
Abstract
Background:
Thyroid hormone (TH) deficiency has been associated with increased cholesterol gallstone prevalence. Hypothyroidism impacts hepatic lipid homeostasis, biliary secretion, gallbladder motility, and gallstone (LITH) gene expression, all potential factors contributing to cholesterol gallstone disease (CGD). However, how TH deficiency may lead to gallstone formation is still poorly understood. Therefore, we performed molecular studies in a CGD mouse model under lithogenic conditions and modulation of TH status.
Methods:
Male, three-month-old C57BL/6 mice were randomly divided into a control (euthyroid) group, a hypothyroid (hypo) group, a gallstone (litho) group, and a gallstone+hypothyroid (litho+hypo) group and were treated for 2, 4, and 6 weeks (n = 8/treatment period). Gallstone prevalence, biliary composition and cholesterol crystals, hepatic expression of genes participating in cholesterol, bile acid (BA), and phosphatidylcholine synthesis (Hmgcr, Cyp7a1, Pcyt1a), and canalicular transport (Abcg5, Bsep, Abcb4) were investigated.
Results:
Increased cholesterol gallstone prevalence was observed in hypothyroid mice under lithogenic diet after 4 and 6 weeks of treatment (4 weeks: 25% vs. 0%; 6 weeks: 75% vs. 37.5%). Interestingly, neither the composition of the three main biliary components, cholesterol, BAs, and phosphatidylcholine, nor the hepatic expression of genes involved in synthesis and transport could explain the differences in cholesterol gallstone formation in the mice. However, TH deficiency resulted in significantly increased hydrophobicity of primary BAs in bile. Furthermore, downregulation of hepatic sulfonation enzymes Papss2 and Sult2a8 as well as diminished biliary BA sulfate concentrations in mice were observed under hypothyroid conditions all contributing to a lithogenic biliary milieu as evidenced by microscopic cholesterol crystals and macroscopic gallstone formation.
Conclusions:
We describe a novel pathogenic link between TH deficiency and CGD and suggest that the increased hydrophobic character of biliary BAs due to the diminished expression of hepatic detoxification enzymes promotes cholesterol crystal precipitation and enhances cholesterol gallstone formation in the bile of hypothyroid mice.
Introduction
Cholesterol gallstone disease (CGD) affects 10–15% of the adult population worldwide and is one of the most common gastrointestinal disorders requiring hospital admission (1,2). A possible association between thyroid dysfunction and CGD in humans has previously been reported. While two epidemiological studies from Taiwan and Italy did not find such an association (3,4), other studies suggested a link between thyroid dysfunction and CGD, in particular for overt or subclinical hypothyroidism (5 –10). Moreover, the population-based Study of Health in Pomerania (SHIP) revealed an independent association between hypothyroidism and CGD in men in contrast to a female predominance for CGD in euthyroidism (6). The pathogenic mechanism of CGD is multifactorial, and thyroid hormone (TH) may modulate potential causal factors for CGD. Hypothyroidism is associated with hypercholesterolemia, high low-density lipoprotein, and low high-density lipoprotein serum levels (11,12). Inconclusive findings were reported in hypothyroid mice and rats, which showed both increased and decreased hepatic cholesterol synthesis due to upregulation or downregulation of Hmgcr (13,14). Furthermore, decreased bile acid (BA) synthesis by downregulated hepatic Cyp7a1 was observed in hypothyroid rodents (11,15). Beside the impact on lipid homeostasis, hypothyroidism was shown to diminish the sphincter of Oddi motility, which could lead to an enhanced storage time of gallbladder bile, thereby promoting biliary supersaturation as well as precipitation of cholesterol crystals (10). Furthermore, TH alters gallstone (LITH) gene expression, for example, Cyp7a1, Abcg5/8, Bsep, Abcb4, or Sult2a1 (12,15 –19). However, most studies addressed the impact of TH as such and not the lack of TH on LITH gene expression.
To more comprehensively address TH action in CGD, we used a mouse model under CGD conditions in which we modulated systemic TH status. In this model, we show that hypothyroidism diminishes biliary BA sulfonation by downregulation of hepatic detoxification phase II enzymes Sult2a8 and Papss2 leading to an increased hydrophobicity of primary BAs, representing a novel and hitherto undescribed pathogenic link between TH deficiency and cholesterol gallstone formation.
