
Abstract
Background:
Even though the clinical features of Graves' orbitopathy (GO) are well known, its exact pathogenesis remains controversial. The imbalance of redox homeostasis in the connective tissue could play a crucial role leading to an inflammatory state and edema of soft orbital tissues, thus contributing to orbital hypoxia and increase in hypoxia-inducible factor (HIF)-1α. This oxidative stress appears to target the orbital cells such as fibroblasts and also adipocytes. This study aims to explore which pathways can lead to the aforementioned oxidative stress in GO adipose cells and therefore offers new plausible therapeutic targets.
Methods:
Orbital fat samples were obtained from patients with GO (Western blot [WB]: n = 8, immunohistochemistry [IHC]: n = 8) and from control patients (WB: n = 5, IHC: n = 3–5). They were processed for WB analysis and IHC of the antioxidants (catalase, superoxide dismutase 1) and for HIF-1α. The expression of caveolin-1 (Cav-1) and deiodinase 3 (DIO3), known to be regulated by HIF-1α, was also analyzed by WB and IHC, as well as the targets of Cav-1: glucose transporter type 4 (Glut-4), NADPH oxidase (NOX)-2, and endothelial nitric oxide synthase (eNOS). Triiodothyronine (T3) expression was also analyzed by IHC.
Results:
In GO adipocytes, the expression of catalase was reduced, whereas that of HIF-1α was strongly increased. A decreased local T3 supply was associated with DIO3 upregulation. The low expression of Cav-1 in GO adipocytes was associated not only with low expression of Glut-4 but also with an increased expression of NOX-2 and active eNOS phosphorylated on serine 1177.
Conclusions:
Cav-1 and DIO3, both sensitive to hypoxia and to the increase of HIF-1α, play a pivotal role in the oxidative stress in GO adipocytes. DIO3 regulates the cellular supply of T3, which is essential for the cell homeostasis. Cav-1 determines the cellular glucose supply through Glut-4 and regulates the activity of NOX-2 generating superoxide anions and that of eNOS generating nitric oxide (NO).
Introduction
Graves' disease is an autoimmune disease leading to hyperthyroidism. It is sometimes associated with thyroid eye disease known as thyroid-associated orbitopathy, Graves' ophthalmopathy, or Graves' orbitopathy (GO). The clinical features observed in GO include exophthalmos, periorbital edema, eyelid retraction, and strabismus (1). The known immune reaction implicating autoantibodies that bind to the thyroid stimulating hormone receptor occurs not only in the thyroid but also in orbital tissues such as fat, extraocular muscles, and lacrimal glands (2). Orbital fibroblasts seem to be the main actors in this immune reaction leading to an infiltration of the orbit by T lymphocytes, which are a mixed type 1 helper (Th1)/type 2 helper (Th2) cells. Th1 lymphocytes that produce IL-2, IFN-γ, and TNF prevail at the onset of the disease, whereas Th2 cells predominate later and produce cytokines such as IL-4, IL-5, and IL-10 (3,4). This inflammation combined with the hyperthyroid state generates an overproduction of reactive oxygen species (ROS), mainly superoxide anion (O2 •−), and hydrogen peroxide (H2O2), therefore inducing oxidative stress in orbital fibroblasts. This mechanism is also amplified by the lowering of antioxidant defenses such as the catalase enzyme, known to detoxify H2O2. The combination of inflammatory cytokines, growth factors, and ROS induces an overproduction of glycosaminoglycans leading to edema and an increased proliferation of orbital fibroblasts (2) among which a subset expressing Thy-1 differentiates into adipocytes increasing the orbital fat volume (5). Later on, fibrosis settles in the orbit, characterizing the chronic stage of the disease.
The imbalance in the redox homeostasis observed in fibroblasts from GO patients has also been described in orbital adipocytes and extraocular muscle cells (6). The aim of this study was to determine the different pathways leading to the oxidative stress in Graves' orbital adipocytes.
Considerable in vivo and in vitro evidence supports a role for caveolae in adipocytes and skeletal muscular cells metabolism and homeostasis. Indeed, the absence of caveolae results in a variety of diseases, such as lipodystrophy, muscular dystrophy, cardiovascular diseases, and cancer. Caveolae are 50–100 nm plasma membrane invaginations that represent up to 50% of plasma membrane surface in adipocytes. They contain significant amounts of proteins such as cavins and three types of caveolin (Cav-1, Cav-2, and Cav-3), but Cav-1 is the principal structural component of caveolae in the plasma membrane (7).
In adipocytes and skeletal muscular cells, Cav-1 is known to regulate cellular trafficking and to be implicated in the insulin-dependent glucose uptake through the translocation of glucose transporter type 4 (Glut-4) vesicles into the plasma membrane (8). In skeletal muscle cells, the expression of Glut-4 is triiodothyronine (T3) dependent, and normal muscle responsiveness to insulin requires conversion of thyroxine (T4) to T3 by deiodinase 2 (DIO2) (9). T3 also plays an important role in adipocytes, its availability being determined by DIO2 and also by deiodinase 3 (DIO3), which transforms T4 into the inactive form reverse T3 (10,11). It is important to note that DIO3 is increased by hyperthyroidism and inflammation, which are hallmarks of Graves' disease (12).
