
Abstract
Background:
Adipogenesis, glycosaminoglycan hyaluronan (HA) production, inflammation, and fibrosis are the main pathogenic mechanisms responsible for Graves' orbitopathy (GO). We hypothesized that disulfiram (DSF), an aldehyde dehydrogenase (ALDH) inhibitor used to treat alcoholism, would have therapeutic effects on orbital fibroblasts (OFs) in GO. This study aimed at determining the therapeutic effects and underlying mechanisms of DSF on these parameters.
Methods:
Primary cultures of OFs from six GO patients and six control subjects were established. The OFs were allowed to differentiate into adipocytes and treated with various concentrations of DSF. Lipid accumulation within the cells was evaluated by Oil Red O staining. Real-time polymerase chain reaction (RT-PCR) and Western blotting were used to measure the expression of key adipogenic transcription factors, ALDH1A1, ALDH2, and mitogen-activated protein kinase (MAPK) signaling proteins. Apoptosis assays and reactive oxygen species levels were evaluated by flow cytometry. HA production was measured by using an enzyme-linked immunosorbent assay (ELISA) kit. The mRNA levels of proinflammatory molecules were measured by using RT-PCR after interleukin (IL)-1β stimulation with or without DSF. The mRNA expression of markers associated with fibrosis was examined by using RT-PCR after transforming growth factor (TGF)-β1 stimulation with or without DSF. The wound-healing assay was assessed by phase-contrast microscopy.
Results:
Under identical adipogenesis conditions, GO OFs effectively differentiated, while normal control (NC) OFs did not. DSF dose dependently suppressed lipid accumulation during adipogenesis in GO OFs. The expression of key adipogenic transcription factors, such as perilipin-1 (PLIN1), PPARγ (PPARG), FABP4, and c/EBPα (CEBPA), was downregulated. Further, DSF inhibited the phosphorylation of ERK by inhibiting ALDH1A1. In addition, DSF attenuated HA production and suppressed inflammatory molecule expression induced by IL-1β in GO OFs and NC OFs. The antifibrotic effects of DSF on TGF-β1 were also observed in GO OFs.
Conclusions:
In the current study, we provide evidence of the inhibitory effect of DSF on GO OFs adipogenesis, HA production, inflammation, and fibrosis in vitro. The results of this study are noteworthy and indicate the potential use of DSF as a therapeutic agent for the treatment of GO.
Introduction
Graves' orbitopathy (GO) is a complex organ-specific autoimmune inflammatory manifestation of Graves' hyperthyroidism, which accounts for ∼80% of thyroid-associated ophthalmopathy (1,2). Approximately 3–5% of GO cases are severe, accompanied by severe eye pain, inflammation, corneal ulcers, or compressive optic neuropathy (3). The main clinical manifestations are exophthalmos, increased orbital pressure, and vision loss caused by the expansion of orbital fatty connective tissues and extraocular muscles (4). This vexing and undertreated ocular disease dramatically affects the quality of life.
The pathogenesis of GO is not completely clear. Studies have shown that GO is caused by imbalanced immune tolerance that is affected by antigen stimulation, the environment, heredity, and other factors, which results in autoimmune T cells (Treg cells, CD8+ cytotoxic T cells, CD4+ helper T [Th] cells, Th1/Th2/Th17 cells) (5,6) and B cell activation (7,8).
Orbital fibroblasts (OFs) are essential GO target cells and effector cells. The OFs can interact with lymphocytes, mononuclear cells, macrophages, and mast cells in orbit (9). They secrete various chemokines and cytokines [interleukin (IL)-1β, IL-2, IL-6, CXCL8, IL-10, IL-17A, COX2, CCL2, CCL5, IFN-γ, TNF-α, transforming growth factor (TGF)-β, etc. (10,11)], and they strengthen the inflammatory and autoimmune response. Thyroid-stimulating hormone receptor (TSHR) was first reported in orbital tissues from individuals with GO by Feliciello et al. (12). Active GO patients had much higher TSHR expression than inactive patients (13). Further, OFs express both TSHR and insulin-like growth factor 1 receptors (IGF-1R) (14). IGF-1R is overexpressed in OFs and lymphocytes in patients with GO (15). IGF-1R forms a functional complex with TSHR and mediates the TSHR downstream signaling (16 –20). In summary, IGF-1R and TSHR can be considered as “partners in crime.”
Clinically, the first-line treatment for active, moderate-to-severe, sight-threatening GO is high-dose intravenous glucocorticoid (GC) therapy (21). However, there are some GC-dependent or GC-resistant patients. Further, systemic application of GCs will produce specific side effects and complications. For inactive or fibrotic GO, surgery (orbital decompression, strabismus correction, etc.) is mainly used to alleviate symptoms, but there is no cure. Even surgical contraindications and complications are thorny problems. Immunosuppressive therapies and orbital irradiation also have limitations. Recently, novel drugs have been gradually developed for the molecular mechanism of pathogenesis, such as teprotumumab (anti-IGF-1R monoclonal antibody) (22,23), TSHR-specific allosteric small-molecule antagonists (24), tocilizumab (anti-IL-6 receptor monoclonal antibody) (25), etanercept (TNF-alpha receptor blocker) (26), and adalimumab (monoclonal antibody against TNF-α) (27).
