
Abstract
Background:
Thyroid hormone (triiodothyronine [T3]) plays an important role in regulating vertebrate developmental, cellular, and metabolic processes via T3 receptor (TR). Liganded TR recruit coactivator complexes that include steroid receptor coactivators (SRC1, SRC2 or SRC3), which are histone acetyltransferases, to T3-responsive promoters. The functions of endogenous coactivators during T3-dependent mammalian adult organ development remain largely unclear, in part, due to the difficulty to access and manipulate late-stage embryos and neonates. We use Xenopus metamorphosis as a model for postembryonic development in vertebrates. This process is controlled by T3, involves drastic changes in every organ/tissue, and can be easily manipulated. We have previously found that SRC3 was upregulated in the intestine during amphibian metamorphosis.
Methods:
To determine the function of endogenous SRC3 during intestinal remodeling, we have generated Xenopus tropicalis animals lacking a functional SRC3 gene and analyzed the resulting phenotype.
Results:
Although removing SRC3 had no apparent effect on external development and animal gross morphology, the SRC3 (−/−) tadpoles displayed a reduction in the acetylation of histone H4 in the intestine compared with that in wild-type animals. Further, the expression of TR target genes was also reduced in SRC3 (−/−) tadpoles during intestinal remodeling. Importantly, SRC3 (−/−) tadpoles had inhibited/delayed intestinal remodeling during natural and T3-induced metamorphosis, including reduced adult intestinal stem cell proliferation and apoptosis of larval epithelial cells.
Conclusion:
Our results, thus, demonstrate that SRC3 is a critical component of the TR-signaling pathway in vivo during intestinal remodeling.
Introduction
Thyroid hormone (triiodothyronine [T3])
When T3 is present, TR/RXR heterodimers recruit coactivators to induce gene expression (8 –10). Coactivators for TR include histone acetyltransferases (HATs) such as steroid receptor coactivators (SRCs) (11 –15) and p300/CREB-binding protein (CBP) (16,17) and the histone methyltransferases such as PRMT1 and CARM1 (18,19). The SRC family contains three isoforms SRC-1, SRC-2, and SRC-3 (20). They bind to TR and other nuclear receptors in an agonist-dependent manner through LXXLL (L, Leucine; X, any amino acid) motifs, and they help to recruit other coactivators such as p300/CBP (16,21 –23) into large complexes that can modify histones. These coactivators possess intrinsic HAT activity, and thus may regulate target gene expression in part through local histone acetylation. However, it remains unclear whether and how TR utilizes these coactivators in vivo, especially during vertebrate development, in part due to the difficulty to study this in mammals as the embryos and neonates depend on maternal supply for survival.
Because anuran amphibians develop externally in a biphasic process that is independent of maternal influence, we have utilized the T3-dependent metamorphosis in Xenopus tropicalis as a model to study the role of coactivators in TR function in development (24). During the second phase of anuran development, the tadpole undergoes metamorphosis to become a frog in a process that affects essentially all organs/tissues and can be easily manipulated by controlling the availability of T3 to tadpole rearing water (8,25,26). Notably, intestinal metamorphosis involves T3-dependent de novo formation of adult stem cells and mimics intestinal maturation during postembryonic mouse development (27). Thus, anuran metamorphosis is an advantageous and unique model to study not only how T3 regulates postembryonic vertebrate development but also how adult organ-specific stem cells are formed during vertebrate development, a critical problem that has been difficult to study in other species, including mammals.
Using the Xenopus model, we have previously reported that SRC3 expression in the intestine peaks at a climax of metamorphosis (stage 62) when larval epithelium is almost completely replaced by the newly formed, proliferating adult epithelial stem cells (28). Further, transgenic expression of a dominant negative SRC3 inhibits metamorphosis by blocking the activation of T3-responsive genes, demonstrating that coactivator recruitment is required for the developmental effects of T3 in vivo (29). However, the function of endogenous SRC3 during intestinal remodeling is still unclear. Here, we used the recently developed CRISPR-mediated gene knockout technology to test the hypothesis that endogenous SRC3 regulates intestinal remodeling, including the formation/proliferation of adult stem cells, by functioning as a TR coactivator.
Materials and Methods
All experiments involving X. tropicalis animals were carried out as approved by the National Institute of Child Health and Human Development Animal Use and Care Committee. The details are in the Supplementary Materials and Methods (Supplementary Data).
