
Abstract
Background:
It is important to properly understand the molecular mechanisms of aggressive tumors among papillary thyroid carcinomas (PTCs) that are often the most indolent. Hypoxia inducible factor-1α (HIF-1α), induced by hypoxia, plays pivotal roles in the development and metastasis of the many tumors, including PTCs. Upregulation of telomerase reverse transcriptase (TERT) activity is found in highly invasive PTCs. Further, previous studies have reported that autophagy serves as a protective mechanism to facilitate PTC cell survival. We, therefore, hypothesized that there was a link between HIF-1α, TERT, and autophagy in promoting PTC progression.
Methods:
Immunohistochemistry staining was conducted to evaluate the expressions of HIF-1α, TERT, and autophagy marker, LC3-II, in matched PTC tumors and corresponding nontumor tissues. Two PTC cell lines (TPC-1 and BCPAP) were used in subsequent cytological function studies. Cell viability, proliferation, apoptosis, migration, and invasion were assessed during hypoxia, genetic enhancement and inhibition of TERT, and chemical and genetic inhibition of autophagy. The protein expression levels of the corresponding biomarkers were determined by Western blotting, and autophagy flow was detected. We characterized the molecular mechanism of PTC cell progression.
Results:
The protein expression levels of HIF-1α, TERT, and LC3-II were upregulated in PTCs and were significantly correlated with high tumor-node-metastasis stage. Further, an in vitro study indicated that HIF-1α induced by hypoxia functioned as a transcriptional activator by binding with sequences potentially located in the TERT promoter and was positively correlated with the malignant behavior of PTC cell lines. Overexpression of TERT inhibited the kinase activity of mammalian target of rapamycin (mTOR), resulting in the activation of autophagy. Functionally, TERT-induced autophagy provided a survival advantage to PTC cells during hypoxia stress.
Conclusions:
We identified a novel molecular mechanism involving the HIF-1α/TERT axis, which promoted PTC progression by inducing autophagy through mTOR during hypoxia stress. These findings may provide a basis for the new treatment of aggressive PTCs.
Introduction
Papillary thyroid carcinoma (PTC) is the most common endocrine malignancy, and most of these tumors have an excellent prognosis (1,2). However, ∼5–10% of PTC patients suffer from locally invasive and metastatic diseases that cannot be controlled by conventional therapy, resulting in a decreased survival rate and quality of life (3). Aggressive PTCs pose a particularly difficult therapeutic and prognostic challenge in today's thyroid cancer medical treatment regimen. Hence, an understanding of the underlying molecular mechanisms involved in the unusually high aggressiveness and mortality of PTCs in these patients is important in predicting the prognoses and the development of alternative treatments.
In various solid carcinomas, including thyroid cancer, accumulating evidence has shown that hypoxia is an important trigger for tumor cell survival and disease progression (4 –7). One of the most important proteins involved in the hypoxic activation cascade is the transcription factor hypoxia inducible factor-1α (HIF-1α). Previous studies have shown that HIF-1α is upregulated in PTCs and plays crucial roles in tumor invasion and metastasis by providing a growth advantage under hypoxic stress (8 –12). Currently, little is known about the hypoxic regulation of HIF-1α and the HIF-1α-dependent activation of target genes in thyroid carcinomas, which has major implications regarding tumor aggressiveness and local invasion or distant metastasis.
The human telomerase reverse transcriptase (TERT) is the catalytic subunit for human telomerase. Its expression is highly correlated with telomerase activity, which is an important factor in cell proliferation (13,14). Previous studies have shown that HIF-1α transactivates TERT expression in solid tumors and embryonic stem cells (15 –20), suggesting that telomerase activation is a mechanism that protects against genetic stress induced by hypoxia. With the discovery of a high expression of TERT in some types of cancers, it was also investigated in thyroid carcinoma and found to be common, especially in tumors with an aggressive behavior (21 –25). However, the mechanisms that regulate TERT levels in PTCs and the crosstalk between HIF-1α and TERT are poorly understood.
Recent reports have shown that cancer cells maintain a higher level of TERT expression to enable the cells to survive a harsh tumor microenvironment that is deprived of sufficient oxygen and nutrients (18). Thus, TERT plays a critical role not only in cell survival but also in metabolism. Studies have indicated that gain of TERT function modulates the activity of the mammalian target of rapamycin (mTOR) and induces autophagy, which is required for metabolic rewiring, to scavenge the nutrients that are necessary for fueling cancer cell growth in a challenging tumor microenvironment (26,27). Increasing studies have supported the central role of autophagy in the maintenance and development of various kinds of cancers, including thyroid carcinoma (28). Specifically, autophagy plays important roles in regulating the dedifferentiation process of PTCs, although the data are limited (29). Although several independent studies have shown that TERT activates autophagy, the specific molecular pathways involved in this process during hypoxia stress are poorly understood; the underlying molecular mechanisms associated with increased aggressiveness and poor prognosis in thyroid cancer remain unexplored.
In this study, we conducted immunohistochemical staining to evaluate the expression of HIF-1α, TERT, and the autophagy-related core molecule, LC3-II, in matched PTC tumors and corresponding nontumor tissues. The results showed that all of the proteins' expression levels mentioned earlier were upregulated in PTCs and significantly correlated with high tumor-node-metastasis (TNM) stage. Further, we examined the effects of hypoxia and TERT overexpression on PTC cell lines (TPC-1 and BCPAP), and identified a novel mechanism of hypoxia-induced TERT activation via HIF-1α in PTCs. Importantly, we found that HIF-1α bound to the promoter of TERT and acted as a potent transactivator. Moreover, overexpression of TERT inhibited mTOR kinase activity, resulting in the activation of autophagy. Functionally, TERT-induced autophagy provided a survival advantage to PTC cells during hypoxia stress. Taken together, our findings suggested a nexus among those signaling molecules, forming an HIF-1α-TERT-mTOR-autophagy axis, which promotes PTC progression during hypoxia stress.
Materials and Methods
Patients and immunohistochemistry
This study was approved by the Clinical Research Ethics Committee of Shanghai Jiao Tong University Affiliated Sixth People's Hospital (Shanghai, China), and written informed consent was obtained from each patient. Seventy-five patients with PTCs, who were admitted to our hospital and underwent thyroidectomy between 2014 and 2017, were identified as eligible for the study. The inclusion criteria for the study were as follows: (i) a histopathological diagnosis of PTC; (ii) no history of other malignant tumors; (iii) no neck external irradiation; and (iv) the availability of an adequate medical history. Data collected from each patient included age, sex, tumor size, and TNM stage, which were classified based on the criteria of the American Joint Committee on Cancer (AJCC, 7th edition) for differentiated thyroid cancer. Thyroid tissue samples consisted of two parts, cancer tissues and adjacent noncancer tissues, which were confirmed by histopathological examination. The corresponding nontumor thyroid tissues were obtained at least 1 cm away from the tumor. The specimens were processed immediately after sectioning and were stored in liquid nitrogen.
Immunohistochemical staining was performed by using a tissue microarray, which was established with formalin-fixed and paraffin-embedded surgical specimens. Primary antibodies against HIF-1α (1:50, Rabbit. no. 36169; Cell Signaling Technology, Inc., Danvers, MA), TERT (1:100 dilution; Rabbit. no. ab216625; Abcam, Cambridge, MA), and LC3-II (1:50, Rabbit. no. 3868; Cell Signaling Technology) were used. The immunohistochemical procedure was performed as previously described (30). The staining intensity and extent were scored independently by two pathologists who were blinded to the patient information, using the semiquantitative immunoreactivity scoring system as previously described (31) The staining intensities were graded as 0 (negative), 1 (weak), 2 (moderate), and 3 (strong), and the staining extent was scored as the percentage of positive cells (0–100%). The staining intensity and extent were multiplied to generate immunohistochemistry scores.
