
Abstract
Background:
The fetal hypothalamic–pituitary–adrenal (HPA) axis plays a key role in the control of parturition and maturation of organ systems in preparation for birth. In hypothyroid fetuses, gestational length may be prolonged and maturational processes delayed. The extent to which the effects of thyroid hormone deficiency in utero on the timing of fetal maturation and parturition are mediated by changes to the structure and function of the fetal HPA axis is unknown.
Methods:
In twin sheep pregnancies where one fetus was thyroidectomized and the other sham-operated, this study investigated the effect of hypothyroidism on circulating concentrations of adrenocorticotrophic hormone (ACTH) and cortisol, and the structure and secretory capacity of the anterior pituitary and adrenal glands. The relative population of pituitary corticotrophs and the masses of the adrenal zones were assessed by immunohistochemical and stereological techniques. Adrenal mRNA abundances of key steroidogenic enzymes and growth factors were examined by quantitative polymerase chain reaction.
Results:
Hypothyroidism in utero reduced plasma concentrations of ACTH and cortisol. In thyroid-deficient fetuses, the mass of corticotrophs in the anterior pituitary gland was unexpectedly increased, while the mass of the zona fasciculata and its proportion of the adrenal gland were decreased. These structural changes were associated with lower adrenocortical mRNA abundances of insulin-like growth factor (IGF)-I and its receptor, and key steroidogenic enzymes responsible for glucocorticoid synthesis. The relative mass of the adrenal medulla and its proportion of the adrenal gland were increased by thyroid hormone deficiency in utero, without any change in expression of phenylethanolamine N-methyltransferase or the IGF system.
Conclusions:
Thyroid hormones are important regulators of the structure and secretory capacity of the pituitary–adrenal axis before birth. In hypothyroid fetuses, low plasma cortisol may be due to impaired adrenocortical growth and steroidogenic enzyme expression, secondary to low circulating ACTH concentration. Greater corticotroph population in the anterior pituitary gland of the hypothyroid fetus indicates compensatory cell proliferation and that there may be abnormal corticotroph capacity for ACTH synthesis and/or impaired hypothalamic input. Suppression of the development of the fetal HPA axis by thyroid hormone deficiency may contribute to the delay in fetal maturation and delivery observed in hypothyroid offspring.
Introduction
Birth, and the successful transition from the intra- to extrauterine environment, is arguably the most important physiological event in life. Survival of the offspring depends on the maturation of a wide range of fetal tissues toward term, including the lungs, gastrointestinal tract, and brown adipose tissue (1). Developmental changes in these fetal organs are essential to activate pulmonary gas exchange, enteral nutrient uptake, and thermoregulation, all physiological processes that are required for the first time at birth. The mechanisms that induce fetal maturation are closely linked to the mechanisms of parturition to ensure the delivery of a viable offspring.
The fetal hypothalamic–pituitary–adrenal (HPA) axis is a key regulator of fetal maturation and the onset of parturition (2). Hypothalamic neurons release corticotrophin-releasing hormone (CRH) and arginine vasopressin (AVP), which stimulate corticotrophs in the anterior pituitary gland to secrete the adrenocorticotrophic hormone (ACTH). In the sheep fetus, there are changes in corticotroph ultrastructure, the molecular capacity to synthesize ACTH, and responsiveness to CRH and AVP with increasing gestational age (3,4). Concentrations of ACTH rise in the fetal circulation toward term and promote the growth and secretory capacity of the adrenal gland. In the zona fasciculata of the adrenal cortex, ACTH binds its receptor (MC2R) to promote the expression of proteins that are responsible for the synthesis of adrenal hormones from cholesterol. The steroidogenic acute regulatory protein (StAR) transports cholesterol from the outer to the inner mitochondrial membrane and a series of steroidogenic enzymes convert cholesterol into glucocorticoids (cholesterol side chain cleavage, CYP11A1; 17α-hydroxylase, CYP17; 3β-hydroxysteroid dehydrogenase, 3βHSD; 21-hydroxylase, CYP21; 11β-hydroxylase, CYP11B1). Increasing plasma concentrations of adrenal hormones stimulate the process of parturition (glucocorticoids in sheep and androgens in humans) and maturational events in key tissues that are essential for neonatal survival (glucocorticoids in both species) (5,6).
