
Abstract
Background:
The development of the thyroid follicular cell has been well characterized as it progresses from the original stem cell, either embryonic or adult, through a series of transitions to form a differentiated and functional thyroid cell.
Summary:
In this review, we briefly outline what is known about this transitional process with emphasis on characterizing the thyroid progenitor stem cell by using data obtained from both in vitro and in vivo studies and both mouse and human cells. It is of particular importance to note the influence of independent factors that guide the transcriptional control of the developing thyroid cell as it is subjected to extracellular signals, often working via epigenetic changes, and initiating intrinsic transcriptional changes leading to a functional cell.
Conclusion:
Thyroid stem cells fall into the category of dispositional stem cells and are greatly influenced by their environment.
Introduction
Stem cells present a unique capacity for self-renewal and differentiation into other cell types (1). Recent studies have paved new pathways for the potential use of stem cells in regenerative medicine and for understanding the biological processes that lead these naive cells into a particular organ-specific differentiation pathway, including the thyroid gland. In theory, functional tissue replacement can now be achieved either by differentiating embryonic stem cells (ESCs) into the required tissue type in vitro or by developing “induced” pluripotent stem cells into the required tissue.
The ESCs, by definition, are pluripotent (or totipotent) cells with an intrinsic ability to generate all tissues. In contrast, adult stem cells (ASCs), which are resident in tissues, have a more limited regenerative ability although like ESCs they too are defined by a clonogenic population with the capacity of self-renewal and immortality. However, identifying resident stem cells and characterizing their role in the thyroid gland has not been as straight forward as it may have once seemed as we move past the initial stages of development (2). Indeed, when it comes to endocrine stem cells, and thyroid stem cells in particular, the situation is increasingly complex because these resident stem cells reside in an extracellular niche that regulates their behavior through epigenetic and nonepigenetic signaling networks. Further, it is not yet fully clear as to what precise spatiotemporal signals are responsible for guiding either ESCs or ASCs into a differentiated thyroid cell in vivo.