Results
Disrupting SRC3 leads to reduction in histone acetylation without apparent effect on overall animal development
We designed an SRC3-specific gRNA targeting exon 1 of X. tropicalis SRC3 (Fig. 1A) and microinjected it together with Cas9 into fertilized egg to obtain mosaic mutant animals. After sex maturation, genomic DNA was extracted from skin to determine the out-of-frame mutation rate induced by the Cas9 RNA-guided endonuclease in the gRNA targeted region in SRC3 by using a color assay (30). Adult frogs with high rates of out-of-frame mutations (30% or more) were thus identified (data not shown), and they were crossed among each other to generate F1 animals. At stage 52 or other indicated stages, we extracted genomic DNA from tail clips and genotyped F1 tadpoles by sequencing to identify tadpoles with one wild type (WT) SRC3 allele and an out-of-frame mutation in the other SRC allele, that is, heterozygous mutants or SRC3 (+/−) tadpoles, and tadpoles with an out-of-frame mutation in each of the two SRC3 alleles, that is, homozygous mutants or SRC3 (−/−) tadpoles (Fig. 1B, C). To verify SRC3 knockout, Western blot analysis of the whole body protein samples from WT, SRC3 (+/−), and SRC3 (−/−) tadpoles at stage 46 was performed. The expected SRC3 protein of 152 kDa was detected in WT tadpoles, and consistent with the mutation in one SRC3 allele, the signal intensity in SRC3 (+/−) was weaker than that in WT (Fig. 1C). Importantly, SRC3 (−/−) tadpoles lack any detectable SRC3 protein (Fig. 1C). Further, when we analyzed acetylated histone H4 in whole animals, we observed that its level was reduced in homozygous SRC3 knockout tadpoles, although no significant difference was observed between WT and SRC3 (+/−) tadpoles (Fig. 1D), consistent with the fact that SRC3 is an HAT.
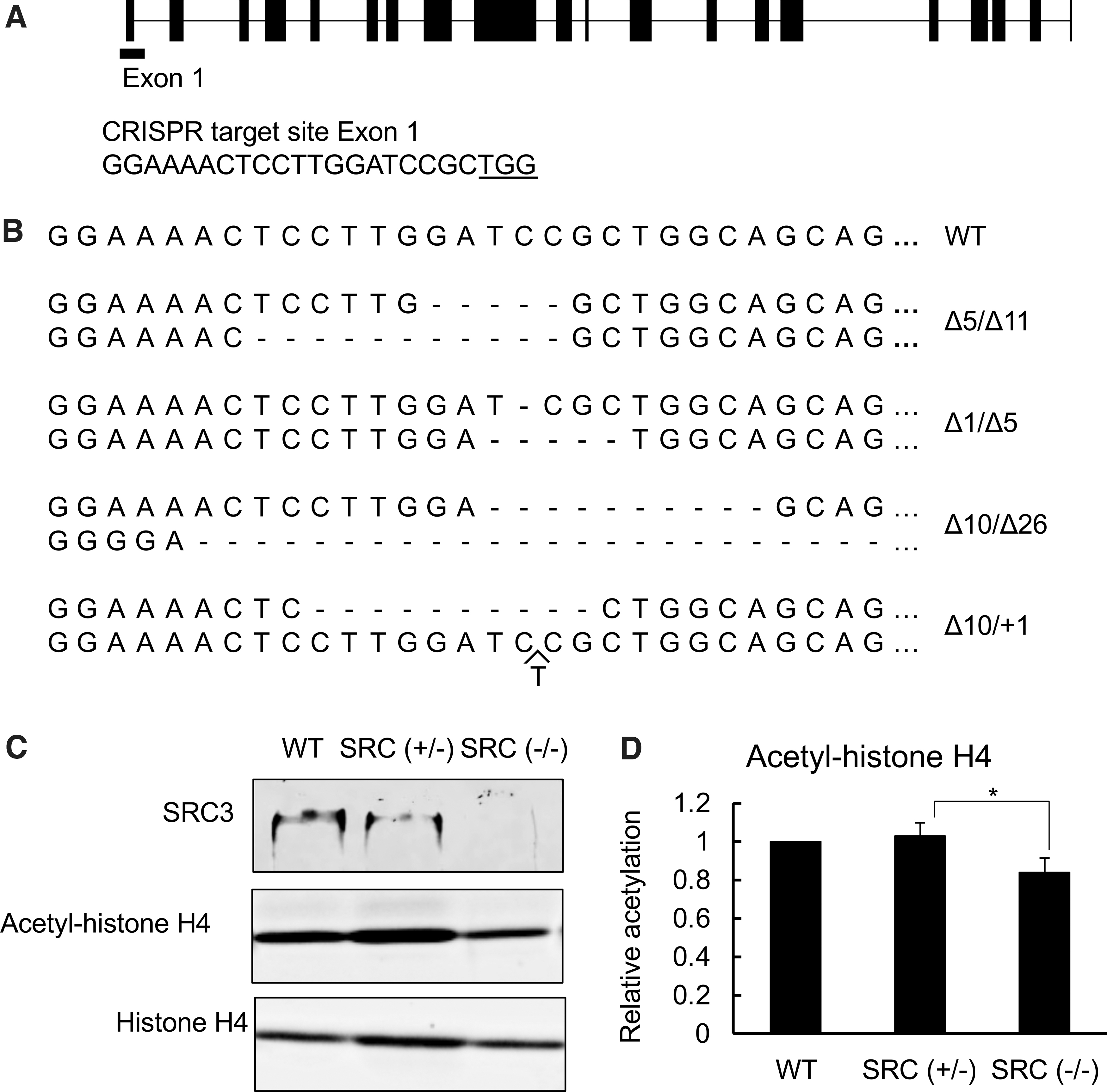
SRC3 knockout reduces histone H4 acetylation. (
To determine the effect of SRC3 knockout, we measured the developmental rate of WT and SRC3 mutant tadpoles. We recorded the time for the animals to develop from fertilization to stage 54 (the onset of metamorphosis), or stage 58 (the beginning of metamorphic climax), and the time from stage 58 to stage 66 (the end of metamorphosis). We found that there was no difference among the three genotypes, WT, SRC3 (+/−), and SRC3 (−/−), for all periods measured (Supplementary Fig. S1), suggesting that SRC3 knockout does not affect gross animal development, at least based on external morphological criteria, possibly due to redundancy among three SRCs.
