
Abstract
Background:
Development of adipose tissue before birth is essential for energy storage and thermoregulation in the neonate and for cardiometabolic health in later life. Thyroid hormones are important regulators of growth and maturation in fetal tissues. Offspring hypothyroid in utero are poorly adapted to regulate body temperature at birth and are at risk of becoming obese and insulin resistant in childhood. The mechanisms by which thyroid hormones regulate the growth and development of adipose tissue in the fetus, however, are unclear.
Methods:
This study examined the structure, transcriptome, and protein expression of perirenal adipose tissue (PAT) in a fetal sheep model of thyroid hormone deficiency during late gestation. Proportions of unilocular (UL) (white) and multilocular (ML) (brown) adipocytes, and UL adipocyte size, were assessed by histological and stereological techniques. Changes to the adipose transcriptome were investigated by RNA sequencing and bioinformatic analysis, and proteins of interest were quantified by Western blotting.
Results:
Hypothyroidism in utero resulted in elevated plasma insulin and leptin concentrations and overgrowth of PAT in the fetus, specifically due to hyperplasia and hypertrophy of UL adipocytes with no change in ML adipocyte mass. RNA sequencing and genomic analyses showed that thyroid deficiency affected 34% of the genes identified in fetal adipose tissue. Enriched Kyoto Encyclopedia of Genes and Genomes (KEGG) and Gene Ontology (GO) pathways were associated with adipogenic, metabolic, and thermoregulatory processes, insulin resistance, and a range of endocrine and adipocytokine signaling pathways. Adipose protein levels of signaling molecules, including phosphorylated S6-kinase (pS6K), glucose transporter isoform 4 (GLUT4), and peroxisome proliferator-activated receptor γ (PPARγ), were increased by fetal hypothyroidism. Fetal thyroid deficiency decreased uncoupling protein 1 (UCP1) protein and mRNA content, and UCP1 thermogenic capacity without any change in ML adipocyte mass.
Conclusions:
Growth and development of adipose tissue before birth is sensitive to thyroid hormone status in utero. Changes to the adipose transcriptome and phenotype observed in the hypothyroid fetus may have consequences for neonatal survival and the risk of obesity and metabolic dysfunction in later life.
Introduction
Congenital hypothyroidism (CH) affects ∼1 in 2000 neonates worldwide (1). At birth, affected infants are more likely to become hypothermic (1), and a small number of studies have reported that they are at greater risk of obesity, insulin resistance, and nonalcoholic fatty liver (NAFL) in childhood and young adult life, even when treated soon after diagnosis (2 –6). Low thyroid hormone status in the newborn is also commonly associated with prematurity and intrauterine growth restriction, conditions that have similar consequences for temperature control at birth and long-term metabolic health (7,8).
Development of adipose tissue before birth is crucial for energy storage, insulation, and thermogenesis in the neonatal period and for metabolic health in later life. In the fetus, adipose tissue comprises a mixture of both white and brown adipocyte types (9). White adipocytes, termed unilocular (UL), contain a large single lipid droplet and secrete a variety of adipokines, such as leptin, while brown adipocytes, termed multilocular (ML), are characterized by the presence of several smaller lipid droplets and an abundance of mitochondria with the capacity for nonshivering thermogenesis. Thermogenesis in ML adipocytes is enabled by the unique expression of uncoupling protein 1 (UCP1) on the inner mitochondrial membrane, which uncouples the electron transport chain to generate heat.
The sheep fetus is commonly used to study adipose tissue development and thyroid hormone biology before birth. In human and ovine fetuses, adipose tissue first appears around midgestation with similar anatomical locations in these and other large mammalian species (10). One of the largest adipose depots is located around the kidneys (perirenal adipose tissue, PAT), which is composed of a mixture of UL and ML adipocytes (10). Toward term, changes in the structure and function of fetal adipose tissue are observed in preparation for the nutritional and thermoregulatory challenges at birth (9). Differential gene expression profiles have been reported in ovine PAT perinatally as the structure of adipose tissue undergoes the transition from predominantly ML adipocytes at birth to UL adipocyte types at 2 weeks of postnatal life (9,11). In fetal sheep, leptin and UCP1 mRNA abundances in PAT increase near term, in association with rising concentrations of cortisol and triiodothyronine (T3) in the circulation (12,13).
Thyroid hormones, thyroxine (T4) and T3, have important roles in the control of growth, metabolism, and development of the fetus (7). In animal models, experimental hypothyroidism in utero modifies fetal growth and impairs the maturation and long-term functioning of organs, such as the heart, brain, and adipose tissue (7). Thyroid deficiency in fetal sheep impairs adipose thermogenic capacity and the ability to maintain body temperature at birth (14). In addition, maternal hypothyroidism during pregnancy increases visceral fat mass and causes glucose intolerance in adult rat offspring (15,16).
Intrauterine programming of adiposity by thyroid hormones has implications for adult insulin sensitivity and metabolic disease (17). Cross talk between adipose tissue, especially PAT (18), liver, and skeletal muscle may be mediated by adipose-derived factors, such as free fatty acids and adipocytokines. In adult humans, PAT thickness has been correlated with increased risk of conditions, including hypertension, fatty liver, and coronary heart disease (18). The molecular mechanisms by which thyroid hormones regulate adipogenesis and adipose function during fetal life, with implications for offspring health in the short and longer term, are, however, poorly understood.
The aims of the current study were to determine the effects of hypothyroidism in utero on the growth and development of ovine adipose tissue and to determine the molecular mechanisms responsible using transcriptome profiling. We tested the hypothesis that thyroid hormone deficiency during late gestation would increase and decrease the amounts of UL and ML adipocytes, respectively, and impair adipose thermogenic capacity near term.
Materials and Methods
Animals
All surgical and experimental procedures were carried out in accordance with UK Home Office legislation and the Animals (Scientific Procedures) Act 1986, after ethical approval by the University of Cambridge Animal Welfare and Ethical Review Body at the Department of Physiology, Development and Neuroscience, University of Cambridge, UK. Nineteen Welsh Mountain pregnant ewes of known gestational age and carrying twin fetuses (15 female and 23 male) were used in this study. The ewes were housed in individual pens and were maintained on 200 g/day concentrates with hay and water ad libitum and access to a salt block. Food, but not water, was withheld from the ewes for 18–24 hours before surgery.
