
Abstract
Background:
Although studies have reported an increased risk for cognitive disorders in Hashimoto's thyroiditis (HT) patients, even in the euthyroid state, the mechanisms involved remain unclear. The hippocampus is a classic brain region associated with cognitive function, among which the formation of long-term potentiation (LTP) in the Schaffer collateral-CA1 pathway plays an important role in the process of learning and memory. Therefore, this study established a euthyroid HT model in mice and investigated whether and how HT itself has the ability to trigger LTP alterations accompanied by learning and memory abnormality.
Methods:
An experimental euthyroid HT model was established in NOD mice through immunization with porcine thyroglobulin (Tg). Morris water maze was measured to determine mice spatial learning and memory. We investigated the effect of HT on synaptic transmission and high-frequency stimulation-induced LTP in the Schaffer collateral-CA1 synapse of mice hippocampus in vivo. Then, animals were sacrificed for thyroid-related parameter measure as well as detection of cellular and molecular events associated with the induction of LTP.
Results:
HT mice showed intrathyroidal lymphocyte infiltration and rising serum thyroid autoantibody levels accompanied by normal thyroid function. The HT mice had poorer performance in Morris water maze than controls. These alterations were mirrored by abnormalities in synaptic plasticity in the Schaffer collateral-CA1 synapses of the hippocampus in vivo. The integrity of the synaptic structure is the premise for the production of LTP. As detected by transmission electron microscopy, the ultrastructure of synapse and astrocyte in the hippocampus were impaired in euthyroid HT mice. Additionally, Western blot and real-time polymerase chain reaction analyses confirmed that in HT mice, GS, GLAST, and GLT-1, key elements in glutamate–glutamine circulation located in astrocyte, were downregulated, accompanied by elevated levels of glutamate in the hippocampus, which impaired the material basis for LTP induction. NMDR2B expression in the hippocampus was also downregulated.
Conclusion:
HT can induce damage of LTP in the hippocampal Schaffer collateral-CA1 pathway in the euthyroid state, and this can be attributed, at least partly, to astrocytes impairment, which may underlie the deleterious effects of HT itself on hippocampal-dependent learning and memory function.
Introduction
Hashimoto's thyroiditis (HT) is a common autoimmune thyroid disease. It is characterized by lymphocyte infiltration in the thyroid gland along with rising antithyroglobulin (anti-Tg) antibody and antithyroid peroxidase (anti-TPO) antibody, which progressively damage the thyroid tissue, and is the main cause of hypothyroidism in areas with sufficient iodine intake (1,2). However, the clinical progression of the disease is slow. In the early stage, only positive antithyroid antibodies can be detected without any thyroid hormone disorders (3). Available data indicate that 79.3% of HT patients present with normal thyroid hormone levels at the initial diagnosis and may only have positive antithyroid antibodies and/or diffuse goiter (4). It is well documented that hypothyroidism can lead to neuropsychological deficits, including depression, anxiety, and cognitive impairment (5 –8). Only in recent years, several clinical studies have reported an association between cognitive impairment and HT, even in euthyroid population (9 –11). Our previous study (12) found no significant ultrastructural damage in the neurons in HT mice in the frontal lobe, which is an important cognition-related brain region. Also, Quinque et al. (13) used multimodal magnetic resonance imaging to evaluate neuronal activity in cognition-related brain regions of HT patients with normal thyroid function and found no significant difference between the HT and control groups. These findings suggest that euthyroid HT-related cognitive impairment appears to have little to do with neuronal damage and is independent of thyroid hormone levels. Therefore, the mechanism responsible for neurological dysfunction in the context of euthyroid HT remains unknown.
It is well known that the hippocampus is a classic “learning and memory” brain region, where neural plasticity known as long-term potentiation (LTP) was first discovered, and this response process is recognized as one of the main mechanisms for storing memories in the brain (14). There are three major excitatory synaptic pathways related to learning and memory in the hippocampus, among which the formation of LTP in the Schaffer collateral-CA1 pathway plays an important role in the process of learning and memory (15). Researchers generally believe that the production of LTP in this synaptic pathway requires the excitatory neurotransmitter glutamate to activate the postsynaptic NMDA glutamate receptor, which then causes a series of changes in signaling molecules and signal transduction pathways inside and outside the cell (16,17). LTP impairments are found in all neurological diseases associated with cognitive impairment in clinic, such as Parkinson's disease (18), Alzheimer's disease (19), and depression (20,21). The establishment of animal model provides opportunities for us to study the influence of HT on learning and memory from LTP in the Schaffer collateral-CA1 pathway.
Tg-induced thyroiditis is a classic model for studying HT and can be established in susceptible mice, such as NOD mice (22 –24). The induction leads to the development of HT-like illness characterized by lymphocyte infiltration in the thyroid gland and presence of autoantibodies against Tg and TPO, which are serological hallmarks of HT. Thus, this model has been widely used to explore the pathogenesis of HT and test therapeutics. The purpose of this study was to use behavioral test program and electrophysiological test to observe the effect of HT itself on hippocampal-dependent learning and memory function in a euthyroid HT mice model. The changes of cell morphology and molecular biology in the hippocampus can be observed by transmission electron microscopy (TEM), immunohistochemistry (IHC), and high-performance liquid chromatography (HPLC), to explore the possible mechanism of HT itself on learning and memory under normal thyroid function.
Materials and Methods
Animals
Female NOD mice (6–7 weeks old; 18–20 g) were obtained from Model Animal Research Center of Nanjing University (Nanjing, China) and randomly assigned into two groups: the control group (Con group, n = 20) and the HT group (HT group, n = 20). These mice were housed in cohorts of three to four under standard laboratory conditions (23°C ± 2°C, a 12-hour light/12-hour dark cycle, 55% ± 5% humidity) and had ad libitum access to tap water and rodent chow. All animal procedures were performed in accordance with the protocols approved by the Institutional Animal Care and Use Committees of Anhui Medical University.
Immunization and experimental design
After 7 days of acclimatizing, mice in the HT group were challenged with 25 μg porcine Tg (Sigma–Aldrich, St Louis, MO) emulsified in complete Freund's adjuvant (Sigma–Aldrich) injected subcutaneously at the base of the tail, and with a booster injection of an equal dose of Tg in incomplete Freund's adjuvant (Sigma–Aldrich) performed 14 days later. Control mice were injected with phosphate-buffered saline (PBS) in adjuvant at the same time intervals as porcine Tg-injected animals. Four weeks after the second challenge, the behavioral and biochemical parameters of all mice were evaluated (Supplementary Fig. S1).
Morris water maze
Behavioral test was performed to examine the cognitive states in animals. Mice were taken to the test room 60 minutes before the test. Behavioral procedures were conducted between 08:30 and 12:00 hours in a dim and quiet room. The observers were blinded to the experimental design. During the test, all animals were tracked and recorded by ANY-Maze™ Video Tracking Software (Stoelting Co.) with a digital camera. The Morris water maze consists of a black circular swimming pool divided into four quadrants of equal area: I, II, III, and IV. The pool was filled with edible melanin diluted with water (20°C ± 2°C). A circular platform (diameter 10 cm) was located 1 cm below the water surface in the middle of the II quadrant. All the mice (HT group: n = 10; Con group: n = 10) swam to accommodate the water maze and platform on the first day. Training began from the second day and continued till the fifth day, with each mouse undergoing four trials each day. In each trial, mice were placed into the water and allowed to search for the platform with a time limit of 60 seconds, and the time spent by the mice finding the platform was recorded and assigned as escape latency time. After staying on the platform for 10 seconds, mice were gently picked up, returned to the cage, and dried under a heat lamp (orientation navigation test). On the sixth day, the platform was removed, the mice were put into the water, and the time taken by each mouse to reach the place where original platform was located within 60 seconds was recorded (spatial probe test). Each mouse swam twice.