SRC3 is important for intestinal remodeling during natural metamorphosis
We next investigated whether SRC3 knockout had any effect on the intestine during metamorphosis. Intestinal remodeling involves near complete degeneration of the larval epithelium with a single epithelial fold and de novo development of a multiply folded adult epithelium via the formation of adult epithelial stem cells and their subsequent proliferation and differentiation, accompanied by drastic shortening of the length of the intestine. Thus, we first measured intestine length during metamorphosis. Since individual animals of the same age and/or stage can vary in size, we normalized the length of the intestine against the length of the body (from tip of the head to the end of the belly). As expected, we observed that the length of the WT intestine was shortened by more than two-fold by the climax of metamorphosis (stage 62) and a little further by the end of metamorphosis (stage 66) (Fig. 2A). Heterozygous SRC3 knockout animals behaved similarly. However, the homozygous SRC3 knockout animals had a longer intestine compared with the other two genotypes at the end of metamorphosis, suggesting that SRC3 knockout delayed or inhibited intestinal remodeling during natural metamorphosis.
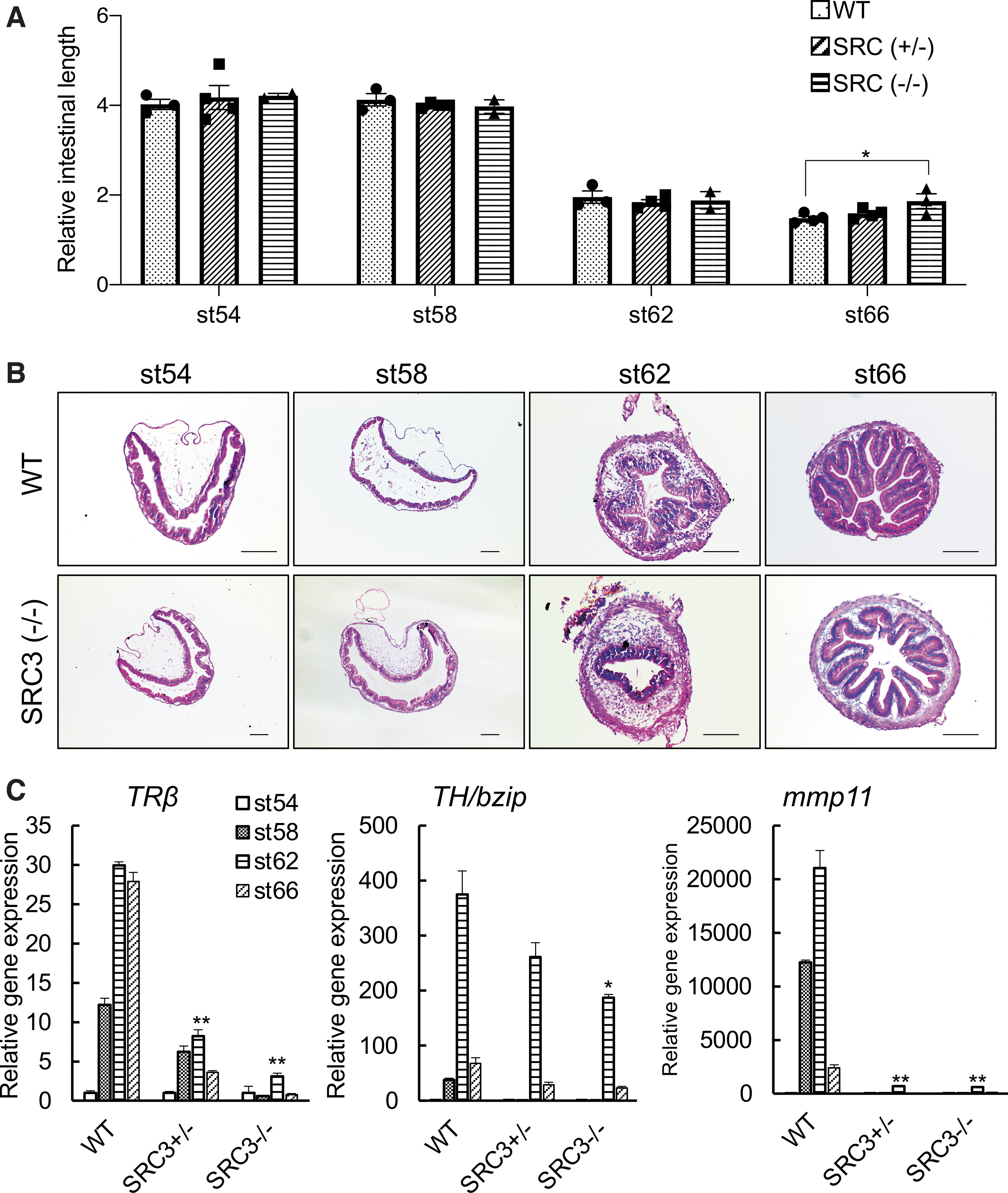
SRC3 knockout reduces TR target gene expression in the intestine and delays intestinal remodeling during metamorphosis. (
To investigate whether SRC3 knockout affected cellular transformation, we stained cross-sections of the intestine from WT and SRC3 (−/−) tadpoles at different stages. The result showed that at stage 62, numerous adult intestinal epithelial folds were formed in the WT but not SRC3 (−/−) tadpoles (Fig. 2B), indicating a delay/inhibition in adult intestinal development by the SRC3 knockout. Further, both epithelial cell death and proliferation were easily detected at the climax of metamorphosis (stage 62) (Supplementary Fig. S2) when drastic intestinal remodeling took place in the WT intestine. The signals were much less in SRC3 (−/−) tadpoles, supporting an important role of SRC3 during intestinal remodeling.