A range of experimental and clinical studies have shown that thyroid hormones also have an important role in the control of fetal maturation and the timing of birth. In the sheep fetus, hypothyroidism prolongs gestation and causes abnormal growth and development of a range of fetal organs (7,8). These effects may be a direct consequence of thyroid hormone deficiency and/or may occur secondary to changes in other hormones in the hypothyroid fetus, including insulin, leptin, and insulin-like growth factors (IGF; 8–10). Prolonged gestational length is also observed in human pregnancy where the fetus has congenital hypothyroidism (CH), that affects 1:2000 human births worldwide. Pregnancies complicated by CH are more likely to extend past 40 weeks of gestation (35–48% CH vs. 15–19% control) and to require induction (11,12). Furthermore, even in studies where infants with CH, prematurity and low birth weight are excluded, high neonatal thyrotropin concentration, indicative of low systemic thyroid hormone levels, is associated with a longer pregnancy (13). Cases of CH and dysmaturity syndrome have also been reported in horses in Canada and Europe; gestational length is increased and the hypothyroid foals show skeletomuscular abnormalities, poor temperature control, and reduced survival (14). The extent to which thyroid hormones influence fetal maturation and gestational length via development of the HPA axis before birth, however, is unknown.
In neonatal and adult rats, hypothyroidism induced by surgical and pharmacological methods decreases the expression of components of the HPA axis involved in the production of ACTH and glucocorticoids (15,16). Little is known, however, about the consequences of thyroid hormone deficiency before birth for the structure and function of the developing pituitary and adrenal gland, especially in a species such as the sheep that has a similar pattern of maturation of thyroid hormone activity to human infants (8). The aims of this study were, therefore, to determine the effect of fetal hypothyroidism on (a) circulating concentrations of ACTH and cortisol, (b) the corticotroph population in the anterior pituitary gland, (c) the zonal structure of the adrenal gland, and (d) the adrenal mRNA abundance of key genes responsible for glucocorticoid synthesis. It was hypothesized that thyroid hormone deficiency in the sheep fetus would lead to lower circulating ACTH due to a reduction in the number of corticotrophs in the anterior pituitary gland and, as a consequence, lower plasma cortisol due to impaired growth and steroidogenic capacity of the adrenal gland.
Materials and Methods
Animals
All surgical and experimental procedures were carried out in accordance with UK Home Office legislation and the Animals (Scientific Procedures) Act 1986, after approval by the Animal Welfare and Ethical Review Body, University of Cambridge, United Kingdom. Sixteen Welsh Mountain pregnant ewes of known gestational age and carrying twin fetuses were used in this study. The ewes were housed in individual pens and were maintained on 200 g/day concentrates (14% crude protein, energy content 12 MJ/kg dry matter; H & C Beart Ltd., Stowbridge, United Kingdom) with hay and water ad libitum and access to a salt block. Food, but not water, was withheld from the ewes for 18–24 hours before surgery.
Experimental procedures
Under general anesthesia (2% isoflurane in O2-N2O) and at 102–110 days of gestation (dGA; term ∼145 ± 2 days), the twin fetuses of each ewe underwent either surgical removal of the thyroid gland (thyroidectomy [TX]) or a sham operation in which the thyroid gland was exposed but not removed (sham), as previously described (7). At surgery, antibiotics were administered to each fetus intravenously and into the amniotic cavity of each fetus (total 600 mg benzylpenicillin in 5 mL of 0.9% saline: Crystapen, Schering-Plough, Welwyn Garden City, United Kingdom). The ewes were treated with antibiotics (30 mg/kg procaine benzylpenicillin I.M.; Depocillin, Intervet UK Ltd., Milton Keynes, United Kingdom) immediately before the start of surgery and daily for three days thereafter. The animals were monitored over the recovery period and resumed normal feeding within 24 hours of surgery.
Between 140 and 145 dGA, the fetuses were delivered by Caesarean section under general anesthesia (20 mg/kg maternal body weight sodium pentobarbitone I.V.). Blood samples were collected by venipuncture of the umbilical artery into tubes containing ethylenediamine tetraacetic acid. Each fetus was weighed and a variety of fetal organs, including the pituitary and adrenal glands, were dissected and weighed after administration of a lethal dose of barbiturate (200 mg/kg sodium pentobarbitone I.V.). One adrenal gland was snap frozen in liquid nitrogen and stored at −80°C for molecular analysis, and the other adrenal gland and the pituitary gland were immersion fixed whole in 4% paraformaldehyde (with 0.2% glutaraldehyde in 0.1 M phosphate buffer, pH 7.4) for 2 days for histological analysis. After washing in phosphate-buffered saline, the fixed adrenal and pituitary glands were processed and embedded in paraffin wax.
Plasma hormone measurements
Umbilical plasma triiodothyronine (T3) and thyroxine (T4) concentrations were determined by radioimmunoassay (RIA) kits (MP Biomedicals, Loughborough, United Kingdom); the intra-assay coefficients of variation were 3% and 5%, and the minimum levels of detection were 0.14 and 7.0 ng/mL, respectively. Plasma concentrations of ACTH, cortisol, and insulin were determined by using ELISA kits (ACTH 1–39; Demeditec Diagnostics GmbH, Kiel, Germany; cortisol: IBL International, Hamburg, Germany; insulin: Mercodia, Uppsala, Sweden); the intra-assay coefficients of variation were all <10%, and the minimum levels of detection were 0.22 pg/mL, 2.5 ng/mL, and 0.025 ng/mL, respectively. Plasma leptin concentration was measured by RIA as previously described (17). The intra-assay coefficient of variation was 5%, and the minimum level of detection was 0.09 ng/mL.