Materials and Methods
Animals and treatment
Three-month-old, male, wild-type C57BL/6JRj mice (Janvier Labs, France) were housed under temperature (23°C ± 1°C) and light-controlled (12:12 hour light–dark cycle) conditions. After acclimatization for one week on standard laboratory chow, all mice were randomly assigned into four groups.
Control group (eu): control diet (MD.1572-97350; ENVIGO, United Kingdom) and regular drinking water supplemented with 0.3% saccharine (Sacc; Sigma–Aldrich) (control drinking water); hypothyroid group (hypo): low-iodine diet (MD.1571-95007; ENVIGO) and drinking water supplemented with 0.04% 2-mercapto-1-methylimidazole (MMI), 0.5% sodium perchlorate (ClO4 −), and 0.3% Sacc; gallstone group (litho): lithogenic diet (MD.1574; ENVIGO) containing 15.5% fat, 1.25% cholesterol, and 0.5% cholic acid (CA) and control drinking water; gallstone and hypothyroid group (litho+hypo): low-iodine lithogenic diet (MD.1573; ENVIGO) containing 15.5% fat, 1.25% cholesterol, and 0.5% CA and drinking water supplemented with 0.04% MMI, 0.05% ClO4 −, and 0.3% Sacc. Mice were treated for 2, 4, and 6 weeks (n = 8). Food and water were provided ad libitum.
All animal experiments were performed in accordance with the German regulations for Laboratory Animal Science (GVSOLAS) and the European Health Law of the Federation of Laboratory Animal Science Associations (FELASA). The protocols for animal studies were approved by the Landesamt für Natur, Umwelt und Verbraucherschutz Nordrhein-Westfalen (LANUVNRW), Germany.
Collection of serum, bile fluid, gallbladder, and liver tissue
At the end of experiment and after 12 hours of fasting, mice were euthanized as described previously (20). Blood samples were harvested by right ventricular heart puncture from deeply anesthetized animals, centrifuged at maximum speed for 20 minutes at 4°C, and serum samples were stored at −80°C. Determination of cholesterol gallstones was performed by examination of intact gallbladders under light (n = 8). Biliary cholesterol monohydrate (ChM) crystals were investigated by light polarization microscopy using Olympus BX51 upright microscope (Olympus, Germany) (n = 4–8). Bile fluid was collected by puncture with an insulin syringe (BD Micro-Fine, 29G) and stored at −80°C. Livers, gallbladders, and pituitaries were isolated from heparinized saline-perfused mice and processed as followed: liver tissue and pituitary were snap frozen in liquid nitrogen and stored at −80°C or liver tissue was embedded in Tissue-Tek O.C.T. compound (Plano GmbH, Germany), snap frozen in liquid nitrogen, and stored at −80°C. For histological analysis, liver and gallbladder tissues were stored in 4% paraformaldehyde in 1 × phosphate-buffered saline (PBS) at room temperature.
Serum measurements
Total thyroxine (TT4) serum concentration (n = 7–8) was measured using an ELISA kit according to the manufacturer's instructions (EIA 1781; DRG Instruments GmbH, Germany). Data below the detection limit of 0.5 μg/dL were derived from the standard curve.
Serum enzyme activities (n = 4) of alanine transaminase (ALT), aspartate transaminase (AST), and alkaline phosphatase (AP) were analyzed by LABOKLIN GmbH & Co. KG (Bad Kissingen, Germany).
Bile measurements
Biliary BA concentrations (n = 4–8) were determined by negative electrospray ionization liquid chromatography–tandem mass spectrometry (LC-MS/MS) in multiple-reaction-monitoring mode on an Agilent 6460 triple quadrupole mass spectrometer (Agilent, Germany) coupled to an Agilent 1290 HPLC system (21). Briefly, 5 μL of diluted bile (1:100) were spiked with 20 μL of internal standard solution in methanol:water (1:1 v/v). Protein precipitation was performed with 30 μL of methanol, followed by centrifugation. The supernatant was used for LC-MS/MS analysis (Supplementary Tables S1–S3).
