
Abstract
Background:
Graves' ophthalmopathy (GO) is an autoimmune eye disease with the characteristic symptoms of eyelid retraction and proptosis. Orbital fibroblast activation induced by platelet-derived growth factor-BB (PDGF-BB) stimulation plays a crucial role in GO pathogenesis, leading to excessive proliferation and extracellular matrix production by orbital fibroblasts. Currently, GO treatment options remain limited and novel therapies including targeted drugs are needed. Histone deacetylases (HDACs) are associated with the development and progression of several cancers and autoimmune diseases by epigenetically controlling gene transcription, and HDAC inhibitors (HDACis) may have therapeutic potential. Nevertheless, the role of HDACs in orbital fibroblasts from GO is unknown. Therefore, we studied the expression of HDACs as well as their contribution to extracellular matrix production in orbital fibroblasts.
Methods:
Orbital tissues were obtained from GO patients (n = 18) who underwent decompression surgery with approval from the Institutional Review Board of the Faculty of Medicine (Protocol number 401/61), Chulalongkorn University (Bangkok, Thailand). Furthermore, orbital tissue was obtained from control patients (n = 3) without inflammatory or thyroid disease who underwent surgery for cosmetic reasons. Orbital fibroblast cultures were established from the orbital tissues. HDAC mRNA and protein expression in orbital fibroblasts was analyzed by reverse transcription–quantitative real-time PCR and Western blot. PDGF-BB-activated orbital fibroblast and orbital tissues were treated with HDACis or HDAC4 small-interfering RNA.
Results:
PDGF-BB-stimulated orbital fibroblasts had upregulated HDAC4 mRNA and protein expression. HDAC4 mRNA expression was significantly higher in GO compared with healthy control orbital fibroblasts. Histone H3 lysine 9 acetylation (H3K9ac) decreased upon PDGF-BB stimulation. Treatment with HDAC4i (tasquinimod) and HDAC4/5i (LMK-235) significantly decreased both proliferation and hyaluronan production in PDGF-BB-stimulated orbital fibroblasts. HDAC4 silencing reduced mRNA expression of hyaluronan synthase 2 (HAS2), collagen type I alpha 1 chain (COL1A1), Ki67, and α-smooth muscle actin (α-SMA), as well as hyaluronan production in PDGF-BB-stimulated orbital fibroblasts. Tasquinimod significantly reduced HAS2 and α-SMA mRNA expression in whole orbital tissue.
Conclusion:
Our data indicated, for the first time, that altered HDAC4 regulation along with H3K9 hypoacetylation might represent a mechanism that contributes to excessive proliferation and extracellular matrix production by orbital fibroblasts in GO. HDAC4 might represent a novel target for GO therapy.
Introduction
Graves' ophthalmopathy (GO) is an autoimmune eye disease characterized by volume expansion of the soft orbital tissues that surround the eyes and develops in ∼25–50% of Graves' disease (GD) patients (1,2). Clinical GO symptoms include upper eyelid retraction, proptosis, dysfunction of extraocular muscles, conjunctiva edema, and diplopia.
Orbital fibroblasts express thyroid-stimulating hormone receptors (TSHRs) on their surface and are activated by TSHR autoantibody stimulation, which contributes to the expansion and remodeling of the orbital soft tissues (3). Several investigators have also reported that GO orbital fibroblasts highly express the insulin-like growth factor 1 receptor (IGF-1R) that may act in concert with the TSHR (4 –6). Moreover, GO orbital fibroblasts can be activated by locally produced platelet-derived growth factor-BB (PDGF-BB) that stimulates proliferation, adipogenesis, cytokine and extracellular matrix production, as well as enhanced TSHR expression (7). All these processes contribute to the typical orbital tissue expansion and clinical signs of GO. Therefore, orbital fibroblasts play a critical role in GO pathogenesis (8).
Current treatments of GO consist of glucocorticoids and orbital decompression surgery lacking specificity in targeting orbital fibroblast activation (9). Treatment with the monoclonal antibody, teprotumumab, directed at IGF-1R showed clinical efficacy in treating GO and was recently approved by the US Food and Drug Administration (US-FDA) for GO treatment (10,11). However, a substantial proportion of patients will remain unresponsive to the limited available treatment options (12). Therefore, a better understanding into the molecular mechanisms underlying the pathophysiology of GO might reveal new targets for therapy.
Histone deacetylases (HDACs) are enzymes that play a role in the alteration of gene expression by removing the acetyl group from lysine residues on histone tails (13). Based on sequence homology and cellular localization, the zinc-dependent HDACs can be divided into different classes: class I HDACs (HDAC1–3 and 8), class IIa HDACs (HDAC4, 5, 7, and 9), class IIb HDACs (HDAC6 and 10), and class IV HDACs (HDAC11) (14).
Alterations in HDAC expression is associated with the development of various diseases, including cancer, fibrosis-associated disorders, and autoimmune diseases (15,16). Generally, class I HDACs are localized in the cell nucleus (17), whereas class II HDACs are present in both the cell nucleus and cytoplasm (18). With regard to autoimmune and fibrosis-associated disorders, class I and II HDACs contribute to the pathogenesis of rheumatoid arthritis and idiopathic pulmonary fibrosis by controlling fibroblast proliferation, and cytokine and extracellular matrix production (19,20). Accordingly, HDACs have become an attractive treatment target, which is currently under investigation in several clinical trials (21).
Also, in GD, increased HDAC1 and HDAC2 mRNA expression was recently reported in peripheral blood mononuclear cells (22). Interestingly, our recent study on DNA methylation in orbital fibroblasts points to a role for HDACs in controlling genes related to proliferation and cell-to-cell interaction in orbital fibroblasts from patients with active GO (23). However, so far no data on HDAC expression and their contribution to pathological process in GO orbital fibroblasts are available. Insight into HDACs' function in GO orbital fibroblasts could potentially lead to the development of therapeutic HDAC inhibition as an effective treatment by targeting pathogenic cells in both GD and GO.