In vivo electrophysiology
The excitatory postsynaptic field potentials (fEPSPs) were recorded as the CA1 area of the hippocampus of anesthetized (urethane 1.2 g/kg, i.p.) mice. The mice (Con group: n = 6; HT group: n = 6) were placed in a stereotaxic apparatus (SR-5M; Narishige, Inc.), and holes were drilled in appropriate regions of the skull. To record the field potential response, a concentric bipolar stimulating electrode was placed in the CA3 area of the left hippocampus [anteroposterior (AP), 2.2 mm; mediolateral (ML), 1.2 mm; dorsoventral (DV), 1.0–1.5 mm] and a glass capillary recording electrode, filled with 1% Fast Green in 2 M NaCl, was placed in the CA1 of the left hippocampus (AP, 1.5 mm; ML, 2 mm; DV, 1.0–1.5 mm) (25). Input/output curves were generated by systematic variation of the stimulus current (0.1–1.4 mA) to evaluate synaptic potency. Stimulus pulses were delivered at 0.05 Hz, and three responses at each current level were averaged. After recording, the mice were given a 20 minute rest. The stimulus intensty selected for baseline measurements was adjusted to yield about 50% of its maximal amplitude to test fEPSPs. Baseline recording was obtained for 20 minutes, followed by application of the high-frequency stimulation (HFS: 200 Hz) and post-tetanic recording period of 2 hours with single pulse.
Tissue preparation
Mice were deeply anesthetized and randomly sacrificed in the morning (09:00–11:30 am). Blood, the thyroid, and the brain were collected immediately. Blood samples were centrifuged for measuring serological parameters. The thyroids were collected for histopathology. The brains were carefully dissected on ice and randomly assigned for later assay: right hippocampus for electrochemiluminescence immunoassay (ECLIA), enzyme-linked immunosorbent assay (ELISA), and TEM and left hippocampus for IHC, terminal deoxynucleotidyl transferase-mediated dUTP-biotin nick end labeling (TUNEL) staining, and real-time reverse transcription PCR (RT-PCR).
Electrochemiluminescence immunoassay
Serum samples were kept at −80°C until use. Serum triiodothyronine (T3), thyroxine (T4), Tg-Ab, and TPO-Ab concentrations were determined by ECLIA on Cobas e411 analyzer (Roche, Germany). The concentration of T3, T4, Tg-Ab, and TPO-Ab in the supernatant of animal hippocampal homogenate was also determined by this method. In addition, the hippocampus tissues were homogenized in PBS at a ratio of 1:9 (weight to volume) and then centrifuged at 12,000 g for 20 minutes at 4°C. The supernatants were collected for an analysis of protein content using the BCA method and then for the detection of thyroid autoantibody levels. Data are expressed as international units per milligram protein of brain issue.
Enzyme-linked immunosorbent assay
ELISA was used to determine the thyrotropin (TSH) level in serum according to the manufacturer's instructions (TSH; USCN Life Science, Inc., Wuhan, China). Data are expressed as picograms per milliliter of serum.
Hematoxylin and eosin
For histopathology, fresh thyroid tissues were fixed and processed for hematoxylin and eosin. From each animal, five noncontiguous coronal sections were used to examine thyroid histopathology. Histological evaluation for thyroiditis was performed according to lymphocytes infiltration extent in accordance with a previous study (26) and scored as: 0 = absence of infiltrate; 1 = interstitial accumulation of inflammatory cells around one or two follicles; 2 = one or two foci of inflammatory cells reaching the size of a follicle; 3 = 10–40% of inflammatory cell infiltration; 4 = greater than 40% inflammatory cell infiltration. The final score was the arithmetic mean of the five sections. A score below 1 was considered as pathologically not significant. The histological scores were evaluated and averaged by two investigators blinded to the experimental design.
Immunofluorescence
Sections were probed with mouse glial fibrillary acidic protein (GFAP) antibody (1:1000; Bioss, Beijing, China) as primary antibodies. The appropriate fluorophore-conjugated secondary antibodies were purchased from Servicebio, Inc. (Wuhan, China), followed by DAPI staining (Servicebio, Inc.). Samples were scanned on a Nikon Eclipse fluorescence microscope. Astrocytes were visualized using antibodies against GFAP, a marker for astrocyte cytoskeleton. In each section, three nonoverlapping fields were randomly selected in the hippocampus at magnification × 100. Astrocyte number was identified by quantifying the number of GFAP-labeled cells.
Immunohistochemistry
The fixed right hemispheres were embedded in paraffin and sectioned coronally with a microtome into 6 μm thick sections. From each mouse, five sections (1 of every 20 serial sections) of the dorsal hippocampus were selected to be mounted on polylysine-coated slides. Sections were dewaxed and hydrated, and antigen retrieval was performed. Endogenous peroxidase was quenched, and sections were blocked in serum. Sections were labeled with the appropriate primary and secondary antibodies. The primary antibody was mouse GFAP antibody (1:1000; Bioss), which was chosen for immunostaining for its specificity to astrocyte cytoskeleton. An image analysis system was used for quantitative analysis. The system includes MetaMorph image acquisition and processing software (JADA 801D) and a Nikon 80i microscope (Nikon, Tokyo, Japan) equipped with a HP computer. In addition to cell counting, GFAP immunoreactivity was analyzed by determining the percentage of GFAP-stained area as described previously (27).
Transmission electron microscopy
TEM was performed to evaluate the alterations in synaptic density and morphology in mice. TEM procedures were performed as previously described (28). Synapses were imaged at × 20,000 and × 50,000 magnifications. Digital images at × 20,000 (15 images per animal) were used to evaluate postsynaptic density (PSD) (29). For the analysis of hippocampal synaptic density, the number of synapses per unit volume of tissue (Nv) was counted on the National Institutes of Health ImageJ software (RRID: SCR_003070) and the numbers were assessed using the stereological equation Nv = 8ENa/π2, as previously described (30), where E designated the mean of the reciprocal values of the observed PSD lengths, and Na was synapse numbers per unit test area. In addition to synapse counting, we randomly selected 120 synapses from each group for analysis of synaptic ultrastructure. Only synapses with clear presynaptic and postsynaptic properties were assigned for the latter assay. For each condition, active zone length and PSD thickness were measured according to previous criteria described by Lenselink et al. (31). In addition, observations of neurons were made using magnifications ranging from 8000 × to 12,000 × , and the observations of astrocytes used magnifications ranging from 15,000 × to 30,000 × .
HPLC-tandem mass spectrometry
Quantitative analysis of glutamate was performed using a Waters ACQUITY HPLC system coupled with a TSQ Quantum equipped with an electrospray ionisation source in the positive mode operating in select response monitoring mode. Water containing 0.1% formic acid/10 mM ammonium acetate aqueous solution (A) and acetonitrile (B) were used as the mobile phase. A 1 μL of sample solution was injected into a Welch Ultimate XB-C18 HPLC column (4.6 × 150 mm, 3 μm) at a flow rate of 1.0 mL/min. The following gradient program was used: 0–1.9 minutes, 10% B; 1.9–2.5 minutes, 55% B; 2.5–3.5 minutes, 55–95% B; 3.5–4.0 minutes, 95% B; 4.0–4.1 minutes, 95–10% B; 4.1–5 minutes, 10% B. The parameters in mass spectrometry analysis were as follows: capillary temperature, 400°C; vaporizer temperature, 450°C; spray voltage, 4.0 kV (positive). High purity nitrogen was used as sheath and auxiliary gas; high purity argon was used as collision gas.