Cholesterol concentrations (n = 4–8) in bile were determined by gas chromatography–mass spectrometry using 10 μL of diluted bile (1:100) (22).
Biliary phosphatidylcholine concentrations (n = 3–8) were measured using a colorimetric coupled enzyme assay (MAK049; Sigma–Aldrich) according to the manufacturer's instructions.
Histological analysis
Liver and gallbladder samples were fixed in 4% paraformaldehyde in 1 × PBS, embedded in paraffin, and 4 μm thick sections were used for hematoxylin and eosin (HE) staining. Gallbladder wall thickness was quantified using the ImageJ software. For each gallbladder (n = 3–4), wall thickness is the mean value of six determinations in the fundus area (10 × magnification).
To determine neutral lipids, Oil Red O staining of frozen sections of Tissue-Tek-embedded liver samples (5 μm) was performed and quantified using Adobe Photoshop CC2017 as follows: lipid intensities were determined by isolation of the neutral lipid channel from the photomicrographs, followed by evaluation of the mean density values using the histogram feature of five field of views per image (60 × magnification, n = 4). HE staining was digitalized by Aperio Scanner (Leica, Germany), and Oil Red O staining was viewed on the Olympus BX51 upright microscope (Olympus).
Quantitative real-time polymerase chain reaction
Total RNA was isolated using RNeasy Kit (Qiagen, Germany) and reverse transcribed into cDNA (Life Technologies, Germany). Quantitative real-time polymerase chain reaction (qRT-PCR) was performed as described previously (20). Sequences of oligonucleotides designed using Primer-BLAST (NCBI) and synthesized by Eurofins (Eurofins MWG, Germany) are provided in Supplementary Table S4. Relative expression of genes was normalized to euthyroid (eu) mice (n = 7–8).
Cholesterol saturation index calculation
The cholesterol saturation index (CSI) of each sample was calculated in bile using Carey's critical tables for cholesterol solubility limits based on %moles of cholesterol, BA, and phospholipids (23).
Statistics
All data are shown as mean ± standard error of the mean. Statistical analysis was performed using GraphPad Prism 7 software. One-way ANOVA followed by Bonferroni post hoc analysis was used. Expression data of qRT-PCR were analyzed using anti-logarithmic data. Values of *p < 0.05, **p < 0.01, ***p < 0.001, and # p < 0.0001 were considered statistically significant.
Results
Hepatobiliary histology and function under hypothyroid and lithogenic conditions
Successful induction of hypothyroidism was determined by analysis of TT4 serum concentrations, and mRNA expression analysis of pituitary Tshb showing significantly reduced TT4 serum levels and increased Tshb expression in hypothyroid mice (hypo, litho+hypo) compared with eu mice (Fig. 1A, B). Hepatic thyroid state was assessed by mRNA expression analysis of TH-responsive genes Dio1 and Tbg, respectively. Hypothyroidism (hypo, litho+hypo) diminished Dio1 expression and increased Tbg expression (Fig. 1C, D). In addition, lithogenic diet (litho) reduced hepatic Dio1 expression compared with eu mice, but to a lesser extent compared with the hypo and litho+hypo groups (Fig. 1C).