Histology and immunohistochemistry
The fixed pituitary and adrenal glands were exhaustively sectioned at 5 μm. For the pituitary gland, ACTH-positive cells were identified in 10 equally spaced sections by immunohistochemistry using a rabbit polyclonal antibody against human ACTH (10 μg/mL; Bio-Rad Laboratories, Watford, United Kingdom). Detection was achieved by using the Vectastain Elite ABC kit (Vector Laboratories, Peterborough, United Kingdom) and diaminobenzidine, and hematoxylin was used to counter-stain the sections. All sections were scanned by using a NanoZoomer digital slide scanner (Hamamatsu Photonics, Welwyn Garden City, United Kingdom) to create digital images for analysis. Section images were analyzed by an observer blinded to the treatment group using NewCAST stereological software (Visiopharm, Hoersholm, Denmark). In each section of the pituitary gland, ∼5–10% of the tissue was sampled to estimate the fractional volume of anterior pituitary and, specifically, ACTH-positive pituitary cell types by point-counting and application of Cavalieri's principle (18). At least 150 points were counted for each variable. Absolute masses of the anterior pituitary gland and the corticotroph population were calculated by expressing the estimated fractional volume as a proportion of the total pituitary weight, and relative mass was calculated by dividing the absolute mass by fetal body weight at delivery.
For the adrenal gland, 8–10 equally spaced sections were stained with hematoxylin and eosin to distinguish the medulla and zones of the cortex. In each section of the adrenal gland, ∼10% of the tissue was sampled to determine the fractional volumes of the adrenal compartments. Using the point-counting method as detailed for the pituitary gland, the regions of the adrenal gland were classified into four different compartments: capsule, zona glomerulosa, zona fasciculata, and adrenal medulla. In sheep, the zona reticularis is not present in the adrenal gland until postnatal life. Absolute masses of the adrenal compartments were calculated by expressing the estimated fractional volumes as a proportion of the total adrenal weight, and relative mass was calculated by dividing the absolute mass by fetal body weight at delivery.
Quantification of adrenal mRNA abundance
Frozen adrenal glands were separated into cortex and medulla regions on the basis of appearance by using a dissecting microscope. Dissected tissue was homogenized in Lysing Matrix-D tubes (MP Biomedicals) by using a MagNA Lyser (Roche Diagnostics, Almere, The Netherlands). The RNA was extracted following the protocol of the RNeasy Plus Mini Kit (Qiagen, Manchester, United Kingdom), and RNA extraction yields were assessed by using a Nanodrop spectrophotometer (Thermo Fisher Scientific, Loughborough, United Kingdom).
Reverse transcription of the extracted mRNA was performed by using the High-Capacity cDNA Reverse Transcription Kit (Thermo Fisher Scientific). Negative controls were prepared where the multiscribe reverse transcriptase and RNase inhibitor were replaced with equal volumes of RNase-free water. Reverse transcription was carried out in the Gene Pro thermocycler (Bioer Ltd., Hangzhou, China) at 25°C for 10 minutes, 37°C for 120 minutes, and 85°C for 5 minutes.
Quantitative polymerase chain reaction (qPCR) was performed on cortex samples for the genes MC2R, StAR, CYP11A1, CYP17, 3βHSD, CYP21, and CYP11B1, and on medulla samples for the gene phenylethanolamine N-methyltransferase (PNMT: the enzyme that converts noradrenaline into adrenaline). Expression of IGFI, IGFII, IGF type 1 and type 2 receptors (IGF-1R, IGF-2R), and the housekeeping gene YWHAZ, was examined in both cortical and medullary regions. The sequences of the primers used are presented in Supplementary Table S1. The primer efficiencies were 83–115%, and all melting curves showed a single product with melting temperatures between 58°C and 62°C.
Each well of a 96-well PCR plate (STARLAB Ltd, Blakelands, United Kingdom) was loaded with sample cDNA, forward and reverse primers, RNase-free water, and MESA BLUE qPCR MasterMix Plus for SYBR® Assay No ROX (Eurogentec, Seraing, Belgium). Separate plates were used to measure mRNA levels for each gene, and each sample or standard was measured in duplicate. Samples were amplified for qPCR by using a DNA Engine Opticon 2 Sequence Detection System (Bio-Rad Laboratories) at 95°C for 10 minutes, followed by 40 cycles of 95°C for 15 seconds and 60°C for 1 minute, followed by a melting curve stage of 60–90°C and held for 10 seconds to read at every 1°C increment. Data were processed by using Opticon Monitor Version 3.1 (Bio-Rad Laboratories). The negative control samples did not generate any detectable amplicons, confirming the absence of genomic contamination. To compare mRNA abundance of target genes between the treatment groups, cycle thresholds (Ct) were expressed relative to YWHAZ and analyzed by using the delta-delta-Ct (ΔΔCt) method, as all standard curves were linear and parallel.