Experimental procedures
At 105–110 days of gestation (dGA; term ∼145 ± 2 days) and under halothane anesthesia (1.5% halothane in O2-N2O), the twin fetuses of each ewe underwent either a thyroidectomy (TX) or a sham operation in which the thyroid gland was exposed but not removed (sham), as described previously (19,20). At either 129 (n = 18) or 143 dGA (n = 20), the fetuses were delivered by cesarean section under general anesthesia (20 mg/kg maternal body weight sodium pentobarbitone intravenously [i.v.]). Blood samples were collected by venipuncture of the umbilical artery into ethylenediaminetetraacetic acid (EDTA)-containing tubes. Each fetus was weighed and a variety of fetal organs, including the PAT, were collected after the administration of a lethal dose of barbiturate (200 mg/kg sodium pentobarbitone i.v.).
Plasma hormone measurements
Umbilical plasma T3 and T4 concentrations were determined by radioimmunoassay (RIA; MP Biomedicals, Loughborough, UK); the intra-assay coefficients of variation were 3% and 5%, and the minimum levels of detection were 0.14 and 7.0 ng/mL, respectively. Plasma insulin and cortisol concentrations were determined using enzyme-linked immunosorbent assay (ELISA) kits (insulin: Mercodia, Uppsala, Sweden; cortisol: IBL International, Hamburg, Germany); the intra-assay coefficients of variation were both 9%, and the minimum levels of detection were 0.025 and 2.5 ng/mL, respectively. Plasma concentrations of leptin, like growth factor (IGF)-I, and IGF-II were determined by RIA as previously described (21,22). The intra-assay coefficients of variation were 4–5%, and the minimum levels of detection were 0.09, 0.08, and 4.0 ng/mL, respectively.
Adipose tissue histology
Fetal PAT was fixed in 4% paraformaldehyde (with 0.2% glutaraldehyde in 0.1 M phosphate buffer, pH 7.4) and embedded in paraffin wax. Each block of PAT was cut into 7 μm sections and stained with hematoxylin and eosin. Sections were scanned using a NanoZoomer digital slide scanner (Hamamatsu Photonics, Welwyn, UK) to create digital images for analysis. All stereological measurements were performed and analyzed blind to the treatment group. The percentage volumes of UL and ML adipocytes were determined using NewCAST stereological software (Visiopharm, Hoersholm, Denmark). A point-counting grid of 25 points was applied over the adipose sections, and meander sampling was used to analyze the adipocyte types. A total of 40 counting frames were used per slide to provide at least 200 points per animal. UL cells were defined as an adipocyte with a diameter larger than 60 μm, after tissue shrinkage.
UL cell size was determined by measuring the perimeter of 60–80 of the largest UL adipocytes using the stereology software
RNA sequencing and bioinformatic analysis
Total RNA was extracted from fetal PAT samples using the RNeasy Lipid Tissue Mini Kit (Qiagen, Manchester, UK), and cDNA libraries were prepared in samples with RNA integrity number >6 (Agilent Bioanalyzer 2100 system; Agilent Technologies TDA UK Limited, Stockport, UK). Briefly, mRNA was enriched from total RNA before reverse transcription, and adenylation and barcode ligation were performed after the synthesis of double-stranded cDNA. Ligated libraries were enriched with a limited amplification. Indexed libraries were normalized, pooled, and sequenced on the Illumina HiSeq 4000 platform with single-end reads (SE50) at the Genomics Core Facility, Cancer Research UK Cambridge Institute (Cambridge, UK).
For each library, original reads files were quantified, trimmed, and aligned to the Ovis aries (sheep) genome assembly Oar_v3.1 from the International Sheep Genome Consortium using ClusterFlow pipeline tool (version v0.5 dev, fastqc_star pipeline) (24), including the following software: fastqc (version 0.11.5) (25), trim_galore (version 0.4.2) (26), fastq_screen (version 0.9.3) (27), MultiQC (version 0.9dev) (28), and read alignment software STAR (version 2.5.1b_modified) (29). Further details can be found in the data report (
As the nucleotide sequences for the thyroid hormone receptor (TR) isoforms, α1 and α2, were not available for sheep, highly similar porcine sequences were used to map their genomic positions in the sheep genome using BLAT (32). From the BLAT results, a bed12 file was created to isolate the exon positions for the TRα1 and TRα2 isoforms. Individual RNA-seq alignment files from each of the treatment groups were merged and loaded into Integrative Genomics Viewer (IGV, genome Oar_v3.1).
Initial quality control included principal component analysis (PCA), and data from two fetuses were removed as outliers before further analysis. Differentially expressed genes were identified using R (version 3.5.3) DESeq2 package (version 1.22.2) (33), using variance stabilizing transformed expression for counts. Genes with more than one read across all samples within a contrast were retained. Additional filtering of genes with low mean read counts was automatically applied by DESeq2. For each contrast, differentially expressed genes with Benjamini–Hochberg (BH) adjusted p-values <0.05 were identified. Log2 fold change in gene expression was plotted against the mean of read counts normalized by library size for each gene in MA plots.
Significant differentially expressed genes from each comparison, within and between treatment and gestational age groups, were plotted in volcano plots and a summary of the numbers of genes in the intersections of the comparisons were plotted using UpSetR (version 1.4.0). For heatmap analysis, gene-level transcripts expression values were derived by normalized transformed values estimated by DESeq2 for each sample.
