
Abstract
Background:
The pathogenesis of Graves' hyperthyroidism (GH) and associated Graves' orbitopathy (GO) appears to involve stimulatory autoantibodies (thyrotropin receptor [TSHR]-stimulating antibodies [TSAbs]) that bind to and activate TSHRs on thyrocytes and orbital fibroblasts. In general, measurement of circulating TSHR antibodies by clinical assays correlates with the status of GH and GO. However, most clinical measurements of TSHR antibodies use competitive binding assays that do not distinguish between TSAbs and antibodies that bind to but do not activate TSHRs. Moreover, clinical assays for TSAbs measure stimulation of only one signaling pathway, the cyclic adenosine monophosphate (cAMP)-protein kinase A (PKA) pathway, in engineered cells that are not thyrocytes or orbital fibroblasts. We determined whether measuring TSAbs by a cAMP-PKA readout in engineered cells accurately reveals the efficacies of stimulation by these antibodies on thyrocytes and orbital fibroblasts.
Methods:
We measured TSAb stimulation of normal human thyrocytes and orbital fibroblasts from patients with GO in primary cultures in vitro. In thyrocytes, we measured secretion of thyroglobulin (TG) and in orbital fibroblasts secretion of hyaluronan (hyaluronic acid [HA]). We also measured stimulation of cAMP production in engineered TSHR-expressing cells in an assay similar to clinical assays. Furthermore, we determined whether there were differences in stimulation of thyrocytes and orbital fibroblasts by TSAbs from patients with GH alone versus from patients with GO understanding that patients with GO have accompanying GH.
Results:
We found a positive correlation between TSAb stimulation of cAMP production in engineered cells and TG secretion by thyrocytes as well as HA secretion by orbital fibroblasts. However, TSAbs from GH patients stimulated thyrocytes more effectively than TSAbs from GO patients, whereas TSAbs from GO patients were more effective in activating orbital fibroblasts than TSAbs from GH patients.
Conclusions:
Clinical assays of stimulation by TSAbs measuring activation of the cAMP-PKA pathway do correlate with stimulation of thyrocytes and orbital fibroblasts; however, they do not distinguish between TSAbs from GH and GO patients. In vitro, TSAbs exhibit selectivity in activating TSHRs since TSAbs from GO patients were more effective in stimulating orbital fibroblasts and TSAbs from GH patients were more effective in stimulating thyrocytes.
Introduction
In patients with Graves' disease (GD), autoantibodies that bind to and activate thyrotropin (TSH) receptors (TSHRs) on thyrocytes and orbital fibroblasts appear to mediate, at least in part, the pathogenesis of Graves' hyperthyroidism (GH) and associated Graves' orbitopathy (GO), which is also referred to as thyroid eye disease. Because production of these antibodies is not regulated, they are chronically present in the patient's circulation and constitutively stimulate these cells. TSHR antibodies that activate TSHRs may be termed TSHR-stimulating antibodies (TSAbs) (1). In general, measurement of circulating TSHR antibodies has been found to correlate with the severity and activity of GH and GO (2 –4).
Although these assays are clinically useful for the rapid diagnosis and monitoring of GH and high serum concentrations of TSH-R-Ab have been associated with GO (2,5), these measurements may not be able to distinguish between GH and GO patients. Therefore, designing assays with these capabilities requires deeper understanding of how TSAbs can lead to GH or GO.
Most clinical measurements of TSHR antibodies use competitive binding assays that do not distinguish between TSAbs and other antibodies that target TSHRs, which may better be termed TSHR-binding antibodies (TRAbs) (6 –8). TRAbs may include antibodies that do not activate TSHRs, and therefore, may not stimulate thyrocytes or orbital fibroblasts. Moreover, clinical assays that do assess TSAbs measure the stimulation of only one of several signaling pathways initiated by TSHR, namely the cyclic adenosine monophosphate (cAMP)-protein kinase A (PKA) pathway, in engineered cells that are not thyrocytes or orbital cells (9,10).
It is now known that TSHR signals via multiple pathways, and at least one other pathway that involves cross talk between TSHR and the insulin-like growth factor 1 receptor (IGF-1R), use a signal transduction pathway that is independent of cAMP in thyrocytes and orbital fibroblasts (11,12). Thus, measurement of TSAbs only by measuring the cAMP-PKA pathway in engineered, nonthyroidal, nonorbital cells may not “tell the whole story” nor accurately reflect all TSAb signaling activities.
