
Abstract
Background:
The pathogenesis of thyroiditis caused by immune-checkpoint inhibitors (ICIs) such as antiprogrammed death receptor-1 (PD-1) and anticytotoxic T lymphocyte antigen-4 (CTLA-4) is incompletely understood. To gain mechanistic insights, we developed a mouse model of ICI-related thyroiditis and assessed the clinical, hormonal, and cytokine profiles.
Methods:
Forty NOD-H2h4 mice, 112 days old at the start of the experiments, were divided into two sequential cohorts. In the first one (No. = 21), mice were injected with both anti-PD-1 and anti-CTLA-4 checkpoint inhibitors while drinking either regular water or iodine-supplemented water. In the second cohort (No. = 19), mice were injected with either anti-PD-1 or anti-CTLA-4 while drinking iodine-supplemented water. Mice were sacrificed two months after the initial injection to collect thyroid gland for histopathology (to assess thyroiditis severity) and flow cytometry (to identify immune cell subsets and tissue-resident memory T cell markers). Mice were also studied before sacrifice to determine thyroid area and structure (by ultrasound), thyroid function (serum total thyroxine, thyrotropin, thyroid antibodies), and cytokine profile (by bead-based Luminex technology).
Results:
Thyroiditis was more severe upon PD-1 than CTLA-4 blockade (p = 0.01) and significantly correlated with the number of CD45+ cells infiltrating the thyroid (cumulative odds ratio [OR] 1.2 [95% confidence interval, CI 1.1–1.3], p < 0.001, that is 20% greater odds of a higher severity score for every 170-unit increase in CD45 infiltrating cells). Thyroiditis was instead more prevalent (100% vs. 63%, p < 0.01) in the anti-CTLA-4 mice, which also showed a larger thyroid area (17 ± 8.2 mm) than those treated with anti-PD-1 (11 ± 4.2 mm) and controls (p < 0.01). Serum IL-6 was markedly increased upon PD-1 blockade (40 pg/mL at baseline, 198 pg/mL on day 172), an increase not seen in the anti-CTLA-4 group (p = 0.01). IL-6 mirrored thyroiditis severity, with highest serum values found in greatest histopathology scores (cumulative OR 1.1 [CI 1.02–1.15], p = 0.009). GM-CSF and MIP1β increased more in the anti-CTLA-4 group (p < 0.001 for both), whereas the other cytokines did not differ among the treatment groups.
Conclusions:
The study reports a mouse model of thyroiditis induced by PD-1 blockade and, comparing it to the anti-CTLA-4 model, uncovers distinctive histopathological, sonographic, hormonal, and immunological features, offering biomarkers, such as serum IL-6, that could be used in the clinical setting.
Introduction
Immunotherapy has long been used to treat cancer but has seen the greatest popularity only in the last decade following the discovery of “immune-checkpoint inhibitors (ICIs),” such as the monoclonal antibodies blocking cytotoxic T lymphocyte antigen-4 (CTLA-4) or programmed death receptor-1 (PD-1). Thyroid dysfunctions secondary to ICIs are among the most common immune-related adverse events (1), and have been classically associated with blockade of the PD-1/PD-L1 pathway. In most human studies, these thyroid dysfunctions are reported dichotomously as either hypothyroidism or hyperthyroidism without information about thyroid autoantibodies or thyroid ultrasound, key pillars in thyroid diagnosis (2). In addition, reports are often assembled by oncologists who may not be familiar with the subtle aspects of thyroid pathophysiology.
To address these shortcomings, we assessed the effects of CTLA-4 and PD-1 blockade in the NOD-H2h4 mouse, a model of iodine-induced autoimmune thyroiditis.
Materials and Methods
Mouse cohorts and experimental groups
The study included a total of 40 NOD-H2h4 mice (21 M and 19 F), 16 weeks (112 days) old at the start of the experiment, which were divided into two sequential cohorts (Supplementary Fig. S1A). The first cohort included 21 mice (9 M and 12 F) that drank either regular water (No. = 11) or water supplemented with 0.05% sodium iodide (No. = 10) for eight weeks. Of them, 10 of 10 in the iodine group and 8 of 11 in the regular water group were injected with two monoclonal antibodies, one blocking PD-1 and the another CTLA-4, while the remaining 3 mice with an isotype control. The second cohort included 19 mice (12 M and 7 F) drinking iodine-supplemented water that were injected with either the monoclonal antibody blocking PD-1 (No. = 8), or the one blocking CTLA-4 (No. = 8), or the isotype control (No. = 3).