Cell lines, reagents, and transfection
PTC cell lines, BCPAP (papillary, BRAFV600E mutation) and TPC-1 (papillary, RET/PTC rearrangement), were purchased from the Institute of Biochemistry and Cell Biology (SIBS, CAS, Shanghai, China). All cell lines were confirmed to contain the BRAFV600E mutation, short tandem repeat, and amelogenin authentication as described in the ANSI Standard by the ATCC Standards Development Organization (SDO). All cell lines were maintained in RPMI 1640 medium (Cat. no. 11875-093; Gibco) supplemented with 10% fetal bovine serum (FBS, Cat. no. 16000-044; Gibco) in a 5% CO2 atmosphere at 37°C. Additional 1% non-essential amino acids (Cat. no. 11140050; ThermoFisher) was supplemented when culturing BCPAP cells.
To simulate the hypoxic environment, cells were cultured under normoxic conditions (20% O2, 5% CO2, 95% humidity) until they were 70–80% confluent and were then subjected to hypoxic conditions (1% O2, 5% CO2, 95% humidity) for 24 hours in an exposure chamber (Biospherix, Lacona, NY). All compounds used in the study were obtained from Selleck (Houston, TX), dissolved in dimethyl sulfoxide, and used at the indicated concentrations.
For in vitro studies, HIF-1α overexpression was achieved by transfection with pcDNA3.1 (Asia-Vector Biotechnology Co. Ltd., Shanghai, China) using lipo2000 Transfection (Thermo Fisher) reagents according to the manufacturer's protocol. After transient transfection for 48 hours, the expression of HIF-1α protein was confirmed by Western blotting. Premade lentiviral particles expressing fusion targets of TERT and RFP-GFP-LC3 were purchased from Genomeditech Co. Ltd (Shanghai, China) and transfected into BCPAP and TPC-1 cells according to the manufacturer's protocol. Lentiviruses containing empty plasmids (vector) and lentiviruses containing nonspecific short hairpin RNA (shRNA; scramble) were used as controls, respectively. TERT and ATG7 shRNA and control shRNA were purchased from Asia-Vector Biotechnology Co. Ltd. The knockdown efficiency was confirmed by fluorescence microscopy and Western blotting.
Cell Counting Kit-8 and the colony formation assay
Cellular proliferation of BCPAP and TPC-1 cells was assessed by using Cell Counting Kit-8 (CCK-8) assay (Yeasen, Shanghai, China), as described in our previous study (32). The colony formation assay was performed by seeding 300 cells per treatment type in six-well plates in triplicates, and culturing for 12 days before Crystal Violet staining of viable colonies.
Transwell migration and invasion assays
Transwell migration and invasion assays were performed as previously described (33).
Flow cytometric analysis
For apoptosis analysis, PTC cells were treated before flow cytometric apoptosis analysis and were then stained with Annexin V/7-AAD (Affymetrix eBioscience). Cells were harvested and washed twice with cold phosphate-buffered saline (PBS), followed by resuspension in a single-cell suspension. The cells were then fixed in cold 75% ethanol overnight at 4°C. The fixed cells were collected, washed twice with PBS, resuspended in PBS containing Annexin V-PE and 7-AAD, and finally incubated for at least 30 minutes in the dark at 4°C. The percentage of apoptotic cells was measured by flow cytometry (Beckman Coulter). The living cells were Annexin V-PE and 7-AAD and were double negative, whereas the late apoptotic or secondary necrotic cells were double positive. The early apoptotic cells were only Annexin V–PE positive, whereas the isolated nuclei or cellular debris were only 7-AAD positive.
Western blotting and the lactate dehydrogenase assay
Western blotting was conducted as previously described (34,35). Anti-HIF-1α (1:1000), anti-TERT (1:500), anti-E-cadherin (1:1000), anti-caspase-3 (1:1000, no. 9662), anti-p-mTOR (1:1000), anti-mTOR (1:1000), anti-p-S6K (1:1000), anti-S6K (1:1000), anti-LC3-II (1:1000), anti-P62 (1:1000), and anti-glyceraldehyde-3-phosphate dehydrogenase (GAPDH) antibodies (1:2000) were purchased from Cell Signaling Technology, Inc. Lactate dehydrogenase (LDH) levels in the culture medium were measured by using colorimetric assay kits (Abcam) according to the manufacturer's instructions.
Dual-luciferase reporter assay
The dual-luciferase reporter assay (no. E1910; Promega) was used to measure luciferase activity. Reporter constructs were introduced into cells by transfection using Lipofectamine 2000 (Invitrogen). All luciferase activity measurements were made at 48 hours post transfection by using a TD-20/20 luminometer (Turner BioSystems, Inc.).
Chromatin immunoprecipitation
To evaluate the interaction between HIF-1α and the TERT promoter region, chromatin immunoprecipitation (ChIP) assays were conducted by using a kit (Millipore) according to the manufacturer's instructions. Polymerase chain reaction was performed by using three primer sets for the three areas containing potential HIF-1α binding sites in the TERT promoter sequence: (i) TERT-F: 5′-GGCCGTGTGGCTTCTACTG-3′ and TERT-R: 5′-CAACATCTGGTCACATCCCG-3′ (ii) TERT-F: 5′-GTCCAGGGAGCAATGCGT-3′ and TERT-R: 5′-CCTGGGTCTCCGGATCAGG-3′ (iii) TERT-F: 5′-ACGTGGCGGAGGGACT-3′ and TERT-R: 5′-ACCCGGGAGGGGTCG-3′. Genomic DNA and IgG were used as controls.
Confocal microscopy
GFP-RFP-LC3-transfected cells were grown on coverslips and treated. Fluorescent microscopy was used for the analysis of changes in various autophagic structures.
Statistical analysis
Results are expressed as the mean ± standard deviations from at least three independent experiments. Graphpad Prism (version 6.01; GraphPad Software, Inc., CA) was used for the statistical analyses. The Kruskal-Wallis test was used for determining the statistical significance of differences in variables with a non-normal distribution. The Student's t-test and one-way analysis of variance followed by Tukey's post hoc test were used to measure comparisons between groups with normal distributions. Differences with a value of p < 0.05 were considered statistically significant.
Results
HIF-1α and TERT are upregulated in PTC tissues, and TERT expression is positively associated with TNM stages
To determine whether hypoxia was a general characteristic in PTC tissues and the TERT activity, we detected HIF-1α (hypoxia-specific indicator) and TERT expressions in 75 paired PTC and adjacent noncancerous tissues by using immunohistochemistry. Representative staining examples are shown in Figure 1A and B. The expression levels of HIF-1α and TERT in the PTC specimens were significantly higher than those in paracarcinoma tissues, with p < 0.0001 and p < 0.05, respectively (Fig. 1C, D). In addition, the HIF-1α expression was positively correlated with the levels of TERT in PTCs (Fig. 1E). The association of TERT expression was next determined in PTC tissues with known TNM stages. The results revealed that TERT expression was significantly greater in stage II than stage I, and stage III showed the highest levels of expression (Fig. 1F).
