
Abstract
Background:
The thyroid hormone (TH) metabolite 3,5-diiodothyronine (3,5-T2) is considered as a potential drug for treatment of nonalcoholic fatty liver disease (NAFLD) based on its prominent antisteatotic effects in murine models of obesity without the detrimental thyromimetic side effects known for classical TH. To expand our understanding of its mode of action, we comprehensively characterized the effects of 3,5-T2 on hepatic gene expression in a diet-induced murine model of obesity by a combined liver proteome and transcriptome analysis.
Materials and Methods:
Male C57BL/6 mice fed high-fat diet (HFD) to induce NAFLD or standard diet (SD) as control were treated with 2.5 μg/g body weight 3,5-T2 or saline for 4 weeks. We performed mass spectrometry analyses and integrated those proteome data with earlier published microarray-based transcriptome data from the same animals. In addition, concentrations of several sex steroids in serum and different tissues were determined by gas chromatography-tandem mass spectrometry.
Results:
We observed limited concordance between transcripts and proteins exhibiting differential abundance under 3,5-T2 treatment, which was only partially explainable by methodological reasons and might, therefore, reflect noncanonical post-transcriptional events. The treatment affected the levels of more and partially different proteins under HFD as compared with SD, demonstrating response modulation by the hepatic lipid load. The hepatic physiological signatures of 3,5-T2 treatment inferable from the omics data comprised the reduction of oxidative stress and alteration of apolipoprotein profiles, both due to decreased liver fat content. In addition, induction of several classical TH target genes and genes involved in the biosynthesis of cholesterol, bile acids (BAs), and male sex steroids was observed. The latter finding was supported by hepatic sex steroid measurements.
Conclusion:
While confirming the beneficial hepatic liver fat reduction by 3,5-T2 treatment, our data suggest that besides the well-known induction of fatty acid oxidation the stimulation of cholesterol- and BA synthesis with subsequent excretion of the latter through bile might represent a further important mechanism in this context. The obvious intensified male sex steroid exposition of the liver in 3,5-T2-treated HFD animals can be predicted to cause enhanced hepatic “masculinization,” with not yet clear but potentially detrimental physiological consequences.
Introduction
Overweight and obesity represent complex disorders that affect all social classes and age groups, cause diverse health problems with increasing prevalence (1), and, more specifically, are positively associated with the metabolic syndrome as well as the disease state of nonalcoholic fatty liver disease (NAFLD). The latter comprises a histological spectrum of liver pathologies ranging from NAFL over nonalcoholic steatohepatitis, often with fibrosis, to cirrhosis with its complications. NAFLD is based on an imbalance between hepatic lipid supply through fatty acid uptake and/or de novo lipogenesis, on the one hand, and hepatic lipid clearance through fatty acid oxidation or triglyceride-rich lipoprotein secretion, on the other hand (2,3).
There are numerous studies characterizing the antisteatotic effect of the thyroid hormone (TH) metabolite 3,5-diiodothyronine (3,5-T2) in diet-induced NAFLD. It is described to be metabolically active and to affect energy as well as lipid metabolism, thereby reducing the body weight under a high-fat diet (HFD). Remarkably, this beneficial effect was reported to occur without any accompanying detrimental thyromimetic side effects as known for classical THs when administered to rats after a specific dose regime (4 –7). Therefore, the endogenous TH metabolite 3,5-T2 represents an interesting candidate molecule for the treatment of obesity and associated diseases such as NAFLD. However, depending on the respective animal model and applied dose of the metabolite, negative effects on hypothalamus–pituitary–thyroid axis and cardiovascular system have also been observed in different studies (8 –12).
So far, only few studies have used a proteomic approach for analyzing the effects of 3,5-T2 in an animal model of HFD-induced fatty liver to obtain specific information regarding its hepatic mode of action as well as therapeutic potential (13,14). Therefore, in this study we performed a comparative state-of-the-art mass spectrometry analysis of the hepatic proteomes of male C57BL/6 mice fed either HFD to induce a fatty liver or standard diet (SD) as control. Both groups were treated with 2.5 μg/g body weight 3,5-T2 or with saline for 4 weeks. This approach allowed for the detection of differentially abundant proteins after treatment with 3,5-T2 in the two specific diet groups.
The protein level represents the final and, therefore, physiologically most relevant stage of gene expression. The fact that RNA prepared from the same liver tissue samples that were used for this proteome study was already used for microarray-based transcriptome analyses (15) allowed us to relate the proteome profiles in this study with the already generated corresponding transcriptome data. This gave us the opportunity to perform a comprehensive proteome-transcriptome analysis for the described study design.
Materials and Methods
Treatment of animals and liver sample collection for omics analyses
Male C57BL6/J mice kept with free access to water received SD or a high-fat diet (HFD) ad libitum for 20 weeks immediately after weaning. Afterward, mice were continued on SD or HFD and daily administered intraperitoneally with 3,5-T2 (2.5 μg/g body weight) in physiological saline solution or with the solvent control for 28 days. Mice were sacrificed and liver tissue was dissected, immediately snap-frozen in liquid nitrogen, and stored at −80°C until use for omics analyses. Details are described in the Supplemental Data.
All experimental procedures described followed established guidelines for the care and handling of laboratory animals and were approved by ethics committee of the State Agency of Environment, Health and Consumer Protection (State of Brandenburg, Germany).
Analysis of the hepatic proteome by nanoLC tandem mass spectrometry
Liver protein extracts were prepared using a TRIzol reagent-based extraction method with sample homogenization in a bead mill dismembrator as previously described (15). Samples were dissolved in urea/thiourea solution and protein concentrations were determined by Bradford assay. For three of the four experimental groups samples from four animals and for one group samples from three animals were used in the proteome analysis. The details and procedures for liver tissue processing, nanoLC tandem mass spectrometry, and data analysis are described in the Supplemental Data.
Sex steroid analysis by high-sensitive gas chromatography-tandem mass spectrometry
Progesterone, androstenedione, testosterone, and dihydrotestosterone (DHT) were assessed in liver, serum, and white adipose tissue (WAT, inguinal and gonadal) from 3,5-T2-treated mice fed HFD or SD and their respective control animals treated with saline as described in the Supplemental Data.
Microarray-based analysis of the hepatic transcriptome
Hepatic RNA extraction (n = 3 independent tissue samples per experimental group), RNA quality control, further processing of RNA samples, including hybridization with GeneChip Mouse Gene 1.0 ST Arrays (Affymetrix), as well as statistical analysis of the generated transcriptome data were as previously described by Lietzow et al. (15).