Overall, the need for new treatments for GO has not been met. It is necessary to deeply study the pathogenesis of GO and develop new targeted therapeutic drugs. To provide additional insight into the pathogenesis of GO, the contribution of specific factors and their interaction with other mediators in adipogenesis and fibrosis requires further evaluation.
This study investigated the therapeutic effects of disulfiram (DSF) on an in vitro GO model. As an aldehyde dehydrogenase (ALDH) inhibitor, the Food and Drug Administration (FDA) approved DSF to treat alcohol abuse in 1951. The good pharmacokinetics, safety, and tolerance of DSF have been widely recognized in the clinic (28). Currently, increasing evidence points to new prospects for old drugs. DSF can reverse diet-induced obesity and metabolic dysfunction, prevent weight gain, eliminate the adverse effects of an obese diet on the insulin response, and reduce liver steatosis and islet hypertrophy (29,30). By regulating lipid metabolism and oxidative stress, DSF significantly reduced liver inflammation, inhibited inflammation-related genes, inhibited macrophage infiltration, reduced hepatic steatosis, and considerably reduced hepatic fibrosis (31). DSF suppresses NLRP3 inflammasome activation to treat peritoneal and gouty inflammation (32).
Based on these mechanisms, we hypothesize that DSF has therapeutic effects on OFs and promising potential for further development as a GO treatment. Here, we first report the antiadipogenic effects of DSF on GO and explore some mechanisms. In addition, we indicate that DSF can attenuate hyaluronan (HA) production, suppress inflammatory molecule expression, and exert antifibrotic effects.
Materials and Methods
Primary cultures of OFs
Orbital adipose/connective tissue samples were collected from six patients with GO who underwent orbital decompression. Further, all cases were euthyroid during the operation. Normal control (NC) orbital/connective tissue samples were collected from six patients without GO who underwent enucleation. Clinical patient information is provided in Table 1. The severity and clinical activity of GO were graded according to the NOSPECS classification (0 = no symptoms or signs; I = only signs, no symptoms; II = soft tissue involvement; III = proptosis; IV = extraocular muscle involvement; V = corneal involvement; VI = sight loss, due to optic nerve involvement) and the 7-item clinical activity score (CAS) scheme proposed by EUGOGO, respectively (21,33). All patients signed informed consent forms. This study was conducted according to the Declaration of Helsinki and approved by the Institutional Review Board of Zhongshan Ophthalmic Center (2016KYPJ028).
Clinical Features of Patient Samples Used in this Study
CAS, clinical activity score; F, female; GCs, glucocorticoids; GO, Graves' orbitopathy; M, male; N, no; R/L, right or left eyes; Y, yes.
Tissue explants were cut into small pieces and plated in 10-cm culture dishes in high-glucose Dulbecco's modified Eagle's medium (DMEM) containing 20% fetal bovine serum (FBS) and 1% penicillin/streptomycin (all from Gibco Laboratories, New York). After the cells had migrated out of the tissue pieces and reached confluence, the cells were passaged with 0.25% trypsin/EDTA (Gibco Laboratories). Then, the cells were grown in proliferation medium (PM) (DMEM containing 10% FBS and 1% penicillin/streptomycin) by using standard cell culture protocols. For experiments, OFs were between passages 3–7, and each experiment was repeated for at least three independent specimens.
Cell viability assays
Cell viability was assessed by using a Cell Counting Kit-8 (CCK-8) assay. Briefly, cells were treated with different concentrations of DSF (Sigma-Aldrich, Shanghai, China) and NCT-501 (Absin, Shanghai, China) for the indicated times in 96-well plates. Measurements were similar to those of the previous study (34).
Adipogenesis and Oil Red O staining
Cells were plated in six-well plates or 6-cm culture dishes and allowed to reach confluence. To induce adipogenic differentiation, PM was changed to commercial adipogenic differentiation medium (DM; SALIAI, Guangzhou, China) with or without DSF (0.5, 1, 2, and 4 μM). The medium was replaced with fresh DM every two to three days according to the instructions. Three days later, OFs were lysed, and adipogenic marker mRNA expression was analyzed based on the time-dependent preliminary assay (data not shown). Ten days later, differentiated OFs were stained with Oil Red O or lysed for subsequent Western blotting. Oil Red O staining and relative quantification were performed as previously described (34).
RNA extraction and real-time polymerase chain reaction
According to the manufacturer's protocol, total RNA was extracted by using a TaKaRa MiniBEST Universal RNA Extraction Kit (TaKaRa, Dalian, China). cDNA was synthesized from total RNA by using PrimeScript RT Master Mix (TaKaRa). Real-time polymerase chain reaction (RT-PCR) was performed on a Roche LightCycler 480 (Roche, Basel, Switzerland) by using TB Green Premix Ex Taq II (TaKaRa). The primer pair sequences for RT-PCR are listed in Supplementary Table S1. GAPDH was used as the housekeeping gene.