Statistical analyses
Data from the two treatment groups are presented as mean ± standard error of the mean. After assessment for normality by using the D'Agostino-Pearson test, data from the treatment groups were compared by the Student's unpaired t-test or Mann–Whitney test, as appropriate. Relationships between variables were determined by Pearson correlation using log10-transformed data where necessary. p < 0.05 was regarded as significant. Statistical tests were not performed on data that included values below the lower limit of assay detection (plasma T4 and T3 concentrations).
Results
Hypothyroidism in utero suppressed circulating ACTH and cortisol concentrations
Umbilical plasma T4 and T3 concentrations decreased to below the limits of assay detection in the TX fetuses (Table 1). Compared with the sham fetuses, the TX fetuses had significantly lower plasma ACTH and cortisol, and higher plasma insulin and leptin concentrations (p < 0.05; Table 1). When data available from all fetuses were combined, regardless of treatment group, negative correlations were observed between log10 plasma ACTH and both insulin (R = −0.64, p < 0.01, N = 16) and leptin concentrations (R = −0.81, p < 0.0005, N = 16).
Mean (±Standard Error of the Mean) Plasma Hormone Concentrations, and Body and Organ Weights, in Sham (n = 15) and Thyroidectomized (n = 12) Fetuses at 143 Days of Gestation
Significantly different from sham fetuses, p < 0.05; ND (limit of assay detection: T4 7.0 ng/mL, T3 0.14 ng/mL).
ACTH, adrenocorticotrophic hormone; ND, not detectable; T3, triiodothyronine; T4, thyroxine; TX, thyroidectomy.
Low plasma ACTH in hypothyroid fetuses was associated with an increase in the pituitary corticotroph population
In the TX compared with sham fetuses, body weight was reduced (p < 0.05; Table 1) and both absolute and relative weights of the total pituitary gland were increased (p < 0.005; Table 1). The fractional volume of corticotrophs in the anterior pituitary gland was reduced by hypothyroidism (sham 31.4% ± 1.2%, TX 27.1% ± 1.7%, p < 0.05; Fig. 1A); however, when expressed as absolute and relative masses, the populations of both corticotrophs (absolute mass: sham 25.0 ± 1.7 mg, TX 33.6 ± 1.9 mg; relative mass: sham 6.8 ± 0.4 mg/kg, TX 10.6 ± 0.5 mg/kg) and other pituitary cell types were greater in TX compared with sham fetuses (p < 0.005; Fig. 1B, C). The estimated absolute and relative weights of the anterior pituitary gland were also increased in the TX fetuses (p < 0.05; Fig. 1B). The relative total pituitary weight correlated negatively with log10 plasma ACTH (R = −0.58, p < 0.01, N = 21) and positively with plasma concentrations of insulin (R = 0.62, p < 0.05, N = 16) and leptin (R = 0.59, p < 0.05, N = 16). Plasma concentrations of insulin and leptin also correlated with the relative masses of the anterior pituitary (both R = 0.59, p < 0.05, N = 16), corticotrophs (insulin: R = 0.53, leptin: R = 0.56, p < 0.05, N = 16), and other pituitary cell types (insulin: R = 0.57, leptin: R = 0.55, p < 0.05, N = 16).

Mean (± SEM) populations of corticotrophs and other cell types in the anterior pituitary gland, expressed as (
Hypothyroidism in utero decreased zona fasciculata and increased medulla sizes in the fetal adrenal gland
There was no difference in total adrenal gland weight, expressed in absolute or relative terms, between the groups of TX and sham fetuses (Table 1). Hypothyroidism caused a decrease in zona fasciculata percentage volume and an increase in medulla percentage volume of the total adrenal gland (p < 0.05; Fig. 2A). When expressed as a percentage volume of the adrenal cortex (zona glomerulosa and zona fasciculata combined), the zona fasciculata in the TX fetuses was a smaller fraction of the adrenal cortex than in the sham fetuses (sham 86.4% ± 0.7%, TX 83.2% ± 0.8%, p < 0.01). The absolute mass of the zona fasciculata was lower, and the relative mass of the medulla was greater in the TX compared with sham fetuses (p < 0.05; Figs. 2B, C). There were no significant differences in the absolute (p = 0.07) or relative mass of the adrenal cortex between the TX and sham fetuses (Fig. 2B, C). Overall, log10 plasma cortisol concentration correlated positively with the absolute and relative masses of both the adrenal gland (R = 0.54 and R = 0.40, respectively, p < 0.05, N = 27) and the zona fasciculata (R = 0.67 and R = 0.64, respectively, p < 0.0005, N = 27), and negatively with the relative mass of the medulla (R = −0.46, p < 0.05, N = 27).