As a result of the changing cellular landscape that stem cells encounter, there appears to be a continuum of cell stages ranging from ASCs to committed progenitors, which are oligopotent cells with a restricted capability, all the way to a fully differentiated follicular cell with limited proliferative ability (Fig. 1). This concept is critical in understanding that a cell may be at a particular stage, with the active genes relating to that stage, but it may be very transient. Eventually, the cell becomes committed and well on its way to a differentiated state.
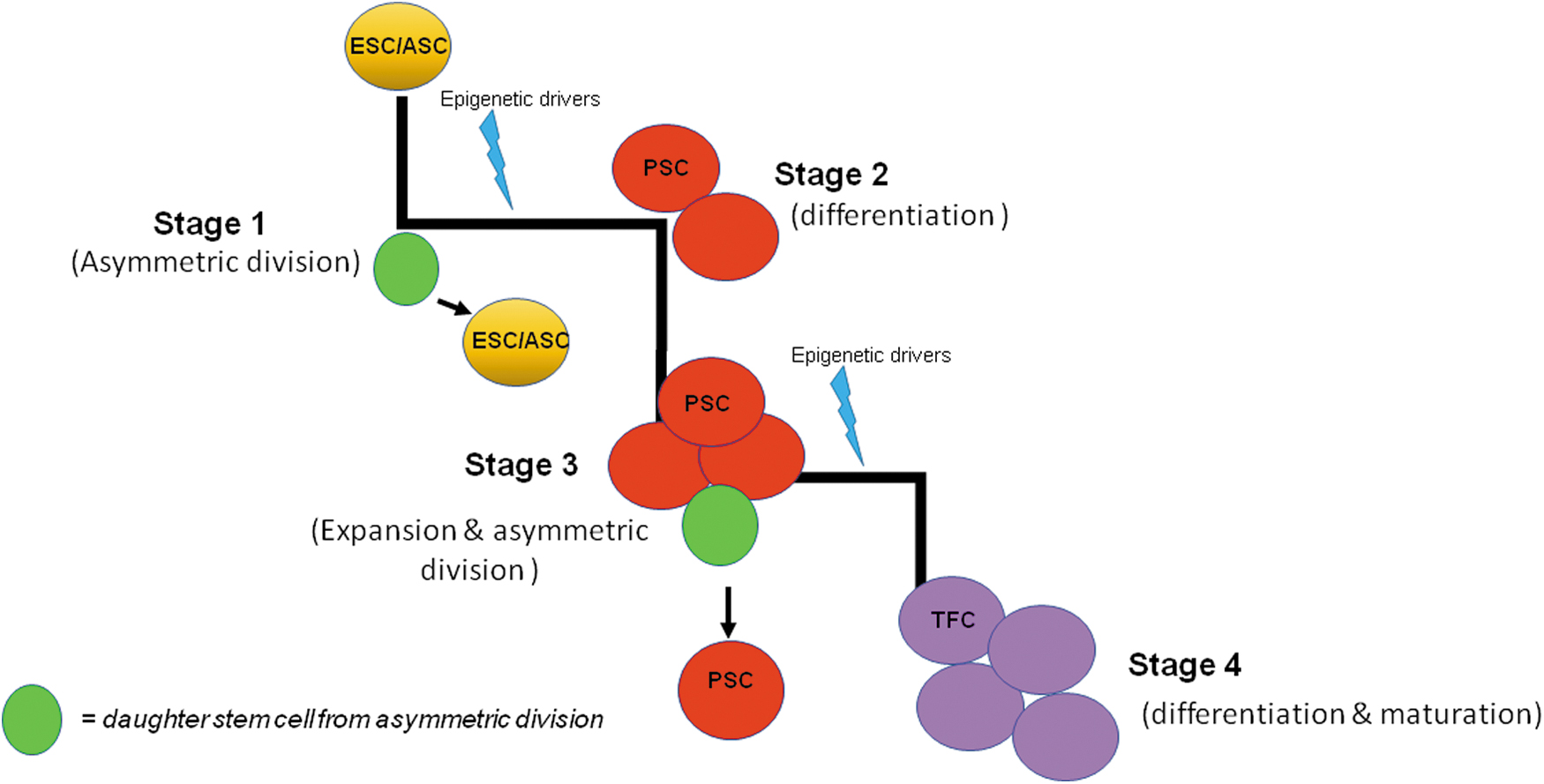
Cascade of “thyroid stem cell continuum.” This diagram illustrates our concept of the thyroid cell continuum. The cascade begins with ESCs or ASCs, which first start dividing asymmetrically to give rise to a daughter stem cell, which maintains the pool and mature stem cells that on receiving epigenetic drivers (natural or induced) propel them to form PSCs. These PSCs further expand by mitosis and also provide cells to the PSC pool by dividing asymmetrically. The PSCs, on receiving further specific epigenetic triggers, then differentiate into TFCs, which are mature differentiated cells that expand by mitosis to form functional thyroid follicles. ASCs, adult stem cells; ESCs, embryonic stem cells; PSCs, progenitors stem cells; TFCs, thyroid follicular cells.
Correctly defining the various developing cell types is a challenge because of their transitional state but one successful approach is via so-called “marker genes”—the genes that the cells express at a particular time in development. For example, in the case of developing thyroid-specific cells, we can say that the increased and combined expression of key transcription factors such as Pax8, NKX2-1 and FoxE1 are the “marker genes.” However, this requires a comprehensive profile for identification and classification. Although the cell populations always show considerable heterogeneity (3), a certain level of clarity has been possible. It remains the best way to understand these stem cell populations if a characteristic gene expression profile can be described. The ESCs have such a characteristic gene expression profile involving mostly genes that are well known to enhance immortality and maintain pluripotency (4,5). However, such a clear distinction has been hard to define for ASCs because of their evasive nature in the organ in which they reside, especially so in the thyroid, with limited ability to isolate and characterize them. Recent attempts (6 –8) have shown the existence, and possible location (9 –11), of thyroid resident ASCs in thyroid tissue (2,12) but they remain poorly characterized.