Automated Western blotting
A protein extraction kit (KeyGEN, Nanjing, China) was used to lyse the cells. Protein concentrations were measured with a BCA kit (Beyotime, Shanghai, China). Protein separation and detection were performed by using an automated capillary electrophoresis system (Simple Western system and Compass software; ProteinSimple). Wes Separation Capillary Cartridges for 12–230 kDa were used for the proteins. The following primary antibodies were used: GAPDH, PPAR-γ, FABP4, c/EBP-α, c/EBP-β, perilipin-1 (PLIN1), ALDH1A1, ALDH2, ERK1/2, phospho-ERK (p-ERK), p38, and JNK (all from Cell Signaling Technology, Boston, MA). Signals were detected with an HRP-conjugated anti-rabbit secondary antibody and visualized by using Compass software.
Immunofluorescence analysis
Immunofluorescence (IF) staining of tissues was performed as previously described (35). The primary antibody was ALDH1A1 (1:500; Cell Signaling Technology), while the secondary antibody was anti-rabbit Alexa Fluor Plus 488 (1:500; Thermo Fisher Scientific, Rockford).
Apoptosis assays and reactive oxygen species measurement
Cells that were 80% confluent were treated with DSF (0, 0.1, 0.5, 1, 2, 4, and 5 μM) for 72 hours. Annexin V (AV)-propidium iodide (PI) staining was performed by using an AV-PI apoptosis detection kit (Vazyme, Nanjing, China) according to the product instructions. Reactive oxygen species (ROS) generation was determined by using the fluorescent probe 2′,7′-dichlorodihydrofluorescein (Beyotime, Shanghai, China). Stained cells were analyzed by a flow cytometer (BD LSRFortessa, Franklin Lakes, NJ), as previously described (34).
HA measurement
Cells were treated with different concentrations of DSF (0, 0.5, 1, 2, or 4 μM) and 1 ng/mL IL-1β (R&D Systems, Minneapolis, MN) in DMEM supplemented with 1% FBS. After 24 hours, the cell culture media were collected and centrifuged at 5000 g for 10 minutes. According to the manufacturer's instructions, the concentrations of HA in the cell culture supernatant were quantified with an enzyme-linked immunosorbent assay (ELISA) kit (R&D Systems).
Wound-healing assay
Confluent cell monolayers in a six-well plate were wounded by mechanical scraping; a straight scratch was made with a pipette tip to simulate a wound. Wound width was assessed at the time of scraping to ensure that all wounds had identical widths at the beginning of the experiment. The cell culture medium was subsequently replaced with fresh medium supplemented with DSF (4 μM) and 10 ng/mL TGF-β1 and incubated for 24, 48, and 72 hours. Wound closure was observed by phase-contrast microscopy (40 × ; Nikon, Tokyo, Japan) and photographed. The results are expressed as the wound width.
Statistical analysis
All experiments were performed at least three times with samples from different individuals, and samples were assayed in duplicate each time. The data are expressed as the mean values and standard deviations. All calculations and statistical analyses were performed by using GraphPad Prism v9 (GraphPad Software, Inc., La Jolla, CA). Statistical analyses were performed with one-way analysis of variance or Student's t-tests. A p-value <0.05 was considered to indicate statistical significance.
Results
DSF exerts antiadipogenic effects on GO OFs
To determine the noncytotoxic DSF concentrations, CCK-8 assays were performed on OFs. The results show that DSF at concentrations <5 μM was safe for GO and NC OFs (Supplementary Fig. S1A). Therefore, we chose 0.5, 1, 2, and 4 μM DSF for subsequent experiments. Cultured OFs from GO and NC patients were allowed to differentiate into adipocytes for 10 days. Oil Red O staining shows that adipogenesis was significantly stimulated by DM induction in GO OFs.
Interestingly, we found that NC OFs did not effectively differentiate even with the same stimulation, which is similar to the results of a previous study (36). As shown in Figures 1A and B, DSF inhibited cellular lipid droplet formation in a dose-dependent manner. Compared with those in the DM group, the number and size of lipid droplets in the DSF group significantly decreased.