HDAC inhibitors (HDACis) consist of pan-, class-selective, and isoform-selective HDACis. Pan-HDACis, such as SAHA and panobinostat, have been used as anticancer drugs with approval by the US-FDA for T cell lymphoma and multiple myeloma, respectively (24). In addition, class- and isoform-selective HDACis with potentially less side effects, such as HDAC1i (quisinostat) and HDAC4i (tasquinimod), were studied in patients with relapsed multiple myeloma and prostate cancer, respectively (25,26). Therefore, insight into the involvement of specific HDACs in the pathogenesis of GO could be helpful for improving effective treatment options for this disease with HDACis.
In this study, different HDACs were investigated in primary orbital fibroblasts isolated from GO patients and compared with control orbital fibroblasts. In addition, the effects of isoform-specific HDACis and HDAC silencing on orbital fibroblast proliferation, extracellular matrix production, and myofibroblast differentiation (alpha smooth muscle actin expression) were examined. This revealed that altered HDAC4 regulation along with histone H3 lysine 9 (H3K9) hypoacetylation occurs in orbital fibroblasts from GO and might represent a mechanism that contributes to excessive extracellular matrix production by orbital fibroblasts in GO. Therefore, the results from this study might potentially lead to HDAC4 targeted treatment for GO.
Methods
Reagents
Isoform-specific HDACis quisinostat (HDAC1i), CAY10683 (HDAC2i), RGFP966 (HDAC3i), tasquinimod (HDAC4i), LMK-235 (HDAC4/5i), tubastatin A (HDAC6i), and PCI-34051 (HDAC8i) were purchased from Selleck Chemicals, Inc. (TX). Recombinant human PDGF-BB was obtained from BioLegend, Inc. (CA).
Isolation of orbital fibroblasts from orbital tissues
Orbital tissues were obtained from GO patients (n = 18) who underwent decompression surgery at King Chulalongkorn Memorial Hospital (Bangkok, Thailand). Characteristics of the GO patients are shown in Supplementary Table S1. Control orbital tissues (n = 3) were obtained from individuals without thyroid or inflammatory disease who underwent cosmetic surgery. For isolation of orbital fibroblasts, the orbital tissues were sliced into small pieces (0.5 cm3/piece) and cultured in medium containing Dulbecco's modified Eagle's medium (DMEM; GE Lifesciences, MA) supplemented with 20% fetal bovine serum (FBS; Life Technologies, Inc., Germany) and 100 μg/mL gentamicin (Life Technologies, Inc., Germany) as described previously (27). After orbital fibroblast isolation, 10% FBS, DMEM, and 100 μg/mL gentamicin were used for further culture at 37°C and 5% CO2.
Orbital fibroblasts were used for experiments between the 4th and 12th passages. The fourth to fifth passages were used for the studies of HDACs and H3K9 acetylation (H3K9ac) expression level in orbital fibroblasts and HDAC4 knockdown experiments. Orbital fibroblasts used for the investigation of isoform-specific HDACi effect on PDGF-BB-induced orbital fibroblast activation were between the 5th and 12th passages.
Approval for the study was given by the Institutional Review Board of the Faculty of Medicine (Protocol number 401/61), Chulalongkorn University (Bangkok, Thailand).
HDAC mRNA expression in orbital fibroblasts
Orbital fibroblasts (control: n = 3 and GO: n = 9) were seeded at 4 × 105 cells/well into six-well plates (Thermo Fisher Scientific, Inc., NY) in DMEM containing 1% FBS and antibiotic overnight. The cultures were stimulated with PDGF-BB (50 ng/mL) for 1, 2, 4, 6, and 24 hours. Total RNA was extracted from the cells using the GenElute™ Total RNA Purification Kit according to the manufacturer's protocol (Sigma-Aldrich, Inc., MO) and converted to cDNA using the iScript™ cDNA Synthesis Kit according to the manufacturer's protocol (Bio-Rad, Inc., CA).
Expression levels of HDAC1, 2, 3, 4, 5, 6, and 8 were determined with TaqMan® Gene Expression Assays (Thermo Fisher Scientific, Inc.; Hs00606262_g1, Hs00231032_m1, Hs00187320_m1, Hs01041648_m1, Hs00608351_m1, Hs00997427_m1, and Hs00218503_m1, respectively) with SsoAdvanced™ Universal Probes Supermix (Bio-Rad, Inc.) by real-time PCR (CFX96 Touch™ Real-Time PCR Detection System; Bio-Rad, Inc.). HDAC expression levels were normalized to ABL. Primers and probe are shown in Supplementary Table S2.
HDAC4, HDAC5, and H3K9ac protein expression in orbital fibroblasts
Orbital fibroblasts (GO: n = 3–6) were seeded at 3 × 105 cells/well into six-well plates in DMEM containing 1% FBS and antibiotic for 24 hours. Subsequently, cells were stimulated with PDGF-BB (50 ng/mL) for 24 and 48 hours. Western blot was performed as previously described (28). Cells were lysed by 1 × RIPA lysis buffer (50 mM Tris-HCl pH 7.4, 150 mM NaCl, 1% Nonidet P-40, 10% sodium deoxycholate, 20% sodium dodecyl-sulfate [SDS]) containing 1 × protease/phosphatase inhibitor cocktail (Cell Signaling Technology, Inc., MA). Total protein concentration was measured by the Pierce™ BCA Protein Assay Kit (Thermo Fisher Scientific, Inc.).
Protein lysates (30 μg for HDACs and 10 μg for H3K9ac) were mixed with 2 × SDS dye (1 M Tris-HCl pH 6.8, 10% SDS, 20% (v/v) glycerol, bromophenol blue, and 10% β-mercaptoethanol) and heated at 100°C for five minutes. Proteins were loaded in 7.5% or 15% sodium dodecyl-sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and run with 100 V for 1.5 hours. Then proteins were transferred to PVDF membrane (Bio-Rad, Inc.) using semidry transfer Trans-Blot SD (Bio-Rad, Inc.) at 15 V for 30–45 minutes. Thereafter, the blotting membrane was blocked with 1× phosphate-buffered saline with 0.1% Tween-20 (PBST; Merck, Inc., Germany) containing 5% nonfat dry milk (Bio-Rad, Inc.).