Results
3,5-T2-mediated alterations of the hepatic proteome in SD- and HFD-fed mice as determined by quantitative proteome analysis
Hepatic proteome analysis of male mice treated with 3,5-T2 or saline for 4 weeks detected 2407 and 2423 proteins in at least three animals fed SD and HFD, respectively, under at least one treatment. Of 2387 proteins detected under both diets, 432 were significantly differentially abundant. In SD and HFD mice, 89 and 134 proteins and 135 and 137 proteins exhibited increased and decreased abundance, respectively, under 3,5-T2 as compared with control (Supplementary Fig. S1), where 62 proteins exhibited the same alteration patterns in both diet groups (Fig. 1).
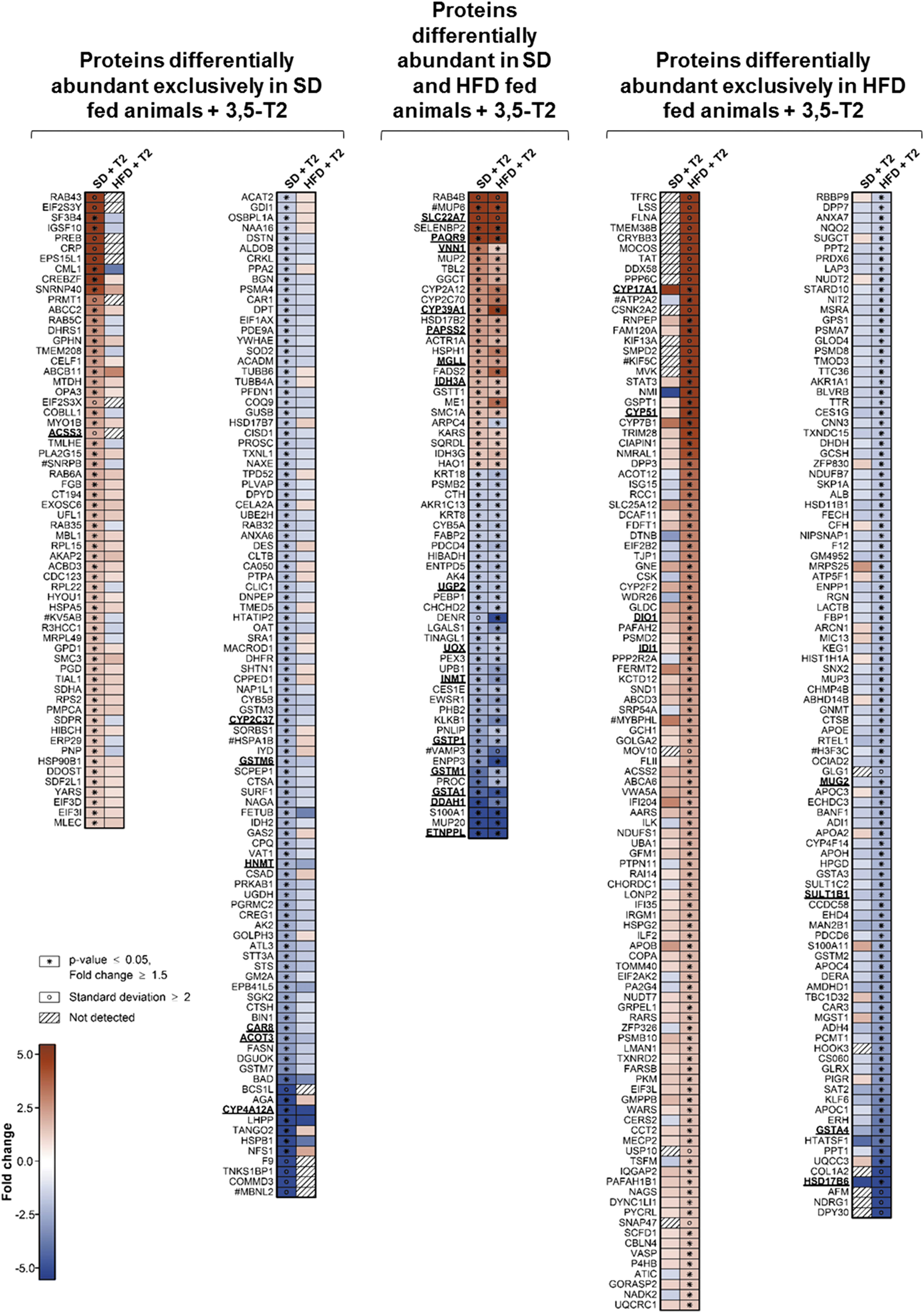
Heat maps summarizing the differential abundance of hepatic murine proteins from animals fed SD or high-fat diet (HFD) under treatment with 3,5-T2 for 4 weeks as compared with corresponding control animals treated with saline. The maps include all proteins fulfilling the criteria for differential abundance applied in this study (p ≤ 0.05, fold change ≥|1.5|). The numbers of animals per group were as follows: SD-saline: 4; SD-3,5-T2: 3; HFD-saline: 4; HFD-3,5-T2: 4. Red and blue indicates increased and decreased protein abundance, respectively, as compared with the respective saline-treated control groups. Underlined protein names indicate that for the respective proteins also significantly altered transcript levels were detected in the corresponding microarray-based transcriptome analysis (p ≤ 0.05, fold change ≥|1.5|). Proteins that could not be identified to be clearly unique but belong to groups of closely related homologs are labeled by #. Diagonally striped boxes indicate proteins not detected at one of the two conditions compared. SD, standard diet; 3,5-T2, 3,5-diiodothyronine.
Comparative analysis of proteome and transcriptome data
The proteome results were compared with complementary microarray-based transcriptome data (15) generated from the same specimens as the protein samples. The transcriptome analysis detected 49 and 54 significantly increased and decreased gene-specific transcript levels under 3,5-T2 treatment compared with vehicle treatment in HFD mice (proteome: 135 and 137 proteins). For mice fed SD, treatment caused 118 and 103 significantly increased and decreased gene-specific transcript levels (proteome: 89 and 134 proteins). Thus, the corresponding protein/transcript ratio was higher in HFD mice (2.6 vs. 1.0) (Supplementary Fig. S2).
Of all genes differentially expressed in HFD and SD mice on at least one level (360 and 423), 24.4% and 47.3% of the alterations were exclusively observed in the transcript level and 71.4% and 47.8% in the protein level, while only 4.2% and 4.9% of the genes showed consistent differential expression in both levels, indicating a rather low degree of concordance (Supplementary Fig. 2). Transcripts and proteins with significant differential abundance under 3,5-T2 treatment in HFD and SD mice are listed in Supplementary Table S1.