To determine whether SRC3 affected intestinal metamorphosis via functioning as a TR coactivator, we analyzed the expression of TR target genes during intestinal remodeling. We chose three well-known, direct, and ubiquitous T3-responsive genes, TRβ (31), TH/bzip (32), and mmp11 (33). As shown in Figure 2C, SRC3 knockout reduced the upregulation of all three genes in the intestine during natural metamorphosis, with stronger effect by the homozygous knockout compared with heterozygous knockout, suggesting that SRC3 functions as a TR coactivator during intestinal remodeling.
SRC3 knockout inhibits gene activation and intestinal remodeling during T3-induced metamorphosis
Natural metamorphosis is a lengthy process, requiring four weeks or longer for the tadpole to develop from stage 54 (onset of metamorphosis) to stage 66 (end of metamorphosis). This may enable various endogenous compensatory mechanisms to reduce the potential effects of SRC3 knockout, consequently leading to relatively small effects due to the knockout. In addition, there is also considerable heterogeneity among sibling animals during such a lengthy developmental process, making it difficult to measure knockout effect. To overcome these issues, we next took advantage of the fact that many processes during anuran metamorphosis, including intestinal remodeling, can be induced by treating premetamorphic tadpoles with physiological concentrations of T3 for as little as three days. We first treated premetamorphic WT and SRC3 knockout tadpoles at stage 54 with 10 nM T3 for 18 hours and isolated total RNA from the intestine for analysis of the three known TR target genes, as studied earlier. All three were strongly induced by T3 in WT animals (Fig. 3A). Their induction was reduced in SRC3 (+/−) and SRC3 (−/−) animals, with a much bigger effect seen in homozygous knockout animals (Fig. 3A).