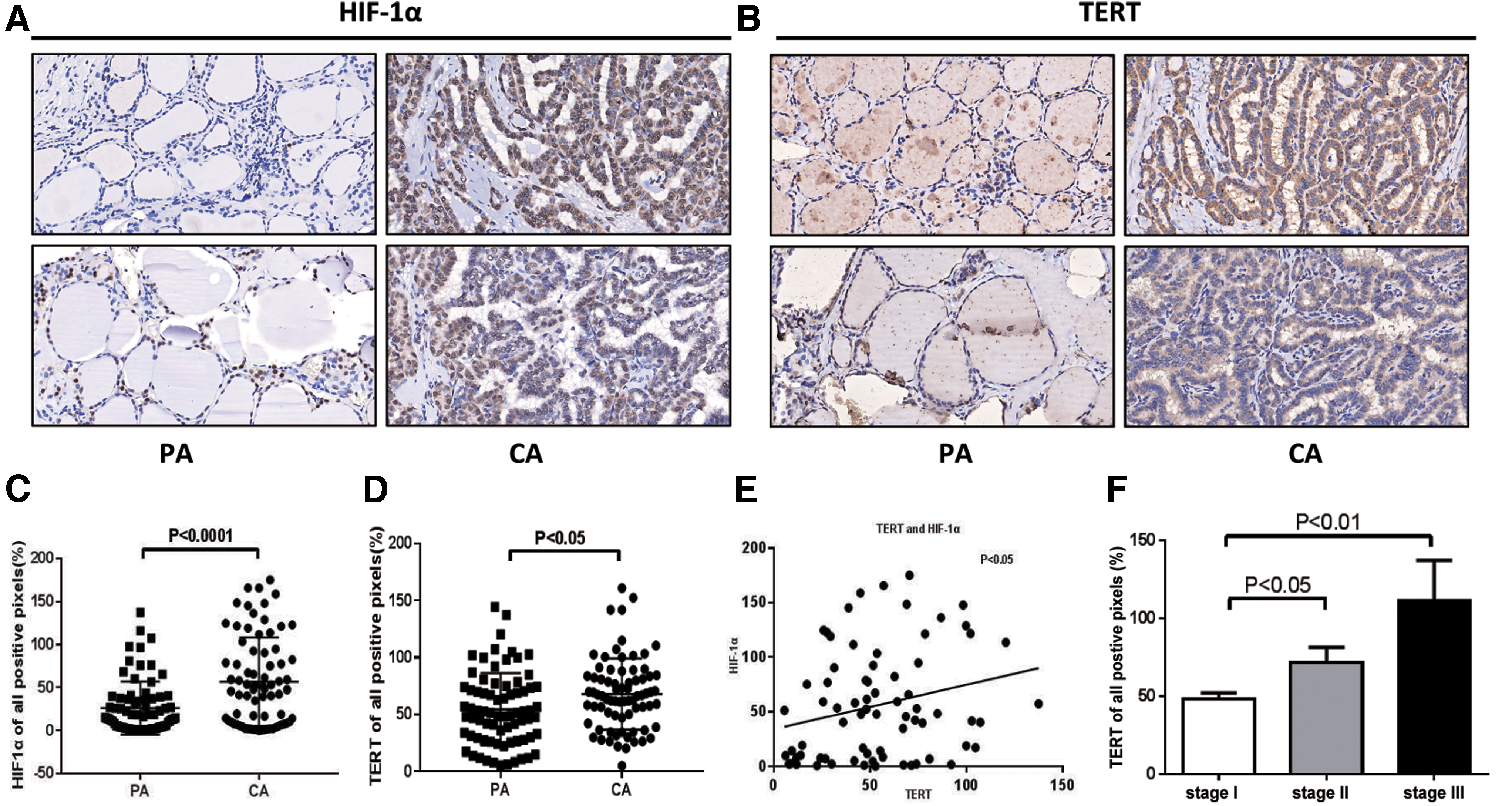
HIF-1α and TERT are relatively overexpressed in PTC tissues compared with paired adjacent normal tissues by IHC. (
Taken together, these results indicated that both HIF-1α and TERT were upregulated in PTC tissues, and TERT expression is positively associated with TNM stages.
Hypoxia and TERT overexpression endow PTC cells with aggressiveness in vitro
To characterize the potential effect of hypoxia and TERT overexpression on cellular proliferation, migration and invasion, and apoptosis, which are required for tumorigenesis and metastasis, we performed a series of investigations to evaluate them in vitro. PTC cell lines, BCPAP and TPC-1, were selected and divided into three groups: control (20% O2), hypoxia (1% O2), and TERT overexpression groups. Figure 2 shows that treatments with hypoxia and TERT upregulation significantly enhanced the ability of cells to proliferate (Fig. 2A), increased migration and invasion (Fig. 2B), increased colony formation (Fig. 2C), and reduced cell apoptosis (Fig. 2D and Supplementary Fig. S1A), when compared with control cells. Increased glucose consumption distinguishes cancer cells from normal cells, and it is referred to as the “Warburg effect” because of increased glycolysis, which partly results from HIF-1α activity (36). LDH is a key glycolytic enzyme and a hallmark of aggressive cancers (37). To gain further insight into the metabolic status of PTC cells, we measured LDH levels, with results indicating a more pronounced increase with hypoxia and TERT overexpression (Fig. 2E).