We determined whether measuring TSAb activities by a cAMP readout in engineered cells would accurately capture the efficacies of stimulation by these antibodies in the pathogenesis of GH and GO. Also, we studied whether there were differences in stimulating activities toward thyrocytes and orbital fibroblasts in TSAbs from GH versus GO patients. We understand that patients with GO likely have GH and GO TSAbs, but we refer to this disease phenotype as GO.
To accomplish these goals, we measured the stimulating activities of these antibodies in normal human thyrocytes and in orbital fibroblasts from patients with GO (GOFs) in primary cultures in vitro (12,13). In thyrocytes, we measured the production of thyroglobulin (TG) because it is the precursor for thyroid hormone production, and in GOFs, we measured the production of hyaluronan (hyaluronic acid [HA]) because it is a major component of the increased extracellular matrix found in patients with GO. We also measured the stimulation of cAMP production in engineered TSHR-expressing cells (HEK-TSHR cells) in vitro (13). This assay served as an alternative to the clinical assays that measure the activity of the cAMP-PKA pathway in other engineered, nonthyroidal nonorbital cells. Furthermore, we determined whether there were differences in stimulation of these cell types by TSAbs from patients with GH alone and patients with GO.
Materials and Methods
Materials
Human M22 was purchased from Kronus (Star, ID). Dulbecco's modified Eagle's medium (DMEM), 1M HEPES buffer, and Hanks' Balanced Salt Solution (HBSS) were obtained from Mediatech, Inc. (Manassas, VA). Hydrocortisone, insulin, cholera toxin, 100 × antibiotic/antimycotic solution, and human TG enzyme-linked immunosorbent assays (ELISAs) were purchased from Sigma-Aldrich (St. Louis, MO). Recombinant human epidermal growth factor was obtained from PeproTech (Rocky Hill, NJ). Y-27632 dihydrochloride was purchased from Bio-Techne (Minneapolis, MN). Fetal bovine serum (FBS), hygromycin B, 100 × penicillin/streptomycin solution, L-glutamine, Ham's F-12 nutrient mixture, trypsin-ethylenediaminetetraacetic acid (EDTA), Slide-A-Lyzer dialysis cassettes, and cAMP-Screen Chemiluminescent Immunoassay were obtained from Thermo Fisher Scientific (Carlsbad, CA).
Bovine serum albumin (BSA) Fraction V was obtained from MP Biomedicals (Santa Ana, CA). One MDa HA was purchased from Lifecore Biomedical (Chaska, MN). Hyaluronic Acid Test Kits were purchased from Corgenix (Broomfield, CO). Amicon® Ultra-15 Centrifugal Filter Units were purchased from Millipore Sigma (Massachusetts, MA).
Acquisition of human thyroid tissue and orbital tissue and sera from patients with GH and GO
Orbital connective tissue was obtained from patients with severe GO who underwent orbital decompression surgery at the joint thyroid-eye-clinic of the Johannes Gutenberg University (JGU) Medical Center, Mainz, Germany. Patients were classified with phenotypically overt and clinically severe GO, according to the Guidelines of the European Thyroid Association/EUGOGO for the management of GO (14). The collection of patients' tissues and sera was approved by the Ethics Committee of the Medical Chamber of the State Rhineland-Palatinate, Germany, and by the Institutional Review Board of the JGU Medical Center. Written informed consent was received from all patients before collection of blood and tissue samples.
Thyroid tissue samples were collected from normal thyroid tissue of patients undergoing total thyroidectomy for thyroid cancer at the National Institutes of Health Clinical Center. Patients provided informed consent on an Institutional Review Board-approved protocol, and materials were received anonymously with approval of research activity through the Office of Human Subjects Research, National Institutes of Health.