Caveolae also host a great number of proteins involved in cellular redox signaling and are known in the cardiovascular system to accommodate two key cellular sources of ROS: NADPH oxidase type 2 (NOX-2) producing O2 •− and endothelial nitric oxide synthase (eNOS) producing nitric oxide (NO) (13). Moreover, Li et al. hypothesized that oxidative stress resulting from cardiac-specific Glut-4 deletion in mice could be related to an overexpression of NOX-2 (14). This would suggest that the oxidative stress due to the decrease in glucose supply could be induced by an overexpression of NOX-2.
This study was designed to analyze in vivo in orbital adipocytes from control and GO patients, the expression of Cav-1 and its target proteins: Glut-4, NOX-2, and eNOS and the expression of DIO2 and DIO3, which determine the cellular T3 content, T3 being a key factor in the metabolic homeostasis of adipocytes. Hypoxia-inducible factor (HIF)-1α expression was also analyzed since it is known to regulate Cav-1 and DIO3 expression.
Patients and Methods
Patients
In this study, we compared orbital fat from patients with GO and from control patients. We identified by immunohistochemistry (IHC) and immunofluorescence several proteins involved in cell homeostasis and we quantified the expression of these proteins by Western blots (WB).
In GO patients (WB: n = 8, IHC: n = 8), orbital fat samples were obtained from orbital fat decompression. In controls (WB: n = 5, IHC: n = 3–5), orbital fat samples were attained from lower lid transconjunctival blepharoplasties or from corrections of subconjunctival fat herniations.
The characteristics of GO patients included in this study are indicated in Table 1 for WB and in Table 2 for IHC. Our local ethics committee approved this study (2013/15AVR/171 and 2017/10OCT/473). The surgical specimens were obtained after patients gave written informed consent. Samples were obtained from the Biolibrary of Cliniques universitaires Saint-Luc, referenced as BB190044, member of the Biothèque Wallonie Bruxelles (BWB) and of biobanking and biomolecular resources research infrastructure in Belgium (BBMRI.be).
Characteristics of Patients with Graves' Orbitopathy for the Western Blot Analysis
GO, Graves' orbitopathy; TSAB, thyroid stimulating antibodies; TSH, thyrotropin.
Characteristics of Patients with Graves' Orbitopathy for the Immunostaining Analysis
Immunohistochemistry
Fat samples were fixed in 4% formaldehyde for 24 hours and embedded in paraffin. Paraffin sections were first dewaxed and rehydrated. Then, sections in citrate buffer (0.01 mol/L, pH 5.7) were heated in a microwave oven once for 3 minutes at 750 W, followed by four times for 3.5 minutes at 350 W. Endogenous peroxidase activity was inhibited in H2O2 solutions (10 μg/mL) for 20 minutes. Paraffin sections were then washed in phosphate-buffered saline with bovine serum albumin (PBS/BSA, 10 μg/mL) for 10 minutes and encountered with a solution of goat serum (20 μg/mL; Sigma, Saint Louis, MO) for 30 minutes at room temperature (RT). Sections were incubated with an appropriate dilution of primary antibody for an appropriate amount of time at RT. Negative controls were performed by omitting the primary antibody. Slides were then washed in PBS/BSA (10 μg/mL) for 10 minutes and incubated with a secondary antibody specific to the appropriate species (33.3 μg/mL) for 1 hour at RT (Pierce® goat anti-rabbit or anti-mouse; Thermo Scientific, Rockford, IL). After washing with PBS/BSA (10 μg/mL) for 10 minutes, the peroxidase activity was revealed with 3-3′ diaminobenzidine tetrahydrochloride (DAB; Sigma) and then washed after color development in PBS/BSA and distilled water. Sections were then counterstained with Mayer's hematoxylin, before being washed under tap water. The slides were mounted in Faramount Aqueous mounting medium (Dako, Glostrup, Denmark) and then observed under a light microscope. The characteristics of antibodies are summarized in Table 3.
Experimental Conditions for Immunohistochemistry and Western Blot
Cav-1, caveolin-1; DIO, deiodinase; eNOS, endothelial nitric oxide synthase; Glut-4, glucose transporter type 4; HIF, hypoxia-inducible factor; IHC, immunohistochemistry; NOX, NADPH oxidase; SOD, superoxide dismutase; T3, triiodothyronine; WB, Western blot.