Source of the anti-PD-1 and anti-CTLA-4 monoclonal antibodies and the administration schedule are available in Supplementary Data and Supplementary Figure S1B. The protocol was approved by the Animal Care and Use Committee of the Johns Hopkins University.
Thyroid ultrasound
We performed thyroid ultrasound using a high-frequency MS700 embryonic probe (30–70 MHz), connected to the Vevo 2100 microimaging system (VisualSonics). For each mouse, we analyzed echogenicity [scored as iso-, hypo-, hyperechoic with respect to the adjacent muscles, considering hyperechoic the normal appearance (3)], homogeneity (scored as homogeneous or inhomogeneous), and thyroid area (ellipse formula).
Serum levels of thyroid hormone and thyroid antibodies
Thyrotropin (TSH), total thyroxine (T4), and total triiodothyronine (4), antibodies directed against mouse thyroglobulin (mTg) (4,5) or thyroperoxidase (mTPO) (6,7) were measured as published, and detailed in Supplementary data.
Thyroid histopathology
Thyroid sections were scored from 0 to 4 according to the severity of mononuclear cell infiltration, as published in Di Dalmazi et al. (4) and Caturegli et al. (8) and detailed in Supplementary Data.
Characterization of immune cell subsets and tissue-resident memory T cell markers by flow cytometry is described in Supplementary Data.
Serum cytokines
Serum levels of cytokines (GM-CSF, INF-g, Mip1-β, TNF-α, IL-6, IL-2, IL-12(p40), MCP-1, IP-10) were measured by Luminex technology, using a customized kit (catalog number MCYTOMAG-70K from Millipore), following the manufacturer's recommendations.
Statistical analyses
The primary outcome of the study was thyroiditis severity and incidence, assessed by histopathological examination of one thyroid lobe, as well as flow cytometric enumeration of the thyroidal CD45+ hematopoietic cells. Secondary outcomes included ultrasonographic features, serum levels of T4, TSH, and thyroid antibodies, distribution of lymphoid subsets, and serum levels of cytokines. Differences in outcomes among the experimental groups were assessed first by Kruskal–Wallis, and then by Wilcoxon rank-sum test to perform pairwise comparisons. Simple ordered logistic regression assessed the relationship between thyroiditis histopathological score and hematopoietic cell counts. Multiple ordered logistic regression was used to define the cytokine signature associated with thyroiditis.
Collinearity was excluded calculating variance inflation factors, and Brant test verified the parallel regression assumption in both single and multiple analyses. Statistical analyses were performed using Stata statistical software, release 15.1 (Stata, College Station, TX).
Results
PD-1 and CTLA-4 blockade induces and exacerbates lymphocytic thyroiditis with distinct pathological characteristics
We began by assessing the ability of the anti-PD-1 and anti-CTLA-4 checkpoint inhibitors to induce thyroiditis in NOD-H2h4 mice, a strain genetically susceptible to develop thyroiditis at low incidence, but where thyroiditis can be markedly exacerbated by the administration of iodine in the drinking water.
When used together, anti-PD-1 and anti-CTLA-4 caused a significantly more severe thyroiditis than isotype-injected controls, especially in mice drinking iodinated water (Fig. 1A, p = 0.02). Notably, the sole injection of the two checkpoint inhibitors promoted thyroiditis even without iodine supplementation, thyroiditis that developed in 5 of 8 antibody-injected mice (63%) and in 0 of 3 isotype-injected controls (p = 0.06 by Pearson chi-squared test).