The ASCs tend to be increasingly oligopotent with restricted potential, often only related to the tissue in which they reside. Under certain conditions, such as inflammation or tissue damage to the organ (13 –15), such ASCs may become activated, due to still mostly unknown cues, and transform into progenitor stem cells (PSCs) which exhibit monopotency and are designated for a single-cell type such as a thyroid follicular cell. The fact that these ASCs have to remain quiescent in some tissues for several years until they are activated back to a proliferative and secretory role would suggest that they remain in a low metabolic state, possibly under hypoxic conditions, and tend to have genes that are upregulated to hold them in their dormant state and ready to differentiate. This situation is not so different to dormant metastatic cancer cells, which may not reveal themselves for many years and which continue to have unidentified micro-environmental cues (16,17).
This logic leads to another way of trying to understand stem cells by considering their intrinsic capacity and their relationship with the environment. Such an approach classifies stem cells into four distinct groups: (i) categorical stem cells—these are intrinsic and independent in nature such as certain cancer stem cells (CSC), (ii) dispositional stem cells—these are intrinsically driven but are highly influenced by external factors such as a stem cell niche, (iii) relational stem cells—these have acquired stemness by their interaction with external stimuli and are not intrinsically driven, and (iv) systemic stem cells—these have acquired stemness without any external stimuli, for example when cancer cells are converted to stem cells (18,19).
Thyroid progenitor cells that reside in the gland, or generated in vitro, are certainly dispositional stem cells and highly reactive to their environment, especially when subjected to injury (9,11), small molecules such as ethacridine (20), overexpression of key genes (14,21,22), or a cocktail of growth factors (23). Nevertheless, these cells are also not well defined in the literature and much confusion has ensued. In this commentary, we attempt to outline the changes that occur as a stem cell moves through its continuum from ESC/ASC to PSC, both in vitro and in vivo, to become a fully differentiated thyroid follicular cell.
ESCs Forming Thyrocytes
Transcriptomic analysis of both human and mouse ESCs combined with gene knockout and real-time polymerase chain reaction has clearly outlined the gene expression profile of ESCs and has largely contributed to the knowledge of the genes needed to maintain pluripotency. SAGE (serial analysis of gene expression) studies on human and mouse ESCs (4) has pointed out that these cell types appear to share a number of gene products but there are notable differences such as the inactive leukemia inhibitory factor and highly enhanced expression of POU5F1 and SOX2 gene products in human ESCs. Such in-depth transcriptomic examinations have contributed to understanding the molecular mechanisms of self-renewal and differentiation in ESCs and initially gave us the ability to effectively culture these cells in the laboratory. This subsequently led to the discovery of induced pluripotent stem (iPS) cells, which can be made from almost any somatic cell (24). However to retain their “stemness”—their ability to self renew and differentiate—a characteristic set of genes is continuously activated and expressed. Detailed expression analysis carried out in human ESCs has indicated a host of core genes. This grouping of genes can be divided into surface markers, identified by antibody staining, and transcription factors, allowing their detection by mRNA analysis. Not all these genes need to be active simultaneously in every ESC and different lines of such cells will express somewhat different profiles, which may also vary with time and culture conditions. Further, the intensity of the signature genes correlates with the plasticity and differentiation stages of the cells (25). Our mRNA-Seq examination of >15,000 transcripts for the human ESC line H9 showed that 13 of the most commonly expressed surface markers and 8 transcription factors were active and formed a signature of stem cell genes (Fig. 2A, illustrated in green text on the X axis).

mRNA-Seq of human ESCs. (
The ESCs require defined culture conditions in vitro to maintain their pluripotentiality. Such ESCs are highly reactive to their environment, and their transformation can be guided by external stimuli. Thus, we (20) and others (21,23) have successfully exploited the nature of ESCs (both human and mouse) to differentiate them into functional thyroid follicular cells. The fact that they can continue to exist to form functional tissue reservoirs in implanted immunocompromised sites, such as under the kidney capsule (21,23), suggests their ongoing plasticity.