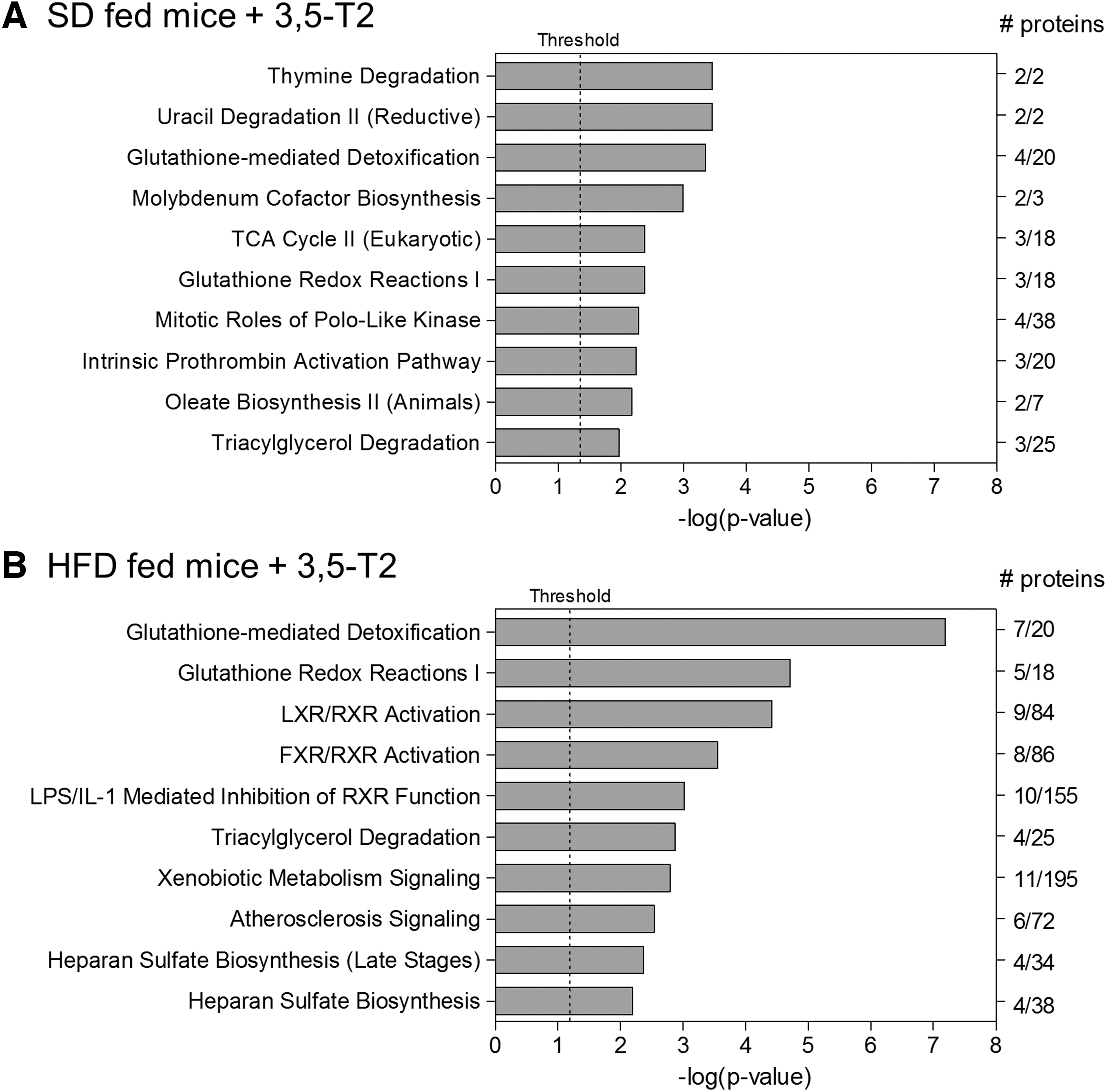
Results of in silico pathway enrichment analyses using the Ingenuity Pathway Analysis software and an input data set encompassing all hepatic proteins exhibiting significant differential abundance (p ≤ 0.05, fold change ≥|1.5|) in mice fed SD (
In case of the 88 and 200 genes exhibiting exclusively transcript level changes under 3,5-T2 treatment in HFD and SD animals, the missing detection of differentially abundant proteins could be attributed to the fact that they were not covered by the proteome analysis because of low abundance or technical reasons. Indeed, 75 (85.2%) and 170 (85.0%) of these proteins were not detected, suggesting that this discrepancy is explained by the lack of coverage in the proteome analysis. In contrast, for most differentially abundant proteins their coding DNA sequences were covered by the microarray analysis, and thus the failure to detect corresponding mRNA level changes could either be due to pronounced technical variation in the transcriptome analysis or specific post-transcriptional regulatory events.
To exclude technical reasons, we determined the coefficient of variance for the transcriptome and proteome measurements of all experimental groups. The demonstrated considerably higher intersample variance in the proteome data (Supplementary Fig. S3), indicating greater interindividual biological and technical measurement differences. The same result was obtained when the analysis was confined to genes exhibiting differential expression under 3,5-T2 treatment exclusively on the protein level, indicating that the relatively large number of such genes was not due to inhomogeneous interindividual mRNA levels in the different experimental groups.
Pathway enrichment analyses of 3,5-T2-mediated alterations of liver protein levels
Pathway enrichment analyses were performed using the Ingenuity Pathway Analysis software and an input data set encompassing all hepatic proteins exhibiting significantly differential abundance in mice fed SD or HFD treated with 3,5-T2 or saline for 4 weeks. Of 40 and 49 significantly enriched canonical IPA pathways detected for SD- and HFD-fed animals, we focused on the 10 respective top pathways (Fig. 2). As the proteins assigned to these pathways partially overlapped, we grouped them to the larger physiologically defined entities Proteins involved in glutathione- and xenobiotic-related metabolism and Proteins involved in lipid metabolism. The pathways Thymine Degradation/and Uracil Degradation II (reductive), Molybdenum Cofactor Biosynthesis, Mitotic Roles of Polo-Like Kinase, and Intrinsic Prothrombin Activation Pathway are discussed in the Supplemental Data.