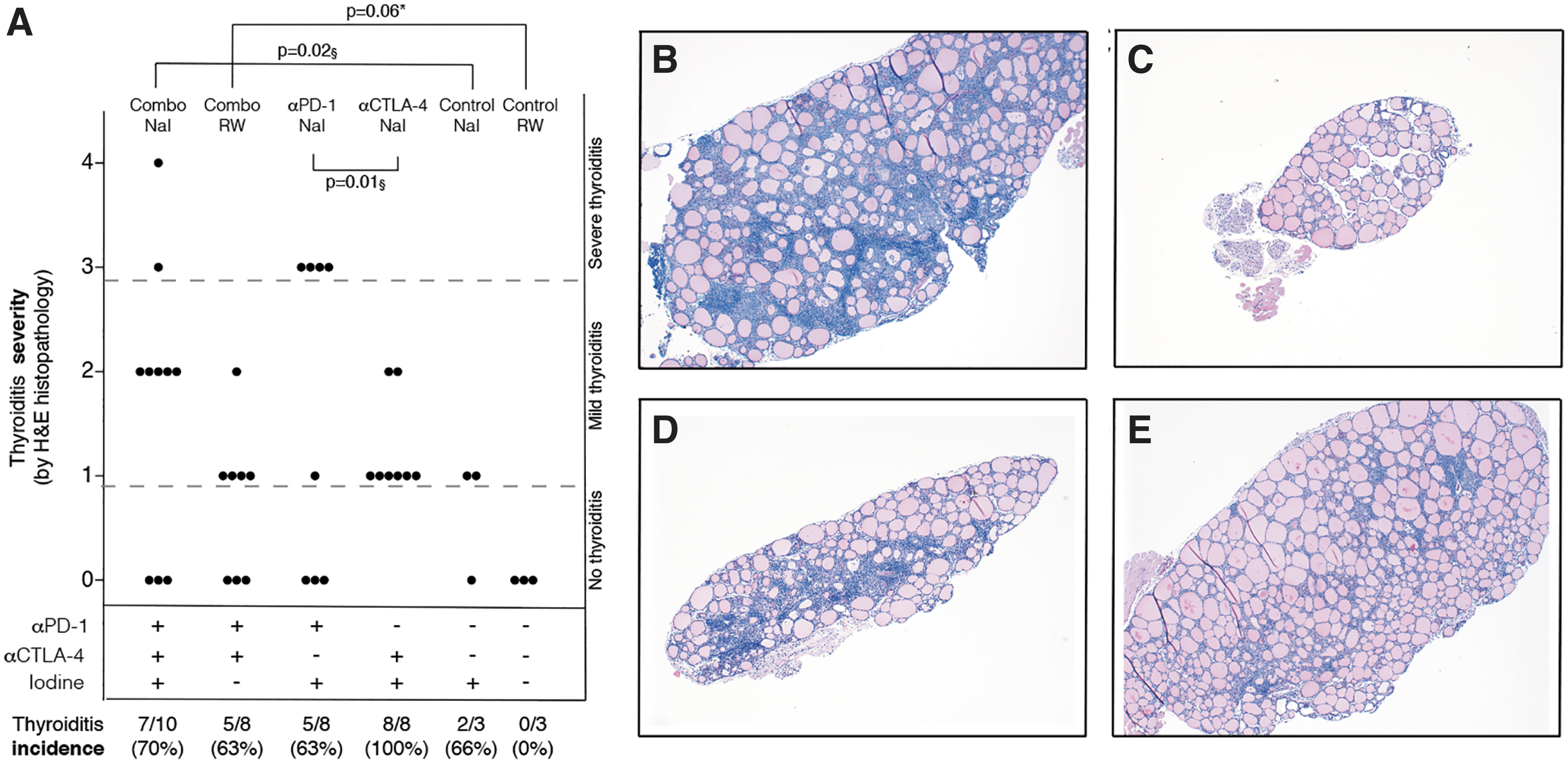
Histological comparison of ICI-treated vs. untreated mice. Histological thyroiditis severity in (i) mice treated with both ICIs and drinking either iodinated water (Combo NaI) or regular water (Combo RW); (ii) mice treated with one ICI and drinking iodinated water (αPD-1 NaI and α-CTLA-4 NaI); (iii) mice injected with the isotype control monoclonal antibody and drinking either iodinated water (Control NaI) or regular water (Control RW). p-Value based on Wilcoxon pairwise rank-sum test. §Comparison between thyroiditis mice. *Comparison of thyroiditis as a categorical variable, p based on Pearson chi-squared test (histopathological severity in thyroiditis mice was not analyzed in this comparison since no mice in the regular water control group developed thyroiditis) (
ICI administration tended to exacerbate thyroiditis severity, which increased from a median of 1 (iqr from 0 to 1) in cases drinking regular water to a median of 2 (iqr from 0 to 2) in those drinking iodinated water (Fig. 1A). Histopathology showed a diffuse mononuclear cell infiltration in cases that often disrupted the normal follicular architecture (Fig. 1B), which instead was preserved in mice injected with isotype control and drinking regular water (Fig. 1C).