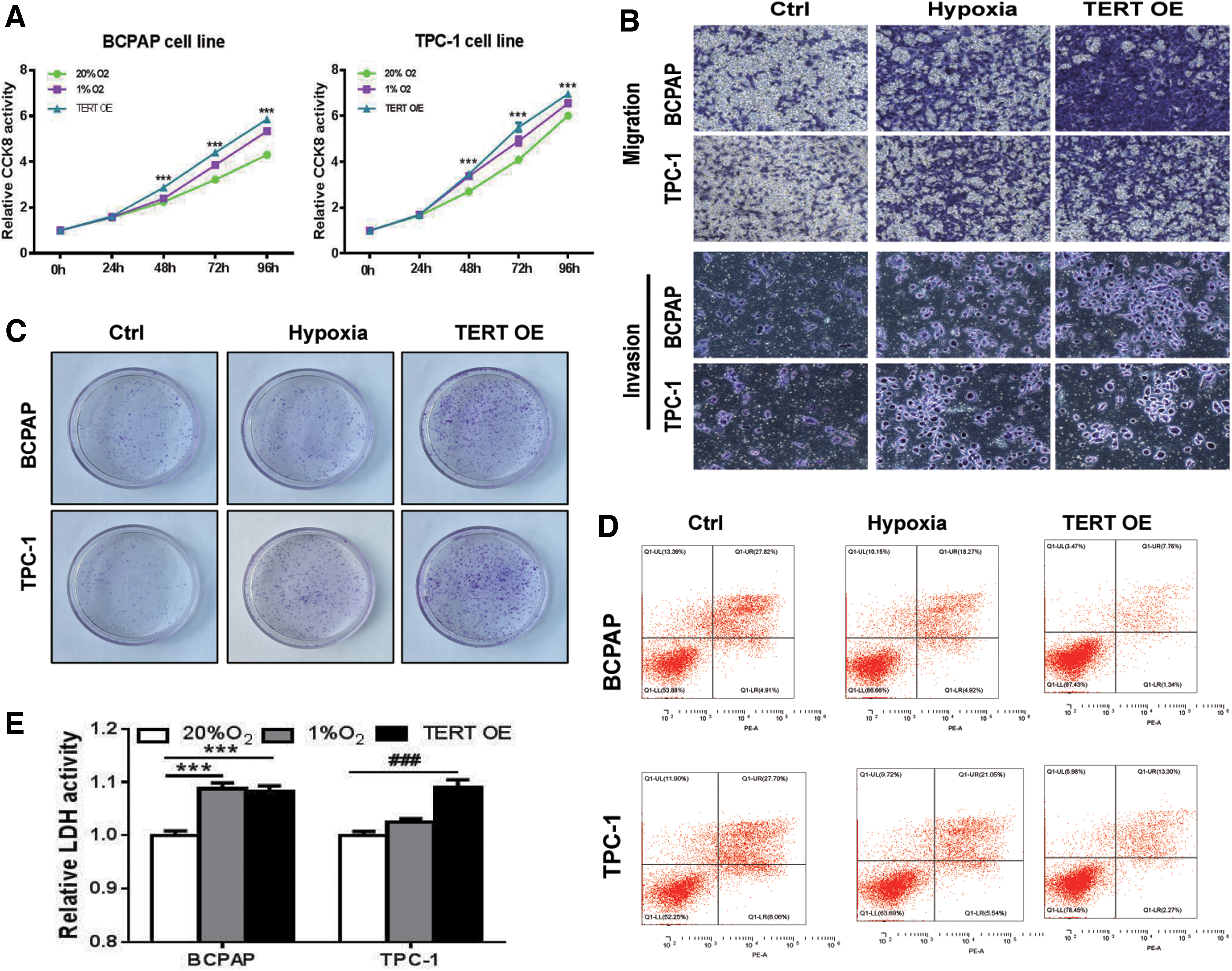
Both hypoxia and TERT overexpression promote tumor growth, invasiveness, and reduce apoptosis in PTC cells. Lentiviral transfection is used to overexpress TERT in the indicated cancer cells, followed by performance of assays of CCK-8 of cell proliferation (
HIF-1α binds and activates the TERT promoter under hypoxia stress, enhancing TERT expression
HIF-1α is a transcription factor that is known to regulate the downstream genes involved in hypoxia. It has been reported that HIF-1α gene expression increases during hypoxic conditions in tumors, including PTC (38). To understand the gene regulatory mechanism of HIF-1α and TERT, we initially measured the protein levels of HIF-1α and TERT in the hypoxia and TERT overexpression models, respectively. In both the BCPAP and TPC-1 hypoxic cellular models, the expression of TERT was significantly upregulated along with the overexpression of HIF-1α. Importantly, in the TERT overexpression models, the expression of HIF-1α did not significantly increase (Fig. 3A, B). Thus, the result indicated that HIF-1α may play a critical role in mediating the expression of TERT. To determine the effects of hypoxia and TERT upregulation on the epithelial–mesenchymal transition, we measured the protein levels of E-cadherin. When HIF-1α and TERT increased, the level of E-cadherin, a protein required for metastasis in multiple models of cancer, was reduced (Fig. 3A, B).
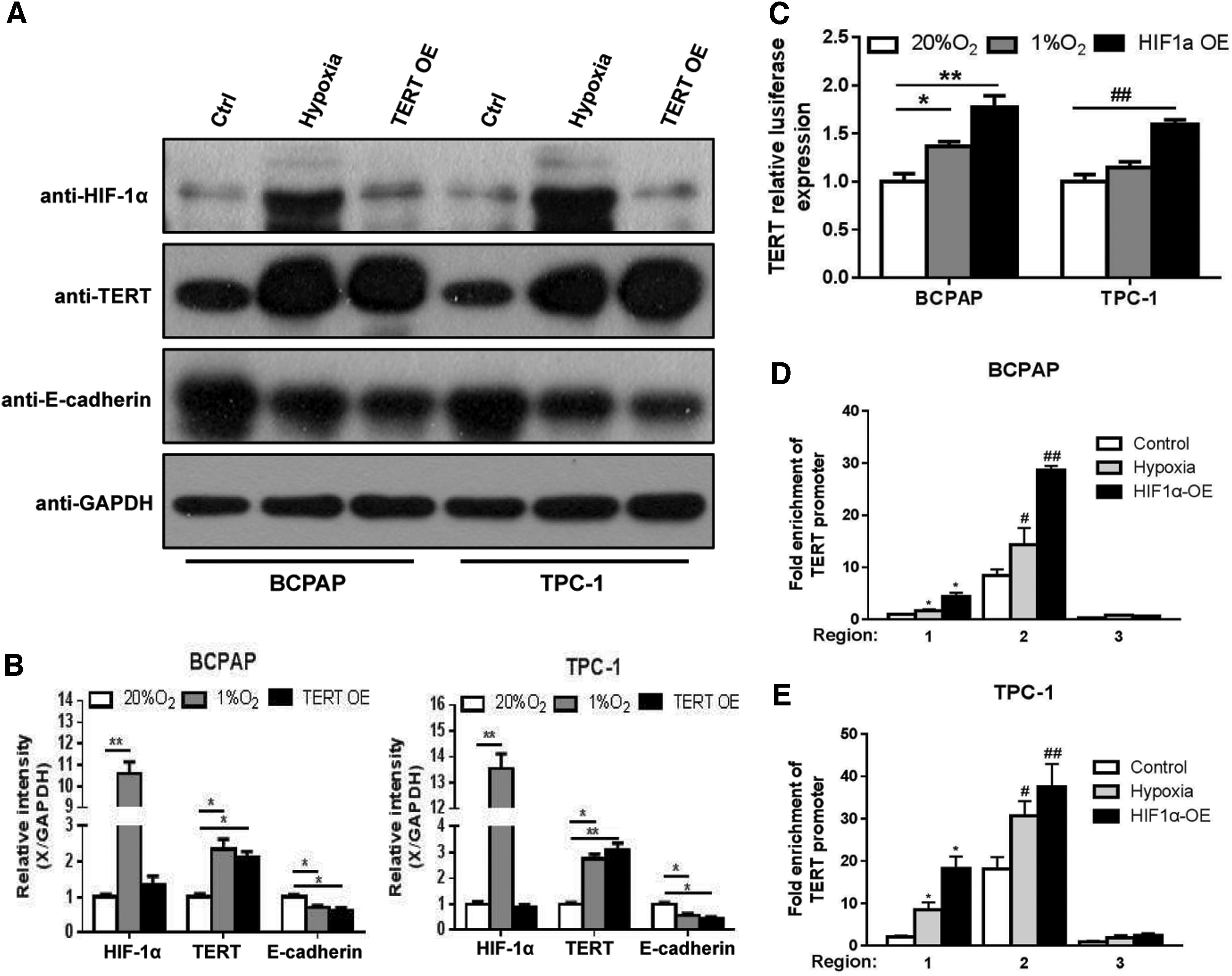
HIF-1α induced by hypoxia acts as a transcriptional activator by binding with sequences located in the TERT promoter in PTC cells. (
To investigate how HIF-1α promoted TERT expression in PTC cells, dual-luciferase reporter assays were performed to determine whether TERT promoter activity was increased in hypoxia treated- and in HIF-1α-overexpressing BCPAP and TPC-1 cells. The results showed that both hypoxia and HIF-1α overexpression enhanced TERT promoter activity (Fig. 3C). As has been demonstrated, HIF-1α is an important transcription factor that regulates TERT. ChIP analyses of the TERT promoter by using an antibody specific to HIF-1α and quantitative real-time polymerase chain reaction primers specific to the TERT promoter region were performed. The results indicated that the sequences at −977 to −850 and −570 to −428 of the TERT promoter were HIF-1α functional binding sites (Fig. 3D, E, region 1 and region 2). Taken together, our in vitro study indicated that HIF-1α induced by hypoxia functioned as a transcriptional activator by binding with sequences potentially located in the TERT promoter, leading to the upregulation of TERT expression.