A Bayesian method (lfcshrink) implemented in DESeq2 was used to moderate the log2 fold changes obtained for genes with low or variable expression levels. Upregulated and downregulated genes in different contrasts (BH-adjusted p < 0.01 and absolute log2 fold change >1) were analyzed for Gene Ontology (GO) term enrichment. Gene sets were analyzed for overrepresentation of biological process (BP) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway using R package clusterProfiler (version 3.10.1). Significantly enriched terms were identified by applying the default clusterProfiler algorithm coupled with Fisher's exact test statistic (p ≤ 0.05, q ≤ 0.05). GO plots were drawn using R package ggplot2 (version 3.2.1). Normalized read counts were used in the statistical analysis of mRNA abundance of key genes. The raw sequencing data and data reports are deposited at ArrayExpress with experimental code E-MTAB-8396. Expression-count data are available online (
Western blotting
Frozen samples of fetal PAT were homogenized in cold lysis buffer (100 mg/mL; 20 mM sodium orthovanadate, 10 mM β-glycerol phosphate, 50 mM sodium fluoride, and protease inhibitor cocktail [Roche, Burgess Hill, UK]) in Lysing Matrix-D tubes using a Super FastPrep 1 homogenizer (MP Biomedicals). Samples were centrifuged at 15,000g for 10 minutes at 4°C. Extracted protein concentration was measured by a bicinchoninic acid protein assay (Sigma, Poole, UK). Before loading, samples were mixed with NuPage 4 × lithium dodecyl sulphate (LDS) loading buffer (2% LDS, 141 mM Tris base, 10% glycerol, 0.51 mM EDTA, 0.22 mM Blue G, 0.175 mM Phenol Red; Life Technologies, Loughborough, UK) and 100 mM DL-dithiothreitol and heated to 70°C for 10 minutes (with the exception of those for phosphorylated S6-kinase [pS6K] quantification, which were heated to 99°C for 5 minutes).
Equal amounts (100 μg) of sample protein were separated using 7.5% Mini-PROTEAN pre-cast gels (Bio-Rad, Hemel Hempstead, UK) for 50 minutes at 150 V and transferred for 10 minutes at 11 V onto a polyvinylidene difluoride membrane (Immobilon P 0.45 μm; Millipore, Sigma) using the Pierce G2 Fast Blotter (Thermo Scientific, Loughborough, UK). The membrane was incubated with 2.5% nonfat milk (or bovine serum albumin for phosphorylated proteins) in Tris-buffered saline with 0.1% Tween-20 for 1 hour at room temperature, followed by incubation overnight at 4°C with primary antibodies: rabbit polyclonal anti-InsRβ (10 μg/mL; Santa Cruz Biotechnologies, Heidelberg, Germany), rabbit polyclonal anti-IGF-1Rβ (10 μg/mL; Santa Cruz Biotechnologies), rabbit polyclonal anti-leptin receptor (1 μg/mL; Biorbyt, Cambridge, UK), rabbit polyclonal anti-pAkt (1:800, Ser473; Cell Signaling Technology, Hitchin, UK), mouse monoclonal anti-Akt1 (1:1000; Cell Signaling Technology), rabbit monoclonal anti-Akt2 (1:1000; Cell Signaling Technology), rabbit polyclonal anti-pmTOR (1:800, Ser 2448; Cell Signaling Technology), rabbit polyclonal anti-pS6K (1:1000, Thr 389; Cell Signaling Technology), rabbit polyclonal anti-GLUT4 (2.5 μg/mL; Abcam, Cambridge, UK), mouse monoclonal anti-PCNA (2 mg/L; Dako, Cambridge, UK), rabbit polyclonal anti-PPARγ (4 μg/mL; Biorbyt), and rabbit polyclonal anti-UCP1 (1:500; Abcam).
Each membrane was incubated with a horseradish peroxidase-conjugated anti-rabbit or anti-mouse secondary antibody (GE Healthcare, Amersham, UK) for 1 hour at room temperature. Protein expression was visualized by addition of Clarity Western ECL chemiluminescence substrate (Bio-Rad) and quantified using Image Lab software (ChemiDoc; Bio-Rad) after normalization to Ponceau S staining (34). All data were normalized to a quality control sample across all gels and expressed as fold changes, relative to the sham group at 129 dGA, in arbitrary units.
Citrate synthase activity
Citrate synthase (CS) activity was measured in homogenized PAT samples by a spectrophotometric enzyme assay. The assay buffer (pH 8) contained 0.1 mM 5,5′-dithio-bis-2-nitrobenzoic acid, 1 mM oxaloacetate, and 0.3 mM acetyl-CoA. Adipose CS activity was determined from the maximum rate of change of absorbance at 412 nm and 37°C (rate of thionitrobenzoic acid production) over 3-minute periods and was expressed as μmoles per minute per mg protein, measured by a bicinchoninic acid protein assay.
Statistical methods
Data were analyzed by three-way analysis of variance (ANOVA) with treatment, gestational age, and sex of the fetus as factors (SigmaStat 3.5; Systat Software, San Jose, CA). The sex of the fetus had no significant effect on any of the variables measured; data from male and female fetuses were, therefore, combined and analyzed by two-way ANOVA followed by the Tukey post hoc test. Relationships between variables were assessed by Pearson's correlation. Significance was regarded as p < 0.05.
Results
Hypothyroidism in utero increases circulating insulin and leptin concentrations
In TX fetuses, plasma T4 and T3 concentrations decreased to below the limits of assay detection, while plasma insulin and leptin were increased to levels above those in sham fetuses at both 129 and 143 dGA (p < 0.05, Table 1). The normal developmental rise in plasma cortisol concentration over this time period was observed in both groups of fetuses (p < 0.05, Table 1); fetal hypothyroidism tended to suppress plasma cortisol concentration (p = 0.052). Plasma T3 increased near term in the sham fetuses (p < 0.05, Table 1), while plasma concentrations of IGF-I and IGF-II were unaffected by TX or gestational age (Table 1).
Mean (±Standard Error of the Mean) Plasma Hormone Concentrations, Body and Perirenal Adipose Tissue Weights, and Adipose Citrate Synthase Activity in Sham and Thyroidectomy Sheep Fetuses at 129 and 143 Days of Gestation
Significantly different from sham fetuses at the same gestational age; +significantly different from fetuses in the same treatment group at 129 dGA; two-way ANOVA, p < 0.05. ND, not detectable (minimum levels of detection: T4 7.0 ng/mL, T3 0.14 ng/mL).
ANOVA, analysis of variance; dGA, days of gestation; IGF, like growth factor; PAT, perirenal adipose tissue; T3, triiodothyronine; T4, thyroxine; TX, thyroidectomy.