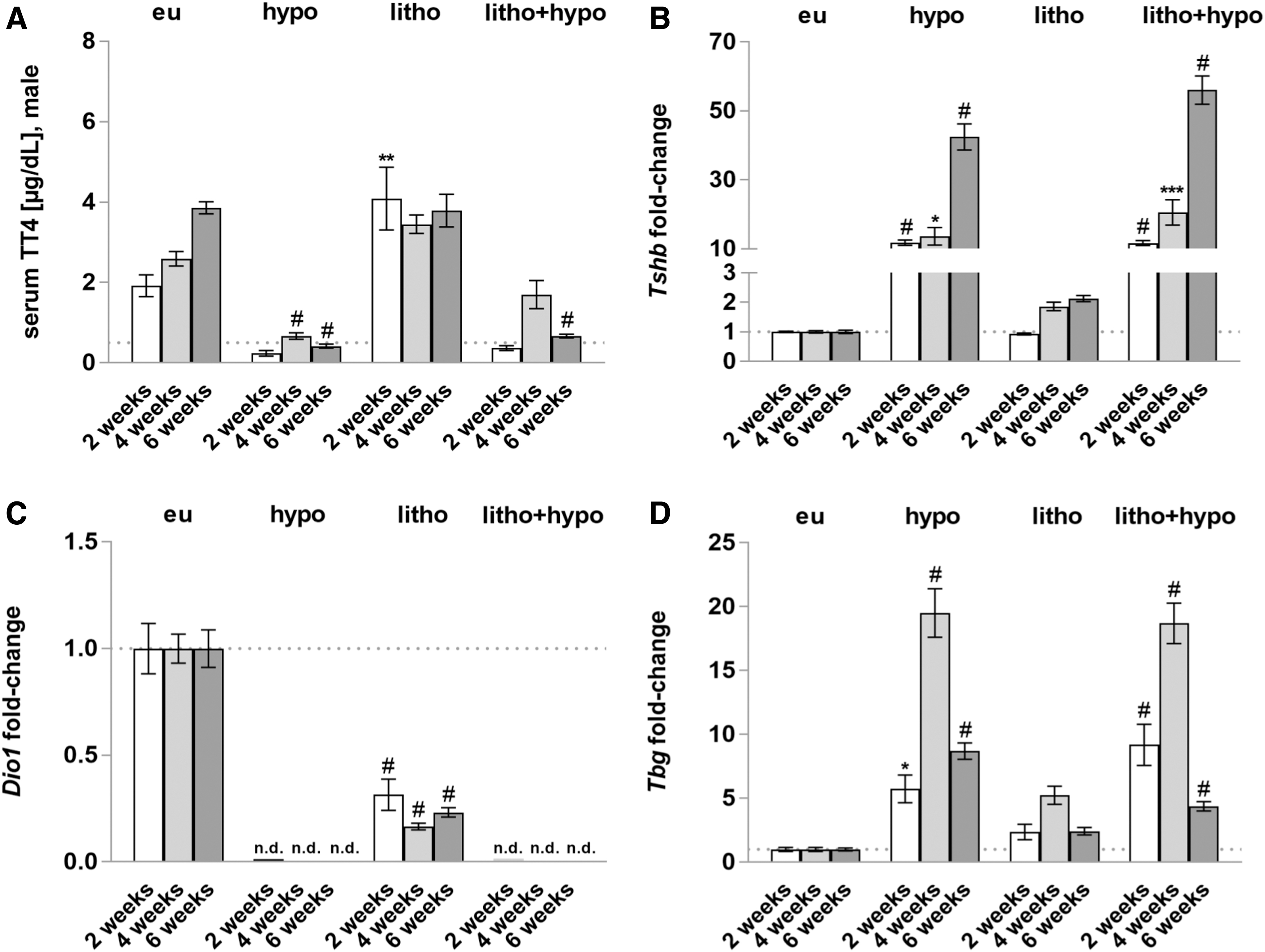
Successful induction of systemic and hepatic hypothyroid state. (
Lithogenic diet increased hepatic lipid content demonstrated by increased hepatocyte lipid droplets formation (Fig. 2A–D). To a minor degree, hepatic lipid droplets were also observed under hypo condition. In addition, increased gallbladder wall thickness under litho and litho+hypo conditions was observed (Fig. 2E, F). Serum enzyme activities of AST and ALT were elevated under hypo, litho+hypo, and litho conditions compared with the control group with the highest transaminase activities found in the litho group (Supplementary Fig. S1A, B). Serum enzyme activity of AP was elevated under litho condition and diminished under hypo and litho+hypo conditions compared with the control group (Supplementary Fig. S1C).

Increased hepatic lipid accumulation and gallbladder wall thickness under lithogenic diet. (
Hypothyroidism increases cholesterol gallstone formation and ChM crystal prevalence in male mice
After 2 weeks, 12.5% of mice under litho and litho+hypo conditions had cholesterol gallstones (Fig. 3A). In contrast, after 4 and 6 weeks, higher cholesterol gallstone prevalence was observed in mice under litho+hypo condition compared with litho condition (4 weeks: 25% vs. 0%; 6 weeks: 75% vs. 37.5%) (Fig. 3A). No cholesterol gallstone formation was observed for eu and hypo mice maintained under normal diet (Fig. 3B). Similar results were obtained when biliary ChM crystal formation was quantified by light polarization microscopy (Fig. 3C, D).