TUNEL staining
Apoptotic neurons in the hippocampus were visualized using a TUNEL kit (Roche, Basel, Switzerland) following the manufacturer's protocol. Serial coronal sections from the left hippocampus were dewaxed in xylene and rehydrated through gradient ethanol. After rinsing with PBS, the sections were incubated with proteinase K solution at 37°C for 20 minutes. Then, they were incubated at 37°C for 2 hours with TUNEL reaction mixture in a dark humidified chamber. Following five PBS washes, the sections were stained with DAB and hematoxylin at room temperature for 10 minutes and slides were coverslipped. TUNEL-positive signals in the hippocampus were examined using a microscope (Nikon Eclipse C1).
RNA purification and real-time RT-PCR
Total RNA was extracted from the hippocampus using TRI reagent (Invitrogen, Carlsbad, CA) and treated with RNase-free DNase, followed by reverse transcription with avian myeloblastosis virus (Promega, Madison, WI) according to the manufacturer's guidelines. The primers used for polymerase chain reaction are listed as follows: GFAP: FP—CGG AGA CGC ATC ACC TCTG, RP—TGG AGG AGT CAT TCG AGA CAA; GLAST: FP—GCC CAC AGA TGA CAT CAC AC, RP—GGA CAA GTG CTC GAC AAT CC; GLT-1: FP—CAA TGT GGT GGG CGA TTC TT, RP—ACT GCG TCT TGG TCA TTT CG; GS: FP—GGG TGA GAA AGT CCA AGC CA, RP—ACA TGG CAA CAG GAT GGA GG; NMDA2B: FP—GGG AGA GGG TGG GAA AAT GG, RP—GGT GCC TCC TCC AAG GTA AC. The analysis method used in this experiment was Relative Quantification Study, and the calculation method was 2−▵▵Ct.
Western blot
Total protein was isolated from snap-frozen tissue homogenized in radioimmunoprecipitation assay buffer. Protein concentration was determined using the Pierce BCA Protein Assay kit (ThermoFisher Scientific, Shanghai, China) according to the manufacturer's protocol, and 40 μg of total protein was fractionated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (Solarbio, Beijing, China) and transferred to the polyvinylidene fluoride membrane (Millipore, Burlington, MA). The following primary and secondary antibodies were used: GLAST antibody (1:500; Abcam, Cambridge, MA), GLT-1 antibody (1:1000; Abcam), GFAP antibody (1:1000; Bioss), GS antibody (1:1000; Abcam), NMDAR2B antibody (1:250; Abcam), β-actin antibody (1:1000; Zs-BIO, Beijing, China).
Statistical analyses
All analyses were conducted using OriginPro 8.0 for Windows. The results are expressed as mean ± standard error of the mean. Thyroiditis scores were compared using the Mann–Whitney U-test. The chromatogram collection and integration of Glu was processed by software Xcalibur 3.0 Thermo, and linear regression was performed with a weighted coefficient of 1/X. The other data were compared by one-way analysis of variance (ANOVA) using least significant difference for post hoc analysis. Following ANOVA, the Student–Newman–Keul's post hoc analysis was used for pairwise comparisons of data. p Value <0.05 was considered significant.
Results
Building a euthyroid HT model in mice
There were no significant differences in the body's and hippocampal weight of HT mice compared with the control ones, suggesting that euthyroid HT did not affect the growth and development of mice (Supplementary Table S1).
As depicted in Figure 1A, histological examination showed that control mice had intact thyroid follicles with an even distribution, and lymphocyte infiltration was hardly found in thyroid tissues. In contrast, HT mice had destruction of thyroid follicles and lymphocyte infiltration in thyroid tissues. The scoring for thyroiditis in HT mice was significantly greater than that in the Con group (Fig. 1B). Serum concentrations of thyroid autoantibodies (anti-TPO and anti-Tg) in the HT group were higher than those in the Con group with no difference in serum T3, T4, or TSH levels between groups (Fig. 1C–G). Moreover, the concentration of anti-Tg in the hippocampus was significantly higher in HT mice than in the controls (Supplementary Fig. S2C). As well, there was a similar, although not significant, tendency for hippocampal anti-TPO levels between groups (p = 0.09) (Supplementary Fig. S2D). No significant difference in hippocampal T3 and T4 levels (Supplementary Fig. S2A, B and Supplementary Table S2) was detected in the two groups. Taken together, these findings indicated that a euthyroid HT model was successfully established in mice. Since NOD mice spontaneously develop autoimmune diabetes, we examined serum glucose levels in all mice and no significant difference was detected in serum glucose levels between control mice and HT mice (5.50 ± 0.55 vs. 6.31 ± 0.52 mmol/L, p = 0.30, n = 10 per group), consistent with a previous study (32). Considering a nonspecific inflammatory response may be induced by the adjuvant itself in the Con group, we measured the concentrations of IL-6, TNF-α, and IFN-γ in the hippocampal supernatant of the two groups of mice, and no significant difference was detected (IL-6: Con 46.57 ± 4.13, HT 47.75 ± 4.25, p = 0.84; TNF-α: Con 2.80 ± 0.21, HT 2.82 ± 0.22, p = 0.95; IFN-γ: Con 15.55 ± 1.11, HT 15.00 ± 1.02, p = 0.72; in pg per mL of hippocampal supernatant, n = 6 per group).