Immunofluorescence
Paraffin sections were dewaxed, rehydrated, and pretreated with a solution of sodium tetrahydroborate (NaBH4) for three cycles of 10 minutes at 4°C, followed by three cycles of 5 minutes of PBS/Triton wash (0.5 μg/mL). Sections in citrate buffer (0.01 mol/L, pH 6) were then heated in a microwave oven once for 3 minutes at 750 W, followed by four times for 3.5 minutes at 350 W. Sections were washed with PBS/Triton (0.5 μg/mL) and then blocked with PBS/BSA (50 μg/mL). Sections were incubated overnight with a mixture of two primary antibodies from different species at a concentration of 3.3 μg/mL for the rabbit polyclonal Cav-1 antibody (BD610059; BD Transduction Laboratories), at a concentration of 20 μg/mL for the mouse monoclonal Glut-4 antibody (Sc53566; Santa Cruz Biotechnology, Dallas, TX), and at a dilution of 1/50 for the rabbit polyclonal pSerine1177eNOS antibody (ab75639; Abcam, Cambridge, United Kingdom) at RT. Negative controls were performed by omitting the primary antibodies. Slides were then washed with three cycles of 5 minutes of PBS/Triton (0.5 μg/mL). They were incubated in the dark for 1 hour at RT with a mixture of two fluorochrome-conjugated antibodies (3.3 μg/mL), each one being mouse or rabbit specific (Alexa Fluor 488 Goat Anti-Rabbit for Cav-1 and Alexa Fluor 568 Goat Anti-Mouse for Glut-4; Invitrogen, Carlsbad, CA). Sections were then washed with three cycles of 5 minutes of PBS/Triton (0.5 μg/mL) and counterstained with 4′,6-diamidino-2-phenylindole (DAPI, 0.1 μg/mL; Sigma) for 5 minutes. Slides were washed with three cycles of 5 minutes of PBS/Triton (0.5 μg/mL) and mounted in Fluorescence Mounting Medium (Dako).
WB analysis and antibodies