Isolation of primary orbital fibroblasts and thyrocytes and cell culture
Tissue was obtained from GO patients who underwent orbital decompression surgery. Orbital fibroblasts were isolated as previously described (15,16). Briefly, tissue explants were minced, treated with collagenase type IV, and plated in culture dishes containing complete growth media comprising high-glucose DMEM with FBS (10% vol/vol), penicillin (100 U/mL), and streptomycin (100 μg/mL). Resulting monolayer outgrowths of adherent fibroblasts/preadipocytes (GOFs) were serially passaged with 0.05% trypsin/EDTA and cultured in F-media designed to prevent the dedifferentiation of isolated cells.
These media comprised DMEM with FBS (10% vol/vol), penicillin (100 U/mL), streptomycin (100 μg/mL), L-glutamine (2 mM), Ham's F-12 nutrient mixture (25% vol/vol), hydrocortisone (25 ng/mL), epithelial growth factor (0.125 ng/mL), insulin (5 μg/mL), cholera toxin (11.7 nM), gentamicin (10 μg/mL), fungizone (250 ng/mL), and Y-27632 (5 μM). Cells were maintained in a humidified 7% CO2 incubator at 37°C. Strains were used at passages 3 and 4.
Thyroid tissue was obtained from patients undergoing thyroid tumor removal. Thyrocytes were isolated as previously described (17). Briefly, tissue explants were minced, treated with collagenase type IV, and plated in culture dishes containing complete growth media that comprised high-glucose DMEM with FBS (10% vol/vol), penicillin (100 U/mL), and streptomycin (100 μg/mL). After 24 hours of incubation, the nonadherent cells were removed and the remaining thyrocytes were serially passaged with 0.05% trypsin/EDTA in the same media. Cells were maintained in a humidified 5% CO2 incubator at 37°C. Strains were used at passages 4–7.
Human embryonic kidney HEK293 cells stably expressing TSHRs (HEK-TSHR cells) were generated as previously described (18) and cultured at 37°C in a 5% CO2 humidified incubator using DMEM with FBS (10% vol/vol), penicillin (100 U/mL), streptomycin (100 μg/mL), and hygromycin (250 μg/mL).
Purification of immunoglobulins from whole serum of patients with GH alone and patients with GO
Antibodies were isolated from whole serum by thiophilic affinity chromatography as previously described (16). Briefly, a 2-mL gravity flow column was packed with Thiophilic-Superflow resin and equilibrated according to the manufacturer's directions. Samples were diluted 1:10 in sample buffer (50 mM sodium phosphate and 0.55 M sodium sulfate; pH 7.0) and applied to the column. Unbound proteins were washed with equilibration buffer (50 mM sodium phosphate and 0.5 M sodium sulfate, pH 7.0).
Immunoglobulins G (Igs) were eluted with 10 mL of elution buffer (20 mM sodium phosphate and 20% glycerol; pH 7.0). The eluant samples were dialyzed with Slide-A-Lyzer dialysis cassettes in HBSS containing 10 mM (4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid [HEPES]) (pH 7.4). The dialyzed samples were then concentrated to their original volume using an Amicon Ultra-15 Centrifugal Filter Unit. This method likely resulted in a heterogeneous mixture of stimulatory, inhibitory, and blocking TSHR antibodies with Ig concentrations that varied among individual samples. Igs were diluted 1:5 before use in experiments, and high concentrations (0.8–2 mg/mL) were used to ensure sufficient TSAb stimulatory activities were exhibited to induce a detectable cAMP, HA, or TG response.
Measurement of cAMP production
cAMP levels were measured in an engineered cell system as previously described (15). HEK-TSHR cells were stepped-down for 30 minutes in 10 mM HEPES in HBSS and then treated with Igs in the presence of 3-isobutyl-1-methylxanthine for 60 minutes. Concentration of cAMP from cell lysates was measured using the cAMP-Screen Chemiluminescent Immunoassay according to the manufacturer's directions.
Measurement of TG response
Thyrocytes were grown to confluence and then plated in DMEM with 10% FBS for the experiment. After 24 hours, the cells were stepped-down to DMEM with 0.1% BSA to differentiate the cells. After 24 hours of being stepped-down, the cells were treated with TSH, M22 (diluted in HBSS), or purified Igs and incubated for 5 days. Conditioned media were collected and stored temporarily at 4°C. Conditioned media were assayed using a human TG ELISA kit as previously described (19).