3,5-T2 strongly increases the levels of cytochrome P450 17A1, a sex steroid biosynthesis key enzyme
Our proteome analysis detected a 20.9-fold increase in the hepatic abundance of cytochrome P450 17A1 (CYP17A1) in HFD-fed animals under 3,5-T2 treatment (Fig. 3, Supplementary Fig. S4). The same trend was observed for SD animals (Fig. 3). Increased CYP17A1 mRNA levels were observed in both diet groups, indicating transcriptional induction. CYP17A1 converts pregnenolone to dehydroepiandrosterone (DHEA) and pregnenolone-derived progesterone to 17-hydroxyprogesterone (16). Because pregnenolone is not hepatically synthesized, pregnenolone sulfate delivered from steroidogenic tissues through the bloodstream represents its most likely source in the liver (17 –19). To catalyze DHEA synthesis, CYP17A1 requires interaction with microsomal cytochrome b5 (CYB5A) (16,20,21). As our proteome data demonstrated a slightly reduced abundance of CYB5A under 3,5-T2 treatment in SD- and HFD-fed mice, it was not clear how this might counteract the predicted induced DHEA synthesis (Fig. 3, Supplementary Fig. S4). DHEA is further processed to testosterone through androstenediol or androstenedione.
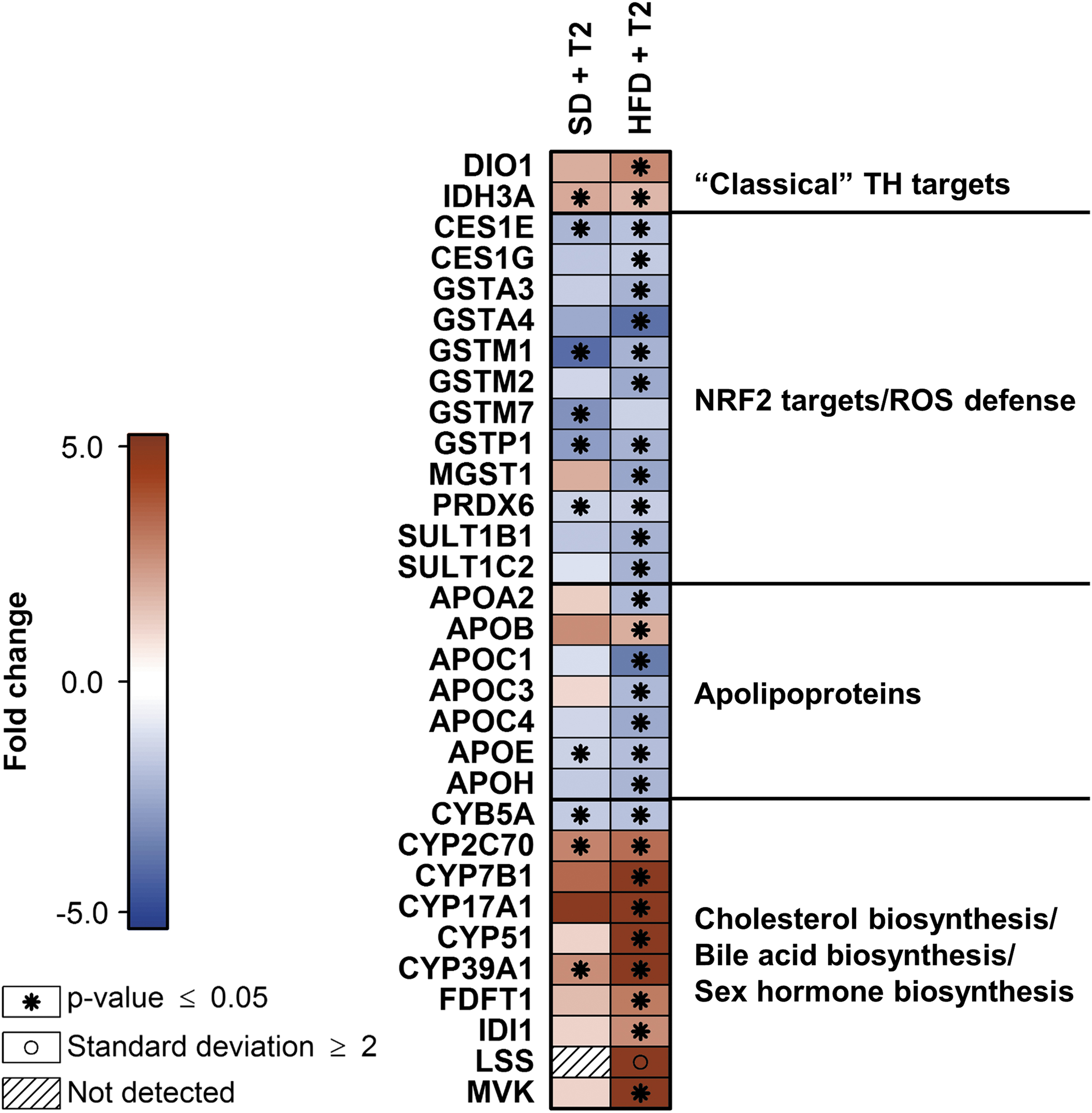
Heat map illustrating abundance alterations of specific hepatic murine proteins assigned to different functional categories under treatment with 3,5-T2 for 4 weeks. Included are proteins that exhibited either significant abundance alterations under treatment (p ≤ 0.05, fold change ≥|1.5|) in at least one diet group (SD or HFD) as compared with the respective control group treated with saline (labeled with *) or, in the single case of LSS, a mean derived protein level of the experimental group classified as “not detected” (here: HFD animals treated with saline) greater or smaller than the twofold standard deviation of that group categorized as “detected” (here: HFD animals treated with 3,5-T2) (labeled with ○). Red and blue indicates increased and decreased protein abundance, respectively, as compared with the respective saline-treated control groups. Diagonally striped boxes indicate proteins not detected under the compared conditions (here: LSS in SD fed animals in both treatment groups). LSS, lanosterol synthase.
Many testosterone effects are finally exerted by its metabolite DHT (22). Physiological considerations based on the estimated hepatic levels of the enzymes involved in sex steroid biosynthesis suggested increased liver concentrations of androstenedione and progesterone as a consequence of the induced CYP17A1 synthesis (see Supplemental Data and Supplementary Fig. S4 for details). Accordingly, both steroids should accumulate in the liver in excess and, therefore, subsequently be released to the circulation. As this implied enhanced systemic male sex steroid effects under 3,5-T2 treatment, we determined sex steroid profiles using gas chromatography-tandem mass spectrometry to validate such far-reaching metabolic consequences directly at the metabolite level.