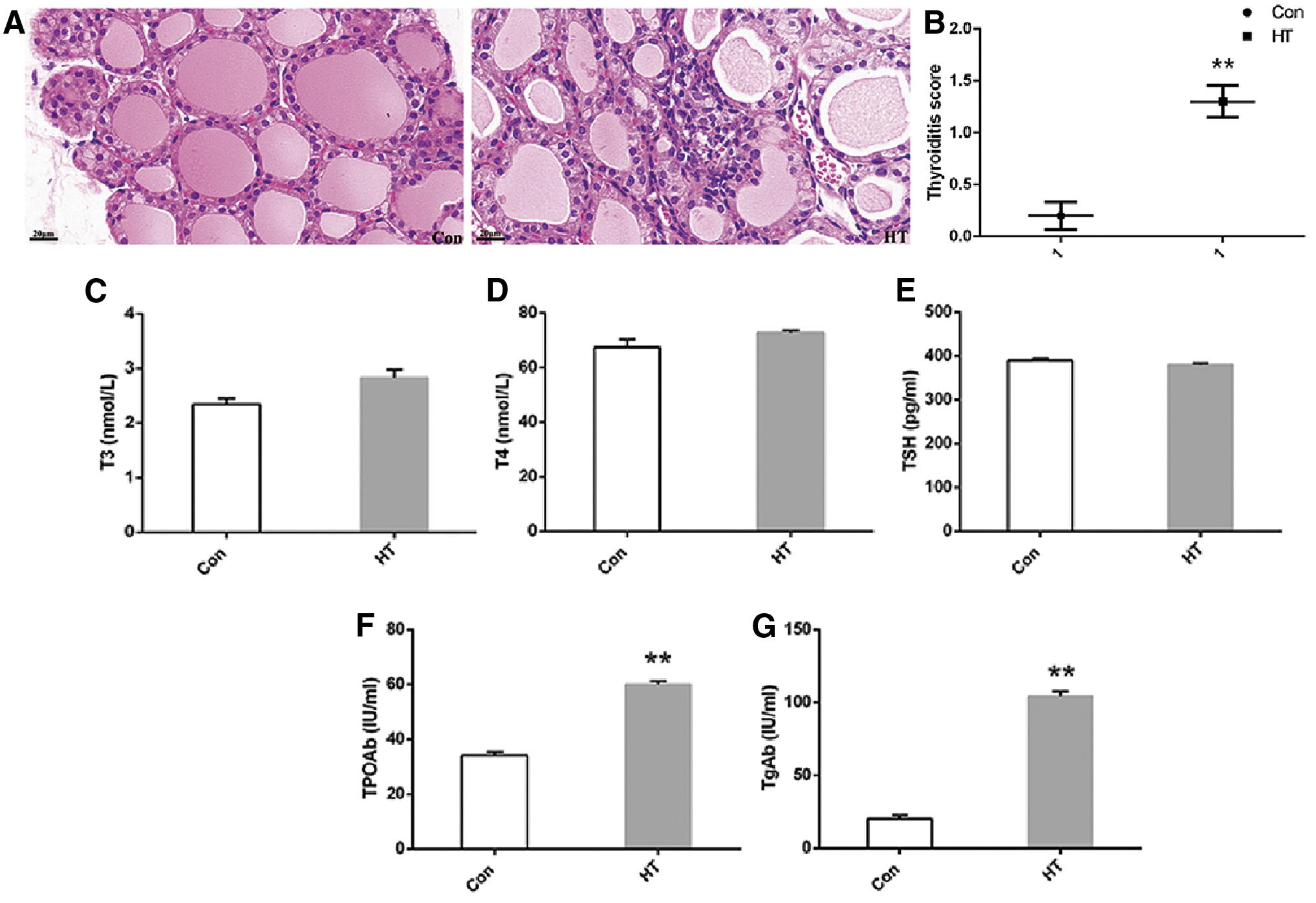
Histological evaluations in euthyroid HT model in mice. (
HT induces hippocampus-dependent learning and memory dysfunction
The escape latency time of two training groups became shorter as training days increased. Meanwhile, there was no statistically significant difference between the Con group (n = 10) and the HT group (n = 10) in swimming speed (p > 0.05). Compared with the Con group, the mean escape latency and swimming distance in the HT mice were prolonged in orientation navigation test (Fig. 2A, B), and in the spatial probe test, and the percentage of distance and time in the target quadrant decreased in the HT group (Fig. 2D, E). The results suggest that the mice in the HT group have learning and memory dysfunction.
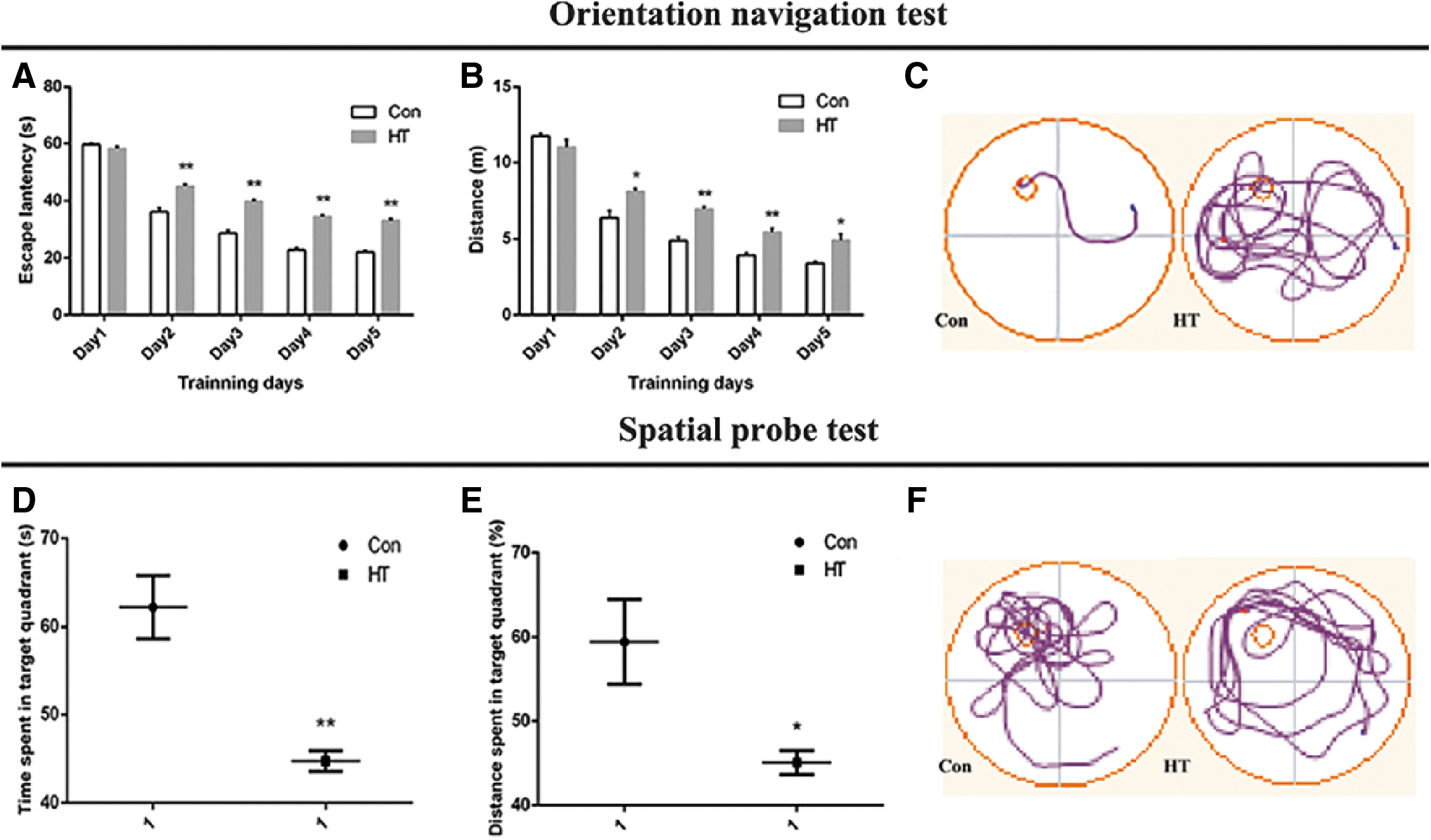
Euthyroid HT induces poor performance in Morris water maze in mice. (
In the first series of experiments, we determined whether euthyroid HT modulates basal synaptic transmission at the Schaffer collateral-CA1 synapse in vivo. We stimulated the Schaffer collateral and recorded the evoked fEPSP in the CA1 region of mouse hippocampus. After establishing a stable baseline for 20 minutes, the Con group (n = 6) and the HT group (n = 6) were measured for 100 minutes. The HT group revealed a significant decrease in the slope of fEPSP (Fig. 3A). The fEPSP slope (mV/mseconds) in the HT group was 0.82 ± 0.08 compared with 1.06 ± 0.09 in the Con group. These results indicate that euthyroid HT can significantly suppress basal synaptic transmission at the Schaffer collateral-CA1 synapse in vivo.