Orbital fat samples were suspended in RIPA buffer containing a protease inhibitor cocktail and Pho-stop. Protein concentration was determined using BCA protein assay kit (Pierce). Ten micrograms of lysate was loaded on 10% or 12% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (Bio-Rad) and transferred onto nitrocellulose membrane (Hybond ECL; Amersham Biosciences, Roosendaal, The Netherlands). Membranes were blocked for 1 hour at RT in a Tris-buffered saline (TBS, pH 7.4), 5% BSA, and 0.1% Tween-20. Membranes were then incubated with primary antibodies (Table 3) overnight at 4°C and peroxidase-conjugated secondary antibodies for 1 hour at RT. Proteins were visualized with enhanced chemiluminescence (ECL) on CL-XPosure Films (Pierce). Proteins bands were evaluated by densitometry. Protein levels were normalized to β-actin.
Statistics
Statistically significant differences (p < 0.05) were determined using unpaired Student's t-test (GraphPad InStat, San Diego, CA).
Results
The antioxidant status is modified and HIF-1α is overexpressed in GO orbital adipose tissue
The antioxidant enzymes, catalase and superoxide dismutase (SOD) 1, were investigated in control and GO orbital adipose tissues. Catalase protein expression was significantly decreased, while SOD1 protein was not different (Fig. 1A, B). The expression of HIF-1α, which is a redox-sensitive protein, was significantly higher in GO orbital adipocytes by WB compared with control orbital adipocytes where there was only weak expression of HIF-1α protein (Fig. 1C). Similar results were obtained for catalase, SOD1, and HIF-1α by IHC in GO adipocytes (Fig. 1D). These results indicate that the HIF-1α pathway is stimulated in GO adipose tissues, and this could potentially be due to the dysregulation of the redox homeostasis.
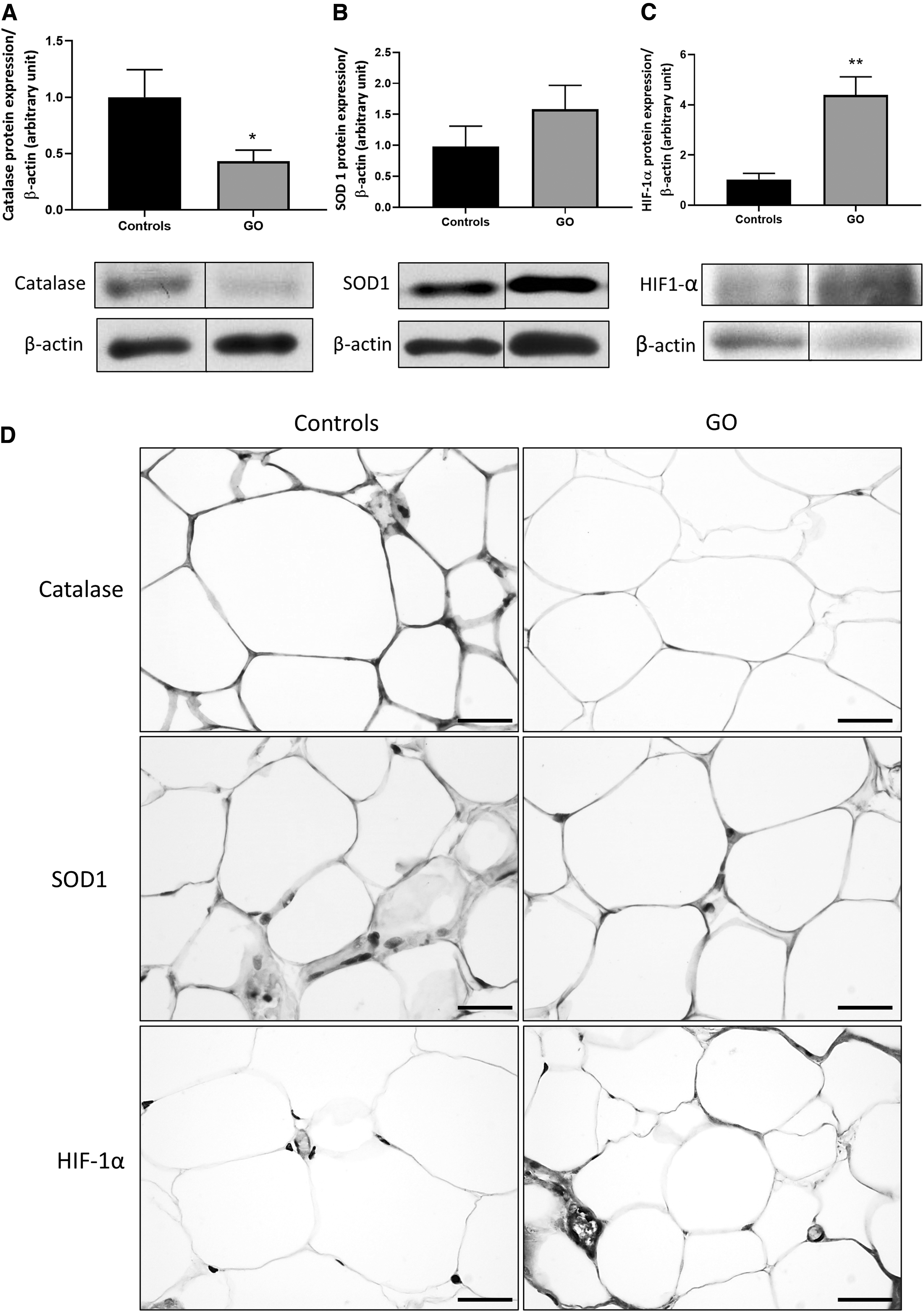
GO orbital adipose tissue antioxidants and HIF-1α subunit expression. (
DIO3 is upregulated in GO orbital adipose tissue