Comparison of thyroiditis in iodine-drinking mice treated with anti-PD-1 or anti-CTLA-4 revealed interesting findings. Disease severity was significantly greater in the anti-PD-1 than anti-CTLA-4 group (Fig. 1A, p = 0.01), featuring a more pronounced and disruptive mononuclear cell infiltration (Fig. 1D, E). Comparison of thyroiditis severity in mice developing the disease treated with anti-PD-1 versus controls in the iodine-supplemented cohort confirmed a substantial significance (p = 0.07). Only mice receiving anti-PD-1, both in mono- and combination therapy, developed severe (severity score ≥3) thyroiditis (Fig. 1A).
Incidence, on the contrary, was greater in the anti-CTLA-4 group, where all mice (100%) developed thyroiditis, than the anti-PD-1 group, where five of eight mice (63%) did. Mice injected with isotype control developed thyroiditis at incidence similar to that of the anti-PD-1 group (66%) if supplemented with iodine, whereas none of the mice drinking regular water developed thyroiditis. In the combination therapy groups, 63% of mice drinking regular water and 70% of mice drinking iodinated water developed thyroiditis.
Thyroid ultrasonography provided additional insights on the pathogenesis of thyroiditis. Thyroid area was markedly increased in mice injected with the two checkpoint inhibitors (12.5 ± 3.6 mm2 in those drinking iodinated water and 11.00 ± 3.1 mm2 in those on regular water) than in those injected with isotype controls (8.4 ± 0.8 mm2 in those drinking iodinated water and 6.9 ± 0.1 mm2 in those on regular water, Fig. 2A). When comparing time of sacrifice with baseline, the thyroid area significantly increased (about 50%) upon injection of anti-PD-1 and anti-CTLA-4, independently of the presence of iodine, an increase that was instead not seen in the isotype controls (p = 0.06 when comparing mice drinking iodinated water and p = 0.01 when comparing mice drinking regular water, Fig. 2A).

Morphological comparison of ICI-treated vs. untreated mice. Thyroid frontal area (sum of width × depth × Π for each lobe) fold change compared with baseline in (i) mice treated with both ICIs and drinking either iodinated water (Combo NaI) or regular water (Combo RW); (ii) mice treated with one ICI and drinking iodinated water (αPD-1 NaI and α-CTLA-4 NaI); (iii) mice injected with the isotype control monoclonal antibody and drinking either iodinated water (Control NaI) or regular water (Control RW). p-Value based on Wilcoxon rank-sum test (
Interestingly, monitoring thyroid areas during disease development showed a clear goiter, with about twofold increase at the time of sacrifice over baseline in the anti-CTLA-4 group (17 ± 8.2 mm2, Fig. 2A, E), an increase that instead was not seen in controls (p = 0.02). In the anti-PD-1 group, this increase was less marked (11 mm2 ± 4.2), so that no significant increase in the thyroid area was seen when compared with the isotype control group (p = 0.3). Differently from the severe thyroiditis typical of anti-PD-1-treated mice, mice receiving anti-CTLA-4, both in mono- and combination therapy, developed goiter (Fig. 2A–E).
Hypothyroidism is more severe upon PD-1 blockade than CTLA-4 blockade
As thyroiditis unfolded in mice supplemented with iodine, serum total T4 began to decline in all groups, reaching statistically lower values than baseline one month after treatment initiation (142 days of age). As disease progressed, T4 continued to decline in the anti-PD-1 group, reaching 50% lower values than baseline on day 172, whereas it began to recover in the anti-CTLA-4 and isotype control groups (Fig. 3A, p = 0.02). TSH followed the T4 changes accordingly, by increasing in the anti-PD-1 and anti-CTLA-4 groups during the development of thyroiditis, with no difference between the two inhibitors (Fig. 3B).

