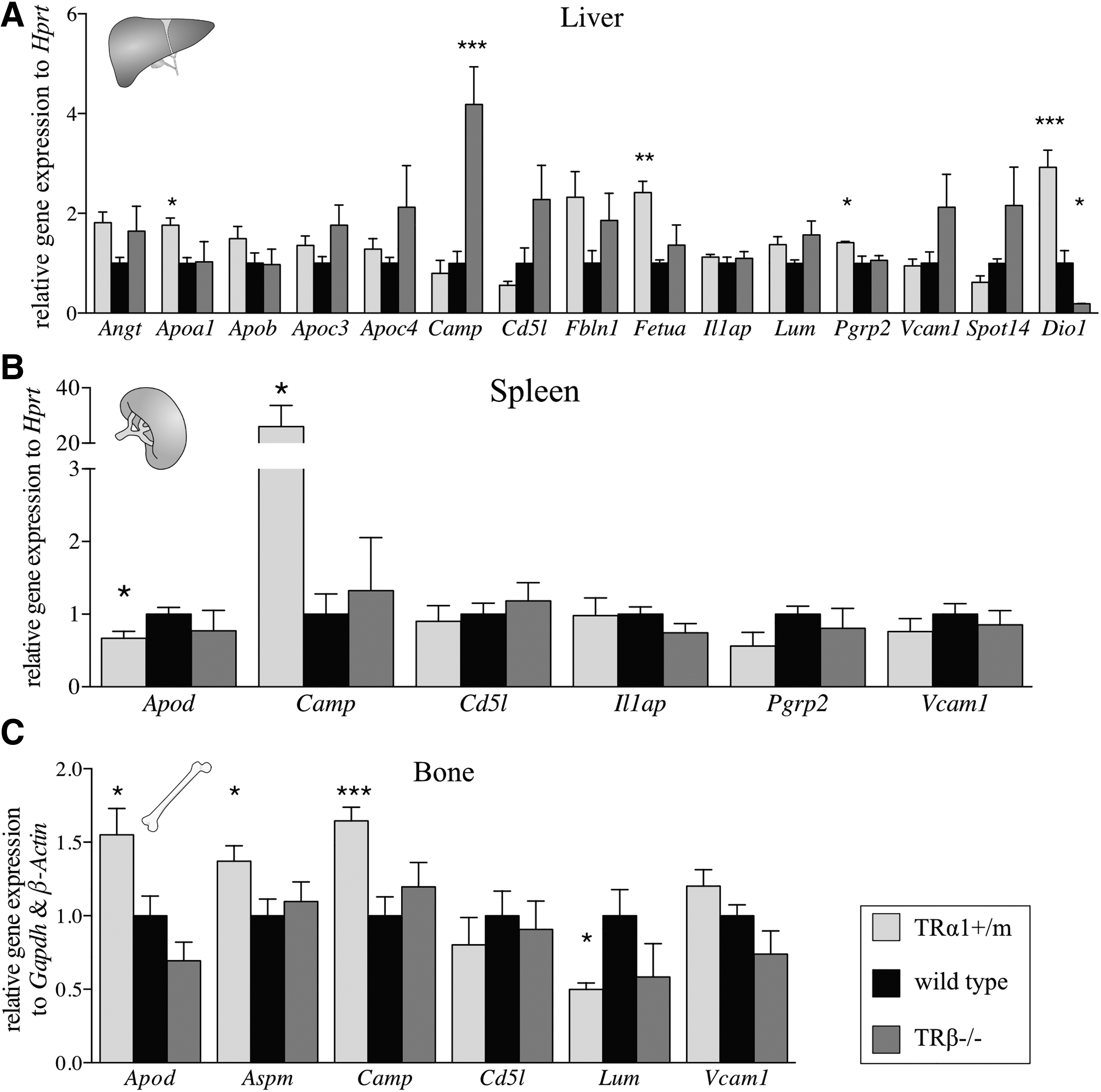
Validation of candidate gene expression in tissues of TRα1+/m and TRβ−/− mice compared with wild-type animals. (
Target gene expression in primary bone cells
As 6 of the 16 initially identified target genes were expressed in bone, including Cd5l and Lum, which were regulated under acute T3 treatment or in TR mutant mice, we used primary bone cell culture, differentiated osteoblasts, from mouse mesenchymal stem cells (Fig. 6A) or osteoclasts from mouse hematopoietic stem and progenitor cells (HSPCs) (Fig. 6B) to identify the cell type expressing the gene and to test for a direct cell-type specific regulation. The gene expression analysis of primary mouse osteoblasts revealed a reduction of Cd5l and Lum mRNAs in cells deficient for TRβ, while only a tendency for decreased Cd5l expression or increased Lum expression in cells carrying a mutation in TRα1 was detectable (Fig. 6C). These genes were also affected in primary mouse osteoclasts where Cd5l transcript amounts were increased in cells lacking TRβ, while those of Lum only showed a tendency toward increase in these cells (p = 0.16). Conversely, in primary osteoclasts from TRα1+/m mice, Cd5l expression was unchanged and Lum expression increased (Fig. 6D). Surprisingly, different TH concentrations did not induce any alterations in the expression of Cd5l and Lum (Fig. 6E), suggesting that those observed in the different animal models are rather the result of pre-existing endogenous dysregulations in these cell types caused by the genetic manipulations, and do not result from direct regulation via these TRs.
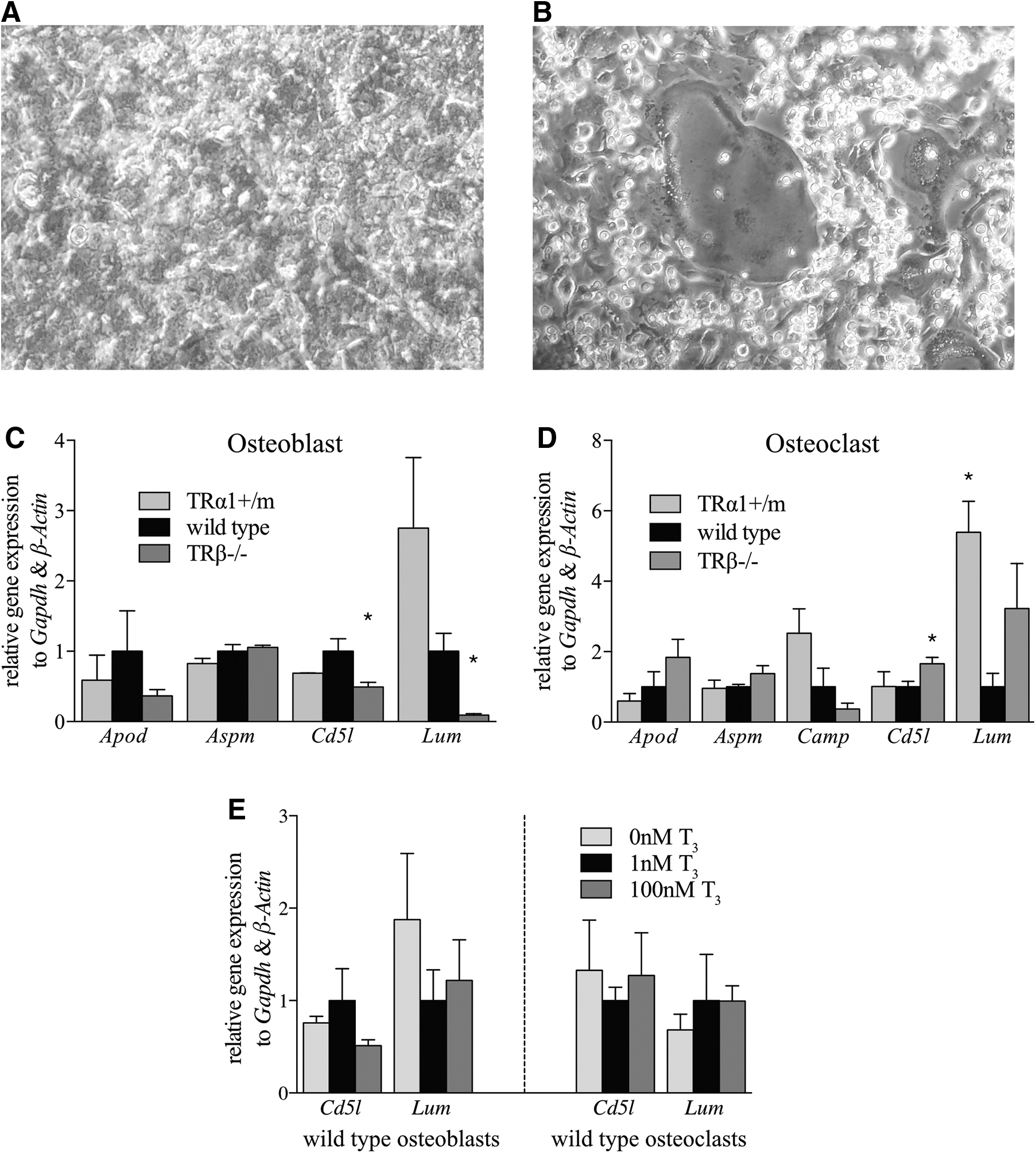
Determination of target gene expression in primary osteoblast or osteoclast culture. (