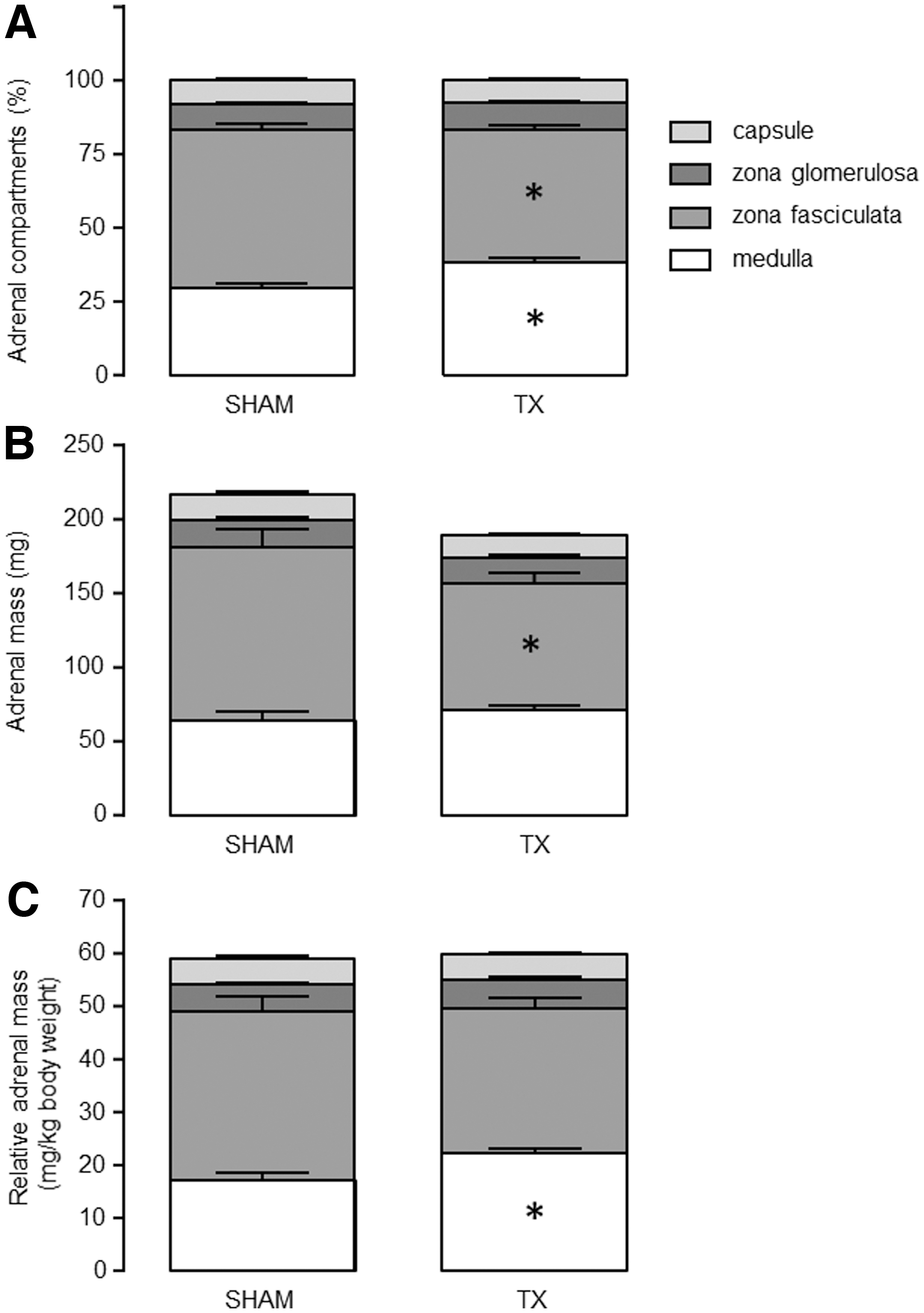
Mean (± SEM) adrenal compartments, expressed as (
Hypothyroidism in utero reduced mRNA abundance of steroidogenic and growth factor genes in the adrenal cortex
In the TX relative to the sham control fetuses, mRNA abundance in the adrenal cortex was reduced for StAR, steroidogenic enzymes CYP11A1, CYP17, 3βHSD, CYP21, and CYP11B1 (p < 0.05; Figs. 3A–F), and for IGFI and IGF-1R (p < 0.05; Fig. 3G, H). Overall, log10 plasma cortisol concentration correlated with the mRNA abundance for StAR and all of the steroidogenic enzymes (R = 0.52–0.58, p < 0.005, N = 26), except CYP21 and CYP11B1. A significant relationship was also observed between log10 plasma cortisol and adrenocortical IGF-1R mRNA abundance (R = 0.44, p < 0.05, N = 26). Fetal hypothyroidism had no effect on MC2R, IGFII, or IGF-2R mRNA abundance in the adrenal cortex, or PNMT, IGFI, IGFII, or IGF receptor mRNA in the adrenal medulla (Table 2).