Measurement of HA response
Orbital fibroblasts (GOFs) were grown to confluence and then plated in DMEM with 10% FBS for the experiment. After 24 hours, cells were treated with TSH, M22 (diluted in HBSS), or purified Igs and incubated for 5 days. Conditioned media were collected and stored temporarily at 4°C. Conditioned media were assayed using a modified Corgenix HA ELISA kit as previously described (15,16).
Statistics
Statistical analyses of the data were performed by using GraphPad Prism version 8.4 Windows (GraphPad Software). p < 0.05 by linear regression analysis was used to determine significance of correlations. Analyses were done with Igs shown to stimulate cAMP production three standard deviations above basal production. This threshold allowed for 49 of the Ig samples from patients with GH only (GH-Igs) and 49 of the Ig samples from patients with GO (GO-Igs) to be used in the analysis.
Results
All 50 patients with GH without clinical signs and symptoms of orbitopathy were female and on antithyroid drugs (ATD) (methimazole [MMI] dose 2.5–10 mg per day). In comparison, 29 of 50 (58%) patients with GO were male, and all GO patients were also on ATD (MMI 2.5–20 mg/day). Mean disease durations of GH and GO were 6.3 months (standard deviation [SD] 2.9) and 11.4 months (SD 5.1), respectively. Mean age of the GH and GO patients was 38.1 years (SD 6.2) and 67.3 years (10.7), respectively. Mean serum levels of the stimulatory TSH receptor autoantibodies (TSAb) were specimen to reference ratio (SRR, cell-based bioassay) 468.7% (SD 35.66) and 745 SRR% (59.1) for the GH and GO subjects, respectively.
All patients who underwent orbital decompression surgery had a clinically severe GO with a clinically activity score of 2–7 points, and all have been pretreated with intravenous steroids according to the EUGOGO guidelines (14,20).
We purified Igs from 50 patients with GH and no clinical evidence of GO (GH-Igs) and 50 patients with GO (GO-Igs) understanding that GO patients likely had GH-Igs and GO-Igs. The activities of these samples were measured in three assays: (1) acute stimulation of cAMP production in HEK-TSHR cells for 1 hour; (2) stimulation of TG secretion by primary cultures of normal human thyrocytes over 5 days; and (3) stimulation of HA secretion by primary cultures of GOFs over 5 days. The assay for stimulation of cAMP production in engineered HEK-TSHR cells (21) was used instead of a clinical assay for TSAbs so as to use conditions more similar to the assays for TG and HA.