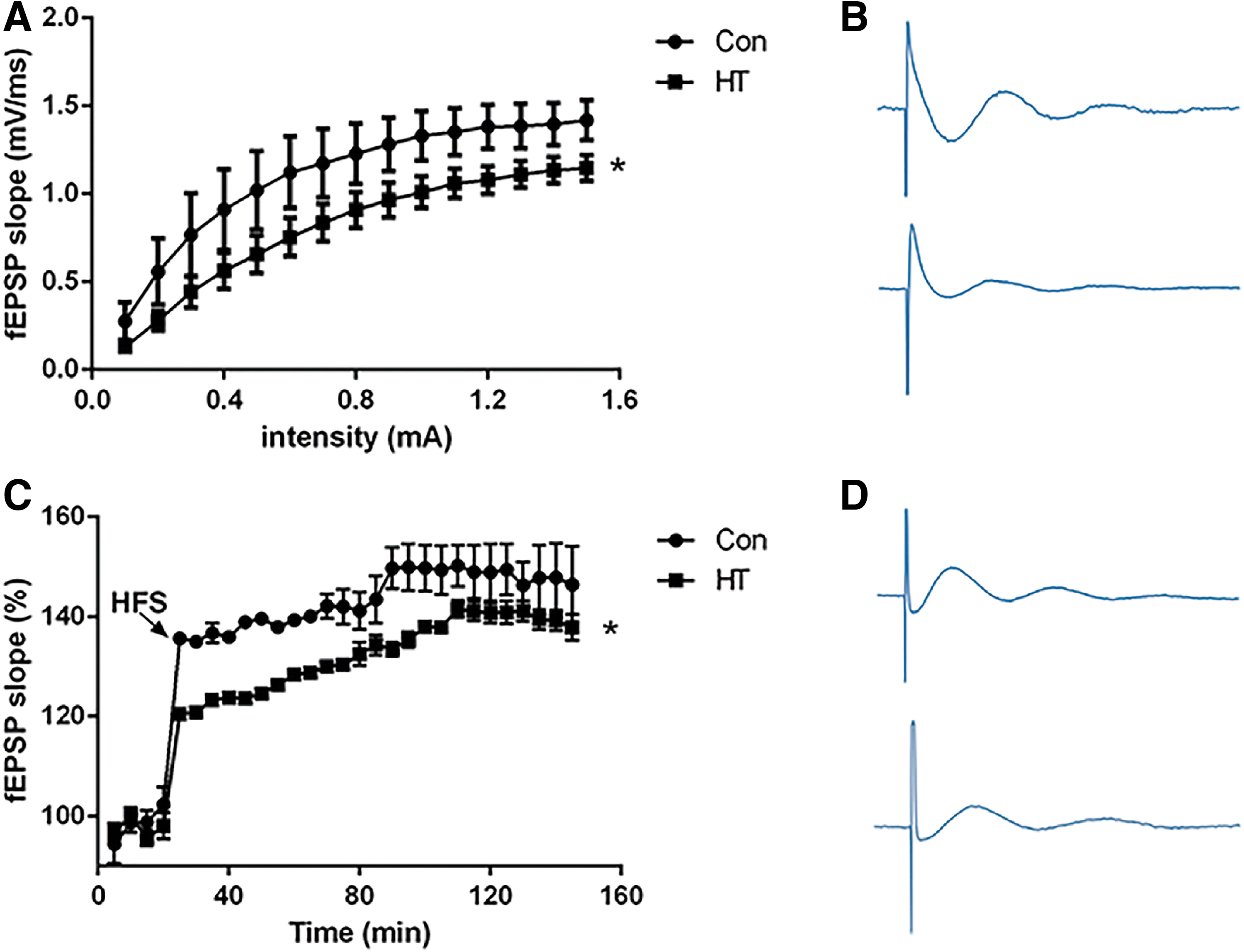
Euthyroid HT has an inhibitory effect on synaptic plasticity of the hippocampal Schaffer collateral-CA1 pathway. (
Next, we determined whether euthyroid HT regulates the induction of LTP at Schaffer collateral-CA1 synapse in vivo. As shown in Figure 3C, application of HFS resulted a marked increase in the amplitude of fEPSP at Schaffer collateral-CA1 synapses in control rats during the 120 minute recording period (fEPSP amplitude was 152.70% ± 11.63% of baseline values at 90 minutes after HFS application, n = 6). While in the HT group, the induction of HFS-induced LTP was inhibited, and this inhibitory effect was maintained without declining throughout the recording period. At 90 minutes after HFS stimulation, the fEPSP amplitude was 136.54% ± 5.47% (n = 6) of baseline value (p < 0.05 compared with the Con group). These results suggest that euthyroid HT can block the induction of LTP at the Schaffer collateral-CA1 synapse in mouse hippocampus in vivo.
HT induces ultrastructural changes in hippocampal synapses
To examine the effect of HT on hippocampal synapses, synapse morphometry was examined using TEM. Con and HT groups showed significant differences in synaptic density and interface parameters (Fig. 4A–F). Quantitatively, the ultrastructure of the hippocampal synapses revealed lower synaptic density (Fig. 4G), shorter active zone length (Fig. 4H), thinner PSDs (Fig. 4I), and decreased synaptic curvature (Fig. 4J) in the HT mice compared with those in the control mice. The other synaptic interface parameters, such as synaptic cleft, did not show differences between groups (Fig. 4K). The morphological data suggest that euthyroid HT could induce synaptic loss and impair synaptic ultrastructure in the hippocampus regions of mice.