Thyroid function and autoimmunity in thyroiditis mice treated with PD-1 or CTLA-4 blockade. Serum T4-fold change compared with baseline in mice supplemented with iodine that developed thyroiditis, treated with PD-1 blockade (closed circles), CTLA-4 blockade (closed diamonds), or isotype control (open diamonds); dashed line represents the line of no difference, that is the value of T4 level observed at baseline; p based on Wilcoxon rank-sum test (
Thyroglobulin antibodies increased in all mice drinking iodinated water. Interestingly, the increase was more pronounced in the anti-PD-1 group where they reached the highest levels on day 172 (Fig. 3C, closed circles). A similar behavior was observed for thyroperoxidase antibodies that significantly increased over baseline in the checkpoint inhibitor groups (Fig. 3D, p < 0.05 for both inhibitors), although with no difference between the two inhibitors.
Distinct lymphoid subsets are found in the thyroid glands of mice treated with anti-CTLA-4 or anti-PD-1
Analysis of hematopoietic infiltration of one thyroid lobe by flow cytometry showed an excellent positive correlation with the histopathological score assigned to the contralateral lobe (Fig. 4A).
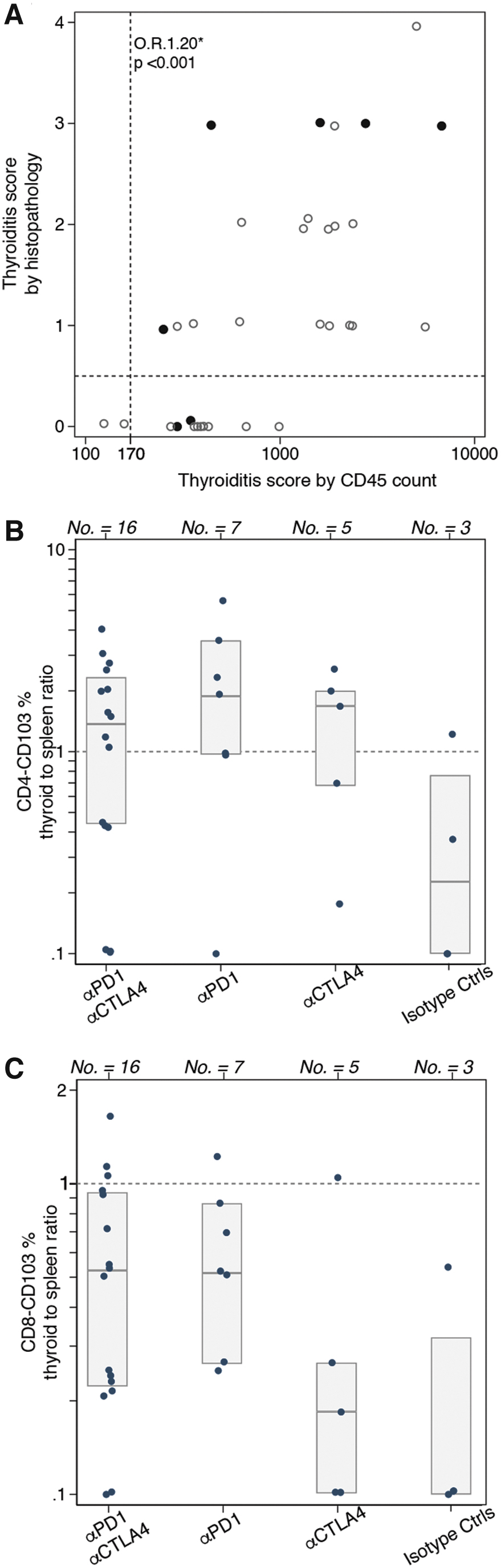
Lymphoid subsets in thyroid gland and comparison with spleen levels as assessed by flow cytometry. Infiltration of CD45-positive cells (count) in one thyroid lobe related to the histological thyroiditis score on the contralateral lobe. *OR and p based on ordered logistic regression for the odds of a higher vs. lower histopathology severity every 170-unit increase in CD45 infiltrating cells; the dashed horizontal line represents the distinction between thyroiditis or no thyroiditis based on histology, while the dashed vertical line represents the same distinction based on a cutoff value (170) of CD45-positive cells infiltrating the thyroid gland (
Using 170 hematopoietic cells per lobe as the cutoff between normal and infiltrated thyroid, an ordered logistic regression model relating the absolute number of CD45+ cells in one lobe to the five levels of the histopathological score yielded a cumulative odds ratio (OR) of 1.20 (95% confidence interval [CI] from 1.1 to 1.3, p < 0.001), that is a 20% greater odds of a higher versus lower severity score for every 170-unit increase in CD45 infiltrating cells. This cutoff proved more sensitive than histopathology since classified as thyroiditis mice that were instead scored normal by microscopy (Fig. 4A, lower right quadrant). It is important, however, to recall that autoimmune pathologies often affect the target organ focally, and thus, it is possible that one thyroid lobe was affected more severely than the other.