ASCs in the Thyroid Gland
Unlike ESCs, ASCs are resident in a particular tissue and are known to be multipotent—they readily form the tissue of that organ—but are not pluripotent. Although they can sometimes be coaxed to form other tissues, it is important to remember that they cannot form an entire multicellular organism—a capability only retained by ESCs. As discussed earlier, such ASCs are generally difficult to identify and isolate from tissues, including the thyroid, because they reside in poorly defined “niche” areas. Studies carried out on intestinal crypt cells and hair follicle bulge cells have, however, identified and located stem cells with signature cell markers (26,27) and have been successfully traced through their developmental stages. Recent attempts (6 –8) have also shown the existence, and location (9 –11), of thyroid resident ASCs in thyroid tissue where they reside between thyroid follicles (11) and are maintained in a steady nondifferentiating state by the niche environment, which may include a variety of molecular controls and a degree of hypoxia (28). These are rare to find, because they exist in very low numbers and are not activated to divide unless there is tissue damage or disease of the organ, which then induces their transformation. In addition, to maintain their cell pool they undergo asymmetric cell division whereby they segregate cell fate determinants into only one of the daughter cells. This method of cell division also helps to orient the cell plane so that one of the daughter cells maintains contact with the niche and thus retains stem cell identity (Fig. 1). We have previously shown that thyroid CSCs undergo asymmetric cell division, contributing to cellular diversity (19). It is believed that slow cycling ASCs can remain dormant for years in some cases but in others they can be highly active. For example, studies mentioned earlier and carried out with intestinal villi crypt cells identified adult Lgr5-positive stem cells as the reservoir that builds the fast depleting intestinal villi cells and are also a source for intestinal cancer forms when they develop genetic aberrations (29). Attempts to isolate ASCs from mouse and human thyroid have included the characterization of side-population cells via flow cytometry (Fig. 3A) (6,8). Side-population cells express the ATP binding cassette–dependent transporter (ABCG2), a drug transport channel, which has been identified as a marker for certain stem cells (30) and studies have established that these side population cells have ASC characteristics (Fig. 3A) (31). Unpublished studies carried out from our laboratory examining mouse side population cells did show them to be Sca1 and Oct4 positive but negative for thyroglobulin (TG) or sodium iodide symporter (NIS) and low in NKX2-1 and Pax8. Whatever the nature of these adult thyroid resident stem cells, to become “thyroid-related,” such cells must acquire an endodermal phenotype from which the thyroid gland differentiates. Hence, we would expect such thyroidal ASCs, with reduced tissue potential, to express an array of endodermal markers as, indeed, shown in the side population cells just described (unpublished data). This can also be mimicked in vitro when human ES cells are treated with activin, a known essential driver of the endoderm. Of importance is the concept that as the cell moves into an endodermal phase it should reduce or even lose the expression of its stem cell genes, which were the drivers of pluripotency. This phenomenon is well illustrated in Figure 2B upper. Hence, while endodermal gene expression and markers are coming on line and expanding, the signs of stemness are lessening and the characteristic ESC profile is disappearing. A typical fluorescence-activated cell sorting analysis of such cells driven into the endodermal state shows high expression of SOX17 and FOXA2 in vitro (Fig. 2B lower). We would designate such in vitro developed cells as ASCs.