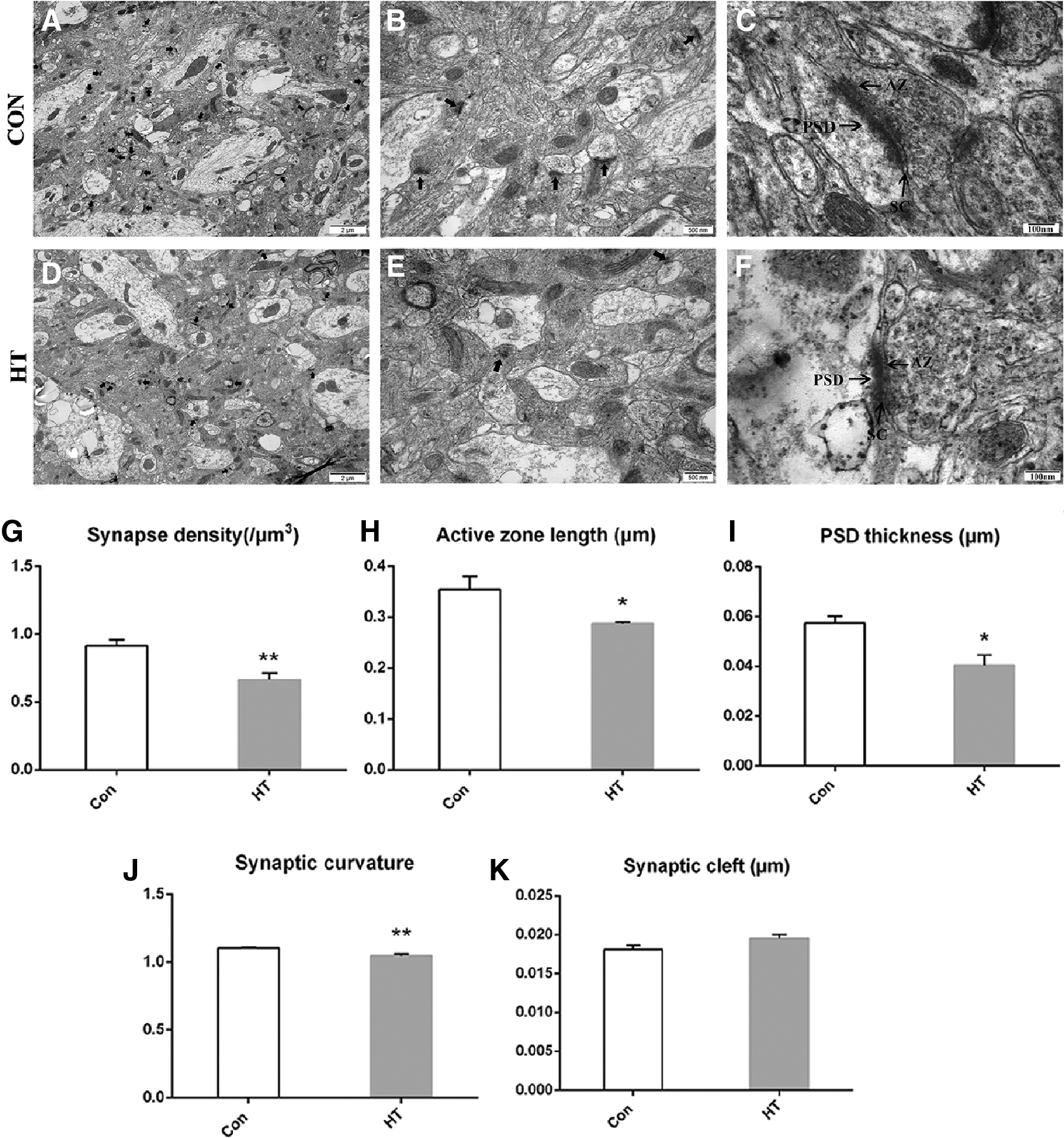
Euthyroid HT induces ultrastructure changes in hippocampal synapses. (
HT does not induce neuronal apoptosis in the hippocampus
In this study, we performed TUNEL staining and TEM to identify neuronal apoptosis in the hippocampus. As shown in Supplementary Figure S3, ultrastructure of hippocampal neurons in HT mice was similar to control mice with the nucleus with intact nuclear membranes and evenly distributed chromatin. In addition, we did not observe any changes in TUNEL-positive neurons in the hippocampus between groups. There were no apoptotic features, such as chromatin margination or nuclear condensation, in the neurons examined.
HT induces impaired astrocyte in the hippocampus
We examined the astrocytes in the hippocampus of mice from both groups by using IHC, immunofluorescence (IF), real-time RT-PCR, Western blot, and TEM.
Representative images of GFAP (astrocyte marker) staining are shown in Figure 5A and B. We found that there was a decrease in astrocytes in HT mice compared with that in the Con group, as demonstrated by the lower percentages of GFAP-stained areas (Fig. 5C) and the fewer numbers of GFAP-positive cells (Fig. 5D) in the captured photographs. These immunohistochemical and IF results demonstrate that euthyroid HT induces astrocyte loss in the hippocampus. A quantitative analysis of GFAP messenger RNA (mRNA) and protein levels in the hippocampus (Fig. 5E–G) confirm these results.
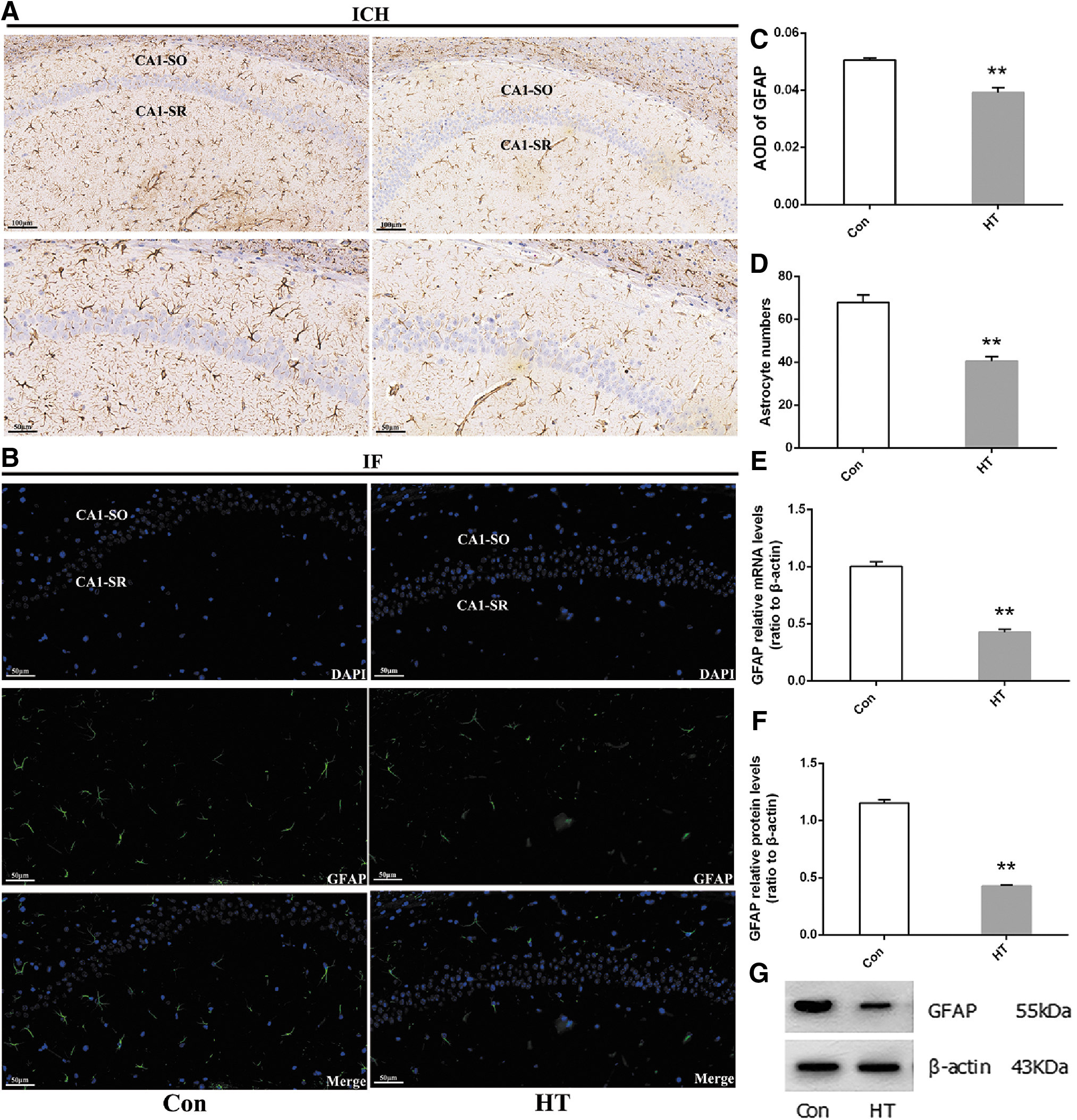
Euthyroid HT induces astrocyte loss in the hippocampus. IHC and IF were performed to assess the condition of astrocytes in animals. (
We also observed the ultrastructural features of astrocytes by using TEM (Supplementary Fig. S4). In HT mice, impaired ultrastructure was observed in the hippocampal astrocytes. The main manifestations were lumpy condensation of heterochromatin in the nucleus and vacuolar changes in the cytoplasm, accompanied by swelling of mitochondria and expansion of the endoplasmic reticulum. According to ultramicropathology, these results suggest that HT induces ultrastructural damage to the astrocytes in the hippocampus.