Protein detection was performed using a primary antibody against HDAC4 (#7628, 1:2000), HDAC5 (#20458, 1:1000), H3K9ac (#9649, 1:1000), H3 (#4499, 1:1000), or β-actin (#4970, 1:2000) (all antibodies from Cell Signaling Technology, Inc.) and incubated overnight. Thereafter, the membrane was washed twice with 1× PBST for 5 minutes followed by two additional washes of 15 minutes. Then, the mouse anti-rabbit IgG (HRP conjugate) secondary antibody (#5127, 1:4000; Cell Signaling Technology, Inc.) was added and incubated for one hour with shaking before washing with 1 × PBST. Then the membrane was incubated with the Amersham ECL Western Blotting Detection Kit (GE Healthcare Life Sciences, Inc., MA) and visualization was done with the Alliance Q9 chemiluminescence imaging system (Uvitec, Inc., United Kingdom).
Lactate dehydrogenase cytotoxicity assay
Orbital fibroblasts were seeded at 6 × 103 cells/well into 96-well plates with 1% FBS DMEM for 24 hours to allow the cells to attach overnight. After that, cells were pretreated with HDACis for 24 hours. As for GO tissues, they were treated by the HDAC4i (tasquinimod) for 24 hours. Then supernatants were collected to determine cell toxicity by lactate dehydrogenase (LDH) assay according to the manufacturer's protocol (BioVision, Inc., CA), and the percentage of cytotoxicity was calculated relative to a negative and positive control.
Orbital fibroblast proliferation assay
Orbital fibroblasts (GO: n = 6) were seeded at 6 × 103 cells/well into 96-well plates with DMEM containing 1% FBS and antibiotic for 24 hours to allow the cells to attach overnight. For prophylactic effect, cells were pretreated with HDACis at concentrations that were not toxic to the cells for 24 hours and followed by further stimulation with PDGF-BB (50 ng/mL) for 24 hours. In an experimental setup more representative of treatment effect, cells were treated with HDACis simultaneously with PDGF-BB (50 ng/mL) for 24 hours. Proliferation was detected by colorimetric assay using methylene blue dye as described previously (29). Briefly, the cell culture supernatant was removed, followed by cell fixation with 50 μL of 20% formaldehyde for 48 hours. After removal of 20% formaldehyde, cells were stained with 50 μL of 1% methylene blue for 30 minutes. Cell bound dye was eluted with ice-cold 1:1 of hydrochloric acid: absolute ethanol solution and the absorbance was measured with microplate reader at 650 nm. The percentage proliferation was calculated relative to the unstimulated control.
Hyaluronan production by orbital fibroblasts
Orbital fibroblasts (GO: n = 7) were seeded at 1 × 105 cells/well into 12-well plates with DMEM containing 1% FBS and antibiotic for 24 hours. Subsequently, HDACis were added into the cultures for another 24 hours. Cells were further stimulated with PDGF-BB (50 ng/mL) for 24 hours. To explore the treatment potential of HDACis, the cells were treated simultaneously with HDACis and PDGF-BB (50 ng/mL) for 24 hours. Cell supernatants were collected, and hyaluronan was measured by enzyme-linked immunosorbent assay (ELISA; R&D system, Inc., MN) according to the manufacturer's protocol.
HDAC4 knockdown in orbital fibroblasts
Orbital fibroblasts (GO: n = 3–6) were seeded at 3 × 105 cells/well into six-well plates in antibiotic-free DMEM containing 1% FBS for 24 hours. Cells were transfected with 50 nM HDAC4 small-interfering RNA (siRNA, ON-TARGETplus Human HDAC4 siRNA; Dharmacon, Inc., Lafayette, CO) or nontargeting siRNA (ON-TARGETplus Non-targeting siRNA #1; Dharmacon, Inc.) as control with DharmaFECT 1 transfection reagent (Dharmacon, Inc.). After 48 hours, cells were further stimulated with PDGF-BB (50 ng/mL) for 48 hours. Then cells were extracted for the examination of either mRNA expression (HDAC4, HDAC5, hyaluronan synthase 2 [HAS2], collagen type I alpha 1 chain [COL1A1], the proliferation marker Ki67 [Ki67], interleukin 6 [IL-6], α-smooth muscle actin [α-SMA], and histone acetyltransferase P300 [p300]) by reverse transcription–quantitative real-time PCR (RT-qPCR) or protein expression (HDAC4 and HDAC5) by Western blot. In addition, cell supernatants were collected for hyaluronan (R&D systems, Inc.) and IL-6 (BioLegend, Inc.) measurements by ELISA.
HDAC4i (tasquinimod) and HDAC4/5i (LMK-235) effect on GO orbital tissues
Orbital tissues (GO: n = 3–8) were sliced into small pieces (0.5 cm3/piece), then placed into 24-well plates with at least five pieces/well. Subsequently, the tissues were cultured with 1% FBS DMEM in the presence or absence of 20 μM HDAC4i (tasquinimod) or 1 μM HDAC4/5i (LMK-235) and incubated for 24 hours, as reported previously (30). Orbital tissues were disrupted with pestle motor mixer (Cole-Parmer, Inc.) for 5 minutes and 200 μL 1-bromo-3-chloropropane (Sigma-Aldrich, Inc.) was added and incubated for 3 minutes followed by centrifugation (11,600 g, 4°C, 15 minutes). Total RNA was subsequently extracted using the RNeasy Mini Kit according to the manufacturer's protocol (QIAGEN, Inc., United Kingdom). Total RNA was reverse transcribed to cDNA using TaqMan™ Reverse Transcription Reagents according to the manufacturer's instructions (Life Technologies, Inc., NY).