3,5-T2 increases hepatic but not systemic male sex hormone concentrations in HFD mice
The concentrations of various sex steroids such as progesterone, androstenedione, testosterone, and DHT were determined in liver, serum, inguinal WAT, and gonadal WAT of HFD and SD mice under 3,5-T2 treatment. As expected, we observed increased hepatic concentrations of progesterone and androstenedione and similar trends for testosterone and DHT (Fig. 4). In the other tissues, no significant alterations could be measured, with the exception of increased progesterone in inguinal WAT, while no significant effects were observed for SD mice (data not shown).
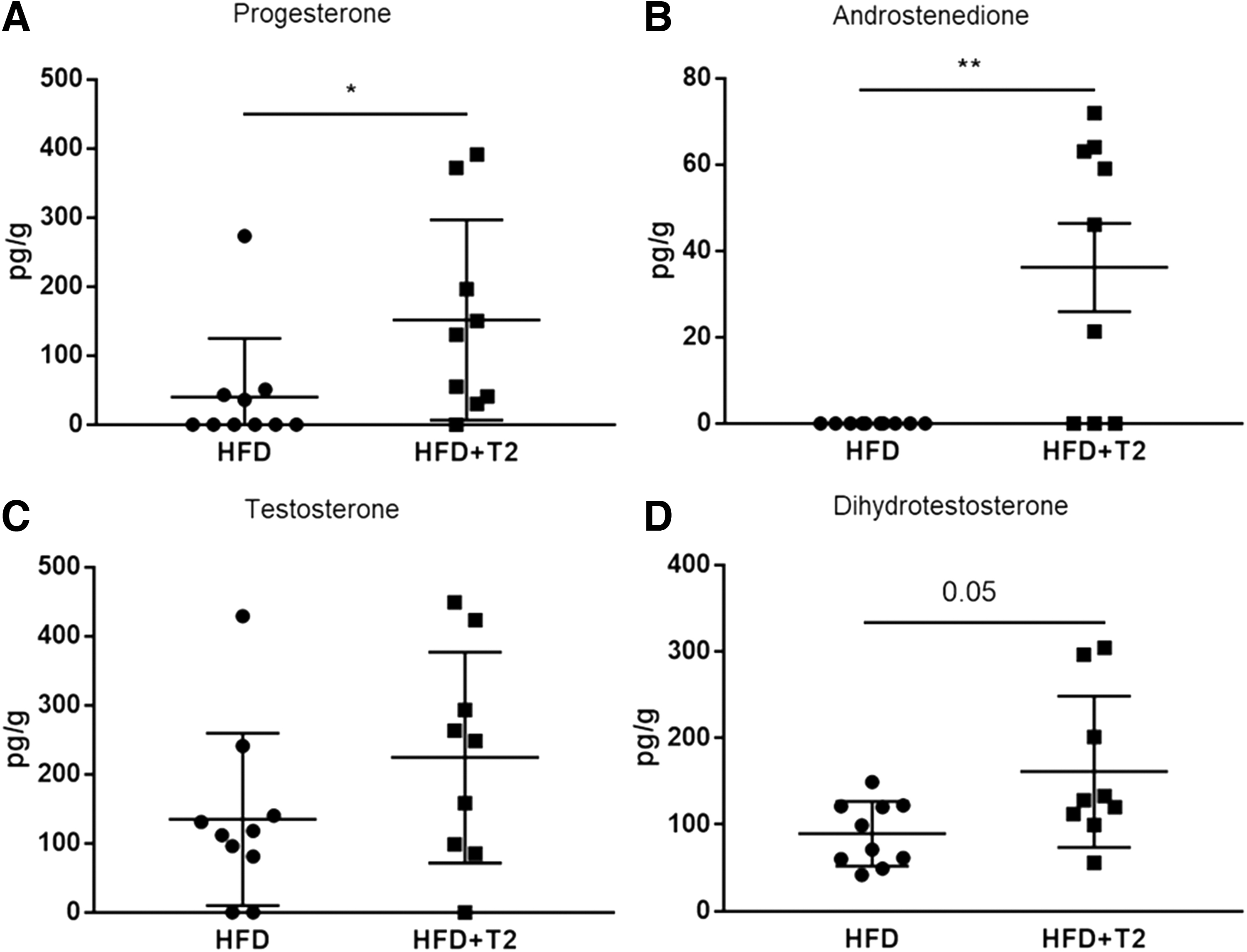
Hepatic concentrations of progesterone (
Discussion
The mechanisms underlying the large number of genes differentially expressed under 3,5-T2 treatment exclusively at the protein level remain hypothetical. As treatment was performed for a comparably long period (4 weeks), early induced transcriptional responses might already be reset to baseline after that time, while resulting proteome alterations are fixed because of resulting changes in hepatocellular proteostasis. Transcriptional desensitization of hepatic murine genes acutely affected by triiodothyronine (T3) treatment has been reported, while other genes were exclusively regulated after treatment for 10 days, and few genes at both times (23). The exclusive proteome alterations could also be based on post-transcriptional regulatory mechanisms modulated by noncanonical signaling of 3,5-T2 through cytoplasmic TH receptors (THRs) or independent of those, according to type 3 and 4 TH signaling as defined by Flamant et al. (24). Regulation by 3,5-T2 involving noncanonical TH signaling has been reported (4,25 –27).
The higher number of proteins exhibiting differential levels under 3,5-T2 treatment in HFD-fed compared with SD-fed animals suggests modulation of the response reflected by the proteome by the hepatic lipid load. Activity modulation of nuclear receptors by the lipid content was described for members of the peroxisome proliferator-activated receptor (PPAR) family (α, β/δ, γ) regulated by fatty acid ligands (28,29). There is considerable hepatic nuclear receptor crosstalk between THRs, farnesoid X receptor (FXR), liver X receptor (LXR), and the PPARs, all sharing retinoid X receptor (RXR) as dimerization partner for DNA binding, and may partially compete for it (30,31). Possibly, the hepatocellular environment in HFD animals with high lipid content provokes a different combination of active receptors compared with SD animals, thus enhancing 3,5-T2 effects as reflected in the higher number of differentially abundant proteins.