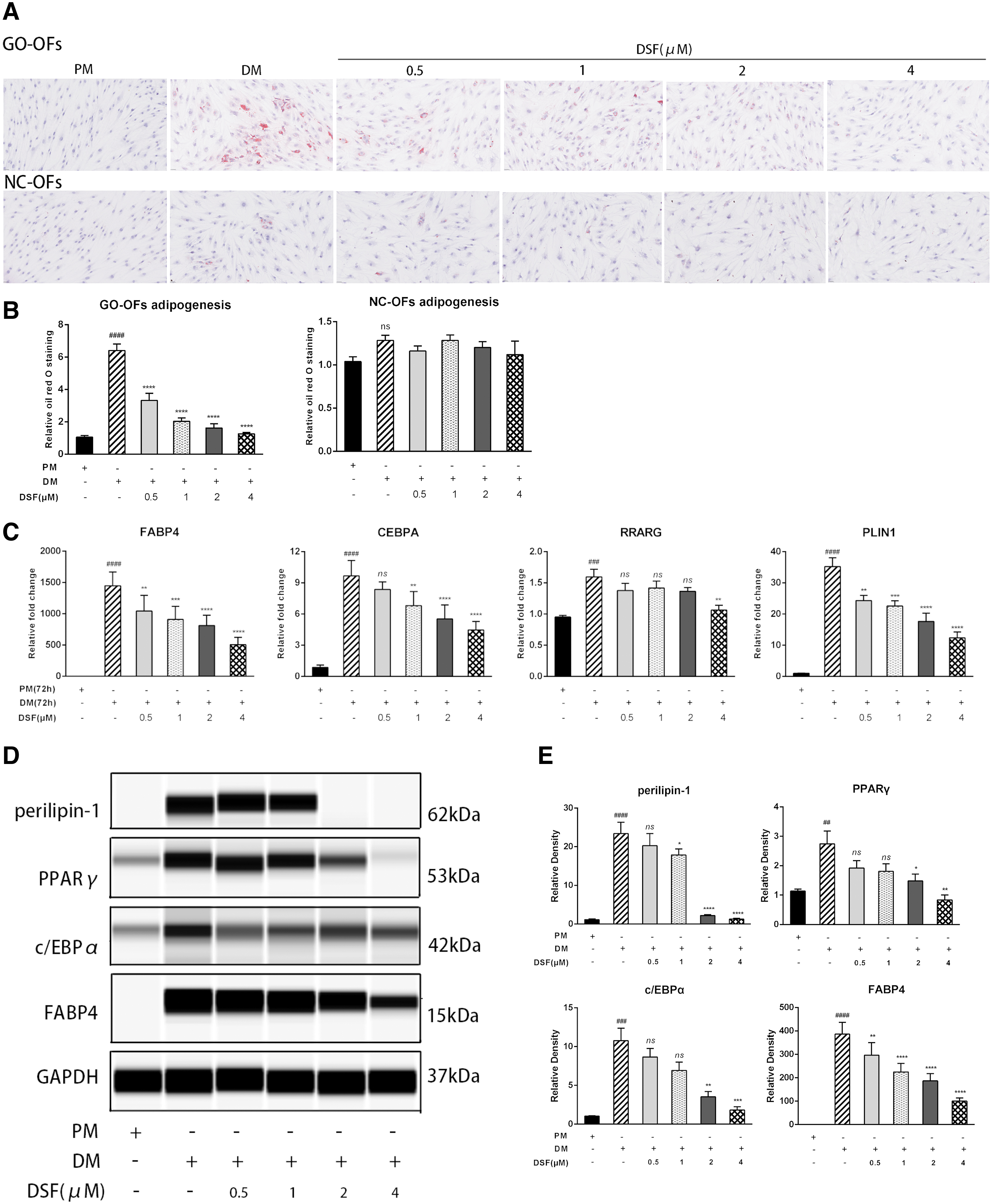
DSF exerts antiadipogenic effects on GO OFs. (
Adipocyte differentiation requires the sequential and cooperative expression of key adipogenic transcription factors, such as PLIN1, PPARγ (PPARG), FABP4, c/EBPα (CEBPA), and c/EBPβ (CEBPB) (37). Further, we performed real-time polymerase chain reaction (RT-PCR) and Western blotting to analyze adipogenic markers. DSF dampened the expression of PLIN1, PPARγ, c/EBPα, and FABP4 at both mRNA and protein levels (Fig. 1C–E), whereas DM did not induce c/EBPβ expression (data not shown).
DSF regulates the ALDH1A1/ERK pathway during adipogenesis
ALDH1A1 and ALDH2 are mitochondrial enzymes whose dysregulation has been associated with many human pathologies, including metabolic disorders and cancer (38 –40). Because the metabolic products of DSF can inhibit ALDH activity in vivo, we investigated whether ALDH1A1 or ALDH2 expression changed during GO OFs adipogenesis by Western blotting. The level of ALDH1A1 was downregulated, while that of ALDH2 did not change (Fig. 2A, B). Moreover, mitogen-activated protein kinase (MAPK)/ERK signaling during the early stage of adipogenesis is responsible for enhancing adipogenic marker expression in preadipocytes (41). We found that ERK was phosphorylated when adipogenesis was stimulated with DM, and DSF dose dependently inhibited p-ERK/ERK levels (Fig. 2A, B). We also examined other MAPK pathways, but there was no effect on p38, and JNK was barely detected (data not shown).