COL1A1, HDAC4, p300, IL-6, HAS2, and α-SMA mRNA levels were determined with SsoAdvanced™ Universal Probes Supermix (Bio-Rad, Inc.) by RT-qPCR as described above. Furthermore, tissue supernatants were collected for measurements of LDH cytotoxicity (BioVision, Inc.) and hyaluronan production (R&D systems, Inc.). The mRNA expression levels and hyaluronan levels in the supernatants were calculated compared with the untreated orbital tissues and expressed as percentage.
Statistical analysis
Data are presented as mean ± the standard error of the mean and analyzed by paired t-test for all GO orbital fibroblast experiments, or unpaired t-test in the experiment that compared GO with control orbital fibroblasts. A p-value <0.05 was considered to indicate a statistically significant difference.
Results
Effect of PDGF-BB stimulation on HDAC expression by orbital fibroblasts
All tested HDACs were expressed by the GO orbital fibroblasts under basal culture conditions, with HDAC2 mRNA being expressed at the highest level and HDAC4 mRNA being expressed at the lowest level. PDGF-BB stimulation slightly, yet significantly, enhanced mRNA expression levels of class I HDACs; HDAC1, HDAC3 and class IIa HDAC; HDAC4 at 6 and 24 hours after stimulation (p < 0.05, p < 0.01, and p < 0.01, respectively; Fig. 1A). On the contrary, the HDAC5 mRNA expression level significantly decreased at 24 hours in PDGF-BB-stimulated orbital fibroblasts (p < 0.05; Fig. 1A). HDAC4 protein significantly increased at 48 hours (p < 0.01) after PDGF-BB stimulation, while HDAC5 protein significantly decreased at both 24 and 48 hours after PDGF-BB stimulation (p < 0.05 and p < 0.05, respectively; Fig. 1B).
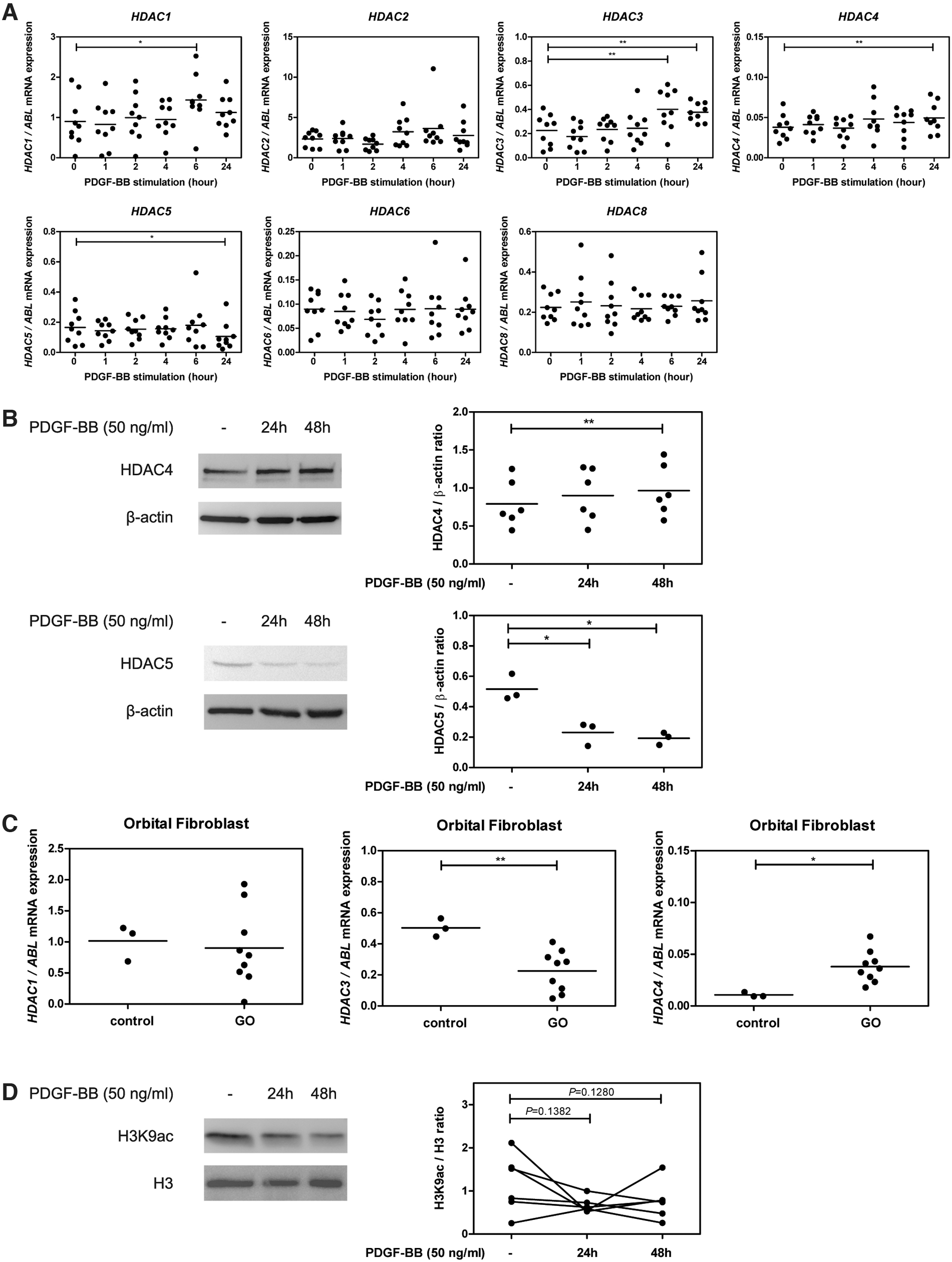
HDAC and H3K9ac expression level in orbital fibroblasts. (
To explore whether differences in basal expression existed for PDGF-regulated HDACs between GO and control orbital fibroblasts, HDAC mRNA expression in primary orbital fibroblasts was further measured. HDAC4, but not HDAC1 and 3, was detected at higher levels in primary orbital fibroblasts isolated from GO compared with healthy control (p < 0.05; Fig. 1C). HDAC4 specifically deacetylates H3K9, and PDGF-BB stimulation did result in H3K9 hypoacetylation in GO orbital fibroblasts at both 24 and 48 hours (p = 0.1382 and p = 0.1280, respectively; Fig. 1D).