HT induces dysfunction in hippocampal glutamate–glutamine circulation between astrocytes and neurons
Since our data clearly showed astrocyte dysfunction in the hippocampus of HT mice, we further extended the work to study glutamate–glutamine circulation, which is an important form of communication between astrocytes and neurons.
In the astrocytes, GS is a key enzyme in the glutamate–glutamine circulation. Meanwhile, GLT-1 and GLAST are responsible for the uptake and clearance of redundant glutamate in the synaptic cleft, which are also mainly located in astrocytes. As expected, the levels of GLT-1, GLAST, and GS measured by RT-PCR and Western blot showed a significant reduction in the hippocampus of HT mice compared with that in controls (Fig. 6). In line with this, the content of glutamate was increased in HT mice compared with that in controls (Supplementary Fig. S5). The downregulation of NR2B expression may be to offset the increased excitatory toxicity of glutamate to neurons. Thus, HT impairs the communication between astrocytes and neurons.
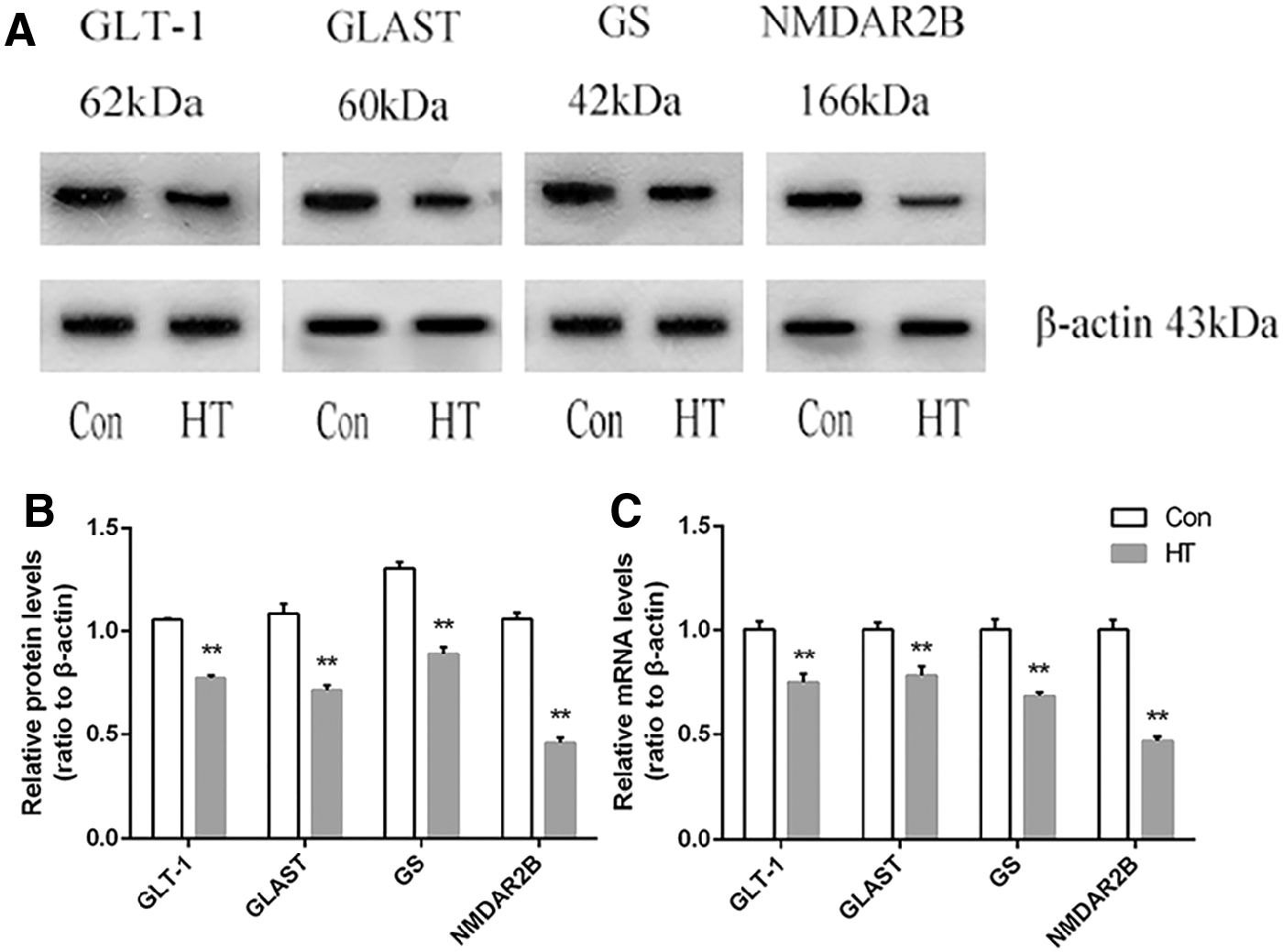
Euthyroid HT induces dysfunction in hippocampal glutamate–glutamine circulation between astrocytes and neurons. (
Discussion
In this study, mice immunized subcutaneously with Tg showed intrathyroidal lymphocyte infiltration and elevated serum anti-Tg antibody and anti-TPO antibody. The levels of T3, T4, and TSH were normal, mimicking HT patients with euthyroid function in clinic. The body weight, hippocampal weight, and hippocampal coefficient of HT mice were not significantly affected, suggesting that HT itself did not affect the normal growth of mice.
Hypothyroidism is known to cause neuropsychological deficits, including depression, anxiety, and cognitive impairment (5 –8). However, the literature on the effects of euthyroid HT on cognitive function remains controversial. Some have shown that compared with the Con group, HT patients have sustained attention impairment (10) and poorer performance in MMSE (mini-mental state examination) test, speech fluency test, attachment test, and MoCA (Montreal cognitive assessment) score even in normal thyroid function (11,33). However, studies using cognitive behavior scale and multimodal magnetic resonance imaging to evaluate the brain function of euthyroid HT patients observed no evidence of cognitive impairment (13). In this study, we first used behavioral experiments to evaluate whether euthyroid HT mice had learning and memory dysfunction. Morris water maze experiment is a classic method to evaluate the ability of hippocampal learning and spatial memory by forcing experimental animals to look for the underwater hiding platform (25). In this study, compared with the Con group, mice in the HT group spent more time in finding the hidden platform and less time in the target quadrant. This suggests that HT itself can induce hippocampal-dependent learning and memory dysfunction regardless of thyroid functions in mice. Our study provides preliminary evidence to support the clinical literature linking euthyroid HT to increased cognitive impairment.
To further support the behavioral test results, we also used the field potential method to record the changes in synaptic plasticity in the hippocampal Schaffer collateral-CA1 pathway, analyzing the alteration in learning and memory function in the two groups. LTP is one of the important manifestations of synaptic plasticity and is considered to be the main mechanism that constitutes the basis of learning and memory (34). The results suggest that HT has an inhibitory effect on the basic synaptic transmission efficiency in the hippocampal Schaffer collateral-CA1 pathway and leads to a decrease in the amplitude of LTP in this pathway even in normal thyroid function. This is consistent with our behavioral results. These positive results prompt us to further explore the possible mechanisms by which HT itself affects brain function.