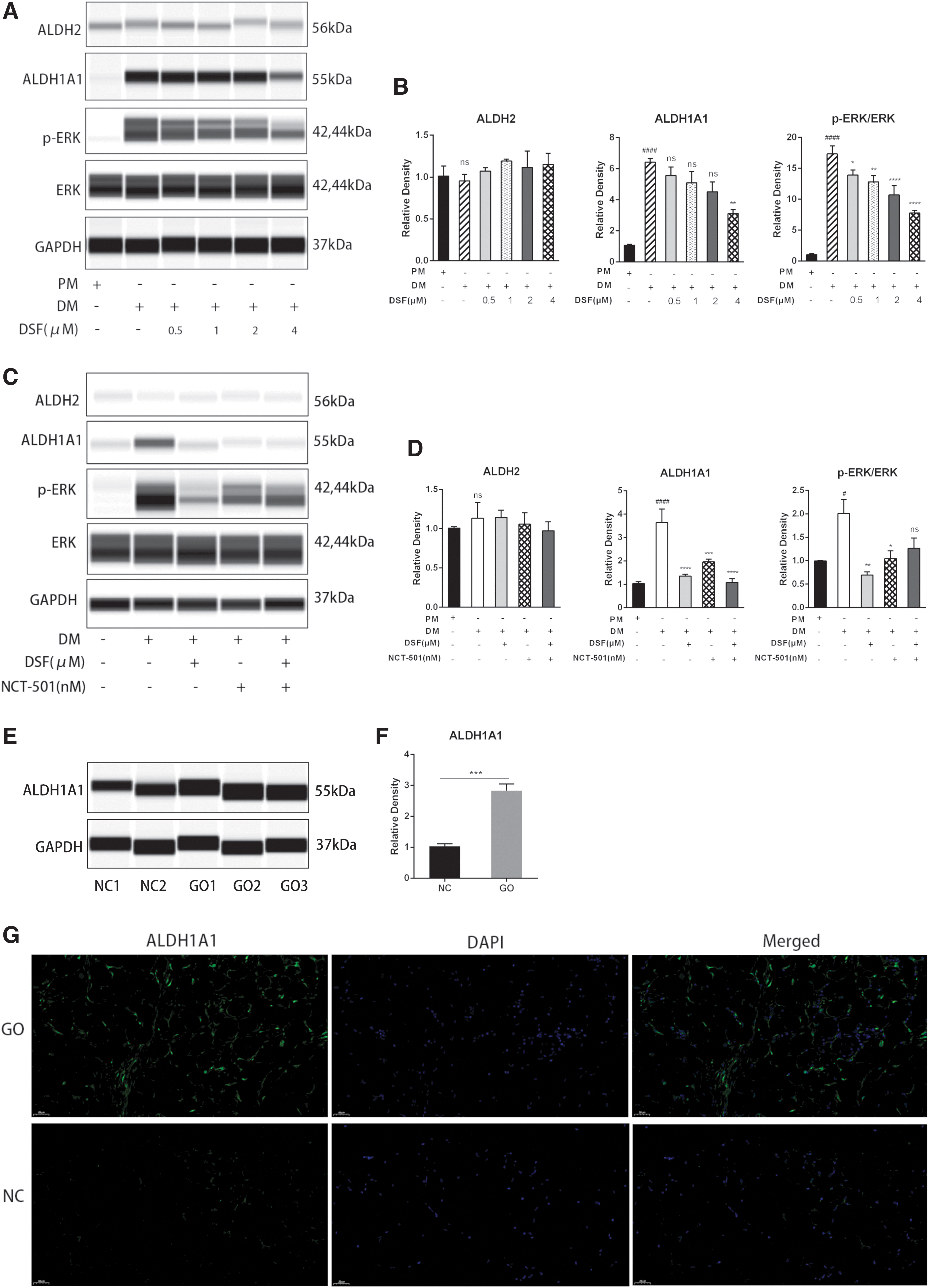
DSF regulates the ALDH1A1/ERK pathway during adipogenesis. (
Further, to confirm that ALDH1A1 and ERK inactivation was responsible for the antiadipogenic effects of DSF, the signaling pathways were examined by using the ALDH1A1 inhibitor NCT-501. CCK-8 assays confirmed the noncytotoxic concentration of NCT-501 in OFs (Supplementary Fig. S1B). Interestingly, NCT-501 blocked the ALDH1A1 and slightly decreased the p-ERK/ERK level (Fig. 2C, D). We examined the effects of NCT-501 on GO OFs. The results show that NCT-501 has a slight antiadipogenic effect but much less than DSF (Supplementary Fig. S2A–D). DSF and NCT-501 were co-administered to evaluate whether ERK phosphorylation was suppressed when ALDH1A1 activation was blocked. Pretreatment with NCT-501 did not show the suppressive effect of DSF on ERK phosphorylation, which suggests that DSF partially functions by inhibiting ALDH1A1. In addition, we examined whether ALDH1A1 expression was different in orbital adipose/connective tissue samples from patients with GO and NC. The results show that ALDH1A1 levels in patients with GO were much higher than those in patients with NC (Fig. 2E, F). Tissue IF analysis also showed that ALDH1A1 expression was significantly higher in the GO group than in the NC group (Fig. 2G).
DSF affects neither apoptosis nor ROS production
DSF/copper has been reported to induce significant cell cycle arrest and apoptosis, cause ROS imbalance, and activate the stress-related ROS-JNK pathway (42). Therefore, we examined apoptosis and ROS levels in DSF-treated OFs. The results show that the noncytotoxic concentration of DSF did not induce cell apoptosis or ROS generation (Supplementary Fig. S3A, B, C). Interestingly, GO OFs expressed higher ROS levels than NC OFs at baseline (Supplementary Fig. S3D, E), consistent with previous research (43).