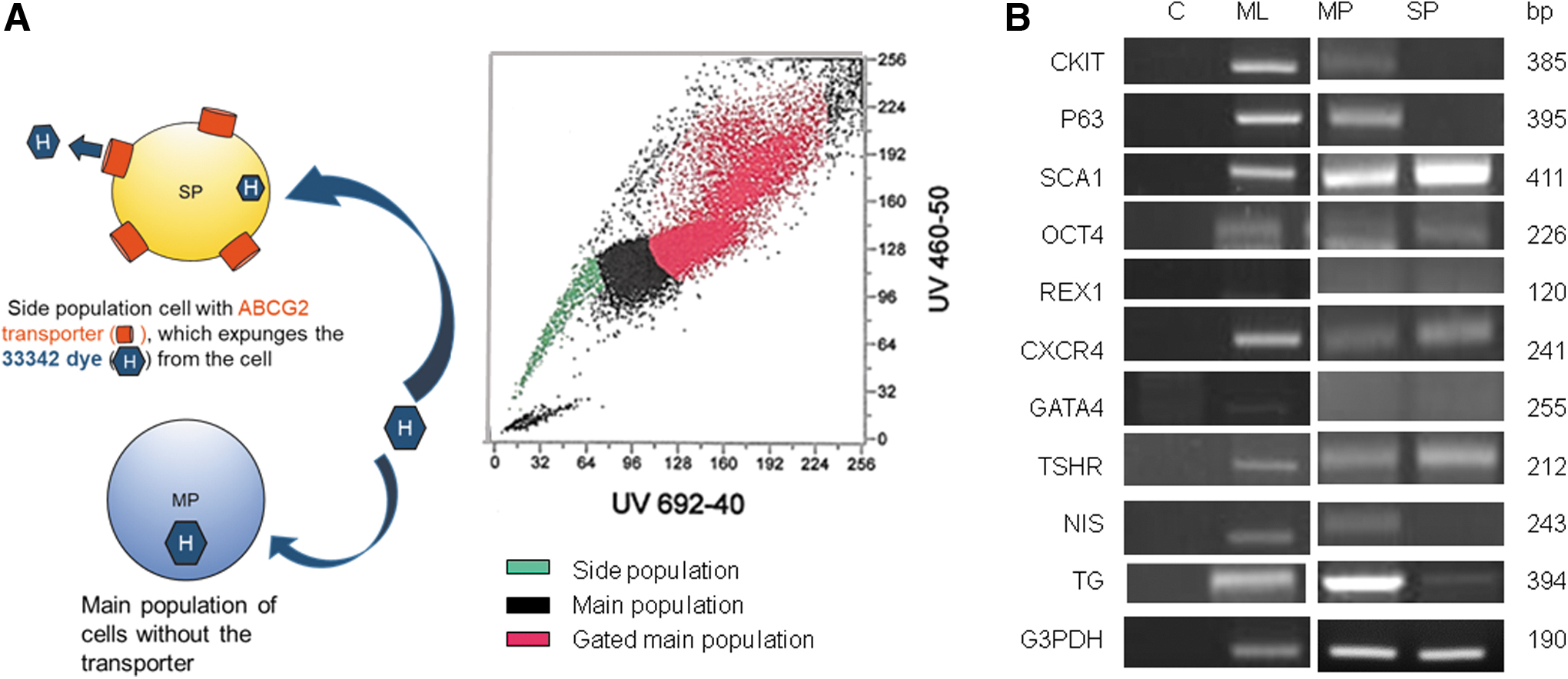
Identifying ASCs in mouse thyroid primary cultures. (
Thyroid Progenitor Cells
As discussed earlier, some of the stimuli capable of initiating the maturation of ASCs into thyroid progenitor cells have now been recognized. These include a major influence by the extracellular matrix in the niche involving a variety of well-defined growth factors and small molecules (11,20 –23), including IGF-1, FGF10 and also including the influence of thyrotropin (TSH) itself. This phenomenon has been known for many years from the use of “Matrigel”—a potent extracellular matrix preparation extracted from the Engelbreth-Holm-Swarm mouse sarcoma, a tumor rich in extracellular matrix proteins on which thyroid cells thrive (32,33). However, the heterogenous mixture of Matrigel and the lot–lot variation has made it clear that providing Matrigel alone is not sufficient to drive the thyroid progenitors further into differentiation. The way in which even defined factors influence thyroid progenitors is only partly understood but is clear from earlier in vitro studies from our laboratory (20,34,35) and others (21) and from tracing thyroid development in mice (36,37) that simultaneous expression of the transcription factors Pax8 and NKX2-1 appears to be unique to thyroid development (21,37) and is highly influenced by epigenetic changes (38). In vitro studies have confirmed that their expression induces such monopotency, leading to further differentiation into thyroid follicular cells (21,35). The expression of these transcription factors may be induced by direct transfection of gene constructs (21,35) or by exposure to a variety of the factors as listed earlier, including TSH. We have also described a small molecule (ethacridine) that is capable of such a stimulus (20). In Figure 2C, we show results from an mRNA-Seq of human stem cells further along on their continuum as they gain less of an endodermal profile and become early thyroid PSCs. As they develop NKx2-1 and Pax8 expression for the first time, it is this combination that signals their oligoclonality and thyroid speciation. This is the definition of a thyroid PSC that expresses both NKx2-1 and Pax8 (and usually Foxe1) and is only at the earliest stages of thyroid gene expression but having lost many of its stem cell markers (Figs. 2C and 4A). Further, such progenitor cells can be recognized in vivo by their larger nuclear size compared with follicular cells and by in situ staining for Pax8 expression with the absence of significant TG expression (Fig. 4B) (39). The time during which a cell passes through the progenitor stage is currently only known from the in vitro differentiation studies where it is ∼10–15 days.