Since DIO3 is known to be a target of HIF-1α, which is overexpressed in GO orbital adipose tissue, we analyzed its expression. A significant upregulation of DIO3 was observed by WB and IHC (Fig. 2A, C). However, no difference in DIO2 expression was observed (Fig. 2B, C). In the cytoplasm of control orbital adipocytes, T3 was highly expressed, whereas no signal was detected in the cytoplasm of GO orbital adipocytes (Fig. 2C). T3 availability is thus reduced in GO orbital adipocytes, and this could therefore be implicated in the generation of oxidative stress.
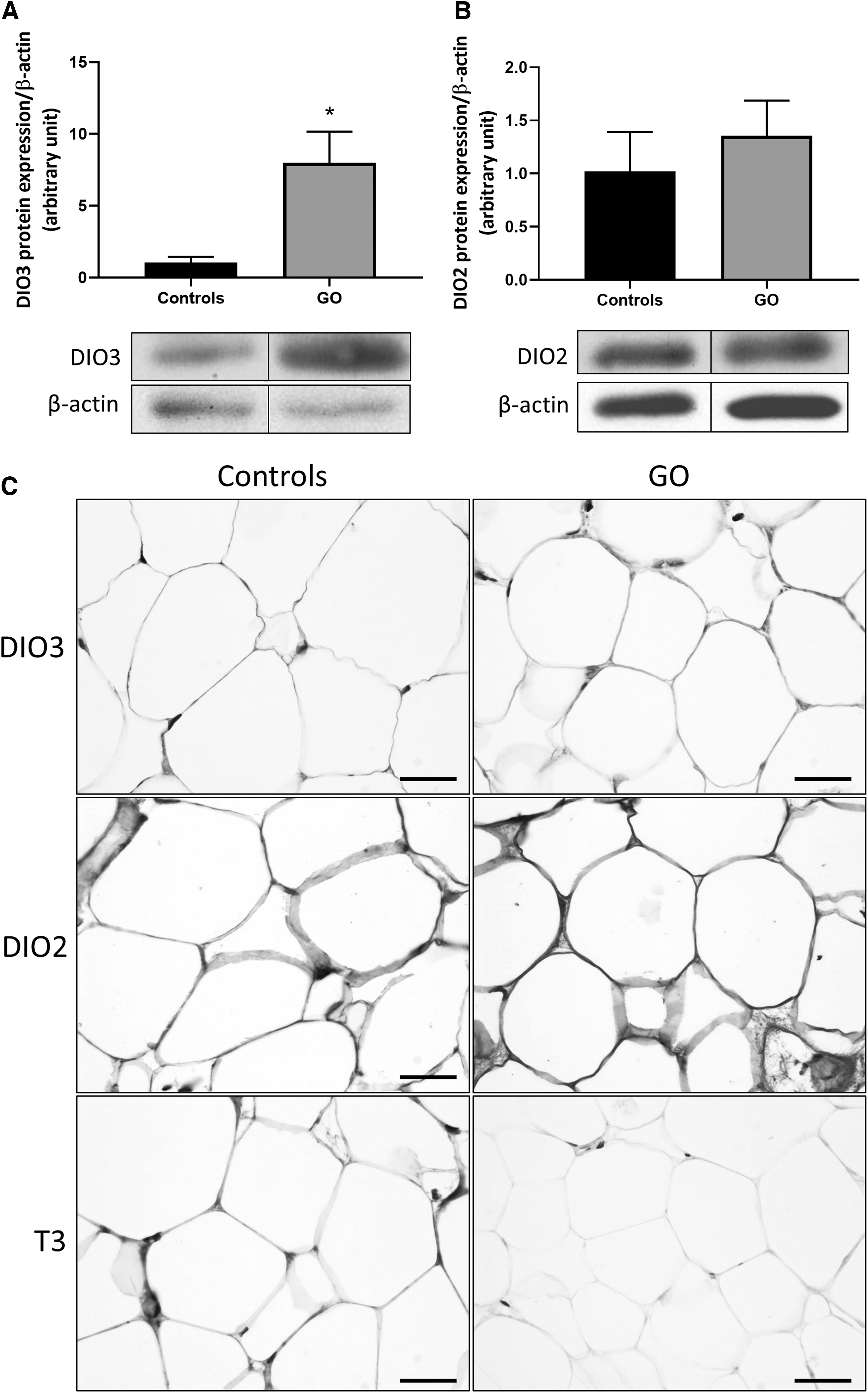
DIO3 and DIO2 expression in orbital fat biopsies from GO patients and T3 availability. (
Cav-1 and Glut-4 are colocalized in control orbital adipocytes
In control orbital adipocytes, Cav-1 (Fig. 3A) and Glut-4 (Fig. 3B) were detected at the plasma membrane. The orange fluorescence (Fig. 3C) resulting from the colocalization of Cav-1 and Glut-4 was detected at the plasma membrane of control orbital adipocytes.
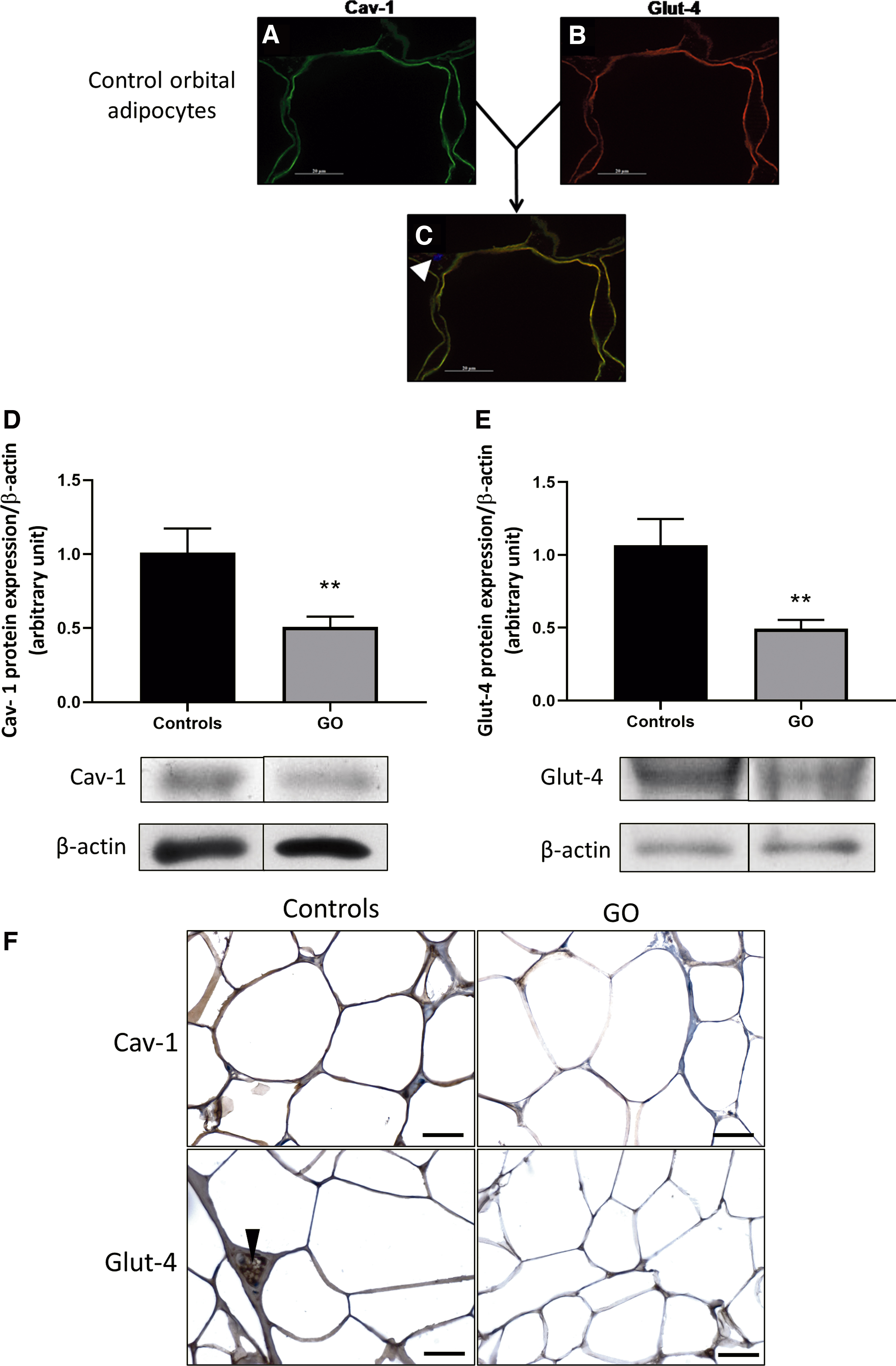
Cav-1 and Glut-4 expression in adipose tissue from GO patients. (
The low expression of Cav-1 in GO orbital adipose tissue is associated with a low expression of Glut-4
Cav-1 protein expression was significantly decreased in GO adipose tissue (Fig. 3D). Cav-1 immunostaining expression was high in the cytoplasm of control orbital adipocytes, whereas a weaker signal was found in GO orbital adipocytes (Fig. 3F).
As it has been demonstrated that Cav-1 depletion causes a decrease in Glut-4 in adipocytes (15), we investigated Glut-4 expression in GO orbital adipose tissue. Compared with control orbital adipocytes, GO orbital adipose tissue had lower Glut-4 protein expression by WB (Fig. 3E) as well as by IHC (Fig. 3F). These results suggest that Cav-1 downregulation in GO adipocytes could lead to a restriction of glucose supply.