DSF attenuates HA production and suppresses inflammatory molecule expression
HA is a type of glycosaminoglycan that plays an essential role in the extracellular matrix (44). We investigated whether DSF inhibited HA production in primary cultures of OFs via ELISA. As shown in Figure 3A, IL-1β increased HA production in GO OFs and NC OFs compared with untreated control cells. In addition, cotreatment with DSF decreased HA production in a dose-dependent manner. The changes in GO OFs were more significant than those in NC OFs. Changes in the mRNA and core genes associated with HA (HAS1, HAS2, HAS3) also confirmed these changes (Fig. 3B, C). However, the mRNA expression levels of HYAL1 were not affected by IL-1β or DSF. Further, DSF significantly suppressed the IL-1β-induced mRNA expression of inflammatory molecules, including IL-6, CXCL8, COX-2, CCL2, and CCL5 (Fig. 3D, E). Similarly, GO OFs reacted more obviously than NC OFs in terms of inflammation.
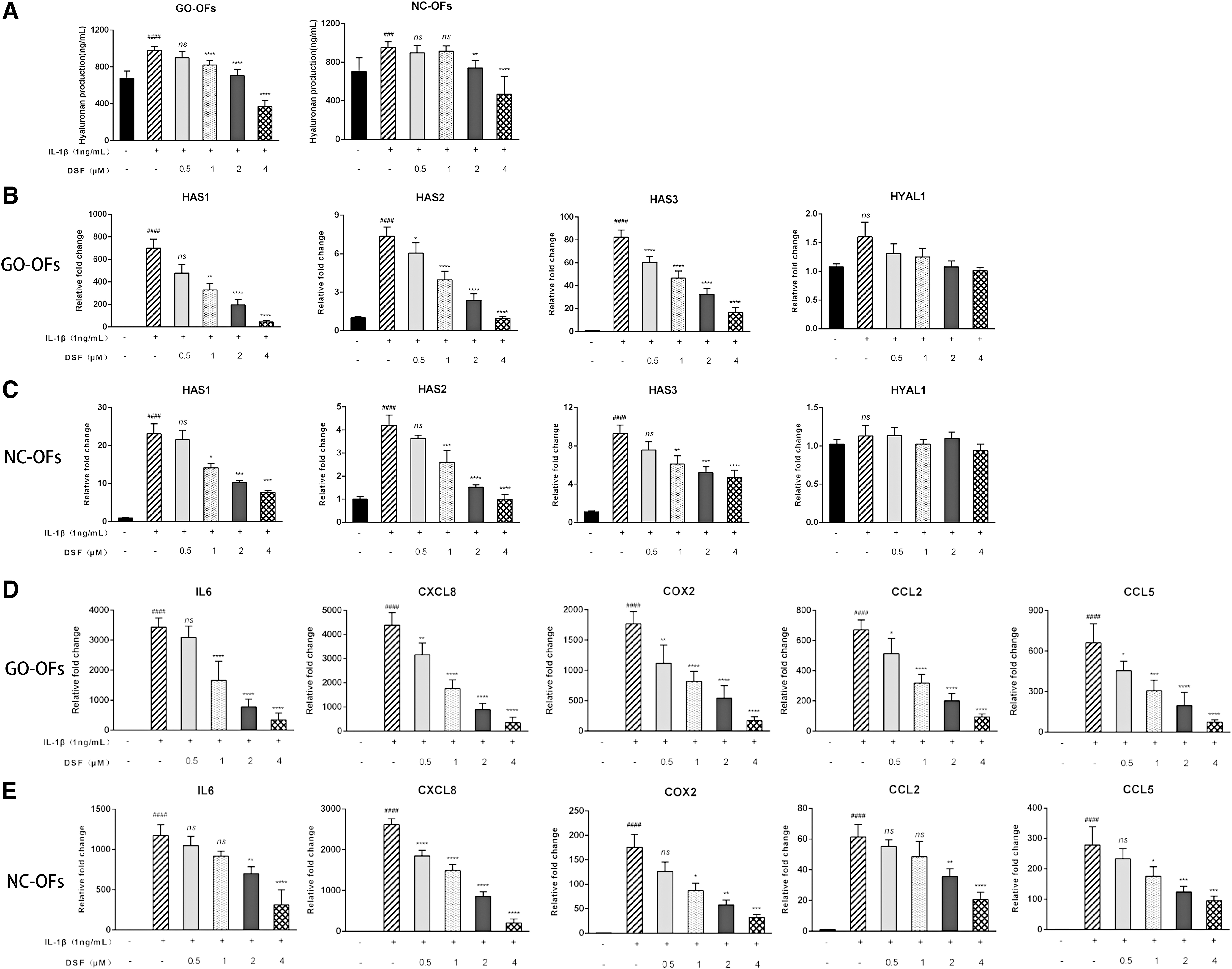
DSF attenuates the production of HA and suppresses the expression of inflammatory molecules. (
DSF exerts antifibrotic effects
Fibrosis is one of the main histopathological changes in GO orbital tissue (45). To explore whether DSF exerts antifibrotic effects on GO OFs, we measured the mRNA expression of markers associated with fibrosis. Cells treated with 10 ng/mL TGF-β1 for 48 hours exhibited increased ACTA2, FN1, COL1A1, COL1A2, and TIMP1. Further, cotreatment with DSF significantly downregulated the levels of these markers (Fig. 4A). As shown in Figure 4B, the effects of DSF on GO OFs migration were assessed by scrape wound assays. At 72 hours, the wounds were clearly closed in all groups. We noticed delayed wound closure in the DSF-treated groups compared with the untreated groups. Over time, quantitation of wound widths revealed a significant inhibitory effect of DSF on GO OFs motility (Fig. 4C).