Prophylactic and treatment effects of HDAC4i on PDGF-BB-induced orbital fibroblast activation
The inhibitory effect of isoform-selective HDACis on PDGF-BB-induced orbital fibroblast activation was investigated. The HDACis were nontoxic at all tested concentrations and did not affect the basal proliferation rate and hyaluronan production of the orbital fibroblasts (Supplementary Figs. S1 and S2).
Pretreatment of the orbital fibroblasts with the maximum concentration of the class I HDACi; HDAC3i (RGFP966) and class IIa HDACis; HDAC4i (tasquinimod) and HDAC4/5i (LMK-235) significantly reduced PDGF-BB-induced orbital fibroblast proliferation (p < 0.01, p < 0.05, and p < 0.05, respectively; Fig. 2A and Supplementary Fig. S3A). In contrast, class I HDACis; HDAC1i (quisinostat), HDAC2i (CAY10683), HDAC8i (PCI-34051) and class IIb HDACis; HDAC6i (tubastatin A) did not affect PDGF-BB-induced orbital fibroblast proliferation (Supplementary Fig. S3A).
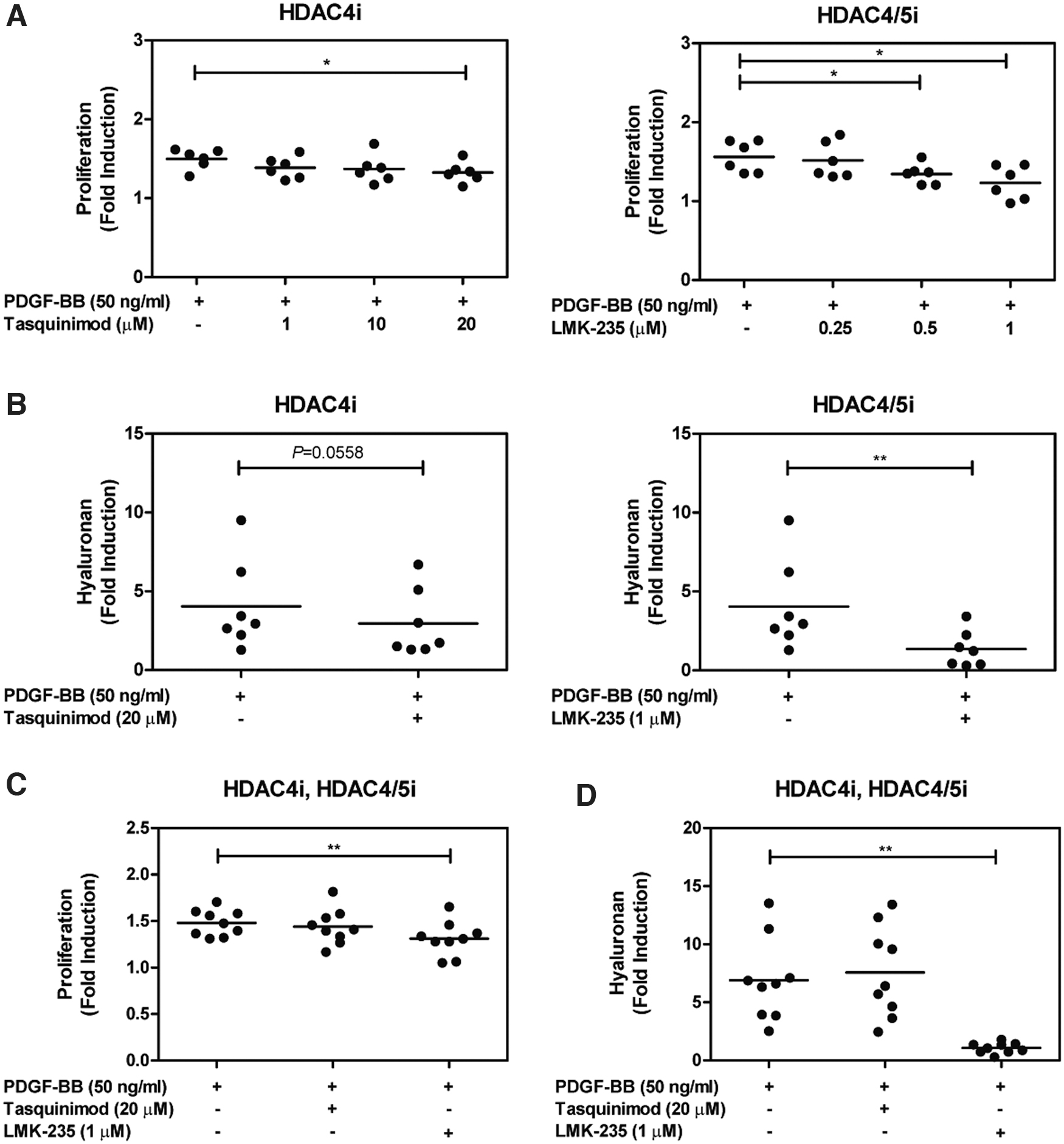
HDAC4 effect on PDGF-BB-induced orbital fibroblast activation. Orbital fibroblasts were pretreated for 24 hours with different HDACi concentrations of class IIa HDACis; tasquinimod and LMK-235, followed by 50 ng/mL PDGF-BB stimulation for 24 hours. (
In addition, HDAC4i (tasquinimod), HDAC4/5i (LMK-235), as well as HDAC2i (CAY10683) significantly decreased hyaluronan production in orbital fibroblasts stimulated by PDGF-BB (p = 0.0558, p < 0.01, and p < 0.05, respectively; Fig. 2B and Supplementary Fig. S3B), while HDAC1i, HDAC3i, HDAC6i, and HDAC8i did not affect PDGF-BB-induced hyaluronan production by orbital fibroblasts (Supplementary Fig. S3B).