NOX-2 and eNOS pS1177 are increased in GO orbital adipose tissue
GO orbital adipocytes had significant overexpression of NOX-2 protein compared with control adipocytes (Fig. 4A, B), therefore suggesting an increased production of O2 •− and H2O2 by NOX-2. In orbital adipose tissue from GO patients, a significant increase of phosphorylated eNOS on their activation site, the Serine 1177 (pS1177 eNOS), was observed (data not shown). Ratio between pS1177 eNOS and total form of eNOS confirmed eNOS stimulation in GO adipose tissue (Fig. 4C). Immunofluorescence for pS1177 eNOS showed upregulation in GO adipocytes (Fig. 4D).
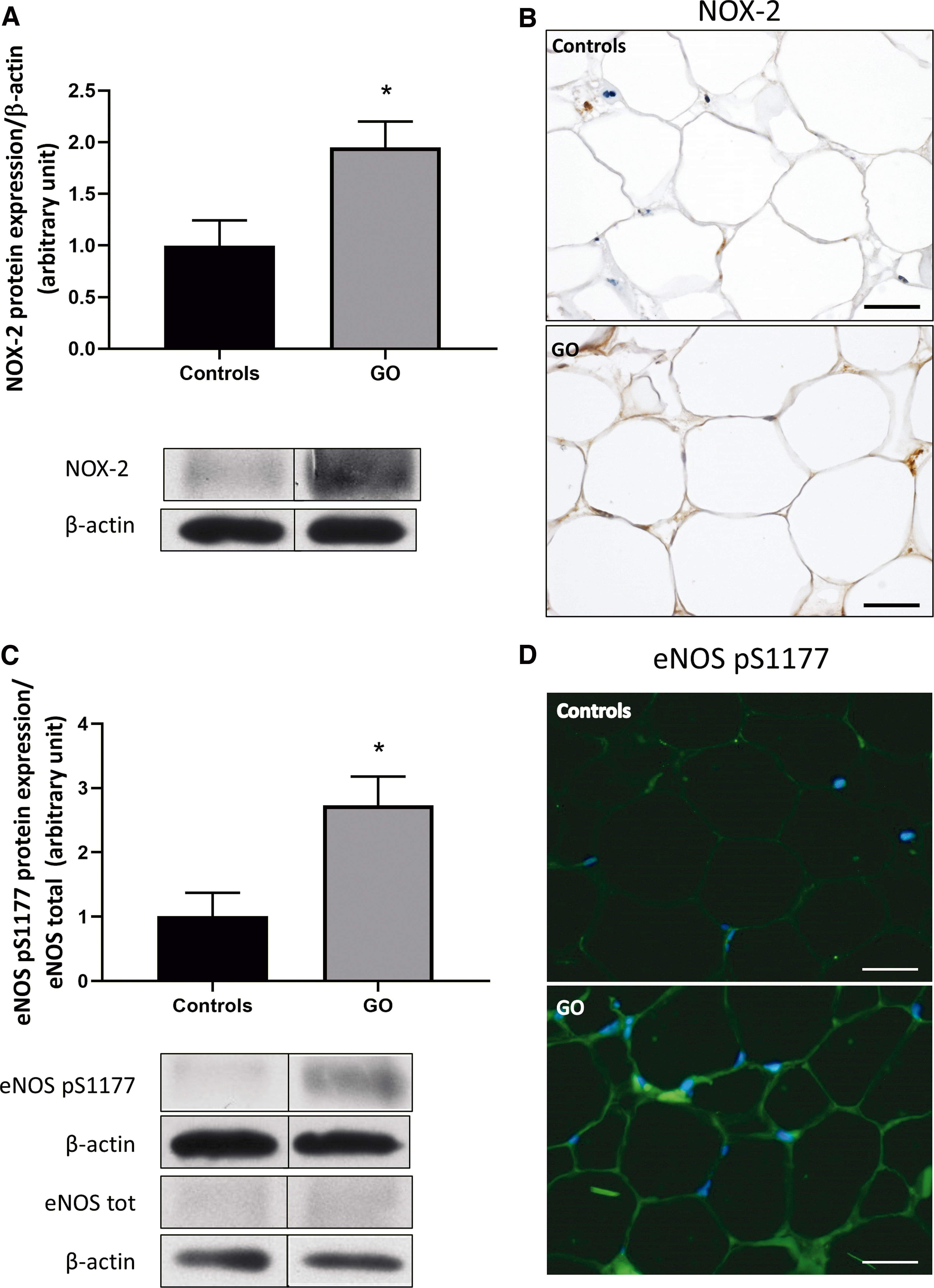
NOX-2 and eNOS pS1177 are overexpressed in GO adipose tissue. (
Discussion
As many inflammatory diseases, GO is associated with oxidative stress due to decreased antioxidant defenses unable to face the increased production of ROS. The oxidative stress has already been demonstrated in vivo in GO orbital adipocytes and extraocular muscle cells (6). Our results confirm a strong reduction of catalase in GO adipocytes, a potent antioxidant that detoxifies H2O2. They also show modifications in the expression patterns of Cav-1 and DIO3, two key proteins in adipocytes homeostasis.
Since the pioneering work of Semenza, it is well accepted that ROS generation is induced by chronic hypoxia and in turn activates HIF-1, which promotes persistent oxidative stress. This oxidative stress further amplifies HIF-1 activation and its consequent effects on gene expression and disease pathogenesis (16,17). The impact of HIF-1α, the O2-dependent subunit of HIF-1, in tissue remodeling in GO has been studied by Gortz et al. (18). They showed, in vitro, an increase of HIF-1α protein expression in response to hypoxia and cigarette