Mice treated with anti-PD-1 developed the highest infiltration also when assessed by flow cytometry (Fig. 4A, closed circles).
Analysis of tissue resident memory T cells (TRMs), labeled by the surface expression of CD103 and/or CD69 in the CD45+-CD4+ gate, showed a significant increase of CD103 expression in mice treated with ICIs compared with controls (Table 1, p = 0.045). This subpopulation, in fact, increased from an average of 0.9% in controls to about 3% in cases. In the CD45+-CD8+ gate, CD103+ cells also tended to increase in cases compared with controls (Table 1, p = 0.055), markedly in the anti-PD-1 or the combination groups. The median percentage of this lymphoid subset was 2% in isotype controls, 5% in anti-CTLA-4, 8% in anti-PD-1, and 7% in the combination group.
Comparison of Key Lymphoid Subsets in the Thyroid Glands (One Lobe) of Mice Treated with Anti-PD-1, Anti-CTLA-4, Both Antibodies, or Isotype Control
CD4, CD8, and CD19 values are percent of the CD45 gate.
CTLA-4, cytotoxic T lymphocyte antigen-4; PD-1, programmed death receptor-1.
Comparison of resident memory T cell variation in the thyroid and spleen revealed that CD45+-CD4+ cells expressing CD103 in treated mice are higher in thyroid tissue compared with spleen (Fig. 4B, ratio >1), whereas CD45+-CD8+ cells expressing CD103 are lower in thyroid tissue (Fig. 4C, ratio <1), enlightening a potential local activation for the former subset and a systemic activation for the latter.
Comparing mice treated with ICIs developing thyroiditis versus mice treated with ICIs that did not develop thyroiditis, we identified in the CD45+-CD4+ gate a significantly higher expression of single positive CD103 (4% in thyroiditis vs. 1% in nonthyroiditis group; Table 2, p = 0.019), of single positive CD69 (30% in thyroiditis vs. 20% in nonthyroiditis group; Table 2, p = 0.041), and double positive CD69-CD103 (6% in thyroiditis vs. 2% in nonthyroiditis group; Table 2, p = 0.002). On the contrary, CD103 and/or CD69 expression in the CD45+-CD8+ gate was similar between ICI-treated mice, independently of thyroiditis development (Table 2, all p > 0.05).
Comparison of TRM Expression in Mice Treated with Immune-Checkpoint Inhibitors Developing Versus Nondeveloping Thyroiditis
CD4, CD8, and CD19 values are percent of the CD45 gate.
ICI, immune-checkpoint inhibitor; TRM, tissue resident memory T cells.
Gating strategy is represented in Supplementary Figure S2.
PD-1 and CTLA-4 blockade is associated with distinct cytokine profiles in the peripheral blood
Administration of the PD-1 blocking antibody caused a fivefold increase in the circulating levels of IL-6, from an average of 40 pg/mL at baseline to 198 pg/mL on day 172, levels that remained instead normal in most mice treated with the CTLA-4 blocking antibody or the isotype control (Fig. 5A, B, p = 0.01). IL-6 mirrored the thyroiditis severity score, with the highest serum values found in mice with the most severe thyroid histopathology (Fig. 5C). On the contrary, serum levels of GM-CSF (Fig. 5D) and MIP1β (Fig. 5E) increased more in the anti-CTLA treatment group, reaching statistically significant higher values on day 172 (p < 0.001 for both).