Characterizing thyroid PSCs. (
Differentiation Into Thyroid Follicular Cells
Thyroid follicular cells express a profile of thyroid-related molecules (including TG, thyroid peroxidase [TPO], NIS, and the TSH receptor), which are only found to be co-expressed in a thyroid cell. For example, the TSH receptor is widely expressed in fibroblasts, adipocytes, bone cells, etc. (40 –42) but is only found with TG, TPO, and NIS in thyroid follicular cells. We, and others, have shown that ESCs and iPS cells, both mouse and human, can be differentiated into functional thyroid cells under well-defined conditions that are believed to mimic thyroid development (20 –23,35,43). Hence, such cells are no longer progenitor cells once they express high levels of thyroid related genes and their products. Figure 4A illustrates a series of quantitative polymerase chain reaction evaluations of selected genes that show the maturation of stem cells via progenitor cells to functional thyroid follicular cells. These induced thyroid follicular cells are capable of iodine uptake, iodine organification, and thyroid hormone synthesis and secretion (20,22,35,43). Such cells can be transplanted to maintain thyroid function in thyroid-deficient mice (21,23). We have used our TSH receptor knockout mice for such studies and have maintained almost normal thyroid function for many months (Fig. 5) (unpublished observations).
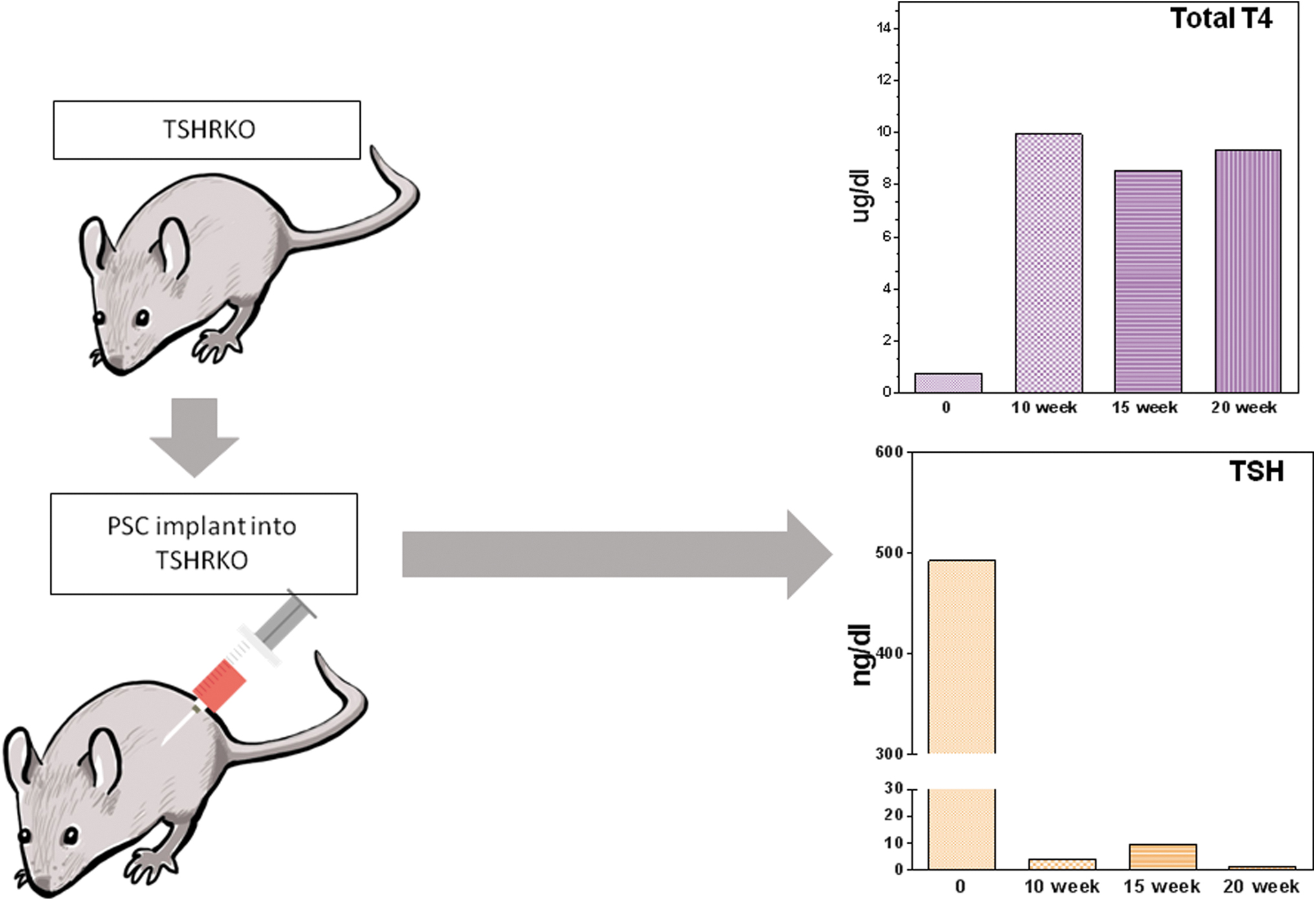