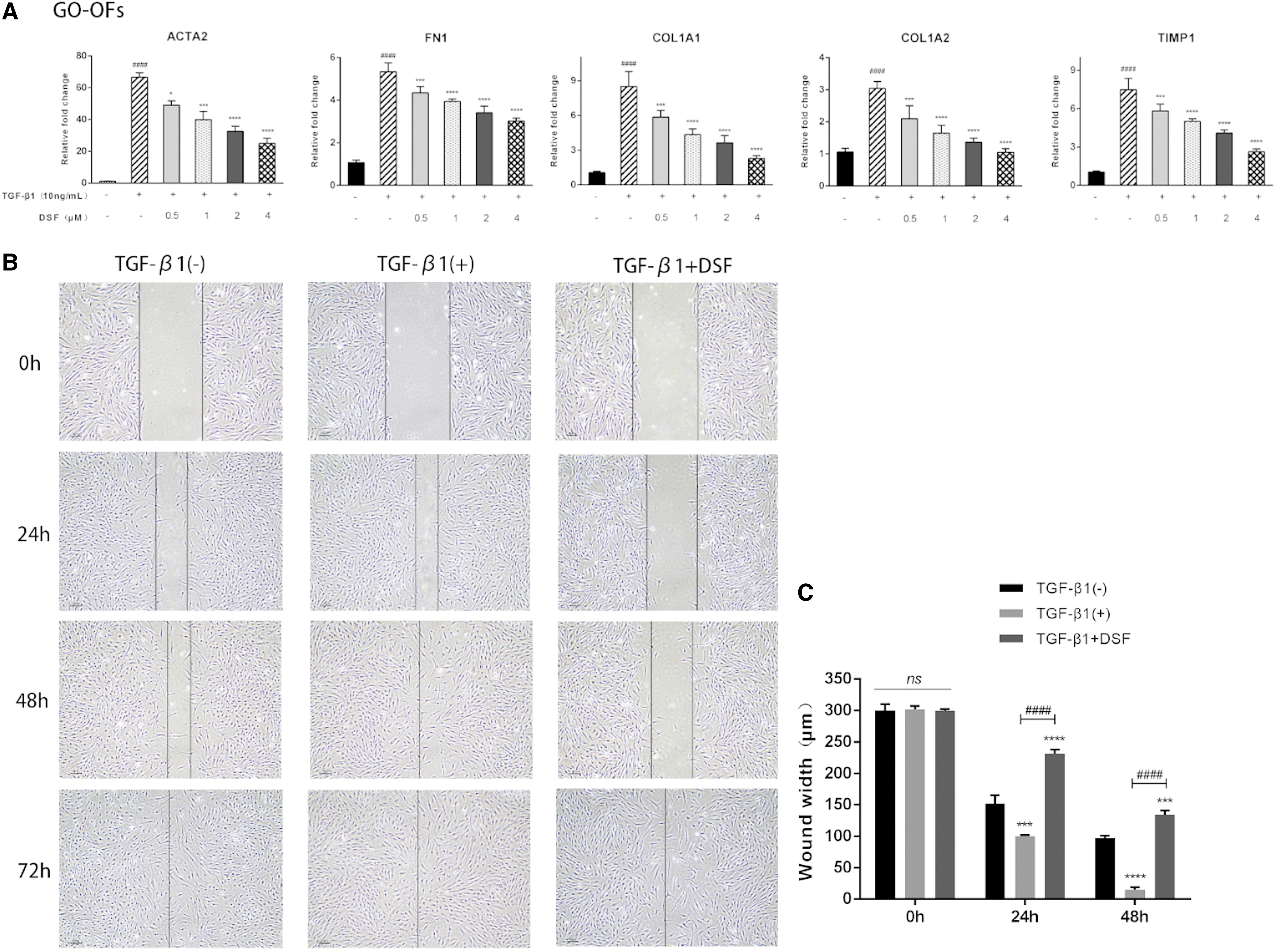
DSF exerts antifibrotic effects. (
Discussion
At present, the treatment options for GO are limited. New types of immunosuppressants and biological agents have become the focus of current research. A combination of mycophenolate mofetil and methylprednisolone shows a better response in patients with active moderate-to-severe GO (46,47). Rituximab, an anti-CD20 monoclonal antibody, has controversial clinical trial data on GO treatment (48 –50). Tocilizumab offers a meaningful improvement in activity and severity in corticosteroid-resistant GO (25,51,52). In two multicenter, randomized, double-blind clinical trials (53,54), teprotumumab had better results than placebo in reducing exophthalmos, CAS, and diplopia and improving quality of life with rare serious adverse events. Moreover, most patients can maintain it for a long time. However, the efficacies and applications of these treatments require more multicenter, randomized controlled trials.