Cytokine profile in mice treated with ICIs and isotype controls. Serum IL-6 (pg/mL) at sacrifice time in mice treated with combination therapy, anti-PD-1, anti-CTLA-4, or isotype controls, p based on Wilcoxon rank-sum test (
The other six cytokines, IL-12, TNF-α, IFN-γ, MCP-1, IL-2, and Rantes, increased similarly in mice treated with the ICIs when compared with isotype controls, but with no difference according to the type of inhibitor (Supplementary Fig. S3). Of them, IL-12 showed the most pronounced increase, going from an average of 8 pg/mL at baseline in all groups to an average of 994 pg/mL in the anti-PD-1 and 1268 in the anti-CTLA-4 groups (Fig. 5F, p = 0.01 vs. isotype controls).
In addition, IL-12 and IL-6 were the cytokines that remained significantly to thyroiditis severity in a multiple ordered logistic regression model that accounted for the other cytokines (OR 1.01 [CI 1–1.02], p = 0.03 for IL-12, and OR = 1.1 [CI from 1.02 to 1.15], p = 0.009 for IL-6, that is 1% and 10% greater odds of a higher vs. lower severity score for every unit increase in IL-12 and IL-6, respectively).
Discussion
ICI therapy triggered thyroiditis in this mouse model with incidence and severity that varied greatly according to the ICI used: thyroiditis was more severe but less frequent after PD-1 blockade and less severe but universal after CTLA-4 blockade. These findings are consistent with the different modes of action of the two checkpoint inhibitors. CTLA-4 acts in secondary lymphoid organs at the early phase of the immune response, and thus, its blockade is predicted to have a broader impact; on the contrary, PD-1 acts in peripheral tissues within an already established immune response, that is, on mice with high predisposition to thyroiditis, yielding a restricted, although more intense, response, causing overt thyroid dysfunction.
Our findings could also explain the discrepancy of why PD-1 blockade thyroid adverse events seem more frequent than the CTLA-4 blockade one (2,9): PD1-blockade triggers a more intense and prolonged effect, finally impairing thyroid function, thus being detectable at functional screening, whereas CTLA-4 blockade may be more involved with alterations in thyroid morphology, not usually screened in trials.
Ultrasound represents the mainstay for diagnostic imaging of the thyroid gland, but this kind of report in thyroid dysfunction secondary to ICIs is rarely described (10,11). In our murine cohort, ICI treatment almost doubled the thyroid area, with a greater effect in mice treated with anti-CTLA-4. Our results support the utility of performing thyroid ultrasound in patients who develop dysthyroidism after ICIs, and could be even useful for screening, since it is a safe, radiation-free, and inexpensive technique. In agreement with this recommendation, a recent study suggested that the thyroid echo pattern at baseline may predict the risk of thyroid dysfunction induced by anti-PD-1 (12).
Our study revealed a marked increase of effector CD4 T cells in the thyroid gland of cases injected with ICIs than controls, consistent with the notion that this population of T cells is the main target of ICIs. The increase was also present at a systemic level, since ICIs also expanded the number of splenic CD4 effector T cells (data not shown), confirming a previous finding (13). CD8 T lymphocytes, on the contrary, did not significantly vary in the subgroups.
Within the T cell compartment, an interesting subset is the TRMs, a specialized memory population that permanently resides in tissues at the epithelial barrier level. Those cells mediate a rapid response to pathogens, independently to the recruitment of T cells from the blood, but an overactive or aberrant response of these cells can cause some human autoinflammatory conditions, such as psoriasis and mycosis fungoides (14). The main cell surface markers that have been associated with both CD4 and CD8 TRM in mice and human tissues are CD69 and CD103. TRMs have been described in many organs such as skin, lung, brain, kidney, and the genitourinary system; as far as we know, this peculiar subclass has never been described in thyroid.