LC3-II is overexpressed in PTC tissues and positively associated with HIF-1α, TERT, and TNM stages
Because TERT and autophagy gain of function have been observed in several cancers, we proposed a model in which TERT activates autophagy, which is important for generating essential nutrients in a metabolically challenged tumor microenvironment, such as hypoxia (26,27). Thus, we further measured the expression level of LC3-II, a microtubule-associated protein that is a key marker of autophagy, by immunohistochemical staining analysis in 75 PTC patients. The expression levels of LC3-II in the PTC specimens were significantly greater than those in paracarcinoma tissues (p < 0.05, Fig. 4A, B). Moreover, the LC3-II expression was positively correlated with the levels of HIF-1α and TERT (Fig. 4C, D). Notably, as in TERT, the expression level of LC3-II was also positively correlated with TNM stage (Fig. 4E).
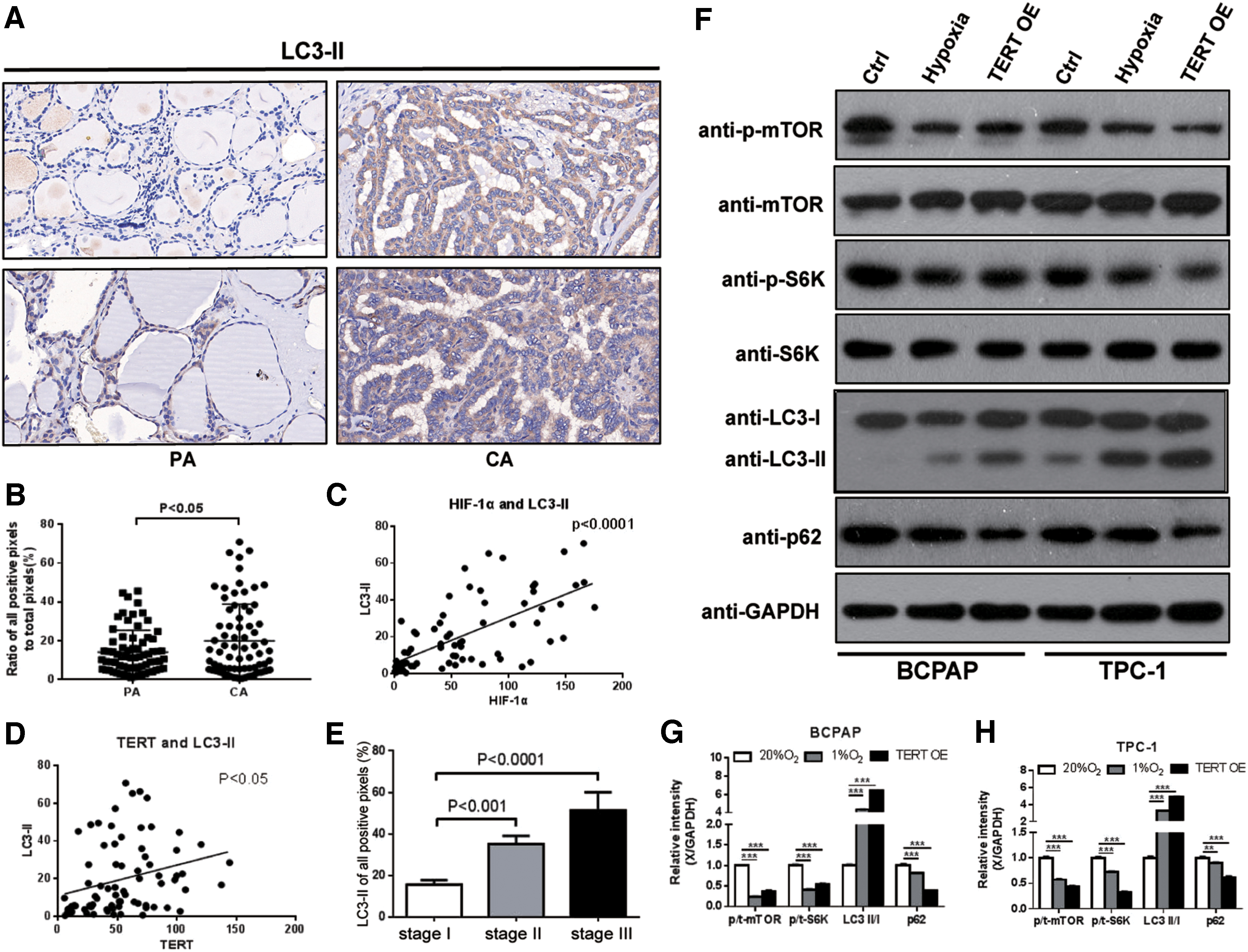
LC3-II is significantly unregulated in PTC tissues and positively associated with HIF-1α, TERT, and TNM stages. Hypoxia and TERT overexpression induced autophagy-related proteins changes. (
Hypoxia and TERT overexpression induce autophagy in PTC cells by decreasing the kinase activity of mTOR
Although independent investigations showed that telomerase activates autophagy, the underlying molecular mechanisms are unclear. mTOR, as one of the most well-characterized central negative regulators of autophagy, has been proposed to have a functional regulatory relationship with TERT (26,27). Next, to determine whether during hypoxia stress, TERT-mediated inhibition of mTOR was translated into activation of autophagy, Western blotting of autophagy-related proteins (mTOR, S6K, LC3-II, and p62) was measured in the hypoxia and TERT overexpression models.
In PTC cells, hypoxia and TERT overexpression decreased the kinase activity of mTOR as determined by decreased phosphorylation of the mTOR target pS6K. Post-translational modification of LC3-I to LC3-II is considered a reliable marker for the induction of autophagy. The ratio of LC3-II/LC3-I was increased both in hypoxia and in TERT overexpression cells, which was accompanied by a decrease in p62, acting as a readout of autophagic degradation. Collectively, these data indicated that both hypoxia and upregulated TERT induced autophagy by suppressing the kinase activity of mTOR (Fig. 4F–H).
Under both normal and amino acid starvation conditions, silencing of TERT reduce the malignant behavior of PTC cells and autophagy levels by increasing the kinase activity of mTOR
We next elucidated the functional significance of TERT-mediated autophagy in PTC cells. For this purpose, TERT-knockdown (TERT-KD) PTC cells were successfully generated by treatment with shRNA. The results showed that under both basal (10% FBS) and amino acid-deprived conditions (Earle's Balanced Salt Solution [EBSS]), TERT downregulation significantly decreased the abilities of cells to proliferate (Fig. 5A), migration (Fig. 5B), invasion (Fig. 5C), the level of LDH (Fig. 5D), and colony formation (Fig. 5E), but increased cell apoptosis (Fig. 5F and Supplementary Fig. S1B, C), when compared with the untreated control cells. Immunoblotting demonstrated the same invasive (decreased) and apoptotic (increased) changes. Knockdown of TERT was observed to enhance the activation of E-cadherin and caspase-3 (Fig. 6A–C). In addition, compared with the control (TERT unknockdown) groups, under both basal (10% FBS) and amino acid-deprived conditions (EBSS), TERT-KD strongly stimulated the kinase activity of p-mTOR and p-S6K, decreased the ratio of LC3-II/LC3-I, which was accompanied by a decrease in p62 (Fig. 6D–F).