smoke extract in orbital fibroblasts from GO patients compared with controls, and they observed, in vivo, an increase of fibroblast-like cells expressing HIF-1α in GO fat biopsies. In our study, we also observed an increase of HIF-1α protein expression in GO orbital adipose tissues.
Gortz et al. also observed that HIF-1-dependent VEGF and adiponectin secretion increases in GO fibroblasts showing that hypoxia stimulates HIF-1-dependent angiogenesis and adipogenesis (18). However, numerous other genes are also regulated by HIF-1 and, in the current study, we focused on DIO3 and Cav-1. In 2008, Simonides et al. demonstrated that hypoxia induces the expression of the DIO3 gene DIO3 by a HIF-1-dependent pathway and that HIF-1α interacts specifically with the DIO3 gene promoter (19). The DIO3 protein increase that we observed in GO adipocytes could thus be induced by HIF-1α and could be a consequence of ROS overproduction and hypoxia. In contrast with Planck et al. (20), we did not observe any difference in DIO2 protein expression. Nonetheless, the increased expression of DIO3 that we observed could be sufficient to reduce local T3 content and action, T3 being known as a critical determinant for adipocytes metabolism and homeostasis. The DIO3 upregulation reported here may be correlated with the low-T3 syndrome or nonthyroidal illness syndrome associated with inflammation and ROS overproduction (21). This could explain why GO features are frequently observed in euthyroid or hypothyroid patients.
Adipocytes homeostasis is also regulated by Cav-1, and it has been shown that hypoxia reduces caveolae-related proteins (7) and that Cav-1 transcription is directly regulated by HIF-1 (22). The reduction of Cav-1 protein expression that we observed in GO adipocytes is also probably due to ROS overproduction and hypoxia.