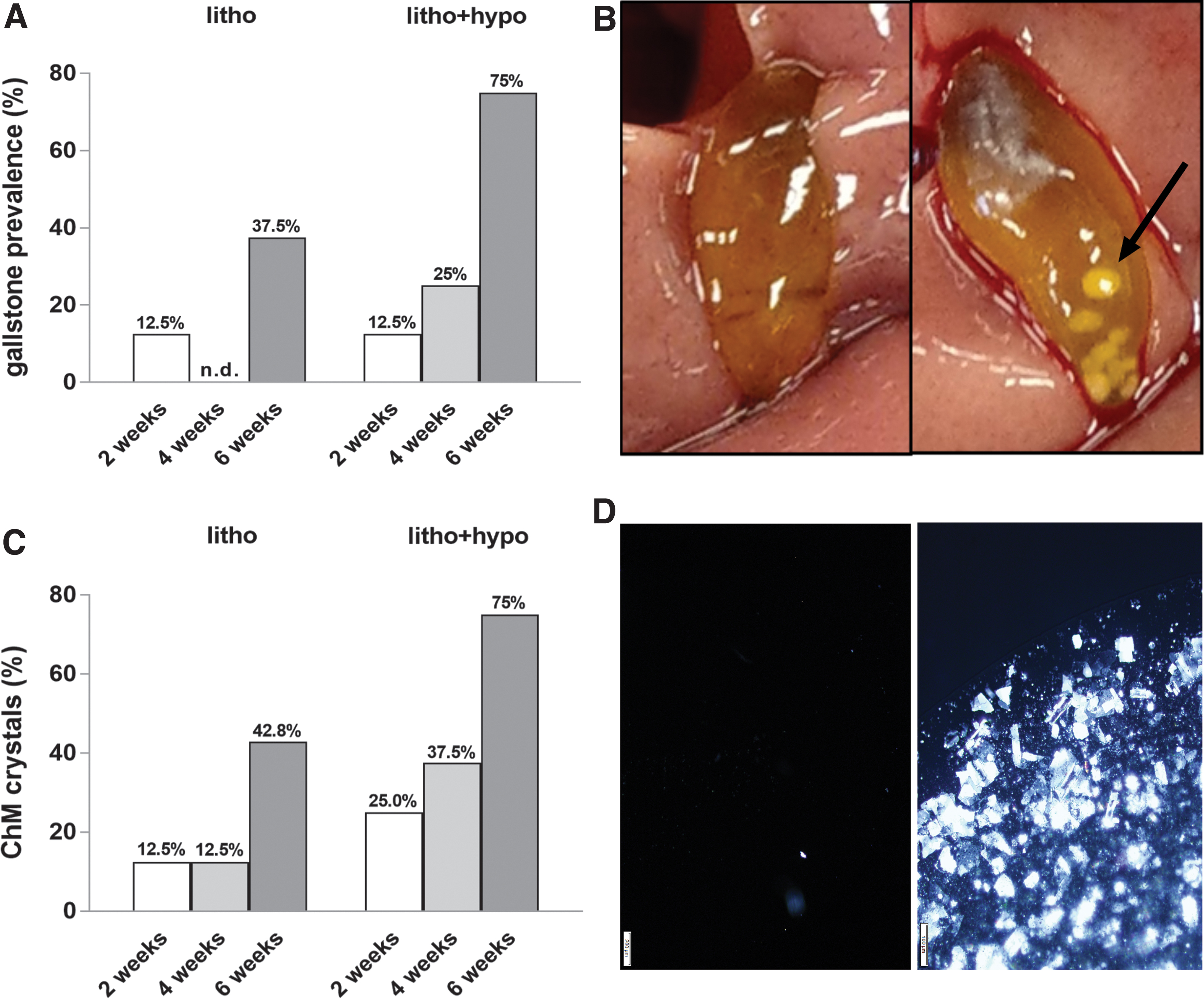
Hypothyroidism increases cholesterol gallstone formation and biliary ChM crystals. (
Biliary balance of cholesterol, BAs, and phosphatidylcholine is not responsible for enhanced cholesterol gallstone formation in hypothyroidism
Because biliary imbalance of cholesterol, BA, and phospholipids and supersaturation of bile with cholesterol are thought to be the main causes for cholesterol gallstone formation (24), we investigated the hepatic gene expression of synthesis enzymes and canalicular transporters (Supplementary Fig. 2) and the biliary concentrations of these components (Fig. 4). Hmgcr and Cyp7a1 were downregulated in litho, hypo, and litho+hypo conditions (Supplementary Fig. S2A, C). The expression of the rate-limiting enzyme in phosphatidylcholine synthesis, Pcyt1a, was unchanged in all treatment groups (Supplementary Fig. S2E). Canalicular BA transporter Bsep and phospholipid transporter Abcb4 were downregulated in hypo condition, but not litho condition (Supplementary Fig. S2D, F). In contrast, expression of the canalicular cholesterol transporter Abcg5 was increased in litho condition after 4 and 6 weeks of treatment, and this was even more pronounced in the litho+hypo group (Supplementary Fig. S2B).