Previous studies discovered that the immune-checkpoint receptors such as CTLA-4 and PD-1 are upregulated in TRMs (15,16); moreover, CD103 TRMs are also present as a subclass of tumor infiltrating lymphocytes in cancer, and even in this setting, a higher expression of immune-checkpoints has been demonstrated (17); in this light, it has been hypothesized that this lymphoid population may be specifically involved in the outcomes of ICI therapies. We further refined this hypothesis by proposing that removal of the immune brake by ICIs expands TRMs and recruits them to mediating thyroid immune-related adverse events. Indeed, ICI treatment significantly increased CD4 expressing CD103 in all treated mice, and CD8 expressing CD103 in PD-1-treated ones. No significant changes in B lymphocytes were found in our study.
Serum levels of GM-CSF and MIP1-β increased after CTLA-4 blockade, while IL-12 after the combined treatment. Nevertheless, the only two cytokines that significantly associated with thyroiditis severity were IL-12 and IL-6, the first increased in all mice treated with checkpoint inhibitors and IL-6 only in those treated with the PD-1 blocking antibody.
IL-12 is produced by activated antigen-presenting cells as they interact with naive T cells, promoting their differentiation into a Th1 phenotype (18). IL-12 is produced in the thyroid gland during the initiation and maintenance phases of lymphocytic thyroiditis (19). In a mouse model, thyroiditis is milder when IL-12 is absent or blocked with specific antibodies (20). In a small cohort of Chinese thyroid patients, IL-12 was increased in those with Graves and Hashimoto diseases (21).
IL-6 is a multifaceted proinflammatory cytokine. It promotes naive T CD4 differentiation to a Th17 profile (22), reduces T-reg differentiation (23), promotes T CD8 cytotoxic activity, and increases antibody production (24,25). Deranged IL-6 levels are found in a variety of diseases, from autoimmunity to viral infections to cancer. Other studies have detected higher levels of IL-6 after PD-1/PD-L1 blockade in patients developing immune-related adverse events such as psoriasiform dermatitis (26,27) and in pneumonitis (28), which were indeed successfully treated with anti-IL-6 therapy. In thyroid autoimmune diseases, IL-6 has been associated with destructive thyroiditis (type 2 amiodarone induced or subacute thyroiditis) (29), a disease that greatly responds to steroid therapy.
In our study, IL-6 increased only in mice treated with the PD-1 blocking antibody, an increase tightly associated with severity of thyroiditis, confirming the presence of a cross talk between the IL-6 and PD-1 pathway. A possible pathogenetic hypothesis is that PD-1 blockade increases IL-6 levels, which in turn, in concert with other cytokines, particularly IL-12, induce thyroid inflammation and loss of functionality. In this light, IL-6 could be used as a potential biomarker. An early treatment with corticosteroid could possibly block this pathogenetic mechanism, thus avoiding long-term dysthyroidism and goiter formation.
Conclusions
ICIs trigger thyroiditis in different ways: CTLA-4 blockade produces a cluster of cytokines having the ability to increase thyroid volume but with a mild effect on thyroid function; conversely, PD-1 blockade increases IL-6, which determines in susceptible subjects a severe inflammatory infiltration responsible for decreased thyroid function.
Acknowledgments
The authors gratefully acknowledge: (1) Paulina Chalan (BS, PhD, Division of Immunology, Department of Pathology, the Johns Hopkins School of Medicine, Baltimore, MD, USA) for her precious help in conceiving experiments on TRMs and assistance with flow cytometry. (2) Kathleen Gabrielson (DVM, PhD, Department of Molecular and Comparative Pathobiology, the Johns Hopkins School of Medicine, Baltimore, MD, USA) for assistance with mice ultrasonography.
Authors' Contributions
P.C., S.I., and G.D.D. conceived the experimental project. S.I., G.D.D., F.P., and E.S. carried out the experiments. P.C. and S.I. performed the statistical analyses, designed the figures, and drafted the article. All authors discussed the results and contributed to the final article.
Abstract Presentation
The work was presented as Abstract at the 89th Annual Meeting of the American Thyroid Association on October 31, 2019, and first author Silvia Ippolito was awarded with the “Women Advancing Thyroid Research Award.”
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Institutes of Health grant RO1 CA-194042 to P.C.
Supplementary Material
Supplementary Data
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3