HDACis targeting HDAC4 and found effective in the prophylactic experimental setup (tasquinimod and LMK-235) were subsequently tested in a set of experiments more representative for treatment. Only HDAC4/5i (LMK-235) significantly decreased PDGF-BB-induced orbital fibroblast proliferation (p < 0.01; Fig. 2C) and hyaluronan production (p < 0.01; Fig. 2D).
Effect of HDAC4 silencing in PDGF-BB-induced orbital fibroblast activation
To test whether the inhibitory effect of HDAC4i (tasquinimod) and HDAC4/5i (LMK-235) on orbital fibroblast activation was mediated through HDAC4 inhibition, HDAC4 siRNA silencing experiments were conducted as tasquinimod did not affect HDAC4 expression (Supplementary Fig. S4). HDAC4 basal mRNA and protein expression levels were lower in comparison with nontargeting siRNA-treated orbital fibroblasts (p < 0.05 and p < 0.01, respectively; Fig. 3A, B). In addition, no off-target effect of HDAC4 siRNA on HDAC5 was observed at both mRNA and protein levels (Supplementary Fig. S5).
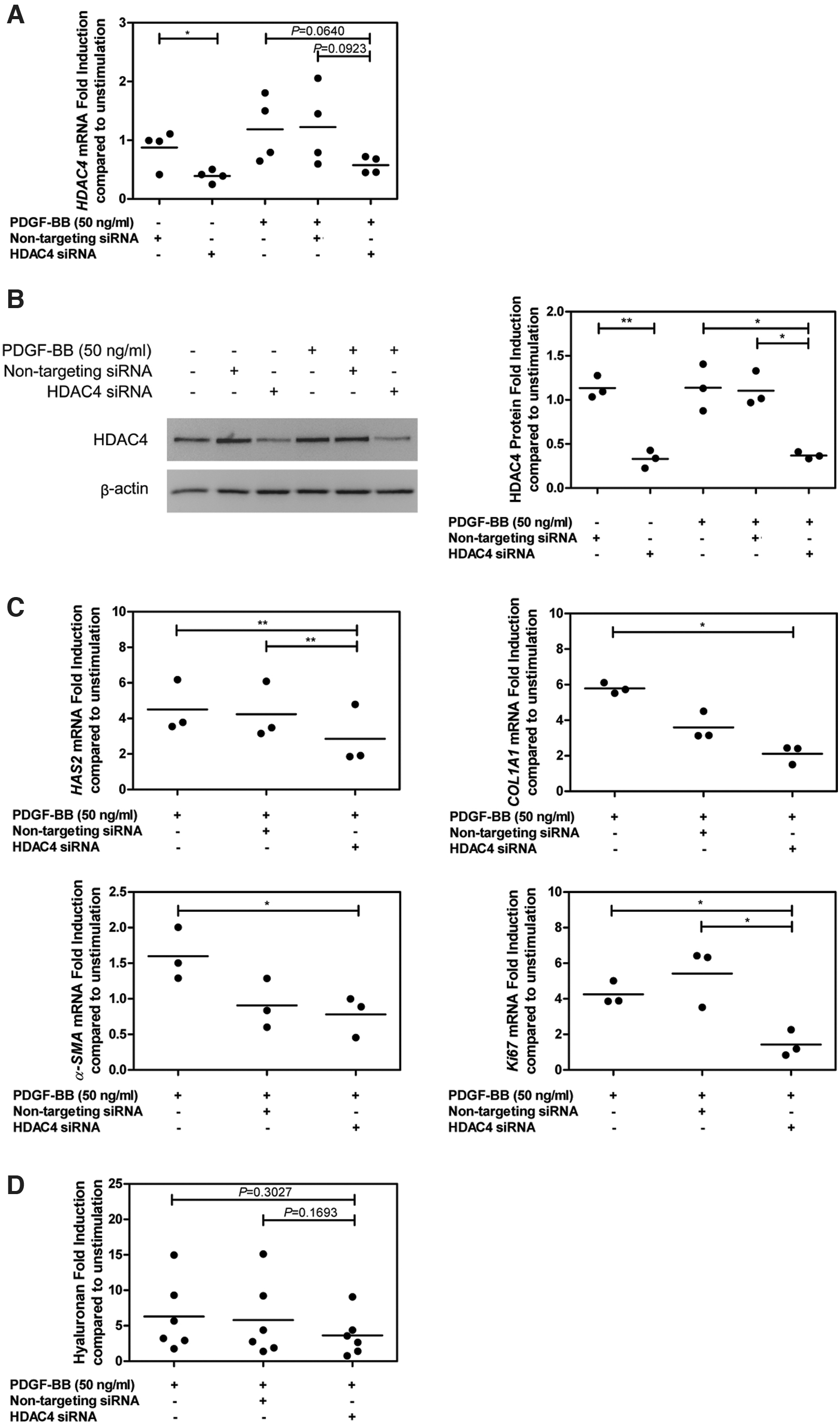
HDAC4 knockdown and its effects in PDGF-BB-stimulated orbital fibroblast activation. Orbital fibroblasts (GO: n = 3–6) were transfected by HDAC4 siRNA or nontargeting siRNA with 50 ng/mL PDGF-BB for 48 hours. (
HDAC4 silencing did not alter PDGF-BB-induced IL-6 and histone acetyltransferase P300 (p300) mRNA expression and IL-6 production (Supplementary Fig. S6A, B). On the contrary, HAS2, COL1A1, Ki67, and α-SMA mRNA expression levels were significantly decreased compared with PDGF-BB stimulation (p < 0.01, p < 0.05, p < 0.05, and p < 0.05, respectively; Fig. 3C). Likewise, the hyaluronan level was slightly decreased in culture supernatants from HDAC4-silenced PDGF-BB-stimulated orbital fibroblasts (p = 0.3027; Fig. 3D). HAS2 mRNA expression also tended to correlate with the level of hyaluronan in culture supernatants from PDGF-BB-stimulated orbital fibroblasts treated with HDAC4 siRNA (Supplementary Fig. S6C).