UL-specific adipocyte growth and proliferation enlarge adipose tissue mass in hypothyroid fetuses
In TX fetuses, absolute PAT weight was greater than in sham fetuses at 143 dGA (p < 0.05), but not 129 dGA (Table 1); the PAT mass relative to body weight was higher in TX fetuses than in sham fetuses at both ages (p < 0.05, Table 1). When expressed as a percentage of total PAT volume, sham fetuses had a greater percentage of ML relative to UL adipocyte types at 129 and 143 dGA (p < 0.001, Fig. 1A). In TX fetuses, there was an increase in the percentage of UL, and a decrease in ML, adipocytes compared with control values at both ages (p < 0.001, Fig. 1A). When the percentage volumes of ML and UL adipocytes were expressed as absolute and relative masses, a 0.95- to 1.30-fold increase in UL adipocyte mass was observed in the TX fetuses at both 129 and 143 dGA (p < 0.05, Fig. 1B). The absolute and relative ML adipocyte masses, and fetal body weight, were unaffected by TX (Table 1, Fig. 1B). Positive correlations were observed between the relative UL adipocyte mass and fetal concentrations of insulin (r = 0.49, n = 37, p < 0.005) and leptin (r = 0.68, n = 38, p < 0.001). The average perimeter of the largest UL adipocytes increased with hypothyroidism (p < 0.05) and gestational age (p < 0.001, Fig. 1C). These data indicate that the increase in PAT mass observed after TX was due to increased UL-specific adipocyte growth and proliferation (Fig. 1D, E).

Mean (±SEM) measurements of
Adipose transcriptome analysis reveals differential gene expression profiles in response to hypothyroidism in utero
RNA sequencing was undertaken on PAT samples from sham and TX fetuses at both 129 and 143 dGA and the distribution of gene expression was assessed initially by unbiased PCA. Using the top 500 most variable genes, PCA showed distinct clustering of data based on treatment group (sham and TX) and common transcriptional profiles between gestational age within treatment groups (Supplementary Fig. S1A). Genes associated with the effect of TX, defining the principal component 1 (PC1), explained 43.5% of the variance. These included LPL, ELOVL6, PLIN1, FBP2, and ADIPOQ (Supplementary Fig. S1B). Genes associated with the effect of gestational age within each treatment group (principal component 2, PC2) explained 14.0% of the variance and included UCP1, DIO1, FABP3, and ADRA1A (Supplementary Fig. S1C). A hierarchical clustering heatmap using 272 differentially expressed genes with an absolute log2 fold change threshold of 2 and p-adjusted <0.05 confirmed that the transcriptome data from the sham and TX groups clustered apart (Supplementary Fig. S1D).
In total, 17,622 genes were identified in the adipose samples from the annotated sheep genome. Of these, 5999 genes were differentially expressed between the sham and TX groups (34.0%, p-adjusted <0.05). When data from all animals were considered with an absolute log2 fold change ≥1, a total of 1472 genes were affected by hypothyroidism (768 upregulated and 704 downregulated by TX; Supplementary Fig. S2A) and 409 were affected by increasing gestational age (180 upregulated, 229 downregulated with increased gestational age; Supplementary Fig. S2B).
When the data were analyzed by age in each treatment group, the expression of 609 genes changed between 129 and 143 dGA in the sham fetuses (232 upregulated, 377 downregulated; Supplementary Fig. S2C), while this number was reduced to 174 in the TX fetuses over the same time period (86 upregulated, 88 downregulated; Supplementary Fig. S2D). When the data were analyzed by treatment at each age, TX influenced the expression of more genes at 143 dGA (1576 total, 869 upregulated, 707 downregulated; Supplementary Fig. S2F) than at 129 dGA (1090 total, 625 upregulated, 465 downregulated; Supplementary Fig. S2E).
Transcriptome profiles were compared between and within treatment and gestational age groups, and the results are summarized in the UpSet plot (Supplementary Fig. S3A). Differentially expressed genes unique to each of the comparisons were also plotted (Supplementary Fig. S3B).
GO and KEGG pathway analyses identify adipogenic, metabolic, thermogenic, and hormone signaling processes influenced by hypothyroidism in utero
A number of biological pathways were identified as enriched in PAT from TX fetuses. Of particular relevance, enriched KEGG pathways were associated with regulation of lipolysis; fatty acid synthesis and metabolism; insulin resistance; AMPK, FoxO, and cAMP signaling; and signaling pathways for insulin, peroxisome proliferator-activated receptor (PPAR), and adipocytokines (Fig. 2A and Supplementary Fig. S4). Biological process pathways overrepresented in the GO analysis included fatty acid metabolism and biosynthesis, and several aspects of thermogenesis and temperature regulation (Fig. 2B and Supplementary Fig. S5). When the data were assessed by treatment and age, an additional number of enriched pathways were identified in TX fetuses at 129 dGA including apelin and thyroid hormone signaling pathways (Supplementary Fig. S4) and lipid metabolism (Supplementary Fig. S5).