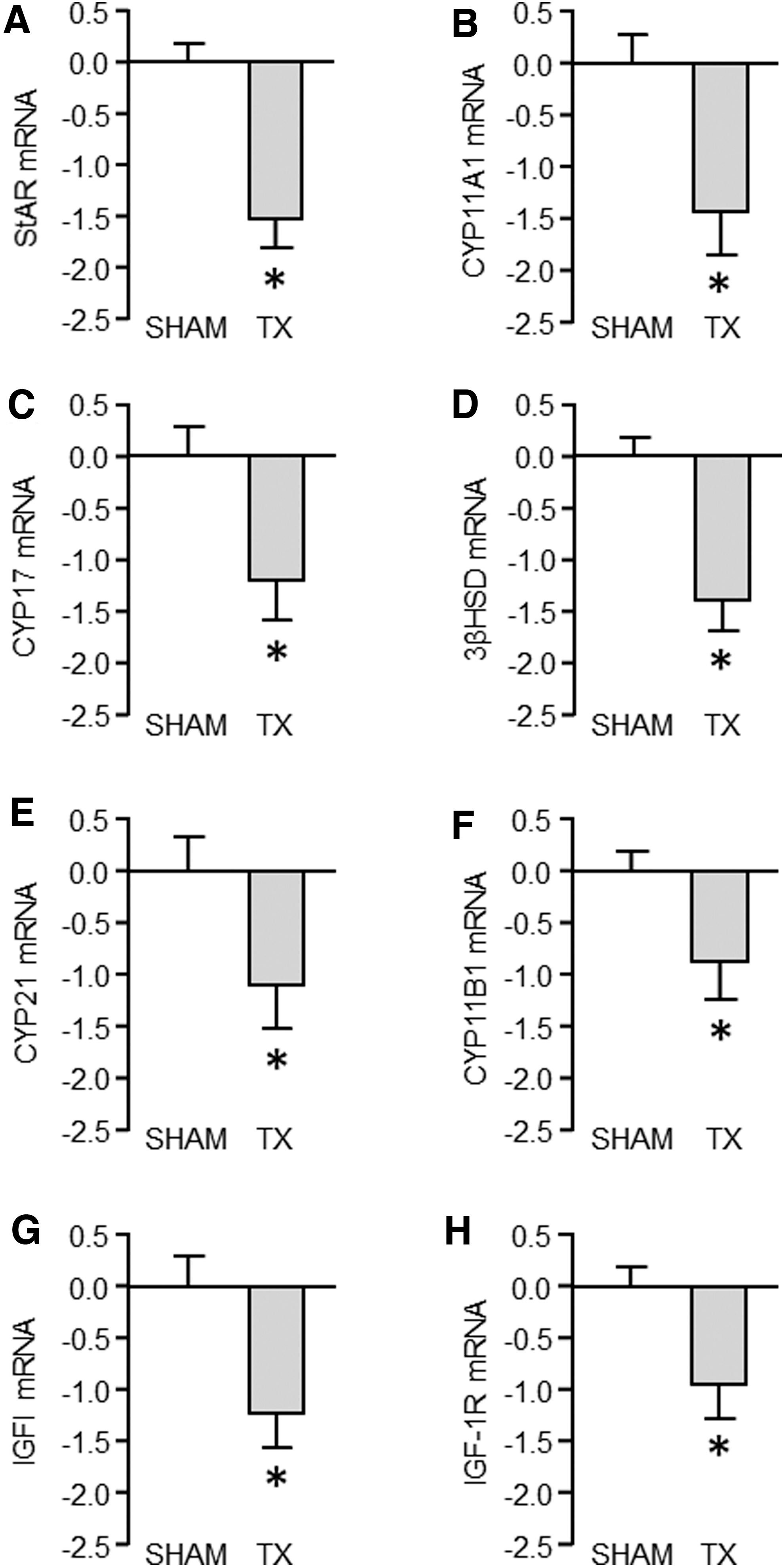
Mean (± SEM) mRNA abundance, expressed as −ΔΔCt values, of adrenocortical StAR and steroidogenic enzymes (
Mean (± Standard Error of the Mean) mRNA Abundance, Expressed as −ΔΔCt Values, in the Adrenal Cortex and Medulla of Sham and Thyroidectomized Fetuses
No significant differences were observed between the treatment groups.