Rescue of the TSHR-KO with differentiated mouse ES cells. Heterozygous mouse ESCs were double transfected with Pax8-GFP and Nkx2-1-mcherry and differentiated as previously described (35). At 21 days, differentiated cells were then transplanted under the kidney capsule of hypothyroid TSHR-KO mouse and followed until 20 weeks with their T4 and TSH monitored periodically, as shown in the graphs on the left. The implanted cells rescued the hypothyroid phenotype of these animals and rendered them euthyroid (unpublished data). T4, thyroxine; TSHR-KO, TSH receptor knockout.
Summary Characteristics During Thyroid Cell Differentiation
As cells transverse their differentiation program, they each have well-recognized characteristics that are summarized in Table 1. While ESCs exhibit rapid proliferation (a doubling time in mice reported as ∼10 hours and human ESCs ∼30 hours) (44), there is progressively reduced proliferative potential as time passes. Indeed, the mature thyroid follicular cell is believed to only divide every 5–10 years (45). In the same way, the phenomenon of asymmetric division, designed to retain the clone of cells while still passing cells for further differentiation, is not seen in normal thyroid follicular cells. So, the time line of cell marker characteristics with the stem cells can be combined with their phenotypes to begin to understand the different potential of each stage of thyroid cell differentiation and the pivotal importance of obtaining a progenitor cell in thyroid development. Orchestrating the right signals at the right stage as seen in normal thyroid development is what can help us decipher their differentiation regulatory controls. This review correlates markers with the various stages of thyroid cell differentiation as windows into this process (Fig. 6) (44, 45, unpublished data).