We recently reported citrate synthase activity induction by 3,5-T2 comparable with T3 in murine HFD liver (9). Consistently, the isocitrate dehydrogenase 3 subunits IDH3A and IDH3G exhibited increased levels in both diet groups under treatment. Succinate dehydrogenase complex flavoprotein subunit A showed higher expression at the protein and transcript level in SD-fed animals and increased mRNA in HFD animals treated for 2 weeks (15). Transcriptional Idh3a activation by classical canonical genomic signaling [type 1 according to Flamant et al. (24)] was demonstrated for different rodents (23,32,33). Our data suggest the same mechanism for 3,5-T2-induced tricarboxylic acid cycle activation in mice.
Established THRβ target genes exhibited similar hepatic expression patterns under 3,5-T2 treatment as known for T3, Dio1 encoding iodothyronine deiodinase I, was significantly higher expressed at both levels in HFD-fed animals; in SD-fed mice, transcript levels were significantly higher, whereas Serpina7 encoding the secreted thyroxine-binding globulin was not detectable in the proteome analysis showed decreased transcript levels in both diet groups (Fig. 3, Supplementary Table S1).
Proteins involved in glutathione- and xenobiotic-related metabolism: after considering overlaps between the enriched IPA-pathways Glutathione-mediated Detoxification, Glutathione Redox Reactions I, Xenobiotic Metabolism Signaling, LPS/IL-1 Mediated Inhibition of RXR Function, Heparan Sulfate Biosynthesis, Heparan Sulfate Biosynthesis (Late Stages), and Triacylglycerol Degradation, 14 of the remaining 19 proteins represented phase I and II detoxification enzymes or with related function, namely glutathione S-transferases, carboxylesterases, sulfotransferases, and peroxiredoxins (34 –40). Of these, 4 and 11 exhibited decreased hepatic amounts under 3,5-T2 treatment in SD and HFD mice, respectively, with transcript level alterations for 4 and 3 proteins.
A number of these proteins are encoded by genes of the nuclear factor erythroid 2-related factor 2 (NRF2) regulon (Fig. 3) (41 –47). NRF2 represents the transcriptional master regulator of the major eukaryotic defense and detoxification system against oxidative cytotoxic stress (48). Increased reactive oxygen species (ROS) levels cause NRF2 activation and upregulation of its target genes encoding proteins involved in glutathione synthesis and detoxification of electrophilic compounds (49). As expression of these genes reflects correspondingly altered ROS levels (50), their 3,5-T2-induced hepatic downregulation might thus be interpreted as the consequence of ROS reduction, based on the well-known association between liver fat and hepatic ROS levels: HFD caused significant weight gain compared with SD (15), and a proportional increase in hepatic fat.
Initially, the liver adapts to such conditions by higher mitochondrial respiratory rates, causing enhanced ROS production that is coped by cellular defense systems, namely the NRF2 regulon. However, if the lipid load further increases and the pathological lipotoxic state becomes chronic, respiration continuously declines, along with pronounced mitochondrial uncoupling and leaking as well as further increased ROS levels finally exceeding the capacities of the protection systems, resulting in cytotoxic oxidative stress (51). In contrast, the antisteatotic 3,5-T2 effects through stimulation of hepatic fatty acid oxidation are well established (52), and treatment of HFD mice as described in our study caused a significant hepatic triglyceride decrease (9). Thus, lower NRF2 regulon protein levels under 3,5-T2 might reflect reduced liver fat and associated ROS levels, and, respectively, lower oxidative stress. As expected, given the clearly higher lipid load and corresponding ROS levels in HFD mice, this signature was more pronounced in this group (Fig. 3).
Proteins involved in lipid metabolism: after considering overlaps between the enriched IPA pathways - LXR/RXR Activation, FXR/RXR Activation, LPS/IL-1 Mediated Inhibition of RXR Function, Atherosclerosis Signaling, Triacylglycerol Degradation, and Oleate Biosynthesis II (Animals), 25 proteins remained, of which 10 are discussed earlier. Of the remaining, 5 apolipoproteins exhibited predominantly lower abundance exclusively in HFD animals under 3,5-T2 without corresponding mRNA level alterations, and two more detected by intensified data inspection shared this profile (Fig. 3). For APOE, this confirmed a reported hepatic abundance decrease in HFD-fed low density lipoprotein receptor (LDLR) knockout mice treated with 3,5-T2 (13).
Strikingly, those apolipoproteins mostly represented components of very LDL (VLDL) particles involved in the fuel transport pathway ensuring triglyceride delivery from the liver to peripheral tissues (53). Most convincingly, this is explained by the 3,5-T2-mediated reduced hepatic triglyceride load: while under HFD hepatic VLDL particle production is increased (54), this should be partially reversed under treatment. Increased hepatic amounts under treatment were only measured for apolipoprotein B (APOB) in HFD mice (Fig. 3). As during LDL internalization hepatic APOB reuptake occurs through LDLR (53) and one of the well-known T3 effects is transcriptional upregulation of Ldlr (55), the elevated APOB levels might be explained by similar 3,5-T2-mediated regulation.
Extended proteome data analysis based on the IPA results revealed five cholesterol biosynthesis key enzymes [mevalonate kinase (MVK), isopentenyl-diphosphate delta-isomerase 1 (IDI1), farnesyl-diphosphate farnesyl transferase 1 (FDFT1), lanosterol synthase (LSS), and lanosterol 14α-demethylase (CYP51) (56)] exhibiting significantly higher levels exclusively or predominantly in 3,5-T2-treated HFD mice, where IDI1 and CYP51 also demonstrated increased transcript levels (Figs. 3 and 5). Of the remaining enzymes of this pathway, 13 were detected in at least three animals in at least one experimental group without differential abundances under treatment (Fig. 5). However, for sterol-4α-carboxylate 3-dehydrogenase (NSDHL), 3-ketosteroid reductase (HSD17B7), and methylsterol monooxygenase 1 (MSMO1) the corresponding transcript levels were significantly increased in HFD-fed 3,5-T2-treated animals. Six enzymes were not detected in all samples, among them 3-hydroxy-3-methylglutaryl-CoA reductase (HMGCR), whose THRβ-mediated transcriptional induction by T3 stimulates cholesterol biosynthesis (30,31) (Fig. 5). In our study, no HMGCR mRNA induction was observed. The squalene monooxygenase (SQLE) transcript levels were significantly increased under treatment in HFD-fed animals, indicating that this could also be the case for the nondetectable enzyme. Exclusively in SD animals, cytosolic acetyl-CoA acetyltransferase (ACAT2) and HSD17B7 exhibited decreased levels, without transcript changes. In summary, the overall protein and mRNA patterns suggested hepatic cholesterol biosynthesis upregulation in HFD mice under 3,5-T2 treatment.