ΔΔCt, delta-delta cycle threshold; MC2R; adrenocorticotrophic hormone receptor, IGFI, insulin-like growth factor I; IGFII, insulin-like growth factor II; IGF-1R, IGF type 1 receptor; IGF-2R, IGF type 2 receptor; PNMT, phenylethanolamine-N-methyltransferase.
Discussion
Thyroid hormone deficiency in the ovine fetus suppressed the activity of the HPA axis near term with actions on the structure and secretory capacity of both the anterior pituitary and adrenal glands. Hypothyroidism in utero caused reductions in circulating ACTH concentration, the size of the zona fasciculata in the adrenal cortex, mRNA levels of steroidogenic enzymes, and plasma cortisol concentration. This study has, therefore, elucidated a potential mechanism by which thyroid hormone deficiency before birth affects the timing of fetal maturation and parturition.
Contrary to the study hypothesis, the reduction in plasma ACTH concentration observed in the hypothyroid fetuses was not due to a deficit in corticotrophs in the anterior pituitary gland. Indeed, although the corticotroph population formed a smaller percentage of the anterior pituitary gland, both absolute and relative corticotroph masses were increased in the thyroid-deficient fetuses near term. Using the estimates based on the fractional volume of total tissue mass, however, it was not possible to determine whether the increase in corticotroph and anterior pituitary mass was due to hyperplasia and/or hypertrophy of pituitary cell types. Overgrowth of the pituitary gland in the hypothyroid sheep fetus was likely to be due to expansion of the thyrotroph and corticotroph populations. Previous studies in hypothyroid adult rats have reported increased numbers of thyrotrophs in the anterior pituitary gland, in response to the lack of negative feedback from thyroid hormones, and a more moderate increase in corticotroph cell number (19). The low circulating concentration of ACTH in the thyroid-deficient sheep fetus may be due, therefore, to impaired corticotroph function. Analysis of the ultrastructure of pituitary corticotrophs in hypothyroid adult rats has shown abnormalities in the formation of endocrine vesicles, which may impair the capacity for ACTH secretion (20). Little is known, however, about the control of corticotroph number and function by thyroid hormones before birth.
Hypothyroidism may delay structural and functional maturation of pituitary corticotroph types, with consequences for the activity of the fetal HPA axis. This may occur via direct actions on the anterior pituitary gland and/or via indirect actions on the neural control of hypothalamic development and neuroendocrine hypothalamic control of pituitary corticotroph function. In the sheep fetus, sub-populations of corticotrophs have been previously described, based on morphology or the expression of genes that are important for ACTH synthesis, which change in relative proportions as the pituitary gland matures toward term and in response to glucocorticoid treatment (3,4,21,22). In this study, however, it was not possible to identify morphologically distinct corticotroph subtypes in the histological images. Moreover, the morphology of corticotrophs in the fetal pituitary gland does not appear to relate to functional characteristics identified in vitro, such as the amount of ACTH stored in CRH-responsive corticotrophs (21). Further studies are required to assess the structural and functional properties of the corticotrophs present in the hypothyroid fetus, including cell ultrastructure and responsiveness to CRH and AVP.
It will also be important to determine the effects of thyroid hormone deficiency in utero on the development of the hypothalamus and its control of corticotroph structure and function. Surgical disconnection of the pituitary gland from the hypothalamus in the ovine fetus increases both thyrotroph and corticotroph numbers and delays the maturation of corticotroph subtypes, adrenal growth, and expression of steroidogenic enzymes usually seen near term (23 –26). Hypothyroidism is likely to influence neural control of hypothalamic function in the fetus, especially over the latter stages of gestation when the fetal HPA axis is activated. Thyroid hormones are well known to play an important role in the development of the central nervous system, including the hippocampus and hypothalamus (27,28), although little is known about the effects of thyroid hormone deficiency on the maturation of the neural networks that regulate the fetal HPA axis in late gestation. In neonatal rats, hypothyroidism blunts ACTH and corticosterone responses to stress and suppresses the developmental increments in plasma ACTH and corticosterone concentrations that usually occur over the early postnatal period in rodents (15, 29). The changes in neonatal pituitary–adrenal function induced by thyroid hormone deficiency were associated with lower CRH mRNA abundance and numbers of CRH-positive neurones in the paraventricular nucleus of the hypothalamus (15). Previous studies in the thyroidectomized sheep fetus have shown that neurotransmitter contents of noradrenaline and serotonin are altered in the hypothalamus and other brain regions, in a manner dependent on the timing of thyroid hormone deficiency (30,31). The consequences of hypothyroidism for the development and activity of hypothalamic CRH and AVP neurone networks toward term, however, and the extent to which changes in these pathways are responsible for suppression of ACTH and glucocorticoid production in the current study remain to be established.