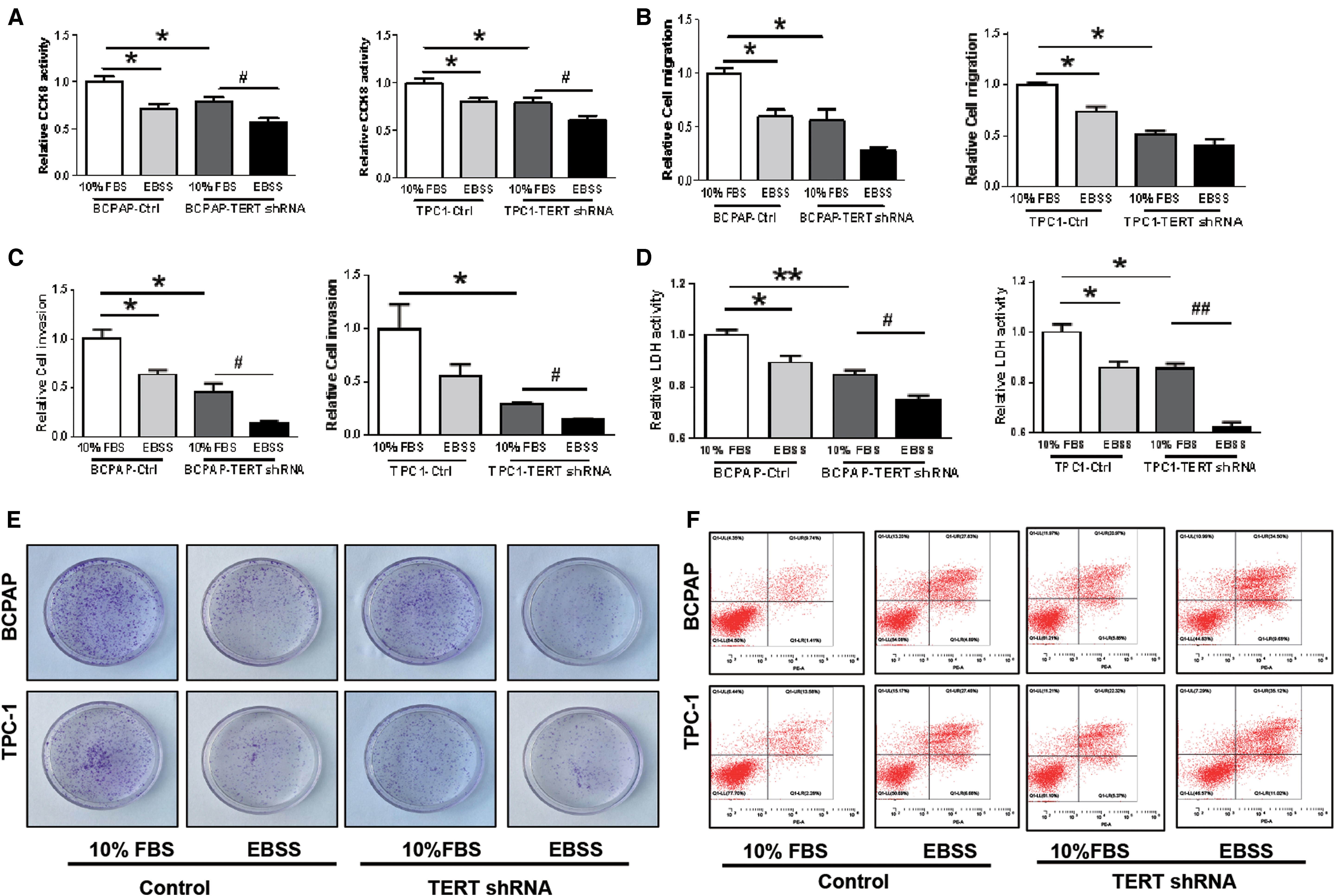
Under both basal (10% FBS) and amino acid-deprived conditions (EBSS), shRNA-TERT significantly decreases the ability of cell proliferation (

Western blotting further indicated that TERT-KD PTC cells decrease aggressiveness, increase apoptosis, and impair autophagy activity through the mTOR pathway under both basal and amino acid-deprived conditions. TERT knockdown results in a significant decrease of TERT and increased expression of E-cadherin and cleaved-caspase3 proteins (
Nutrient (including amino acids and glucose) depletion is one of the most potent physiological inducers of autophagy, which can be mimicked in vitro by withdrawing nutrients, such as amino acids. Therefore, as our experimental results showed, compared with FBS-treated cells, the ratio of LC3-II/LC3-I expression was increased in EBSS-treated groups, regardless of whether TERT was knocked down.
Taken together, these results demonstrated that TERT-KD PTC cells exhibited decreased aggressiveness and impaired autophagy activity through the mTOR pathway during both basal and amino acid-deprived conditions.
Genetic and chloroquine inhibition of autophagy attenuates PTC cell proliferation, migration, and invasion and promotes apoptosis while upregulation of TERT enhances autophagy activity through the mTOR pathway
Because PTC cells showed higher levels of autophagy than normal thyroid cells, we next examined the role of autophagy in PTC cells. Two methods of inhibiting autophagy were used, gene knockdown and chloroquine (CQ). PTC cells were treated with shRNA targeting autophagy-related (ATG) protein ATG7, a ubiquitin E1-like enzyme essential for autophagosome formation, to genetically impair the autophagy pathway. CQ is a lysosomotropic drug that inhibits autophagy at the last stage by preventing autophagosome–lysosome fusion. As shown in Figure 7, treatment with both ATG7-KD and CQ significantly inhibited the ability of PTC cellular proliferation (Fig. 7A), colony formation (Fig. 7B), migration (Fig. 7C), and invasion (Fig. 7D), but it made the cells more susceptible to apoptosis (Fig. 7E and Supplementary Fig. S1D). Immunoblotting showed the same changes by the dramatic decrease of LC3-II and accumulation of p62 (Supplementary Fig. S2). In addition, we transfected PTC cells by using GFP-RFP-LC3 lentiviral particles and monitored autophagy flux. Compared with the control group, ATG7-KD inhibited the formation of autophagosomes (decreased LC3-II puncta, Figs. 8 and 9) while CQ prevented the fusion process of autophagy lysosomes (increased LC3-II puncta, Figs. 8 and 9), both of which suggested a defective autophagy flux.