Cd5l expression in BMDMs
As the main source of CD5L are hepatic Kupffer cells and other tissue residing M1 macrophages (50,51), we hypothesized that the effect of TH on hepatic gene expression would be on liver-residing macrophages rather than epithelial cells, circulating immune cells, or hepatocytes, which did not express high levels of Cd5l as assessed by qPCR analysis in sorted cell populations (Supplementary Fig. S2C, D). Therefore, BMDMs were differentiated from HSPCs by treatment with M-CSF for six days, followed by an LPS+IFNγ-induced polarization for 24 hours into M1 BMDM (Fig. 7A). To test which TR isoform could be involved, HSPCs from TRα1+/m and TRβ−/− mice were also differentiated into M1 BMDM and compared with wild-type BMDM Cd5l expression. BMDM from animals carrying a mutation in Thra encoding TRα1 exhibited an 82% reduced expression of Cd5l, whereas BMDM lacking TRβ showed a similar expression as wild-type cells (Fig. 7B). Surprisingly, T3 itself did not affect the expression of Cd5l in wild-type BMDMs (Fig. 7C), suggesting that the changes in the TR mutant mice are likely caused by general alterations in BMDM formation capacity in vitro but do not result from direct regulation via TH. We, therefore, tested whether the BMDMs would be generally capable of inducing Cd5l expression by using liver X receptor (LXR) and retinoid X receptor (RXR) agonists, which are known transcriptional inducers of Cd5l (52 –54). Treating BMDM with LXR agonist T0901317 (T1317) and RXR agonist 9-cis retinoic acid (9cRA) resulted in increased Cd5l transcript levels in all groups independently of T3 in the culture medium, demonstrating that the BMDM of all animal models