The effects of thyroid hormone deficiency before birth on the developing adrenal gland appear to be mediated, at least in part, by suppression of plasma ACTH. Toward term, the rising plasma concentration of ACTH promotes the growth and secretory function of the fetal adrenal gland (32). In this study, the mRNA abundance of the ACTH receptor was unchanged by hypothyroidism in utero, but the expression of the enzymes responsible for the conversion of cholesterol to glucocorticoids was reduced. Thyroid hormone deficiency also decreased the size of the zona fasciculata in association with lower expression of IGFI and the IGF type 1 receptor in the adrenal cortex. IGFI infusion in the ovine fetus stimulates adrenal growth without affecting plasma cortisol concentration or the expression of steroidogenic or catecholamine-synthesizing enzymes (33) and IGFI may, therefore, mediate the growth-promoting effects of ACTH during late gestation. Thyroid hormones may influence adrenocortical IGF mRNA levels in a manner similar to that reported in other fetal tissues such as the liver and skeletal muscle, and indeed, interactions with the IGF system may be responsible, in part, for the reduction in fetal body weight seen in response to hypothyroidism in this and previous studies (8,34,35).
Although thyroid hormone deficiency impaired the growth of the zona fasciculata in the adrenal cortex, the relative mass of adrenal medulla was increased in the hypothyroid fetuses. Overgrowth of the adrenal medulla occurred without any changes in the expression of the IGFs or their receptors, or in the mRNA level of PNMT, the enzyme that converts noradrenaline to adrenaline. While adrenomedullary and circulating concentrations of the catecholamines were not measured in the current study, it has been previously shown that thyroid hormone deficiency in fetal sheep impairs the plasma catecholamine response to hypoxemia (30). Thyroid hormone deficiency may influence the structural and functional innervation of the adrenal medulla that occurs during late gestation, with consequences for adrenomedullary growth (36,37). It is also possible that the lower levels of glucocorticoids synthesized within the adrenal cortex of the hypothyroid fetus may impact growth of the adrenal medulla. Indeed, glucocorticoids have been shown to suppress proliferation of chromaffin cells in fetal and neonatal rats both in vivo and in vitro (38,39). Overgrowth of the adrenal medulla may, therefore, be a consequence of reduced levels of cortisol perfusing the adrenal medulla from the cortex.
The effects of thyroid hormone deficiency in utero on the developing HPA axis may be direct and/or secondary to other endocrine changes. Hypothyroidism in the sheep fetus is associated with increased circulating concentrations of insulin and leptin, which originate from changes in the structure and function of the fetal pancreas and adipose tissue, respectively (9,10). In fetal sheep, intravenous infusion of leptin to supraphysiological concentration has been shown to prevent the normal increments in plasma ACTH and cortisol seen toward term (40), and to suppress adrenal responsiveness to ACTH challenge and decrease the adrenal mRNA and protein content of the ACTH receptor, StAR and CYP21 in mildly hypoxic fetuses (41). Further, intracerebroventricular infusion of leptin suppresses the amplitude of ACTH and cortisol pulses in the fetal circulation without any change in responsiveness to CRH and AVP administration (42). This suggests that leptin may inhibit HPA activity in utero via central mechanisms, although the extent to which increased systemic levels contribute to the consequences of thyroid hormone deficiency in the current study remains to be determined. A moderate increase in circulating leptin has no effect on basal plasma cortisol concentration in the thyroid-intact sheep fetus (43).
The findings of this study contribute to the understanding of the endocrine control of fetal maturation and parturition. Although not assessed in the current study, prolonged gestational length and delayed fetal maturation have been reported in previous research using the same sheep model of fetal hypothyroidism (7,8). Impaired development of the anterior pituitary and adrenal glands, and suppression of steroidogenic enzymes, observed in the hypothyroid fetus will have consequences for the initiation of labor and delivery. While the structure of the adrenal gland differs between ovine and human species, adrenal hormones induced by ACTH are key regulators in the timing of parturition in both species (2,5). Indeed, gestation is prolonged in both human and other animal models of CH (7,11,12,14). Impaired adrenal steroidogenesis will also impact the development of fetal organs in preparation for birth since many of the maturational processes that take place over the perinatal period are known to be glucocorticoid dependent (6). Dysmaturity observed in hypothyroid fetuses and neonates, therefore, may be due to suppression of the HPA axis and a delay in the prepartum cortisol surge (8). Through interactions with the developing HPA axis, thyroid hormones produced by the fetus appear to have a role in the coordinated delivery and survival of the neonate.
Footnotes
Acknowledgments
The authors would like to thank all the technical staff of the Department of Physiology, Development and Neuroscience, University of Cambridge, Department of Biological and Medical Sciences, Oxford Brookes University and School of Animal Biology, University of Western Australia who assisted in this study.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The study was part-funded by the Biotechnology and Biological Sciences Research Council (BB/P019048/1 and BB/H01697X/1), and Isabella Inzani was supported by a Vacation Studentship from the Society for Reproduction and Fertility.
Supplementary Material
Supplementary Table S1