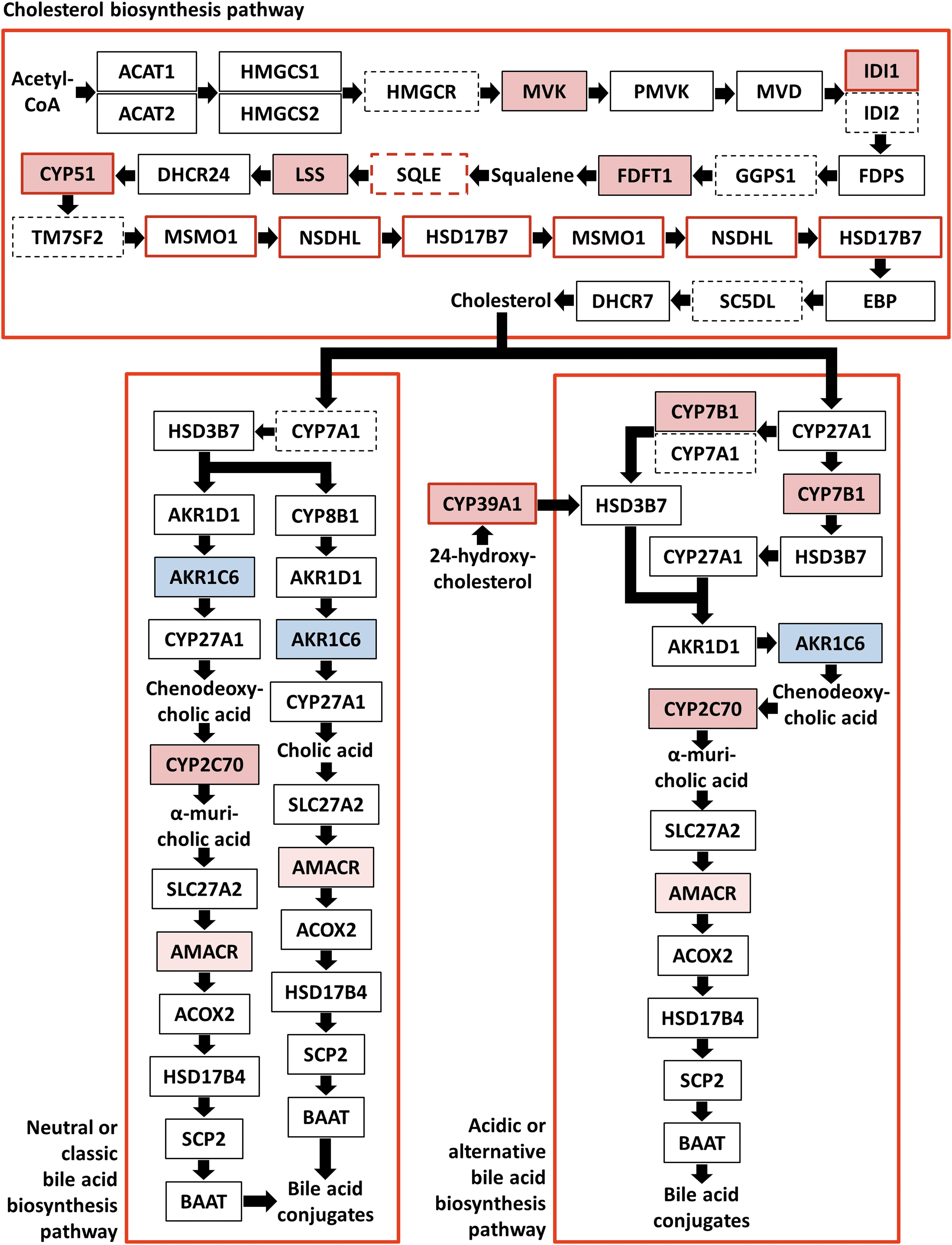
Presentation of the differential abundance of hepatic murine proteins and respective transcripts involved in cholesterol and bile acid biosynthesis in animals fed HFD under treatment with 3,5-T2 for 4 weeks as compared with corresponding control animals treated with saline. Squares represent enzymes where continuous and dashed frame lines indicate detected and not detectable proteins in the present proteome analysis, respectively. Arrows visualize the directions of the metabolite flux and red and blue square areas indicate increased and decreased protein levels, respectively, under treatment. Red frame lines mark increased levels of the respective transcripts. Criteria for differential abundance: p ≤ 0.05, fold change ≥|1.5|, with the exceptions of AKR1C6 and AMACR, where the protein amount fold change amounted only to 1.4 and 1.2, respectively. The following enzymes are depicted: ACAT1 (mitochondrial and peroxisomal acetyl-CoA acetyltransferase), ACAT2 (cytosolic acetyl-CoA acetyltransferase), HMGCS1 (cytosolic hydroxymethylglutaryl-CoA synthase), HMGCS2 (mitochondrial/peroxisomal hydroxymethylglutaryl-CoA synthase), HMGCR (3-hydroxy-3-methylglutaryl-CoA reductase), MVK (mevalonate kinase), PMVK (phosphomevalonate kinase), MVD (mevalonate diphosphate decarboxylase), IDI1 (isopentenyl-diphosphate delta-isomerase 1), IDI2 (isopentenyl-diphosphate delta-isomerase 2), FDPS (farnesyl diphosphate synthase), GGPS1 (geranylgeranyl pyrophosphatase synthase), FDFT1 (farnesyl-diphosphate farnesyl transferase 1), SQLE (squalene monooxygenase), LSS, DHCR24 (Δ24-sterol reductase), CYP51 (lanosterol 14α-demethylase), TM7SF2 (Δ14-sterol reductase; transmembrane 7 superfamily member 2), MSMO1 (methylsterol monooxygenase 1), NSDHL (sterol-4α-carboxylate 3-dehydrogenase; NAD(P) dependent steroid dehydrogenase-like), HSD17B7 (3-ketosteroid reductase), EBP (3β-hydroxysteroid-Δ8, Δ7-isomerase; emopamil-binding protein), SC5DL (lathosterol oxidase; sterol-C5-deasturase), DHCR7 (7-dehydrocholesterol reductase), CYP7A1 (cholesterol 7α-hydroxylase), HSD3B7 (3β-hydroxy-Δ5-C27 steroid oxidoreductase), CYP8B1 (sterol 12α-hydroxylase), AKR1D1 (Δ4-3-oxosteroid 5β-reductase), AKR1C4 (3α-hydroxysteroid dehydrogenase), CYP27A1 (sterol 27-hydroxylase), CYP2C70 (chenodeoxycholic acid 6-hydroxylase), SLC27A2 (bile acid CoA ligase/very long-chain acyl-CoA synthetase), AKR1C6 (estradiol 17 beta-dehydrogenase 5 [aldo-keto reductase family 1, member C6]), AMACR (α-methylacyl-CoA racemase), ACOX2 (peroxisomal branched-chain acyl-coenzyme A oxidase 2), HSD17B4 (peroxisomal multifunctional enzyme type 2), SCP2 (nonspecific lipid-transfer protein), BAAT (bile acid-CoA:amino acid N-acyltransferase), CYP7B1 (oxysterol 7α-hydroxylase), and CYP39A1 (24-hydroxycholesterol 7α-hydroxylase). In addition, the positions of the key metabolites acetyl-CoA, squalene, cholesterol, chenodeoxycholic acid, cholic acid, and 24-hydroxycholesterol within the pathways are shown.