Biliary balance of cholesterol, BAs, and phosphatidylcholine under hypothyroidism. (
Lithogenic diet increased biliary total cholesterol, total BA, and phosphatidylcholine concentrations as well as the CSI under litho and litho+hypo conditions compared with eu mice (Fig. 4A–D). In contrast, hypo mice showed a diminished proportion of total cholesterol and phosphatidylcholine and an increased proportion of total BA compared with eu mice (Fig. 4E).
Increased hydrophobicity of primary BAs results from downregulated phase II detoxification enzymes
Beside the biliary composition, BA hydrophobicity may contribute to cholesterol gallstone formation (24). We identified 20 BAs subdivided into 12 primary and 8 secondary BA in bile of the mice (Supplementary Tables S1–S3). The ratio of hydrophobic versus hydrophilic BA revealed an increased amount of hydrophobic primary BA under hypo and litho+hypo conditions compared with eu condition (Supplementary Fig. S3). To increase BA solubility, primary BAs are conjugated by hepatic detoxification phase II processes. Further analysis revealed diminished BA sulfate concentrations in hypo, litho+hypo, and litho conditions (Fig. 5A). Moreover, after 4 and 6 weeks, the observed decrease in BA sulfate concentrations was even more pronounced in litho+hypo condition compared with litho condition (Fig. 5A). In addition, concentration of glycine- and taurine-conjugated BA was reduced after 6 weeks of treatment under hypo and litho+hypo conditions (Fig. 5B). The expression of Papss2 (sulfonation), Sult2a8 (sulfonation), Baat (amidation), and Ugt2b1 (glucuronidation) was diminished either in hypo or in litho condition (Fig. 5C–F). Most prominent changes were found for Papss2 and Sult2a8 with significant reduction in hypo, litho+hypo, and litho conditions (Fig. 5C, D).

Downregulated gene expression of hepatic detoxification phase II enzymes under hypothyroidism. (
Discussion
To understand how TH deficiency may impact gallstone formation, we conducted mouse studies and show that hypothyroidism increases cholesterol gallstone formation in male C57BL/6 mice due to elevated hydrophobicity of primary BAs. Importantly, this lithogenic change in bile character may arise from downregulation of hepatic detoxification phase II enzymes reducing BA sulfonation and concentrations of glycine- and taurine-conjugated BAs in hypothyroid mice.
Under experimental conditions, an increased cholesterol gallstone formation in litho+hypo condition compared with litho condition after 4 and 6 weeks, but not after 2 weeks, of treatment was observed (Fig. 3A). In contrast to previous studies, the observed gallstone prevalence under lithogenic diet was lower, which could be explained by differences in diet composition and/or mice strains used (25,26). The lack of representative cholesterol gallstones after 2 weeks is consistent with previous study (9) and could be explained by the short treatment period, which may not be sufficient time to lead to CGD.
Interestingly, mice in litho condition showed reduced hepatic Dio1 expression (Fig. 1C) suggesting a local hepatic hypothyroid state in contrast to a systemic eu condition (Fig. 1A). The impact of cholesterol-rich diet on systemic TH levels and hepatic deiodinases has already been described in several studies (27 –29). However, Dio1 downregulation was more pronounced under litho+hypo condition than under litho condition (Fig. 1C).