We investigated first the factors that can affect the formation of LTP in the hippocampal Schaffer collateral-CA1 pathway. The lateral branches of axons from pyramidal cells in hippocampal CA3 region, known as Schaffer branches, pass through the CA3 to the CA1 region and form synapses with the dendritic branches of pyramidal cells in the CA1 region. This synaptic pathway is the structural basis for the generation of LTP, and the integrity of the synaptic structure is the premise for the production of LTP (35). Through TEM, we found a decreased synaptic density, reduced active zone length, PSD thickness, and synaptic curvature in the hippocampus at the ultrastructural level in the HT mice. The changes indicate that euthyroid HT induces synaptic loss and impairs synaptic structures.
Thyroid dysfunction can lead to structural damage to synapses, and our previous studies have highlighted that experimental hypothyroidism induces synaptic deficit in the hippocampus (8) and prefrontal lobe (28). However, in this study, there was no statistically significant difference in thyroid hormone levels in serum and hippocampal tissues in the HT group compared with the Con group, so we believe that the synaptic damage is independent of thyroid hormone levels.
So is the impaired synapses caused by damage or even death of neurons in the hippocampus? Interestingly, the ultrastructure of hippocampal neurons in HT and control mice was similar, and no features of neuronal apoptosis such as chromatin aggregation, nuclear shrinkage, and apoptotic corpuscle formation were observed in the neurons of the two groups. Further TUNNEL staining also supported that euthyroid HT did not cause neuronal apoptosis in the hippocampus. This suggests that HT itself does not cause structural damage to hippocampal neurons, which is consistent with the results of our previous findings showing a lack of neuronal apoptosis or ultrastructural damage in the frontal lobe of HT mice (12). Quinque et al. (13) used resting-state functional magnetic resonance imaging to find that the neuron activity in the hippocampus of HT patients with normal thyroid function was not different from that of the Con group, which is also consistent with our results. Thus, euthyroid HT might have less effect on neurons and the synapse loss is unlikely to be due to local neuronal cell damage in HT mice.
While using electron microscopy to observe the ultrastructure of neurons, we found that the structure of astrocyte in the hippocampus of the HT group was abnormal. The main manifestations were lumpy condensation of heterochromatin in the nucleus, few organelles in the cytoplasm, and even vacuolar changes in the cytoplasm, accompanied by swelling of mitochondria and expansion of the endoplasmic reticulum. Ultramicropathology suggests that when cells are subjected to various types of damage, their characteristic changes are nonspecific expansion and rupture of the endoplasmic reticulum. Our morphological results suggested that HT induced ultrastructural damage to the astrocytes in the hippocampus. Meanwhile, immunohistochemical staining of cytoskeletal-related proteins of astrocyte (GFAP) was used in this study to confirm that the number of GFAP-positive cells in the hippocampus of HT mice was low and the optical density value decreased, indicating that the number of astrocytes decreased.
Astrocytes are the most numerous glial cells in the central nervous system of mammals, and form the tripartite synapse with neurons and participate in synaptic plasticity, playing an important role in the synaptic formation, elimination, and neurotransmitter transmission (36,37). In vitro studies (38) revealed that the highly purified retinal ganglion cells in rodents only formed a few synapses without the influence of astrocytes, while the number of synaptic formation and synaptic activity increased significantly under the condition of astrocytes. Although spinal cord neurons in the medium with high levels of neurotrophic factors could survive, synaptic connections between them could be formed only in the presence of astrocytes (39). These results indicate that the presence of astrocytes in the formation of synapses is a common mechanism in the nervous system. However, our results showed that the number of astrocytes in the hippocampus of the HT group mice decreased that was accompanied with impaired ultrastructure. Therefore, we believe that the synaptic loss in euthyroid HT could be at least partially attributed to astrocyte damage.
If synaptic integrity is the structural basis for the generation of LTP in the hippocampal Schaffer collateral-CA1 pathway, then glutamate and NMDA receptors that induce LTP in this pathway can be said to be the material basis for the generation of LTP. Glutamate is an important excitatory neurotransmitter in mammalian central nervous system, which plays a key role in the maintenance of synaptic plasticity. However, under pathological conditions, glutamate can be over-released or insufficiently cleared, which causes a large amount of glutamate to accumulate in the synaptic cleft, leading to neuronal excitatory toxicity. Thus, glutamate is a double-edged sword, undergoing a transition from neurotransmitter to neurotoxin under certain adverse conditions (40).
There are various glutamate-scavenging mechanisms in the synaptic cleft, and they work together to ensure the normal synaptic transmission. The most important mechanism is the glutamate–glutamine cycle, which relies on glutamate transporters GLAST and GLT-1 in astrocytes. Therefore, maintaining the balance of glutamate in the synaptic cleft is the main way for astrocytes to regulate synaptic plasticity (41). We found impaired astrocytes in the hippocampus of HT mice and GLAST and GLT-1 levels in the hippocampus of the HT group decreased. Many neuropsychiatric disorders with cognitive impairment in clinic, such as Parkinson's disease (42), Alzheimer's disease (43), anxiety, depression, and schizophrenia (40) have been shown to be associated with the excitatory toxicity of glutamate, whose overaccumulation causes neuronal damage and even death. In this study, glutamate content was elevated but no structural damage or death of neurons was found, which is not surprising since synaptic dysfunction always occurs before neuronal damage or even cognitive impairment (37).
NMDA receptor is anionotropic glutamate receptor, which is closely related to learning and memory. In particular, its NR2B subunit plays an important role in the induction and maintenance of LTP. Therefore, Bliss named NR2B as “smart gene” (44,45). NR2B is mainly distributed in the hippocampus and highly enriched in PSD (46). We found that the NR2B level in the HT mice was lower than that in the Con group, which may explain the decreased PSD thickness we observed. This downregulation of NR2B expression may be due to homeostasis regulation mechanism that reduces synaptic NMDAR to counteract the increased glutamate excitatory toxicity to neurons (47).
There are some limitations in the present study. First, although the hippocampal Schaffer collateral-CA1 pathway plays an important role in learning and memory function, the changes of “PP-DG” pathway and “MF-CA3” pathway, the other two excitatory synaptic pathways in the hippocampus, still need to be studied. Second, in addition to glutamate, there are also a variety of neurotransmitters involved in the cognitive process in the central nervous system that need to be studied. Finally, this study mainly focused on astrocyte, which plays an important role in neurotransmitter transmission, the formation and elimination of synapses. The other two important glial cells in the brain, oligodendrocytes and microglia, need to be evaluated in future studies using the HT model.
Conclusion
In summary, our study used animal experiments to confirm that HT itself can lead to hippocampal-dependent learning and memory dysfunction in mice regardless of thyroid function, and the reason can be attributed at least partly to astrocytes impairment, which results in decreased synaptic number, impaired synaptic structure, dysfunctional glutamate–glutamine cycle in the hippocampus, and damage to LTP in the hippocampal Schaffer collateral-CA1 pathway.
Footnotes
Author Disclosure Statement
No competing financial interests exist for any author.
Funding Information
This study was supported by funding from the Natural Science Foundation of Anhui Province (No. 1908085QH339).
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5