The enzymes oxysterol 7α-hydroxylase (CYP7B1), 24-hydroxycholesterol 7α-hydroxylase (CYP39A1), and chenodeoxycholic acid 6-hydroxylase (CYP2C70), involved in the cholesterol downstream metabolism toward bile acid (BA) synthesis, also exhibited increased abundances under 3,5-T2 in HFD mice. For CYP39A1, transcript levels were also higher. In SD animals, these effects were clearly blunted (Fig. 3). CYP7B1 catalyzes biosynthesis of two chenodeoxycholic acid (CA) precursors in the alternative or acidic pathway of BA synthesis, which contributes 25–30% of BA in mice, while CYP39A1 mediates 7α-hydroxylation of its substrate 24-hydroxycholesterol (24-HC) in the 24-HC-pathway, with further processing toward CA (57 –59). CYP2C70 catalyzes the mouse-specific CA hydroxylation at position 6 to form α-muricholic acid (60).
Of the other enzymes involved in the neutral and alternative or acidic BA biosynthesis pathways, as well as of those catalyzing the subsequent steps toward BA conjugation, 12 were detected in at least 3 animals per experimental group, without significantly differential abundances under treatment. For α-methylacyl-CoA racemase (AMACR) and estradiol 17 beta-dehydrogenase 5 (aldo-keto reductase family 1, member C6, weak but significant increased and decreased abundances were found exclusively in HFD animals under 3,5-T2 treatment. CYP7A1 was not detected in all samples. No additional transcript level alterations were measured (Figs. 3 and 5).
To the best of our knowledge, the concerted distinct increase of the hepatic CYP7B1, CYP39A1, and CYP2C70 abundance in HFD animals under 3,5-T2, strongly suggesting upregulation of BA synthesis, has not been described before. For T3, stimulation of cholesterol conversion to BAs is known (30,31), and transcriptional induction of the gene encoding cholesterol 7α-hydroxylase CYP7A1, the rate-limiting enzyme of this pathway, represents the central regulatory step (30,31). While CYP7A1 was not detectable in our proteome analysis, no transcript level alterations were observed for it. However, of several studies of TH treatment effects on the hepatic transcriptome in rodents, only one in rats showed increased CYP7A1 transcript levels after 3 hours of T3 treatment (32) while in mice and in other rat analyses, comparable and longer TH treatment periods failed to demonstrate corresponding mRNA alterations (23,32,33,61 –65).
In the end, while CYP7A1 catalyzes the initial cholesterol hydroxylation in the neutral or classic pathway of BA biosynthesis, the detected higher amounts of namely CYP7B1 and CYP2C70 should also provoke increased BA synthesis through accelerated metabolite flux especially through the acidic or alternative BA biosynthesis pathway (Fig. 5) (57,58).
The increased levels of several key enzymes of cholesterol and BA synthesis in HFD mice under 3,5-T2 treatment might be explained most convincingly by the assumption that one of the mechanisms of 3,5-T2 mediating the well-known reduction of hepatic fat is, besides induction of fatty acid oxidation, stimulation of cholesterol- and BA synthesis, with subsequent excretion of the latter through bile. Accordingly, acetyl-CoA generated by hepatic fatty acid breakdown serves as initial building block for these energy consuming synthesis pathways. The underlying regulation is not clear, but as of nine enzymes participating in cholesterol and BA synthesis present in increased amounts in HFD-fed animals under 3,5-T2 treatment only three also demonstrated increased mRNA levels, suggesting the involvement of post-translational regulatory mechanisms. In this context, the recent observation that noncanonical T3-mediated TRβ signaling is responsible for maintenance of normal serum and liver triglyceride concentrations and that inactivation of this noncanonical signaling causes hepatic and serum hypertriglyceridemia is noteworthy (66).
Remarkably, cholesterol and BA synthesis coupling is principally known for rodents: the LXR LXRα is activated by oxysterols whose levels are increased under conditions of cholesterol accumulation. Activated LXRα, in conjunction with liver receptor homolog-1 (LRH-1), then stimulates transcription of the gene encoding CYP7A1 (57 –59). However, CYP7A1 mRNA was not increased under 3,5-T2 treatment in our analyses, and the higher abundances of CYP7B1, CYP39A1, and CYP2C70 indicate different regulatory mechanisms.
Hepatic abundance of CYP17A1, a sex steroid biosynthesis key enzyme, was preferentially increased in 3,5-T2-treated animals fed HFD. Supporting sex steroid profile measurements (Fig. 4) demonstrated no systemic but liver-specific intensified sex steroid exposition, predicted to cause pronounced hepatic “masculinization.” The resulting physiological effects are not yet clear but might be of detrimental nature, thus this finding emphasizes the need of additional analyses to gain a deeper understanding of the involved mechanism(s) and health-relevant consequences. Therefore, while the potentially limited transferability of data obtained using a murine model to humans has to be kept in mind, these results once more demonstrate that besides the well-established health-promoting effect of 3,5-T2 in decreasing hepatic fat also putatively harmful effects such as the observed liver “masculinization” must be taken into account.
Footnotes
Acknowledgments
We thank Elke Hammer for expert help in the proteome analyses, Ulrike Mäder for valuable input on the article, Anja Fischbach for excellent technical assistance, and Stephan Michalik for essential support in data analysis.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was generously supported by German Research Foundation (DFG) grant KFO 218/2 TP3 to J.K. and, as part of the DFG priority program “SPP 1629-ThyroidTransAct”, by the grants KO 922/16-1/2 to J.K. and HO 2140/6-2 to G.H.
Supplementary Material
Supplementary Data
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Table S1