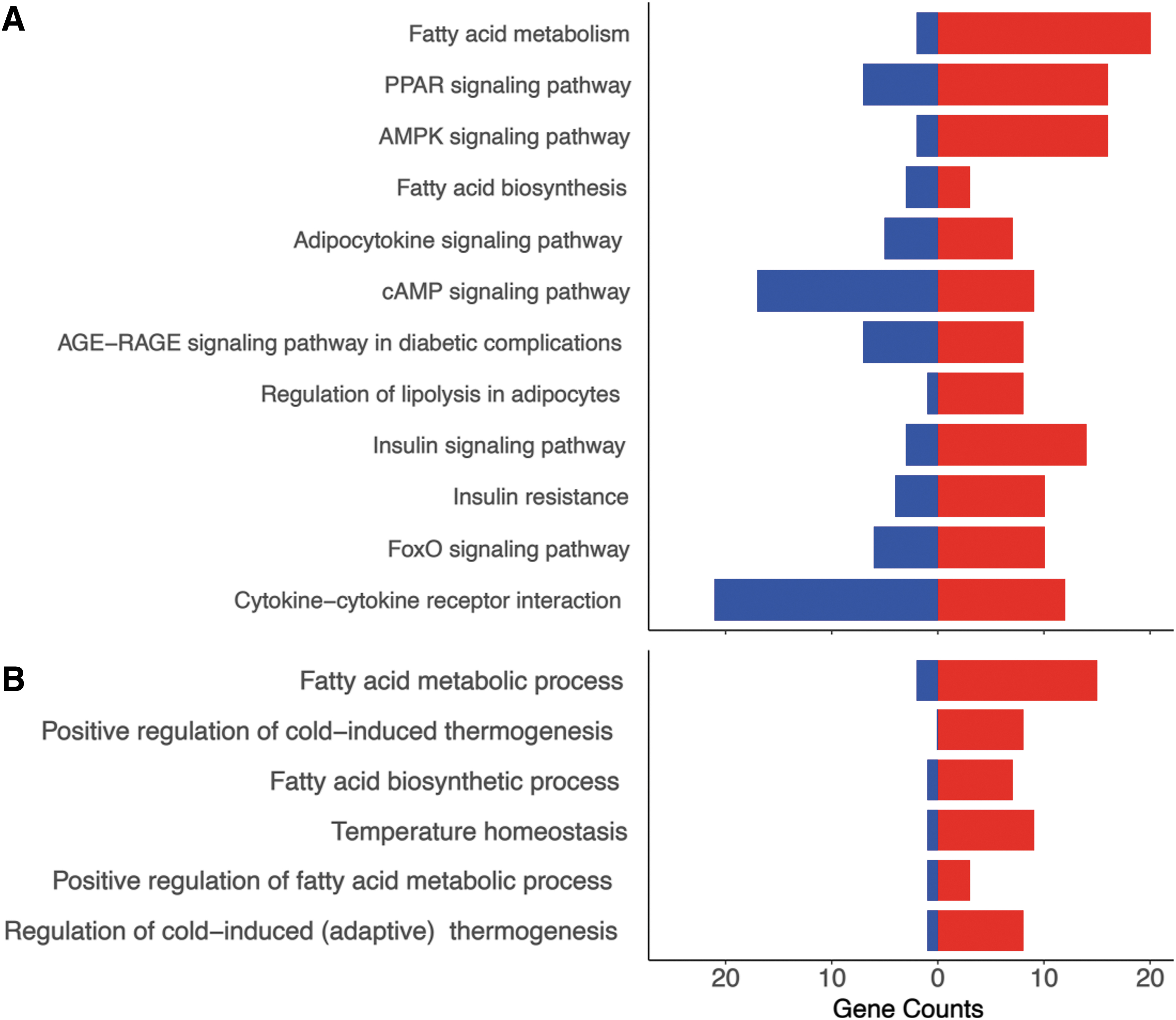
KEGG pathway and BP bar plots using RNA sequencing data from PAT taken from sham and thyroidectomized (TX) fetuses at 129 and 143 dGA. Selected relevant KEGG
Hypothyroidism in utero activates adipose PPAR and insulin-IGF signaling
Key genes in some of the enriched pathways were examined in more detail, and protein content was quantified by Western blotting. Increased mRNA and protein contents of the mitotic marker, proliferating cell nuclear antigen (PCNA), and PPARγ, an important regulator of adipocyte differentiation, were observed in response to fetal hypothyroidism (p < 0.005, Supplementary Fig. S6A, B). Adipose PCNA mRNA abundance decreased between 129 and 143 dGA in sham fetuses (p < 0.05), and PCNA mRNA and protein contents were higher in TX fetuses compared with sham fetuses at 143 dGA (p < 0.05, Supplementary Fig. S6A). Compared with control values, both the mRNA and protein contents of PPARγ at 129 dGA, and mRNA abundance at 143 dGA, were greater in TX fetuses (p < 0.05, Supplementary Fig. S6B). Adipose mRNA abundances of IGF-I and IGF-II and leptin were also increased by TX (p < 0.001, Supplementary Fig. S7A–C). An increase in adipose IGF-I mRNA was observed between 129 and 143 dGA in TX fetuses (p < 0.05, Supplementary Fig. S7A); over the same period, IGF-II mRNA abundance decreased in both sham and TX fetuses (p < 0.05, Supplementary Fig. S7B).
To examine if hyperinsulinemia and increased adipose IGF mRNA abundance observed in TX fetuses were responsible, at least in part, for the greater PAT mass, the expression of insulin-IGF and adipokine signaling pathways was investigated in sham and TX fetuses. At 129 dGA, the mRNA abundance of the insulin receptor was higher, while insulin receptor β-subunit (InsRβ) protein content was lower in TX fetuses compared with sham fetuses (p < 0.05, Supplementary Fig. S6C). Between 129 and 143 dGA, a reduction in InsRβ protein was seen in sham fetuses, but not in TX fetuses (p < 0.05, Supplementary Fig. S6Cii). At 129 dGA, protein kinase β 1 (Akt1) and Akt2 mRNA, and Akt1 protein content, were greater in TX fetuses compared with sham fetuses (p < 0.05, Supplementary Fig. S6D, E). In the TX fetuses, Akt2 mRNA abundance decreased between 129 and 143 dGA, and Akt2 protein content was lower at 143 dGA compared with that observed in sham fetuses (p < 0.05, Supplementary Fig. S6E). The total amount of phosphorylated Akt (pAkt) protein did not change with age or fetal hypothyroidism (data not shown).
A developmental rise in mRNA abundance of the mammalian target of rapamycin (mTOR) was observed in sham fetuses, but not in TX fetuses (p < 0.05, Supplementary Fig. S6Fi); at 143 dGA only, mTOR mRNA abundance was lower in TX fetuses than in sham fetuses (p < 0.05, Supplementary Fig. S6Fi). Fetal hypothyroidism reduced phosphorylated mTOR protein content (p < 0.05), although post hoc analysis failed to identify significant differences at either age (Supplementary Fig. S6Fii). At 129 dGA, pS6K protein content was higher in TX fetuses than in sham fetuses (p < 0.05) and decreased with increasing age in TX fetuses, but not in sham fetuses (p < 0.05, Supplementary Fig. S6Gii); S6K mRNA abundance, however, was unaffected by fetal TX and age (Supplementary Fig. S6Gi).