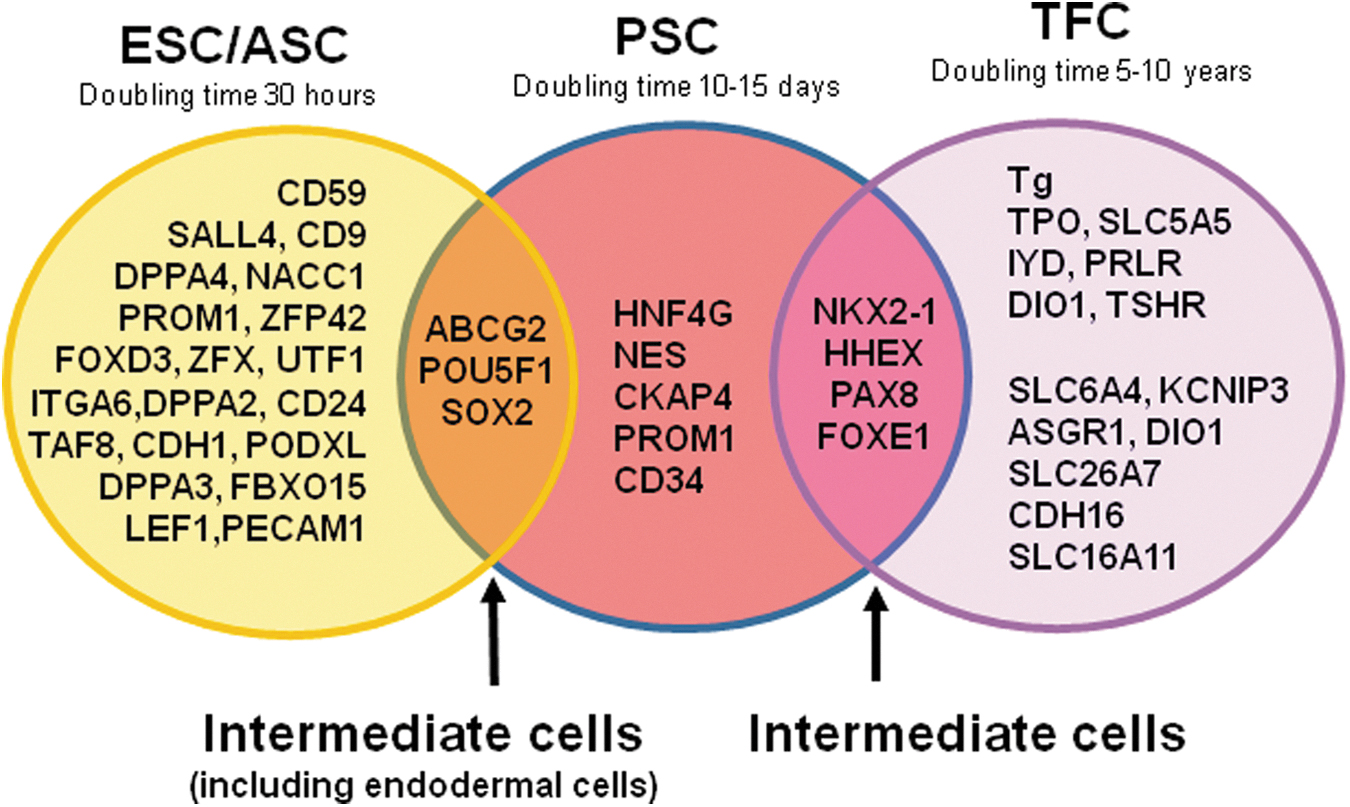
Venn-diagram illustrating the thyroid stem cell continuum by using gene expression profiles: ESC/ASCs, as outlined here on the left, express a cluster of genes that are unique to this cell population and that maintain their pluri- or multi-potent state. When they are stimulated intrinsically or extrinsically to form PSCs, they pass through a transition of gene expression (middle) that has overlaps with their parent of origin and also with the final destined cell (TFCs) that they are programmed to form (right). Hence, the complement of genes that determine their final phenotype is only formed after progressing through their transitional stages (44, 45, unpublished data).
Summary of Characteristics of the Thyroid Stem Cell Continuum
−, Negative; +, positive; +++, very positive.
ASC, adult stem cell; ESC, embryonic stem cell; PSC, progenitor stem cell; TFC, thyroid follicular cell.
Footnotes
Acknowledgments
The authors thank Drs. Syed Morshed and Mone Zaidi for continuous advice and support.
Authors' Contributions
T.F.D.: Designed experiments, performed data analysis, and article editing; R.L.: Designed experiments, data analysis, and article writing; R.S.: Designed experiments, data analysis; R.M.: Designed and performed the experiments, analyzed the data, and wrote the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was funded in part by the Segal Family Fund, Kronus, Inc. (Star, ID), a Department of Veteran Affairs Merit Award to Terry F. Davies and by NIH-NIDDK (DK069713).
Supplementary Material
Supplementary Data