are capable of inducing Cd5l expression (Fig. 7D). We, therefore, hypothesized that the effect of T3 on Cd5l mRNA expression would be indirect through hepatocytes, which would be supported by the fact that the increase in gene expression in the intact animal depends on TRβ, although in macrophages TRα1 represents the predominating TR (55). To test whether a factor secreted by hepatocytes might regulate Cd5l expression, we exposed BMDMs to medium primed by T3-treated primary hepatocytes for 48 hours (Fig. 7E). To control for effects that could be caused by a general depletion of medium ingredients through the cellular metabolism, we used medium primed by T3-treated human hepatic stellate cell line LX-2 for 48 hours as a negative control. In contrast to the LX-2 medium, the hepatocyte primed medium was, indeed, capable of increasing Cd5l expression in mouse M1 BMDM in both TRα1+/m and wild-type mice, but not in TRβ−/− BMDMs (Fig. 7F)—an effect detectable across all tested TH concentrations (Supplementary Fig. S2E–G). The effect of medium primed by hepatocytes of TRβ−/− mice was comparable to that of wild-type mice (Supplementary Fig. S2H). Analyses of pro- and anti-inflammatory macrophage polarization markers did not reveal changes in M1 marker profile of BMDMs from TR mutant mice compared with wild-type mice, but M2 marker Il10 expression was increased in BMDMs from TRα1+/m mice, indicating a more M2-like polarization, although Argi1 expression was not affected (Fig. 7G). These findings demonstrate that a TRβ-dependent hepatocyte-derived factor is required to induce Cd5l expression in BMDMs. The positive regulation of Cd5l by TH, which can be detected in serum proteome and clinical samples, liver transcriptome and cell culture, therefore may provide an integrated readout of TH action on the liver-macrophage crosstalk.