In TX fetuses, mRNA abundance of the insulin-sensitive glucose transporter, glucose transporter isoform 4 (GLUT4), was greater than that observed in sham fetuses at 129 dGA and decreased toward term, unlike sham fetuses (p < 0.05, Supplementary Fig. S6Hi). Adipose GLUT4 protein content was also greater in TX fetuses than in sham fetuses at both ages (p < 0.05, Supplementary Fig. S6Hii).
Developmental increments in adipose adrenergic receptor (ADR) α1A mRNA abundance were observed between 129 and 143 dGA in both sham and TX fetuses, without any effect of TX (p < 0.05, Supplementary Fig. S7D). Adipose ADRβ2 mRNA abundance also increased toward term in sham fetuses, but not in TX fetuses; ADRβ2 mRNA abundance was lower in TX fetuses compared with sham fetuses at 143 dGA (p < 0.05, Supplementary Fig. S7H). At 129 dGA, the mRNA abundance of ADRα1D was lower, while that of ADRβ1 and ADRβ3 were all higher, in TX fetuses than in sham fetuses (p < 0.05, Supplementary Fig. S7E, G, I). Adipose ADRα2A was increased by TX at both 129 and 143 dGA (p < 0.05, Supplementary Fig. S7F). There were no effects of TX or age on the amount of the long-form leptin receptor protein, ADRα1B mRNA abundance, or the mRNA or protein abundance of the IGF type 1 receptor in PAT (data not shown).
Hypothyroidism in utero impairs adipose thermogenic capacity
Although the absolute and relative masses of ML adipocytes were unaffected by TX, the capacity for nonshivering thermogenesis was impaired in the PAT of TX fetuses. Adipose CS activity, as a proxy measure of mitochondrial density, increased between 129 and 143 dGA in sham fetuses, but not in TX fetuses (p < 0.05, Table 1); CS activity in the TX fetuses was lower than control values at both ages (p < 0.05, Table 1). When observations from all fetuses were considered, regardless of gestational age or treatment group, a positive relationship was observed between adipose CS activity and the percentage volume of ML adipose tissue (r = 0.53, n = 37, p < 0.001).
Both adipose UCP1 mRNA and protein content (both absolute values and when expressed relative to CS activity or percentage volume of ML adipose tissue) increased near term in sham fetuses, but these developmental changes were abolished by TX (Fig. 3). Absolute and relative UCP1 mRNA abundance was lower in TX fetuses compared with sham fetuses at 143 dGA (p < 0.05, Fig. 3A, B); relative UCP1 protein content was reduced by TX at both 129 and 143 dGA (p < 0.05, Fig. 3C, D) in line with previous findings at term (7).

Mean (±SEM) abundance of
Hypothyroidism in utero alters adipose thyroid hormone metabolism and signaling
Adaptive changes in adipose mRNA abundance for the iodothyronine deiodinases (DIO) and TR were observed in response to TX. Toward term, significant increments in the mRNA abundance of DIO1 and DIO2, which both metabolize T4 to the biologically active T3, were observed in sham fetuses but not in TX fetuses (p < 0.05, Fig. 4A, B). In TX fetuses, lower DIO1 mRNA level at 143 dGA, and higher DIO2 mRNA level at 129 dGA, were observed compared with sham fetuses (p < 0.05, Fig. 4A, B). Adipose mRNA levels of TRα1, TRα2, and TRβ were increased by TX at both ages (p < 0.05, Fig. 4C–E); a significant increase in adipose TRβ mRNA was observed in sham fetuses near term (p < 0.05, Fig. 4E). A Sashimi plot was constructed to show the expression of the splice variants TRα1 and TRα2 (Supplementary Fig. S8), and adipose TRα1 and α2 mRNA levels were highly correlated when data from all animals were combined (r = 0.99, n = 35, p < 0.0001).
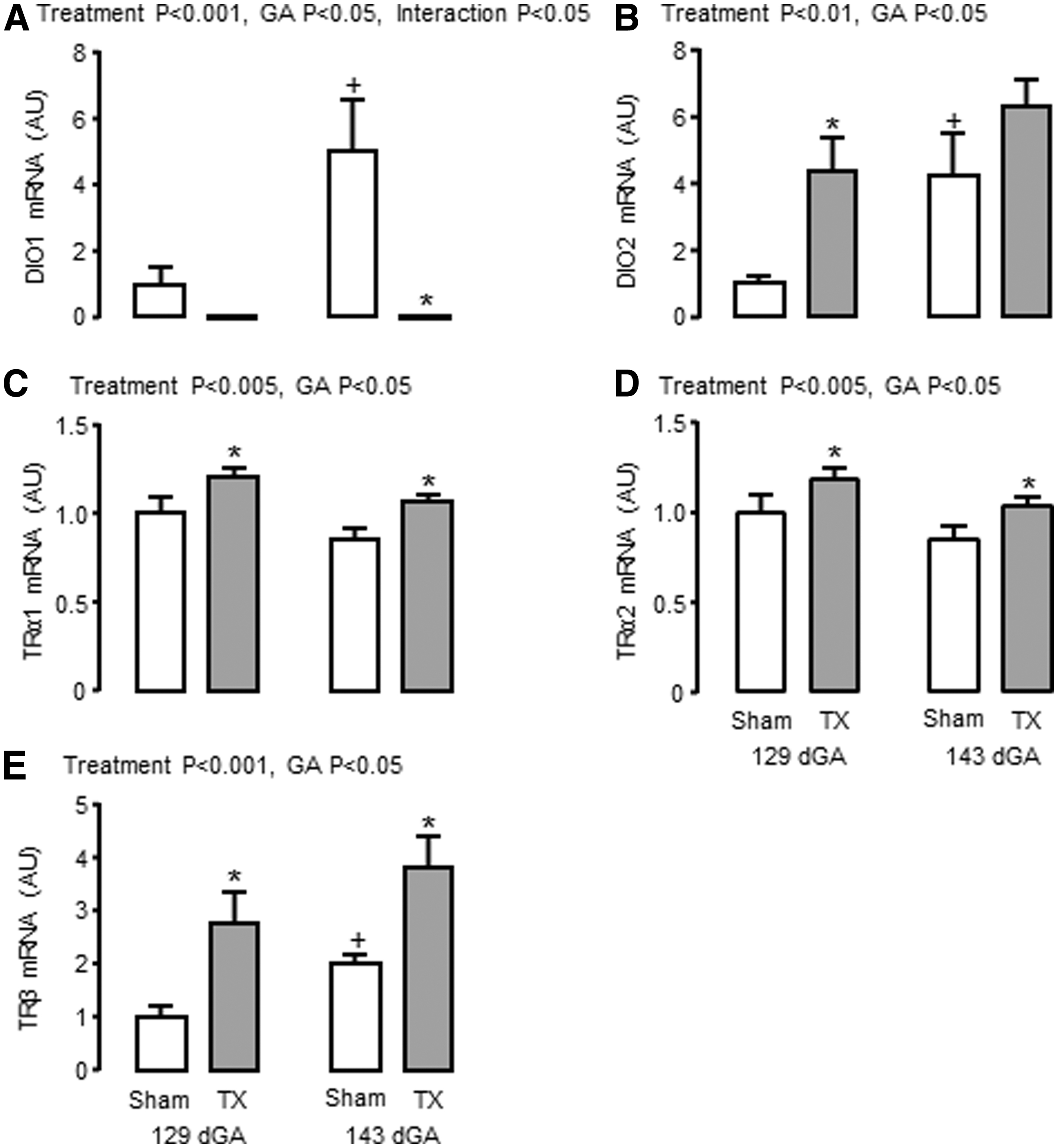
Mean (±SEM) mRNA abundance of
Discussion
This study has shown for the first time that thyroid hormone deficiency modifies the transcriptome, and hence, the growth and development of fetal adipose tissue, in a manner that is likely to compromise the ability of the neonate to maintain body temperature at birth and to increase its risk of metabolic dysfunction in later life. Fetal hypothyroidism caused a shift in the relative composition of UL and ML adipocyte types toward an increase in UL adipocyte mass due to hyperplasia and hypertrophy. Gene markers of UL adipocyte type, such as leptin, adiponectin, and lipoprotein lipase, were increased in TX ovine fetuses, and adipocyte proliferation was indicated by elevated levels of the mitotic marker PCNA and enrichment of gene pathways responsible for PPARγ and insulin-IGF signaling. The percentage of ML adipocytes in PAT was reduced by hypothyroidism in utero and, although the total amount of ML adipose tissue did not differ between sham and TX fetuses, UCP1 expression was impaired, when normalized to mitochondrial density estimated by CS activity, indicating a deficit in thermogenic capacity. Furthermore, bioinformatic analysis showed that, for a substantial number of genes, hypothyroidism prevented the maturational changes normally seen in the transcriptome of ovine PAT near term.