Inhibition of autophagy weakens malignant behavior of PTC cells; meanwhile, TERT overexpression endowed cells with aggressiveness in vitro. Genetic (ATG7 KD) and CQ inhibition of autophagy attenuates PTC cell proliferation (
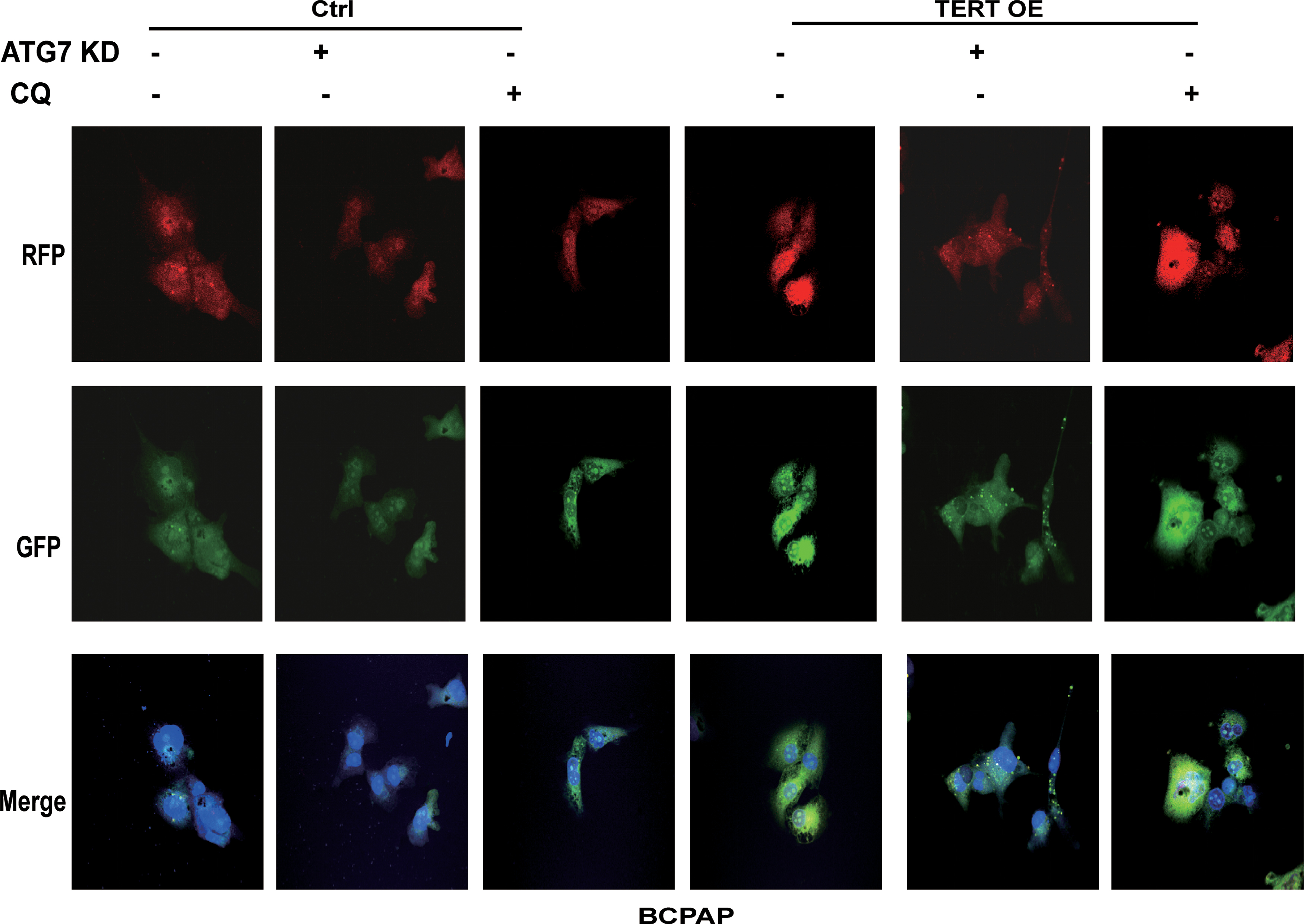
Live-cell confocal microscopic images of RFP-GFP/LC3-II–transfected BCPAP cells showed that ATG7 KD decreased LC3-II-positive puncta and CQ increased LC3-II-positive puncta, while TERT overexpression induced formation of cytoplasmic LC3-II puncta. Color images are available online.
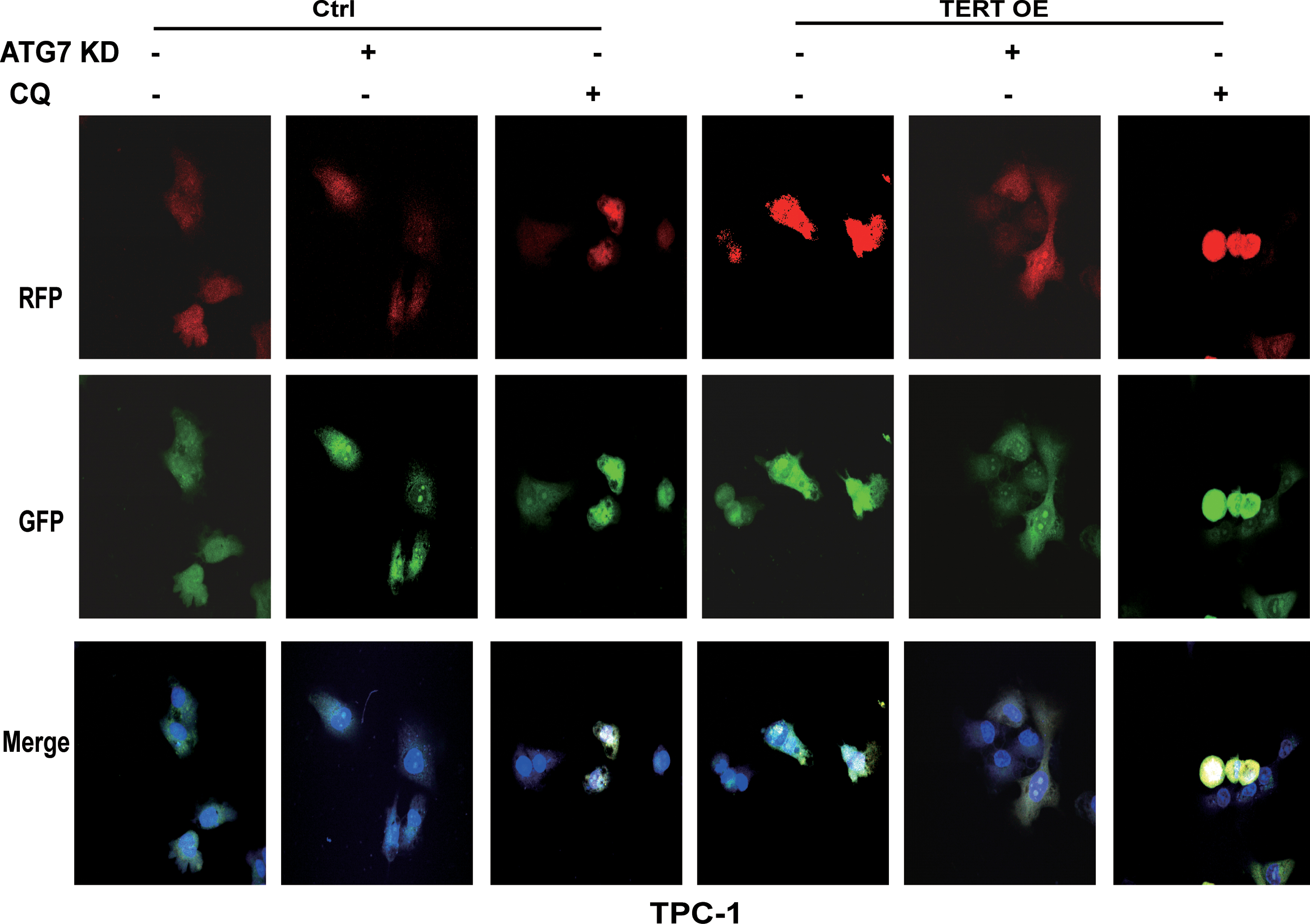
Live-cell confocal microscopic images of RFP-GFP/LC3-II–transfected TPC-1 cells showed that ATG7 KD decreased LC3-II-positive puncta and CQ increased LC3-II-positive puncta, while TERT overexpression induced formation of cytoplasmic LC3-II puncta. Color images are available online.