The solubility of biliary cholesterol is maintained by mixed micelles containing bile salts and phospholipids (23). A biliary imbalance and supersaturation with cholesterol are thought to be the main causes for cholesterol gallstone formation (30). Therefore, we investigated biliary concentrations of cholesterol, BA, and phosphatidylcholine, the main phospholipid, as well as hepatic expression of genes participating in cholesterol, BA, and phosphatidylcholine synthesis (Hmgcr, Cyp7a1, Pcyt1a) and canalicular transport (Abcg5, Bsep, Abcb4). Downregulation of Hmgcr and Cyp7a1 was observed in hypothyroidism (Supplementary Fig. S2A, C) and is consistent with other studies (16,31 –33). In our mouse study, expression of Pcyt1a was not altered by hypothyroidism (Supplementary Fig. S2E). Canalicular cholesterol transport is maintained by the ABCG5/8 heterodimer, which is regulated by triiodothyronine (17). TH deprivation had no effect on Abcg5 or Abcg8 expression in mice (9), and this was also confirmed in our study (Supplementary Fig. S2B). However, we observed a significant increase in Abcg5 expression under litho condition, which was more pronounced in the litho+hypo group. Furthermore, we found downregulation of hepatic bile salt and phosphatidylcholine canalicular transporters Bsep and Abcb4 under hypothyroid conditions, while the lithogenic diet had no impact (Supplementary Fig. S2D, F).
Considering these findings of the altered hepatic expression of canalicular transporter genes, we expected changes in biliary composition. Due to the more prominent upregulation of Abcg5 and downregulation of Bsep and Abcb4 under litho+hypo condition compared with litho condition (Supplementary Fig. S2B, D, F), we expected higher biliary cholesterol and diminished biliary BA and phosphatidylcholine concentrations in the litho+hypo group. However, no significant differences of cholesterol, BA, and phosphatidylcholine concentrations between both treatment groups were present (Fig. 4A–D). In addition, hypothyroidism did not impact the biliary ratios of cholesterol, BA, and phosphatidylcholine under lithogenic conditions (Fig. 4E). In conclusion, increased cholesterol gallstone prevalence under TH deprivation could not be explained by changed biliary composition in our mouse study model.
CGD is also associated with elevated hydrophobicity of BA (34,35). Biliary BAs originate from newly synthesized primary BA and secondary BA returned to the liver via the enterohepatic circulation (Supplementary Fig. S3A). The biliary LC-MS/MS analysis (Supplementary Tables S1–S3) revealed increased hydrophobicity of primary but not secondary BA under hypothyroidism (Supplementary Fig. S3C, D). It has been demonstrated that CA feeding alters the qualitative pattern of BA in mice to 83% of taurocholic acid in total bile pool (36), which was TH independent in our study (Supplementary Fig. S3B). However, hypothyroidism shifted the BA pool toward increased hydrophobicity (Supplementary Fig. S3B). Furthermore, CA feeding has been shown to increase cholesterol intestinal absorption and biliary secretion (36), a factor possibly affecting cholesterol gallstone formation. Thus, further studies on intestinal mechanisms in relation to the TH status should be performed.
The question arises if the chemical compound MMI impacts BA hydrophobicity. However, MMI treatment increases the bile flow and biliary output of BAs in rats (37), whereby it is known that mainly hydrophilic BAs reveal choleretic properties leading to an increased bile flow (38). Moreover, Wang et al. revealed increased biliary ChM formation in subtotal thyroidectomized C57BL/6 mice under lithogenic diet compared with eu mice (9). Therefore, we assume that MMI does not contribute to the increased hydrophobicity of primary BA.