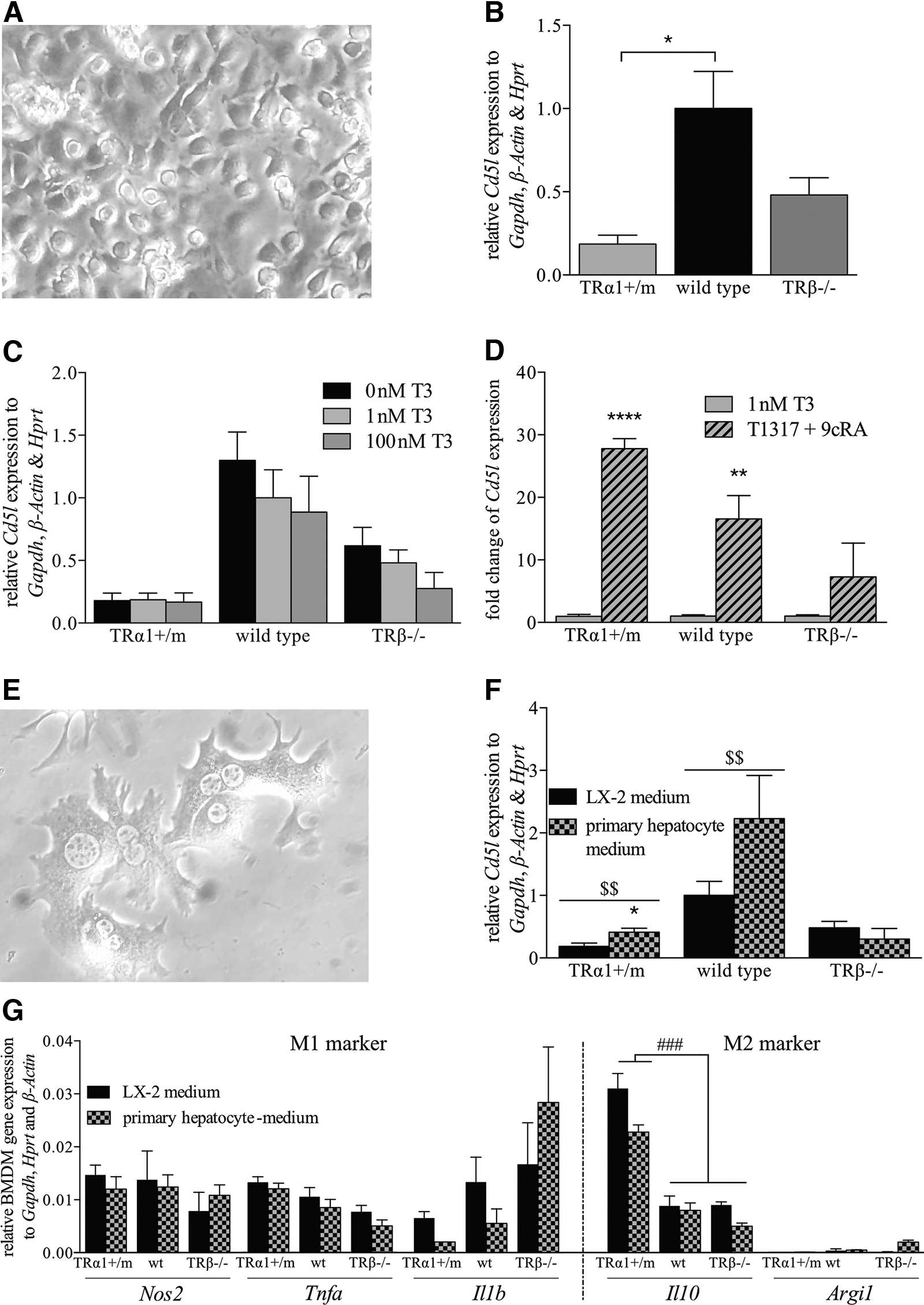
Determination of target gene expression in primary BMDM. (
Discussion
The identification of new specific biomarkers for TH status is a key issue for the development of improved diagnostics and personalized treatment strategies. Several approaches, including Omics techniques, have been used to generate further mechanistic insights into thyroid disorders and TH action (56,57); however, these techniques often lacked sensitivity and revealed inconsistent data across similar experiments depending on the study design used, and therefore always need independent validation (58,59). Here, we used human and mouse plasma proteome datasets obtained from experimentally induced thyrotoxicosis models to screen for robust candidates, which display correlating data across species and thus constitute valid targets for further studies. Of the 32 proteins identified, 16 exhibited concordantly altered abundance in human and mouse, whereas others such as the components of the complement (60 –62) and blood coagulation system (63) differed in both species and thus were excluded from further analysis. An in silico analysis using the Expression Atlas of the EMBL-EBI identified liver, spleen, and bone as the putatively dominant tissues for the remaining 16 target genes. All three tissues are well known to respond to TH. On a genome-wide level, between 2.8% and 13% of hepatic genes are regulated by T3 (57,58). Subsequent independent validation experiments could not confirm significant changes in liver, spleen, or bone expression of Angt, Apob, Apod, Camp, Il1ap, and