To further explore the contribution of TERT to the activation of autophagy, with the inhibition of autophagy, we examined the effects of overexpression of TERT on PTC cells. Overexpression of TERT increased the proliferation of PTC cells (Fig. 7A), as well as their colony formation (Fig. 7B), migration (Fig. 7C), and invasion (Fig. 7D), and it decreased cell apoptosis (Fig. 7E and Supplementary Fig. S1D), when compared with the untreated control. Meanwhile, the expression levels of LDH were also significantly increased (Fig. 7F, G). Western blotting analysis showed that overexpression of TERT increased the level of LC3-II and decreased the kinase activity of p-mTOR and p-S6K, indicating the activation of autophagy by mTOR suppression (Supplementary Fig. S2). Consistently, the number of LC3-II-stained puncta increased, suggesting an important role for TERT in the process of autophagy flux (Figs. 8 and 9).
Collectively, these results indicated that autophagy served as a prosurvival mechanism in PTC cells. Chemical or genetic inhibition of autophagy in PTC cells dramatically decreased cell survival and aggressiveness, but it promoted apoptosis. Functionally, TERT modulated mTOR activity and induced autophagy, which provided a survival advantage to PTC cells in a hypoxic tumor microenvironment.
Discussion
In this study, our results suggest that HIF-1α, TERT, and LC3-II are highly expressed in PTC tissues and are associated with higher TNM staging. Further, we provide experimental evidence showing that the hypoxia regulatory factor, HIF-1α, was a potent TERT transactivator in PTC cells. We also identified the role of TERT as a positive regulator of autophagy by inhibiting the kinase activity of mTOR, to provide a survival advantage to PTC cells during hypoxic stress. Thus, we propose a novel protective mechanism model, the HIF-1α-TERT-mTOR-autophagy axis, which is involved in PTC progression during hypoxic stress.
The definitive molecular mechanisms contributing toward the PTC invasion and metastasis remain poorly understood. Consistent with previous reports, our data confirmed that HIF-1α was highly expressed in PTC tissues, and its upregulation was associated with more aggressive tumors. Moreover, compared with normal thyroid cells, the expression levels of proteins HIF-1α, TERT, and LC3-II in TPC-1 and BCPAP cells are increased, and their migration and invasion capabilities are enhanced (Supplementary Fig. S3). Thus, we next explored the mechanisms by which HIF-1α was involved in PTC progression. The results showed that exposure of PTC cell lines to hypoxic growth conditions induced HIF-1α-dependent upregulation of TERT expression through binding sites located at −977 to −850 and −570 to −428 in the TERT promoter. Until our present report, in thyroid cancer, the mechanisms involved in hypoxia-induced TERT expression were unknown.
TERT, the catalytic subunit of telomerase, has attracted particular attention because it has been consistently detected in 80–90% of malignant tumors (39). Investigators have shown that the presence of TERT promoter mutations was associated with distant metastases, worse response to treatment, and poor survival in several types of cancer, namely in melanoma (40), gliomas (41), as well as in thyroid carcinomas (42). Previous studies directly showed an association of TERT promoter mutations with increased TERT protein expression in thyroid cancers (21,23). In this study, we focused on the expression level of TERT in PTC. Consistent with previous studies (21,23,43), immunohistochemical analysis indicated that TERT was successfully upregulated in PTC tissues and its expression was positively associated with TNM stages. Further, TERT overexpression endowed PTC cells with an aggressive behavior in vitro. Our study emphasized the importance of the tumor microenvironment, hypoxia, on the regulation of TERT expression. It has previously been shown that HIF-1α can transactivate TERT expression in immortal human cell lines (15,18,19), murine embryonic stem cells (20), trophoblast cells (17), and fish cells (44) on exposure to a hypoxic environment. These earlier studies further demonstrated that activation of telomerase on exposure of cells to a hypoxic environment was accompanied by an HIF-1α-dependent transactivation of TERT expression, as observed in PTC cells.
Historically, research on TERT was mainly focused on its ability to maintain telomere length and continually promoted cell proliferation. However, TERT has also been found to have nontelomere-dependent functions in recent years, such as involvement in cell signaling pathway (45), and protection of mitochondrial DNA (46). In this study, our results indicated that overexpression of TERT inhibited the mTOR kinase activity, resulting in the activation of autophagy. Functionally, TERT-induced autophagy provides a survival advantage to PTC cells during hypoxia stress. In our study, autophagy activity was elevated in PTC tissues and was positively associated with TNM stages. Moreover, inhibition of autophagy attenuated the malignant behavior of PTC cells and promoted apoptosis. The underlying mechanism may be that autophagy can benefit PTC cells in a harsh tumor microenvironment (deprived of sufficient oxygen and nutrients) by eliminating wastes, preventing accumulation of toxins, and supplying energy and components for cell survival and metabolism (26,27,47). Nutrient depletion is the most potent physiological inducer of autophagy, which can be mimicked in vitro by withdrawing nutrients, such as amino acids. We found an increase in cell apoptosis, despite the increase in the levels of autophagy-related proteins (LC3-II). To explain this, we further explored the effect of EBSS on PTC cells at different time points (0, 3, 6, 12, and 24 hours; Supplementary Fig. S4A–C). Apoptosis of PTC cells decreased at 0, 3, and 6 hours, but it increased at 12 and 24 hours. The expression level of LC3-II gradually increased. This can be explained by EBSS-treatment activating autophagy of PTC cells, which has a protective effect, so the apoptosis decreases at the beginning; and as time goes on, the nutrients are eventually exhausted, resulting in increased apoptosis of PTC cells.
The TERT-mTOR interaction in tumorigenesis has been investigated in other malignancies (26,27), but the underlying molecular mechanisms in PTC are unclear. mTOR kinase is one of the most well-characterized negative regulators of autophagy, which acts as a signaling hub for integrating upstream nutrient cues. Although tumors alter their metabolism to predominantly use glucose, the Warburg's effect, cancer cells still require amino acids as building blocks to support proliferation. TERT may play a regulatory role in the tuning of mTOR activity to an optimal level, so that these building blocks can be generated by TERT-induced autophagy (27).
In summary, our findings suggest a direct effect of HIF-1α on TERT regulation in PTC cells. HIF-1α binds the promoter of TERT and transactivates TERT expression. Further, upregulation of TERT inhibited mTOR kinase activity, resulting in the activation of autophagy. Functionally, all of these processes provide energy and building blocks to PTC cells during hypoxia stress and to promote tumor progression. This study provides experimental evidences for the role of HIF-1α, TERT and autophagy in PTC progression, as well as the regulatory mechanism. These findings may result in novel therapeutic approaches being developed for the treatment of aggressive PTCs.
Footnotes
Acknowledgments
The authors thank Asia-Vector Biotechnology Co. Ltd (Shanghai, China) and Genomeditech Co. Ltd (Shanghai, China) for their technical support.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was sponsored by the National Natural Science Foundation of China (no.: 81501504), Shanghai Key Discipline of Medical Imaging (no.: 2017ZZ02005), and Shanghai Key Clinical Specialty (no.: shslczdzk03203).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4