The conjugation of BA with sulfate, amino acids, as well as glucuronate increases their hydrophilicity, decreases their intestinal absorption, and enhances their fecal and urinary excretion (39). To investigate if hypothyroidism impacts hepatic detoxification processes, we analyzed the gene expression of hepatic detoxification enzymes. The expression of Papss2 and male predominant murine sulfotransferase Sult2a8, both enzymes involved in sulfonation, was downregulated under litho, hypo, and litho+hypo conditions (Fig. 5C, D). This suggested reduced concentrations of biliary BA sulfates due to the lithogenic diet and TH status and could in fact be confirmed by biliary LC-MS/MS analysis (Fig. 5A). Moreover, the reduction in BA sulfate concentrations was more pronounced after 4 and 6 weeks under litho+hypo condition compared with litho condition (Fig. 5A). This finding strongly correlates with the increased cholesterol gallstone prevalence in hypothyroid mice. Interestingly, Sult2a8 has been identified as a BA sulfotransferase with high activity toward primary BA, but poor activity toward secondary BA (40). This is in accordance with our results showing that only primary BA hydrophobicity is affected by hypothyroidism.
Altered amidation and glucuronidation enzymes were reported in cholestatic patients (41 –44). Our results show evidence of TH impact on expression of the glucuronidation enzyme Ugt2b1 (Fig. 5F). Furthermore, we could detect a trend of downregulated Baat and Ugt2b1 in litho+hypo condition (Fig. 5E, F). While this suggested that the decrease in amidation and glucuronidation enzymes could also contribute to the increase of primary BA hydrophobicity (Fig. 6), significant changes were only found for biliary glycine- and taurine-conjugated BA concentrations after 6 weeks of treatment in hypothyroid mice (hypo, litho+hypo) (Fig. 5B). Although biliary concentrations of sulfated BA are low (45) and their physiological role in cholesterol solubilization is doubtful, together with glucuronidated and amidated BA, they could impact cholesterol gallstone formation.

Hypothyroidism decreases gene expression of hepatic detoxification phase II enzymes leading to increased hydrophobicity of primary BAs promoting gallstone formation. Color images are available online.
In conclusion, we demonstrate that severe hypothyroidism shown by profoundly reduced TT4 serum levels and highly elevated Tshb expression increased cholesterol gallstone formation due to elevated hydrophobicity of primary BA in male C57BL/6 mice. While this represents a novel mechanism of TH-induced biliary disease, we acknowledge limitations of our study. First, experiments were conducted in male mice and group sizes were limited. Thus, whether the identified changes in BA hydrophobicity under hypothyroidism also apply for gallstone formation in female mice, and whether minor changes and trends found, for example, for amidation and glucuronidation enzymes could become significant findings in larger cohorts, needs to be determined. Second, our approach of using the well-established low-iodine/MMI/perchlorate treatment regimen, which causes a profound state of hypothyroidism in mice, triggers an extreme pathophysiology. Thus, it will be highly interesting whether more subtle changes in TH serum concentrations (as seen, e.g., in subclinical hypothyroidism) could also impact cholesterol gallstone formation. While these issues will be the subject of future research, our study describes a novel pathogenic link between TH deficiency and CGD.
Manipulation of BA composition and pool size by BA sequestration have already found clinical application using ursodeoxycholic acid (UDCA) in patients with cholesterol-enriched noncalcified gallstones <20 mm in diameter (46), underlining the relevance of BA hydrophobicity in the pathogenesis of CGD. In case of hypothyroidism, the foremost prevention treatment is adequate TH replacement (47). This raises the question whether simultaneous application of local TH and UDCA in patients with CGD may improve the dissolution of cholesterol gallstones >20 mm in diameter. Further studies are needed to elucidate in how far minor alterations in systemic and hepatic TH availability will contribute to CGD in mice and humans and in how far other co-founders such as obesity inducing a hepatic hypothyroid state (48) may also exert increased CGD risk by the same mechanisms.
Footnotes
Acknowledgments
We thank Kathrin Strumann and Sabrina Kwapik (University of Duisburg-Essen, Department of Endocrinology, Diabetes and Metabolism and Clinical Chemistry–Division of Laboratory Research, Essen, Germany) for competent technical assistance. We also thank the Imaging Core Facility Essen (IMCES) and the Institute of Pathology for its support with the Olympus microscope and histological analysis.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The authors were financially supported by the DFG in the framework of the priority research program SPP1629 (ZW221/2-1) and TRR 296/1-P18. U.H. was supported by the Robert-Bosch Foundation (Stuttgart, Germany).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4