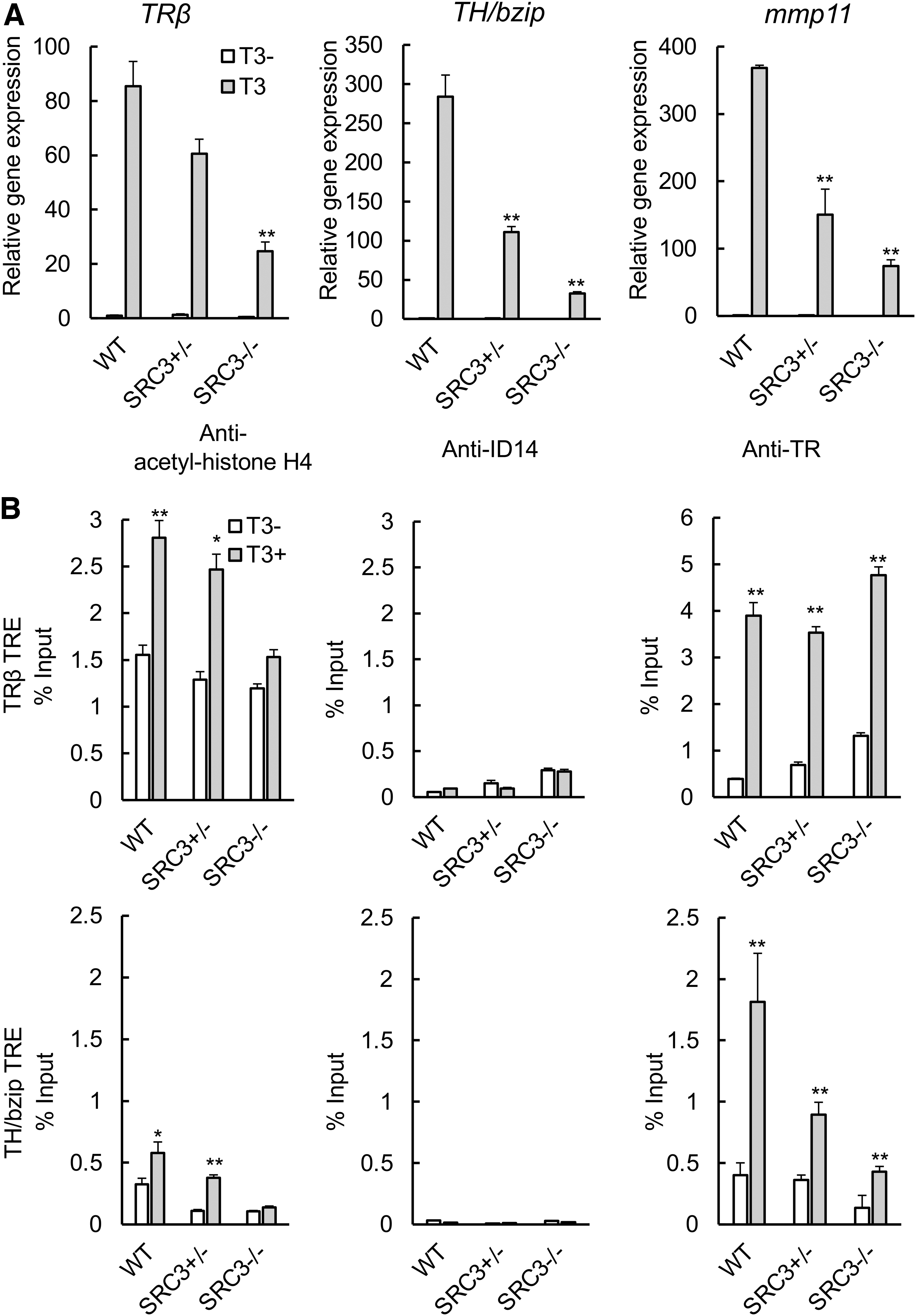
SRC3 knockout reduces T3-induced target gene expression, accompanied by reduced histone H4 acetylation but without affecting TR binding. (
To investigate how SRC3 knockout affected TR target genes, we carried out ChIP assays to measure the binding of TR to and association of histone H4 acetylation with the TREs of two well-known target genes, TRβ and TH/bzip, in the animals. As shown in Figure 3B, H4 acetylation was specifically increased by T3 treatment at the TRE regions of both genes but not the negative control region, the exon 5 of TRβ gene that lacks any TRE (Supplementary Fig. S3), in the WT intestine. This increase was reduced in the SRC3 (+/−) intestine and essentially abolished in the SRC3 (−/−) intestine (Fig. 3B). As a control, we also carried out ChIP assay by using an antibody against an extracellular protein encoded by X. laevis gene ID14 and observed only low background signals (Fig. 3B and Supplementary Fig. S3). We further analyzed the binding of TR to the TREs and found that expectedly, TR was specifically bound to the TREs of both genes but not the exon 5 of the TRβ gene in the intestine and this binding was not affected by SRC3 knockout (Fig. 3B and Supplementary Fig. S3). These results suggest that SRC3 is an important TR coactivator in the intestine.
To study the effect of SRC3 (−/−) on T3-induced metamorphosis, WT and knockout siblings at premetamorphic stage 54 were treated with 10 nM T3 for up to three days. As shown in Figure 4A, three day of T3 treatment led to dramatic metamorphic changes in the WT animals, most noticeably limb development (digit formation) and reshaping of the head structure, which is, in part, due to resorption of the gills (Fig. 4A and Supplementary Fig. S4). In SRC3 knockout animals, T3-induced limb development was inhibited (Fig. 4A). In addition, structural changes in the head region, including gill resorption, was also reduced compared with the WT ones (Fig. 4A and Supplementary Fig. S4). Thus, SRC3 is important for T3-induced external metamorphic changes.