The downregulation of Cav-1 impacts the expression and activation of other proteins, and here, we showed an increased expression of NOX-2 and activation of eNOS in GO adipocytes. Chen et al. have already demonstrated in endothelium and adventitia of pulmonary arteries that Cav-1 acts as a negative regulator of NOX-2 activity, supporting the role of Cav-1 in the pathogenetic mechanism underlying cardiovascular diseases (23). Interestingly, Lei et al. have shown that Cav-1-knockout mice had elevated intraocular pressure and reduced outflow compared with control mice (24). This was associated with an increase of phosphorylated eNOS, suggesting that Cav-1 deficiency was responsible for increased eNOS activity, as also suggested by our results in GO. As Cav-1 proteins are downregulated in GO adipocytes, this could lead to caveolae disruption inducing eNOS uncoupling and NOX-2 (Nox/p22phox membrane complex) release in the cytoplasm, which then promotes its activation by cytoplasmic p47phox resulting in O2 •− production. Thus, the overexpression of NOX-2 and the activation of eNOS via downregulation of Cav-1 may induce the release of pro-oxidant molecules in GO adipocytes.
The oxidative stress in GO orbital adipocytes could result from several interconnected pathways, as summarized in Supplementary Figure S1. First, the downregulation of Cav-1 could lead to a reduced glucose supply to the cell, as suggested by our observations of the colocalization of Cav-1 and Glut-4 and the low expressions of both proteins. The oxidative stress due to the reduced glucose level could be due to superoxide anion production by NOX-2. This is supported by a study by Li et al., which suggests that cytosolic oxidative stress, probably originating from a source of ROS such as NOX-2, can be found in cardiac muscle cells from mice with a specific cardiac Glut-4 deletion (14). Our findings indicate that the expression of NOX-2 is also increased in the cytoplasm of GO orbital adipocytes. Superoxide anions produced by NOX-2 could interact with NO produced by phosphorylated eNOS whose expression is increased in GO orbital adipocytes. The interaction of O2 •− and NO could lead to peroxynitrite production, modifying proteins and interfering with their function through tyrosine nitration (24).
The oxidative stress in orbital adipocytes of GO patients could also originate from local T3 deprivation. Indeed, thyroid hormones seem to reduce the oxidative stress, as demonstrated in a rat model of myocardial infarction (25). Furthermore, it has been highlighted that a decrease in local thyroid hormone may directly decrease Cav-1 expression during postnatal development in rat heart (26) and also Glut-4 expression in various tissues. Indeed, studies show evidence of a positive regulation of Glut-4 expression and translocation to the plasma membrane in the heart, brown adipose tissue, and skeletal muscle of rat (27,28). Therefore, these downregulations of Cav-1 and Glut-4 induced by local T3 deprivation could, in Graves' disease pathogenesis, impair insulin sensitivity and glucose uptake as well. This could explain why GO is more frequent and severe if patients with Graves' disease have type 2 diabetes (29).
Our study shows a correlation between the decrease in local T3 concentrations and the increase of DIO3 in the cytoplasm of orbital adipocytes. This suggests that DIO3, acting as a major inactivator of thyroid hormone action (12), could play an important role in the genesis of the aforementioned oxidative stress.
This process, involving a heightened production of ROS, may eventually lead to an overproduction of proinflammatory cytokines. Indeed, a study by Seo et al. showed that oxidative stress in pancreatic acinar cells could act through NF-κB to induce a rise in the levels of proinflammatory cytokines (30). It is then possible that proinflammatory cytokines such as IL-6, IFNγ, or IL-1 could originate from the same pathway in orbital adipose tissues from GO patients. IL-6 production has been shown to be increased in nonthyroidal illness syndrome (31). Hence, we speculate that the diminished local supply of T3 observed in GO orbital adipocytes could result in an increase of IL-6 through the pathway explained above. Other cytokines could be involved in this pathogenesis such as IL-1 and IL-2 and also IFNγ, which might constitute the main element of an important feedback loop (3). IFNγ increases NOX-2 expression during the maturation phase of myeloid cells (32) and, as a consequence, we speculate the overproduction of NOX-2 that we have observed in orbital fat might be intensified through the same pathway.
Our study findings suggest prospective several treatment targets for GO. First, is the use of antioxidants, supported by the significant decrease of catalase that we observed in GO orbital adipocytes. Numerous studies have already shown the benefits of selenium supplementation on the quality of life and the slowing of the progression of disease in patients with mild GO (33 –35). In addition, it appears that a treatment combining insulin and selenium could restore the translocation of Glut-4 vesicles to the plasma membrane in cardiac muscle of diabetic rats and therefore counteract the glucose deprivation, which was leading to oxidative stress (36). Given their inflammation regulatory properties, corticosteroids could be used to break the proinflammatory cytokines feedback loops and have already proven to be effective and safe (37). NOX-2 inhibitors are also a very promising therapeutic alternative to prevent the progression of GO. It has been described that not only could NOX-2 inhibitors slow down the production of ROS, but they could also induce an attenuated inflammatory response (38,39). Finally, the pivotal proteins responsible for oxidative stress in GO adipocytes are Cav-1 and DIO3. Thus, these hypoxia-sensitive proteins could be considered as very interesting potential therapeutic targets.
Footnotes
Acknowledgments
The authors wish to thank Marc de Bournonville and Christine de Ville de Goyet for their technical help.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received.
Supplementary Material
Supplementary Figure S1