The effects of thyroid hormone deficiency on adipose tissue development before birth may be direct, via thyroid hormone response elements (TREs) on target genes (17), and/or indirect, via changes to energy expenditure and by interactions with other nuclear receptors or changes in fetal hormone concentrations, such as insulin and the IGFs. Basal metabolic rate is difficult to measure in utero, but previous studies have shown that thyroid hormone deficiency in the sheep fetus reduces the whole-body rate of oxygen consumption and is normalized by T4 replacement (35). The extent to which changes in global energy expenditure induced by hypothyroidism contribute to the modifications in the growth and development of fetal PAT in the present study remains to be determined.
The increased circulating concentration of insulin seen in the hypothyroid sheep fetus, secondary to pancreatic β-cell proliferation (20), is likely to be responsible, at least in part, for UL-specific PAT overgrowth. Indeed, a positive correlation was observed between fetal insulin concentration and relative UL adipocyte mass in the present study. Before birth, insulin stimulates growth of the axial skeleton and tissues such as adipose tissue (36). In the ovine fetus, hyperglycemia and hyperinsulinemia induced by fetal glucose infusion have also been shown to promote UL adipocyte growth with no change in ML adipocyte mass (37).
A variety of signaling pathways responsive to insulin and the IGFs, and known to be involved in the control of adipogenesis, were enriched in the adipose transcriptome by hypothyroidism in utero. These included PPARγ, apelin, and FoxO signaling. Furthermore, measurement of selected downstream target proteins showed upregulation of pS6K, GLUT4, and PPARγ in TX fetuses. Phosphorylation of S6K, without any change in mRNA abundance, indicated activation of the PI3-kinase pathway, which is known to regulate adipogenesis via a range of transcription factors and interacting molecular pathways (38,39). Transgenic mice with mutation in the S6K gene are growth retarded from embryonic life with reductions in pancreatic β-cell size and insulin content (40). This phenotype persists to adulthood and is associated with impaired adipogenesis, increased insulin sensitivity, and resistance to diet-induced obesity (41). Increased adipose GLUT4 expression in TX sheep fetuses may also contribute to adipogenesis via enhanced glucose uptake and lipid storage and are consistent with findings in rat pups hypothyroid in fetal and neonatal life (42).
While circulating IGF levels remained unchanged in TX sheep fetuses, adipose mRNA abundances for IGF-I and IGF-II were elevated, indicating potential upregulation of local synthesis and paracrine actions of the IGFs. Thyroid hormone deficiency in utero has been shown previously to modify IGF expression in the liver and skeletal muscle with tissue-specific effects on growth and development in fetal sheep (43,44). Insulin-IGF signaling pathways can also induce the synthesis of adipokines, such as leptin, apelin, and adiponectin, in part via interactions with PPARγ signaling. Previous studies have shown that hyperinsulinemia, in the presence of euglycemia, increases adipose leptin mRNA abundance in fetal sheep (45). High circulating levels of thyroid-stimulating hormone associated with TX may also stimulate leptin secretion, as reported in human adipose tissue cultured in vitro (46). The extent to which the increase in adipose adipokine expression and circulating leptin levels in the hypothyroid sheep fetus results from the greater UL adipocyte mass and/or greater capacity for adipokine synthesis and secretion in individual UL adipocytes remains to be established.