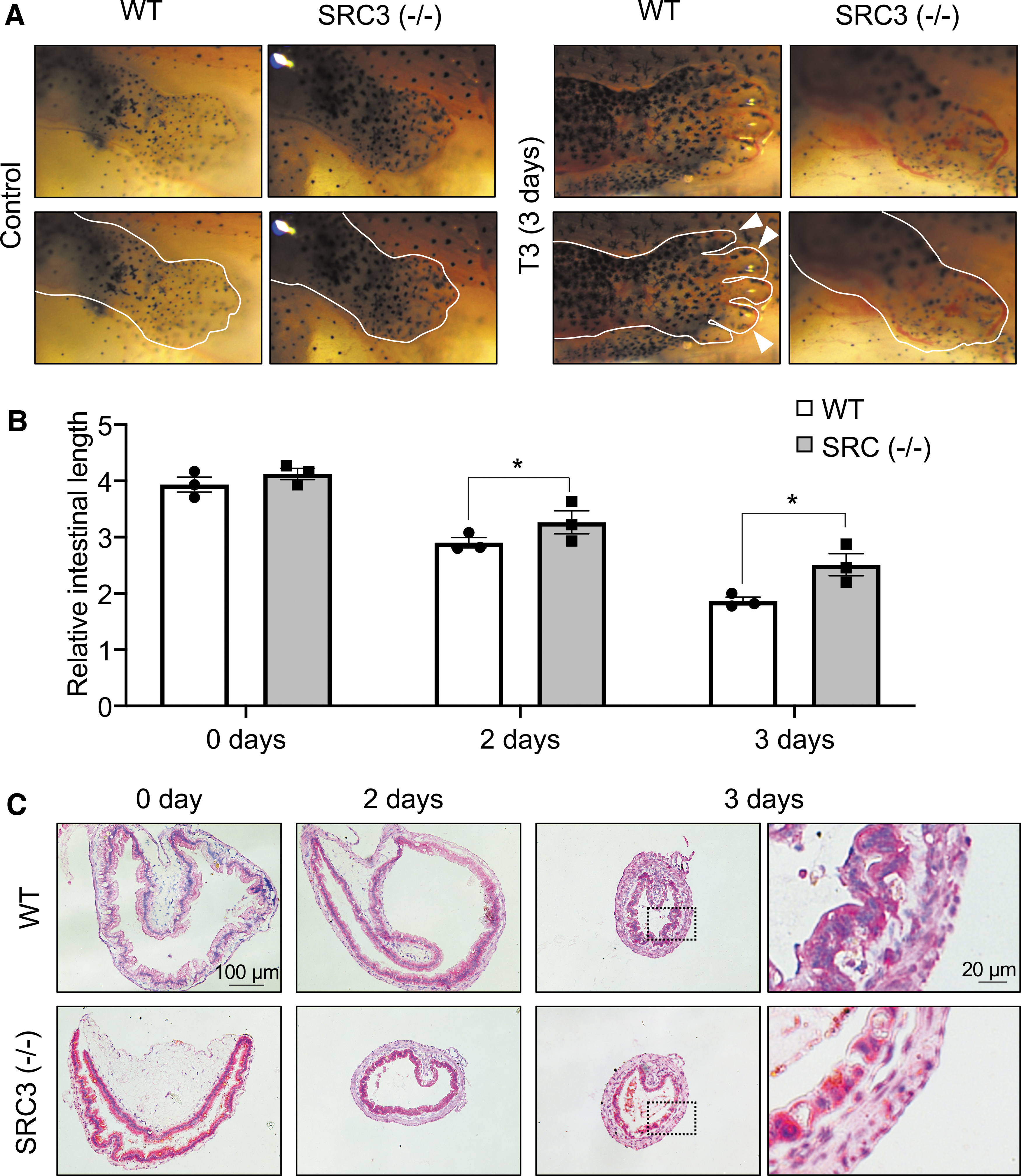
SRC3 knockout inhibits T3-induced limb development and intestinal remodeling. (
We next first measured intestine length. As expected, T3 induced intestine shortening in WT animals within two days of treatment (Fig. 4B). Importantly, homozygous SRC3 knockout reduced this shortening after two or three days of T3 treatment (Fig. 4B). We further stained intestinal cross-sections and found that after three days of T3 treatment, numerous adult epithelial folds were formed in WT but SRC3 (−/−) tadpoles (Fig. 4C), suggesting that SRC3 is important for T3-induced intestinal remodeling.
Intestinal metamorphosis involves near total degeneration of larval epithelium via apoptosis and development of adult epithelium via de novo formation of adult epithelial stem cells followed by their proliferation and differentiation. Thus, we next analyzed cell proliferation with EdU labeling. The results showed that expectedly, intestinal epithelial cell proliferation as detected by EdU labeling was induced during T3-induced metamorphosis in WT animals (Fig. 5). In SRC3 knockout animals, in contrast, T3-induced cell proliferation was drastically reduced (Fig. 5A, B). This was further confirmed by quantification of the percentage of EdU-positive cells (Fig. 5C).