HDAC4i (tasquinimod) and HDAC4/5i (LMK-235) effect on GO orbital tissues
Orbital tissues from GO patients were treated with HDAC4i (tasquinimod) or HDAC4/5i (LMK-235) for 24 hours to test its efficacy in an ex vivo disease setting. HDAC4i (tasquinimod) and HDAC4/5i (LMK-235) concentrations used were not toxic to the orbital tissues (Supplementary Figs. S7A and S8A). HDAC4i (tasquinimod) significantly reduced HAS2 and α-SMA mRNA expression levels in whole orbital tissue culture compared with untreated orbital tissues (p < 0.05 and p < 0.01, respectively; Fig. 4A), while it did not affect HDAC4, p300, COL1A1, and IL-6 mRNA expression (Supplementary Fig. S7B). Besides, tasquinimod slightly, yet nonsignificantly, reduced hyaluronan production (mean = 74.93%) compared with untreated orbital tissues (p = 0.0914; Fig. 4B). HDAC4/5i (LMK-235) did not affect the expression of any of the mRNA transcripts measured nor did it affect the hyaluronan production (Supplementary Fig. S8B, C).
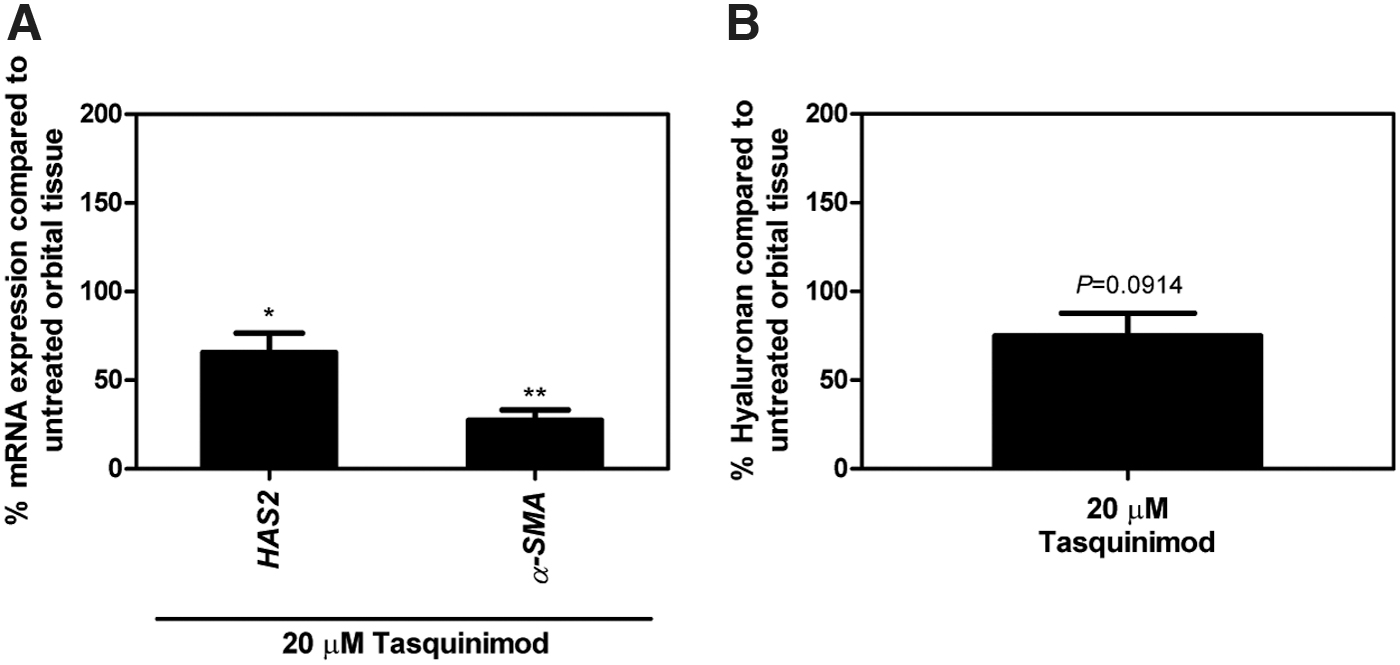
HDAC4i (tasquinimod) treatment in GO orbital tissues. Tissues from GO patients were treated with or without 20 μM HDAC4i (tasquinimod) in 1% FBS DMEM for 24 hours. (
Discussion
In the last decade, histone deacetylation has been shown to have a role in the pathogenesis of several diseases, including cancer, fibrosis-associated disorders, and autoimmune diseases, which led to the development of HDACis to control inflammation, cellular proliferation, apoptosis, as well as myofibroblast differentiation/activation (16,21,31). Although GO shares a pathological overlap with such diseases, there are currently no data available on HDACs in GO pathogenesis. Hence, this is the first study to reveal altered HDAC regulation in orbital fibroblasts in relation to GO pathogenesis.
Both class I and II HDACs contribute to several autoimmune and fibrosis-associated disorders, such as in idiopathic pulmonary fibrosis, rheumatoid arthritis, and systemic sclerosis, through enhancement of local fibroblast activity (19,20,32). In addition, several fibrosis-related growth factors such as TGF-β and PDGF have been found to induce aberrant HDAC expression in fibroblasts (33,34). Therefore, we here investigated class I and II HDACs in orbital fibroblasts in relation to the pathogenesis of GO.