Thyroid deficiency in utero reduced adipose UCP1 expression and affected several genes in the thermogenic pathway, without changing ML adipocyte mass. Previous studies have shown that hypothyroid lambs are unable to maintain normal body temperature at delivery, and their PAT contains less UCP1 and more lipid (14). Maternal hypothyroidism in rats led to low adipose UCP1 mRNA abundance in the fetuses, which correlated with adipose T3 levels and was corrected by maternal thyroid hormone treatment (47). Furthermore, in cultured brown adipocytes taken from fetal rats, T3 causes an increase in UCP1 gene transcription, mRNA stability, and mitochondrial protein content (48). Moreover, a TRE has been reported upstream of the promoter region of the UCP1 gene (49). Suppression of adipose UCP1 levels were observed in TX sheep fetuses despite upregulation of other factors known to stimulate UCP1 expression, such as ADRβ3, IGF-I, leptin, and PPARγ, possibly as compensatory mechanisms.
The sympathomedullary system is primarily activated at birth by delivery into a cold environment and normally interacts with thyroid hormones to promote UCP1 expression and nonshivering thermogenesis. Although plasma catecholamine concentrations were not measured in the present study, PAT catecholamine content has been reported to be unaffected by TX in fetal sheep (50). Previous studies have also shown that noradrenergic-induced cellular respiration in PAT is suppressed in TX sheep fetuses, compared with those infused with T3, which suggests that functional adrenergic signaling may be impaired, despite elevated PAT mRNA abundance of some ADR isoforms (51). The effects of hypothyroidism in utero may also originate from abnormal formation of hypothalamic pathways responsible for adipose thermogenesis, especially since thyroid hormones have a key role in the development of the fetal brain (52).
Maturational changes in thyroid hormone metabolism and signaling were observed in fetal adipose tissue during late gestation, which were modified by thyroid hormone deficiency. In sham fetuses, mRNA abundances of iodothyronine deiodinases DIO1 and DIO2, and the TRβ, increased toward term. Upregulation of DIO1 and DIO2 enzyme activities has been demonstrated previously in ovine fetal PAT over the same period of gestation, in part due to the prepartum rise in plasma cortisol (53,54). Increased DIO1 enzyme activity in the PAT, and the liver and kidney, of the sheep fetus near term is likely to be responsible for the increase in plasma T3 seen close to term (54).
Hypothyroidism in utero had contrasting effects on the expression of DIO1 and DIO2 mRNA in ovine fetal PAT: DIO1 was reduced to negligible levels and DIO2 was increased in TX fetuses, in line with previous findings on deiodinase enzyme activity in adipose and other tissues of hypothyroid sheep and rat fetuses (47,53,55). Indeed, bioinformatic analysis identified DIO1 as the top-ranked gene affected by fetal TX in the current study with a 7.4-fold decrease in expression levels. Although DIO2 enzyme activity is much lower than DIO1 in ovine fetal PAT (54), the increase in DIO2 mRNA abundance may be an adaptive response to maintain local T3 production in hypothyroid conditions. The molecular mechanisms responsible for the tissue-specific control of deiodinase expression by thyroid hormone deficiency before birth remain to be established, although a TRE has been identified in the human Dio1 gene (56). Within the PAT of TX sheep fetuses, the mRNA abundances of both TRs, TRα and β, were increased in an apparent attempt to maintain local sensitivity to thyroid hormones in the face of systemic hypothyroidism. Fetal thyroid hormone deficiency appeared to upregulate TRα gene expression without affecting the relative proportions of the splice variants α1 and α2.
During hypothyroidism in utero, activation of adipogenesis, suppression of thermogenic capacity, and exposure of the fetus to high circulating levels of insulin and adipocytokines may have consequences for adipose function and insulin sensitivity in the longer term (8). Human infants exposed to hyperinsulinemia before birth, such as those born to obese mothers, have greater percentage body fat, higher umbilical cord leptin concentration, and raised indicators of insulin resistance compared with those born to lean mothers (57). In the present study, genomic pathways associated with insulin resistance were identified as enriched in PAT from TX fetuses. Furthermore, since there is a link between adiposity in the neonate and child (58), these findings suggest that the development of fetal adipose tissue and enhancement of insulin resistance pathways may predispose offspring hypothyroid in utero to obesity and metabolic disease in later life.
Several studies worldwide have shown that children born with CH have a greater body mass index and are more at risk of obesity, insulin resistance, and NAFL in early and young adult life compared with the general population, even when diagnosed and treated with T4 soon after birth (2 –6). Moreover, infants with more moderate reductions in thyroid hormones associated with prematurity or intrauterine growth retardation are also at greater risk of obesity and cardiometabolic dysfunction in later life (7,8). Collectively, these findings suggest that exposure to hypothyroidism in utero permanently alters adipose tissue development with consequences for adult health. Further investigations are required, however, to determine whether these programming effects arise directly from the altered adipose phenotype and/or indirectly from other changes in endocrine activity, metabolism, or appetite regulation that affect adult adiposity. For example, antithyroid drug treatment in pregnant rats leads to hyperleptinemia in the adult offspring and alterations in hypothalamic leptin signaling molecules indicative of leptin resistance (59). Hyperinsulinemia and overgrowth of UL adipose tissue in sheep fetuses infused with glucose were also associated with changes to the expression of neuropeptides in the appetite-regulatory regions of the hypothalamus (37). Elucidation of the molecular pathways influenced by thyroid deficiency in utero, and the long-term consequences for physiological function in a range of tissues, will enable greater understanding of the health outcomes in offspring exposed to hypothyroidism before birth.
Footnotes
Acknowledgments
The authors would like to thank technical staff at the Universities of Cambridge, Oxford Brookes, and Western Australia for their assistance in the study.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The project was funded in part by the Biotechnology and Biological Sciences Research Council, and a Research Excellence Award from Oxford Brookes University to A.J.F. S.E.H. was supported by a Nigel Groome PhD Studentship, Oxford Brookes University. Sample library preparation and RNA-seq work were performed at the Genomics and Transcriptomics Core, which is funded by the UK Medical Research Council (MRC) Metabolic Disease Unit (MRC_MC_UU_12012/5) and a Wellcome Trust Major Award (208363/Z/17/Z).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Supplementary Figure S7
Supplementary Figure S8