Adult stem cell proliferation is reduced in T3-treated SRC3 knockout tadpoles. (
We also investigated apoptosis in the intestine by using terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL). We found that after two days of T3 treatment, high levels of apoptotic signals were present in the WT intestinal epithelium (Fig. 6A, B) and that the signals decreased in the WT intestine after three days of treatment (Fig. 6C) when adult cell proliferation took place (Fig. 5). In SRC3 (−/−) animals, T3-induced apoptosis in the intestinal epithelium was much lower than that in WT animals after two days of T3 treatment (Fig. 6A, B). This was again confirmed by quantifying the TUNEL signals (Fig. 6C). Apoptosis was higher in SRC3 (−/−) animals compared with WT animals after three days of T3 treatment (Fig. 6C), suggesting that T3-induced apoptosis was delayed in SRC3 (−/−) tadpoles.
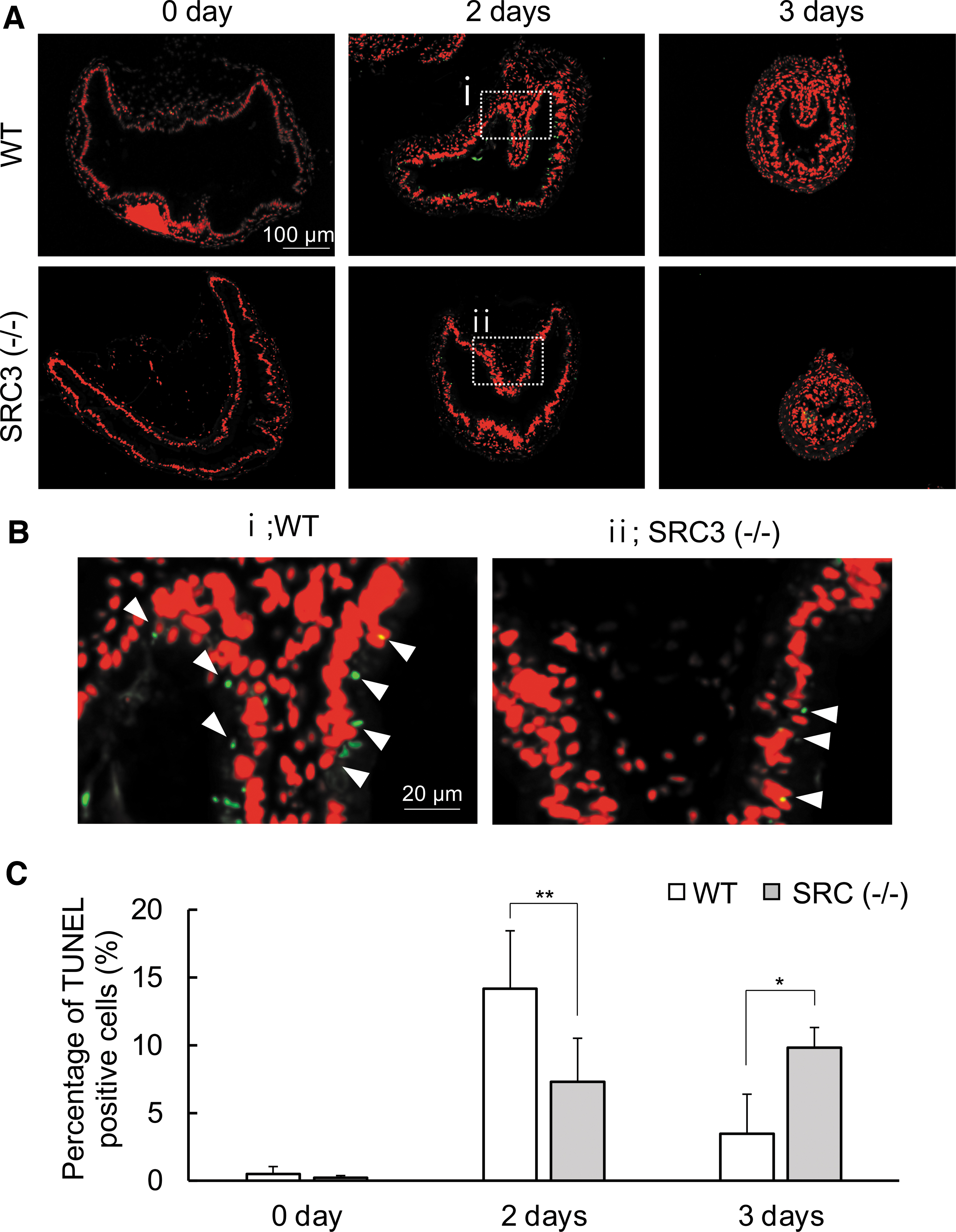
Apoptosis is reduced in SRC3 knockout tadpoles. (
Discussion
The T3-dependent Xenopus intestinal remodeling during metamorphosis as a model of mammalian postembryonic development has provided new insights on the de novo formation of adult organ-specific stem cells in vertebrates (34). Importantly, this process is similar to the development of mammalian adult intestine around the neonatal period (27). Earlier studies of Xenopus intestinal metamorphosis have identified many T3 response genes that are likely involved during Xenopus intestinal remodeling. Among them is the TR-coactivator SRC3 (28,29,35). There are many known TR-coactivators, which often function as coactivators for other nuclear hormone receptors as well. Gene knockout studies in mouse have shown that removing coactivators causes various developmental defects (36). However, it is difficult to determine the molecular mechanisms since the coactivators are often involved in gene regulation by many nuclear hormone receptors and other transcription factors. This is further compounded by the difficulty to manipulate mammalian postembryonic development. Taking advantage of many unique properties of anuran metamorphosis, our results here thus provide for the first time in vivo evidence to support an important role of an endogenous coactivator, the HAT SRC3, in intestinal metamorphosis by functioning as a TR coactivator and increasing local histone acetylation either directly through its own acetyltransferase activity or indirectly through recruitments of other acetyltransferases such as CBP/P300 as a TR-coactivator complex.