Ttr, thus questioning their general validity as putative biomarkers of acute thyrotoxicosis. In contrast, hepatic Apoc3, Apoc4, Cd5l, Fetua, and Lum mRNA levels demonstrated a significant correlation with serum tT4 and tT3 concentrations. This was true as well for Cd5l, which again correlated significantly with serum tT3 and tT4 concentrations not only after long-term exposure but also in the short-term treatment paradigm. To test whether this correlation still exists on the protein level, the relative CD5L abundance in serum was determined and a significant correlation with basal TSH and fT3 in hypo- or hyperthyroid patients could be detected in a proof-of-concept study. However, there is still a need to verify and refine these findings in large study cohorts with different pathological conditions, which will also define the clinical limitations of CD5L as a putative TH biomarker.
As the increase of Cd5l gene expression between control and T4-treated animals was comparable after four days or two weeks of treatment, TH appears to have a rapid and prolonged effect. It argues against a desensitization effect in TRβ−/− mice to explain the negative effects despite approximately comparable serum TH concentrations as in T4-treated wild-type mice. Thus, we believe that the lack of Cd5l induction in TRβ−/− mice in liver or spleen is explained by a far more complex regulatory mechanism. Downregulation in primary mouse TRβ−/− osteoblasts and the upregulation in primary mouse TRβ−/− osteoclasts is not under direct control of T3, as different concentrations had no effect. Moreover, the in vivo altered bone structures in TRα1+/m and TRβ−/− mice (64) might also play a role in this context.