Orbital tissue volume expansion in GO is determined by several factors, including excessive OFs production of the glycosaminoglycan HA, OFs differentiation into adipocytes (adipogenesis), and tissue remodeling during the late phase of GO (fibrosis) (55). In the current study, we provide evidence for the inhibitory effect of DSF on OFs adipogenesis. Further, DSF potently suppresses HA production and exerts anti-inflammatory and antifibrotic effects in vitro. To the best of our knowledge, this is the first study to investigate the therapeutic effect of DSF on GO OFs.
Due to high consumption rates, rising costs, and long new-drug discovery times, conventional drugs in current use are further examined. Increasingly, many studies have shown that DSF has therapeutic effects on inflammation, autoimmune and fibrotic diseases. Their safety has been ensured through clinical trials and their widespread use in patients suffering from alcoholism (56). Our data show that DSF significantly inhibits adipogenesis in GO OFs. This antiadipogenic effect was confirmed by Oil Red O staining of intracellular lipid droplets, RT-PCR, and Western blotting of adipogenic transcription factors, including PLIN1, PPARγ, c/EBPα, and FABP4. The mechanism of the antiadipogenic effect of DSF was examined by Western blotting. The results indicate that DSF inhibited ALDH1A1 but not the ALDH2 enzyme.
Consistent with previous studies, Bernier et al. (29) suggested that DSF treatment prevented body weight gain and mitigated liver steatosis and pancreatic islet hypertrophy in mice. The loss of ALDH2 did not interfere with DSF-mediated metabolic benefits in mice fed an obesogenic diet. ALDH1A1 regulates adipogenesis, abdominal fat formation in adipocytes (57,58), and paracardial fat remodeling (59). ALDH1A1-null adult mice have been shown to resist high-fat (HF) diet-induced weight gain (39,60), which suggests that ALDH1A1 is necessary for HF diet-induced obesity (61,62). Jia et al. reported that DSF could inhibit ethanol-induced osteoclastogenesis via ALDH1A1 (63). ERK activation is essential in the early stage of adipogenesis (64), and DSF-mediated inhibition of ERK phosphorylation was confirmed in our study. Pretreatment with NCT-501, a known ALDH1A1 inhibitor, did not exhibit the suppressive effect of DSF on ERK phosphorylation. In summary, DSF mediates antiadipogenic effects on GO OFs partially by inhibiting ALDH1A1 through the suppression of p-ERK. Our data do not show that the experimental concentrations of DSF influence apoptosis or ROS levels in OFs.
Central to orbital tissue remodeling in GO is the disordered accumulation of HA (65). This study demonstrates that treatment with DSF dose dependently reduces IL-1β-induced HA production. In addition to inhibiting adipogenesis, DSF exerts multiple anti-inflammatory and antifibrotic effects. DSF inhibits IL-17-associated autoimmune inflammation by upregulating miR-30a (66), lobular inflammation in nonalcoholic steatohepatitis (67), mucosal fibrosis in mucous membrane pemphigoid (68), and epithelial–mesenchymal transition (69). As expected, IL-1β-induced mRNA levels of proinflammatory molecules in OFs were significantly suppressed by DSF. Moreover, the antifibrotic effects of DSF were at least partially explained.
There are several limitations in the present study. First, due to the absence of relevant animal orbital models of GO, we could not evaluate in vivo effects or determine the optimum dose of DSF. Second, in vitro, we only focused on OFs without considering crosstalk between immunologically active cells and OFs or the influence of the microenvironment. Third, we only explored the mechanism of the antiadipogenic effect in the current study. The remainder of the mechanism of DSF has not yet been elucidated, which is our future plan.
In conclusion, DSF is a good disease-modifying drug candidate for treating GO and should be investigated in clinical trials. The results of this study are noteworthy and indicate the potential of DSF as a therapeutic agent for the treatment of GO.
Footnotes
Acknowledgments
The authors thank Peng Wang, MD, PhD (Department of Ophthalmology, the First Affiliated Hospital of Chongqing Medical University, Chongqing Key Laboratory of Ophthalmology, Chongqing Eye Institute, Chongqing, China) for his help with the illustrations.
Authors' Contributions
X.W., S.Y., and H.Y. are jointly responsible for experimental design, experimental implementation, and article writing. J.C., L.S., and L.F. are responsible for the guidance of experimental operation and correcting the details of the experimental design. X.W. and T.Z. are responsible for collecting sample tissues and conducting primary cell culture. R.C., W.X., and H.Y. are responsible for the overall control of the experiment and revision of the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The material presented in this work was supported with funding from the National Natural Science Foundation (81670887 and 81870689) of China and the Fundamental Research Funds of the State Key Laboratory of Ophthalmology.
Supplementary Material
Supplementary Table S1
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3