PDGF-BB represents an important pathogenic growth factor in GO (7). In this study, we demonstrate that PDGF-BB increases the expression of HDAC1 and HDAC3 (class I HDACs) and HDAC4 (a class IIa HDAC) in orbital fibroblasts isolated from GO patients (Fig. 1A). Consistent with a previous report in rheumatoid arthritis fibroblast-like synoviocytes (35), downregulation of HDAC5 (a class IIa HDAC) was observed in the current study (Fig. 1A, B). HDAC1, 3, and 4 activities have previously been shown to decrease H3K9ac level (36 –38). In line with this, we found that PDGF-BB reduced H3K9ac level in orbital fibroblasts (Fig. 1D). Therefore, these data suggest that HDAC4-induced H3K9 hypoacetylation aggravates orbital fibroblast activation by regulating global transcription involved in the pathogenesis of GO.
Screening libraries of HDACi in PDGF-BB-induced orbital fibroblast activation revealed that HDAC isoform inhibitors, including HDAC3i (RGFP966), HDAC4i (tasquinimod), and HDAC4/5i (LMK-235), significantly decreased PDGF-BB-induced orbital fibroblast proliferation in a prophylactic setting (Fig. 2A and Supplementary Fig. S3A), while HDAC2i (CAY10683), HDAC4i (tasquinimod), and HDAC4/5i (LMK-235) significantly decreased PDGF-BB-induced hyaluronan production by orbital fibroblasts (Fig. 2B and Supplementary Fig. S3B). In the experimental setup more representative of a treatment scenario, only HDAC4/5i (LMK-235) significantly reduced proliferation and hyaluronan production (Fig. 2C, D). Therefore, HDAC4 seems to be the most interesting HDAC in controlling orbital fibroblast activation.
Although there are no data on the IC50 of HDAC4i (tasquinimod), our results suggest that HDAC4/5i (LMK-235) was more effective in controlling PDGF-BB-induced orbital fibroblast activation than HDAC4i (tasquinimod). Yet, based on the clinical observation of HDAC4i (tasquinimod) to induce minor side effects in prostate cancer patients, including fatigue, nausea, and constipation (39,40), HDAC4i (tasquinimod) was further tested for its efficacy in another experimental setting using GO whole orbital tissue culture. In the orbital tissues, HDAC4i (tasquinimod) did not alter mRNA expression levels of HDAC4, p300, COL1A1, and IL-6, but significantly decreased HAS2 and α-SMA mRNA expression and slightly reduced hyaluronan production compared with untreated orbital tissues (Fig. 4 and Supplementary Fig. S7).
In contrast, HDAC4/5i (LMK-235) did not show any significant effect in whole orbital tissues (Supplementary Fig. S8B, C). This might be due to the small number of samples tested (especially in the case of HDAC4/5i (LMK-235), n = 3) as well as the clinical stage of disease in these patients. The concentration of HDAC4/5i (LMK-235) was also increased to 10 μM; however, it caused cytotoxicity to GO orbital fibroblasts and tissues (data not shown). As GO orbital tissues contain different cell types apart from orbital fibroblasts, including T lymphocytes, B lymphocytes, macrophages, and mast cells, this might interfere with the effects of inhibitors on orbital tissues (7). Nevertheless, our data do show the ability of HDAC4i (tasquinimod) to inhibit the expression of molecules involved in GO pathogenesis in orbital tissues. Therefore, this finding could provide a rationale for clinical testing of HDAC4i (tasquinimod) efficacy in GO patients.
The involvement of HDAC4 in PDGF-BB-induced GO orbital fibroblast activation was confirmed by HDAC4 silencing experiments. Although HDAC4 and HDAC5 display high homology (over 70% similarity) (41), HDAC4 knockdown did not alter HDAC5 mRNA and protein expression (Supplementary Fig. S5). HDAC4 silencing did, however, not prevent PDGF-BB-induced p300 and IL-6 mRNA levels as well as IL-6 production (Supplementary Fig. S6), while it did inhibit HAS2, COL1A1, α-SMA, and Ki67 mRNA expression (Fig. 3C). This result is in line with a previous study indicating that class I and II HDACs interfered with the synthesis of collagen type I and α-SMA protein expression in TGF-β1-induced skin myofibroblast differentiation (42,43).
Although hyaluronan production was significantly inhibited in PDGF-BB-stimulated orbital fibroblasts in the presence of HDACi, statistically significant inhibition was not reached upon siRNA-mediated HDAC4 silencing. This latter might be because of incomplete knockdown of HDAC4 (see Fig. 3B, which demonstrates ∼75% knockdown) even at seven days after siRNA transfection. Nevertheless, we did observe a trend toward a positive correlation between HAS2 mRNA and hyaluronan production in HDAC4-silenced PDGF-BB-stimulated orbital fibroblasts (Supplementary Fig. S6C). Further experiments on promoter activities of genes associated with hyaluronan production, HAS2, and its transcription factors; sp1 and sp3 would provide insight in HDAC4 involvement in hyaluronan synthesis (44).
Taken together, these are the first data to show that HDAC4 plays vital roles in regulating the proliferation and production of extracellular matrix molecules by orbital fibroblasts in GO. We propose that HDAC4 regulates both extracellular matrix production and proliferation in PDGF-BB-induced orbital fibroblasts via H3K9ac, as well as other lysine positions, which require further study. Therefore, HDAC4 could be a new potential target of GO treatment.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported by the Ratchadapisek Sompoch Endowment (760001-HR), the 72nd anniversary of his Majesty King Bhumibol Adulyadej, and the 90th anniversary of Chulalongkorn University Funds, Chulalongkorn University, Thailand as well as the TSRI Fund (CU_FRB640001_01_23_1) and the National Research Council of Thailand.*
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Supplementary Figure S7
Supplementary Figure S8
Supplementary Table S1
Supplementary Table S2