The correlation of Cd5l expression on acute thyrotoxic conditions with serum TH concentrations in various tissues suggested a circulating source, especially as its expression had been previously reported in Th17 cells (65,66). However, we did not observe Cd5l mRNA expression in isolated circulating monocytes, B cells, or T cells. Moreover, previous studies suggested that the main sources for circulating CD5L might be tissue-residing macrophages (67,68), suggesting that in the liver the expression would occur in Kupffer cells (51) rather than hepatocytes. This idea was confirmed by the almost complete absence of Cd5l expression in a macrophage-depleted liver cell suspension, with only residual expression remaining likely due to an incomplete removal of macrophages (69). This was corroborated by our findings of substantial Cd5l expression in primary mouse M1 BMDMs, which are an in vitro model highly similar to Kupffer cells despite being differentiated from different progenitor cells (70,71). Interestingly, Cd5l expression was reduced in these cells when isolated from TRα1+/m mutant mice, an effect not detectable in TRβ−/− mice, which concurs with the predominant role for TRα in macrophages (72). However, the upregulation of Cd5l expression by the LXR agonist T0901317 (T1317) and the RXR agonist 9cRA still occurred in BMDMs of TRα1+/m mice, suggesting that the response is not generally impaired in these cells. In contrast, BMDMs from TRβ−/− failed to respond to the LXR/RXR agonists, indicating a role of TRβ for correct BMDM differentiation and function. Most importantly, different concentrations of T3 did not affect Cd5l expression in BMDMs, demonstrating that the effect of TH on these cells is indirect. This was subsequently confirmed by the induction of Cd5l expression in BMDMs by primed medium from T3-treated hepatocytes. However, the TH dependency of this process was not replicated in the ex vivo system, suggesting that either additional factors, cell
The TRβ dependency of Cd5l induction is certainly of high relevance, as it will allow assessing hepatic/macrophage TRβ crosstalk under hyperthyroid conditions: In the case of intact TRβ signaling, elevated CD5L abundance and a correlation to serum TH levels would be expected, whereas the response would be blunted in TRβ-mediated resistance to TH.
Taken together, our study identified several novel TH target genes in the liver, which represent valuable tools to assess hepatic TH action on the transcriptome level in addition to Dio1 and Spot14, both of which are known to also respond to metabolic stimuli (73,74). In addition, with CD5L we identified an interesting protein with high serum abundance and a robust correlation with TH concentrations, which requires intact TRβ signaling. Future studies will, however, be required to test its usefulness in clinical practice, and especially whether other conditions such as hepatocellular carcinoma, liver cirrhosis, chronic liver disease, atherosclerosis, and some other mostly infectious or autoimmune disease may also influence the circulating CD5L levels. Nevertheless, even under those conditions, the responsiveness to TH through experimental induction of short-term thyrotoxicosis could be used to test for hepatic/macrophage TH resistance.
Footnotes
Acknowledgments
The authors gratefully thank the Gemeinsame Tierhaltung (GTH) of the University of Lübeck for housing and breeding of their animals and they also thank Marius Richter for sharing his knowledge about macrophage cell culture. They also like to thank Julia Resch, Maria Heier, and Anja Fischbach for excellent technical support and Sebastian Mehl for essential help in establishing the clinical cohort.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was generously supported by the German Research Foundation DFG as part of the priority program “SPP 1629—Thyroid Trans Act” (grant numbers Mi1242/5-1, Mi1242/4-1, HO5096/2-1, RA1923/6-1, and BR915/12-1) to J.M., C.S.H., M.R., and G.B., the Heisenberg Program Mi1242/2-2 to J.M., the GRK1957 “Adipocyte-Brain-Crosstalk,” and Research Unit FOR-2558 “TraceAge” (Scho 849/6-1) to J.H. and L.S.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Table S1