Intestinal remodeling during Xenopus metamorphosis involves near-complete degeneration of the larval epithelial through apoptosis and de novo formation of adult epithelial stem cells in a process totally regulated by T3 (37 –40). Earlier studies have shown that the HAT SRC3 is upregulated during Xenopus intestinal metamorphosis and can function as a TR coactivator, suggesting that SRC3 is upregulated by T3 and, in turn, feeds back positively by functioning as a TR coactivator to further enhance T3 signaling during intestinal metamorphosis. Consistently, we have shown here that SRC3 knockout leads to a delay in intestinal remodeling with a reduction of intestinal stem cell proliferation and apoptosis of larval epithelial cells during both natural and T3-induced metamorphosis. Mechanistically, we found that the expression of TR response genes is strongly reduced in SRC3 knockout tadpoles and that SRC3 knockout results in reduced histone acetylation at a TR response gene without affecting TR binding to the TRE. Thus, endogenous SRC3 appears to function as a TR coactivator to increase local histone acetylation to enhance the promoter activity, thereby ensuring proper intestinal remodeling during metamorphosis. It is worth noting that SRC3 is likely recruited to TR target genes during metamorphosis as a large complex containing P300/CBP (29,35), which have stronger HAT activities. Thus, the effects of SRC3 knockout on local histone acetylation might be due to the failure to recruit P300/CBP via the SRC3-P300/CBP complex.
Unliganded TR binding to the TRE region recruits corepressors such as N-CoR and SMRT to a target promoter, whereas ligand-bound TR recruits co-activators such as SRC1, SRC2, and SRC3. Among the three members of the SRC family of coactivators, SRC3 expression has been shown to be drastically increased during intestinal metamorphosis (28). Thus, somewhat surprisingly, SRC3 knockout has little effect on overall tadpole development, at least based on external morphology. Even for the intestine, the effects of SRC3 knockout appear to be fairly modest during natural metamorphosis, with a slightly longer intestine at the end of metamorphosis, suggesting a slight delay/inhibition of the remodeling process. Such mild effects of the knockout likely reflect potential redundant functions among the three SRC family members, as also suggested by genetic studies in mouse. Total knockout of mouse SRC3 also has a relatively mild effect, with the resulting animals having retarded growth, delayed puberty, decreased reproductive function, and blunted mammary gland development (36). In addition, the mice lacking SRC1 have even less defect, although having blunted mammary gland development (41). On the other hand, double knockout of SRC1 and SRC3 leads to embryonic lethality (42), supporting compensatory effects of the two members. Thus, it would be interesting to investigate the role of other SRC family members during anuran metamorphosis.
By using T3-induced metamorphosis under physiological levels of T3, we were able to observe much bigger effects of the SRC3 knockout on metamorphosis, based on both external morphology (limb development and gill resorption) and intestinal remodeling. This was likely due to the fact that T3-induced metamorphosis occurs in a much shorter time frame and more synchronous manner, thus reducing the variations among animals and the likelihood of compensatory events that knockout animals may have in a much longer natural metamorphic process. Our findings here, therefore, not only reveal, for the first time, an important role of an endogenous TR coactivator in T3 signaling and the formation/proliferation of adult organ-specific stem cell, but also further highlight the advantages of the anuran metamorphosis for studying adult organ development/maturation and gene function during postembryonic development.
Footnotes
Acknowledgment
The authors thank Nga Luu for experimental help and laboratory management.
Author Contributions
Y.T. and Y.B.-S. designed the research plan, L.B. generated the F0 mutant animals, and Y.T. carried out the research and data analyses. All authors participated in the article preparation and approved the final version of the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the intramural Research Program of NICHD, NIH. Y.T. was supported in part by a Japan Society for the Promotion of Science Research Fellowship for Japanese Biomedical and Behavioral Researchers at the National Institutes of Health (No. 29-71715).
Supplementary Material
Supplementary Data
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4