We measured TSAb activities in purified Igs rather than in serum samples. Using this approach, the effects of other agonists found in sera that do not signal via TSHR were greatly reduced. Figure 1A shows the data from the individual samples using GH-Igs, and Figure 1B shows the data using GO-Igs. To normalize and combine the data using different orbital GOF and thyrocyte strains, the data were calculated as percent of the maximal level stimulated by 10 mU/mL TSH (% TSH Max) in each assay.
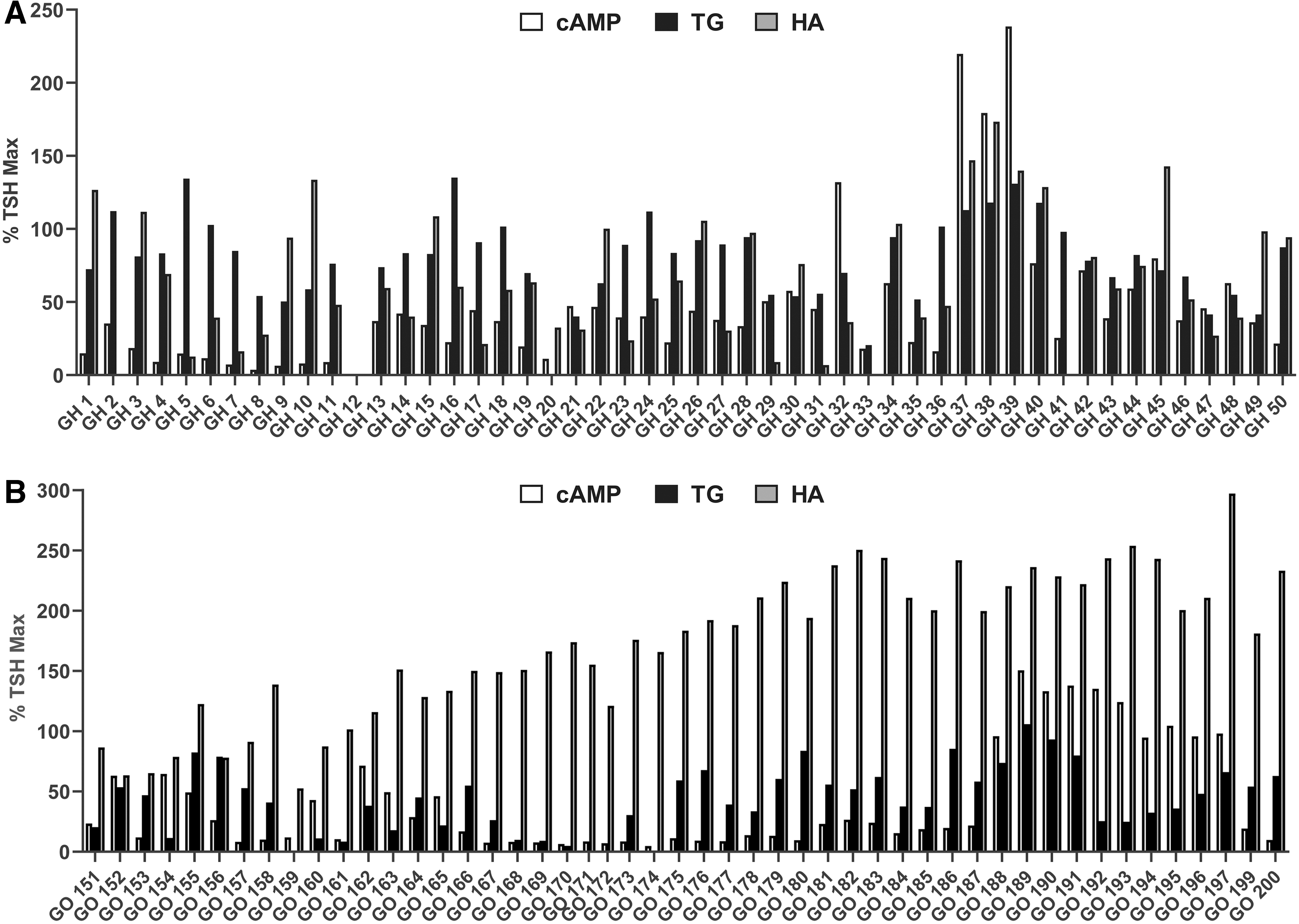
Purified GH-Ig
Figure 2 presents these same data comparing stimulation by GO-Igs with that by GH-Igs. Stimulation of cAMP by GH-Igs and GO-Igs was not significantly different (47.0 ± 7.0% vs. 41.0 ± 6.1%) (Fig. 2A). A major finding was the differences in stimulation of TG and HA secretion by GO-Igs versus GH-Igs. Stimulation of TG secretion was increased to higher levels by GH-Igs than by GO-Igs (80.8 ± 3.7% vs. 44.7 ± 3.8%, p < 0.0001) (Fig. 2B), whereas HA secretion was increased to higher levels by GO-Igs than by GH-Igs (172.5 ± 8.6% vs. 68.7 ± 6.2%, p < 0.0001) (Fig. 2C).

Comparison of cAMP, HA, and TG stimulation by GO-Igs and GH-Igs. GO-Igs (filled circles) and GH-Igs (unfilled squares) were used to stimulate
Based on these findings, we conducted a more detailed analysis of the correlations between stimulation of cAMP production and stimulation of TG secretion or HA secretion by individual GH-Ig and GO-Ig preparations. Figure 3 shows the correlations between stimulation of cAMP production and TG secretion (Fig. 3A) and between stimulation of cAMP production and HA secretion (Fig. 3B) of the individual GO-Ig and GH-Ig preparations. Linear regression analyses showed significantly nonzero correlations between cAMP levels and TG secretion for both GH-Igs (R 2 = 0.11, p = 0.0212) and GO-Igs (R 2 = 0.09, p = 0.0406) (Fig. 3A) and between cAMP levels and HA secretion for both GH-Igs (R 2 = 0.23, p < 0.001) and GO-Igs (R 2 = 0.14, p = 0.0073) (Fig. 3B).
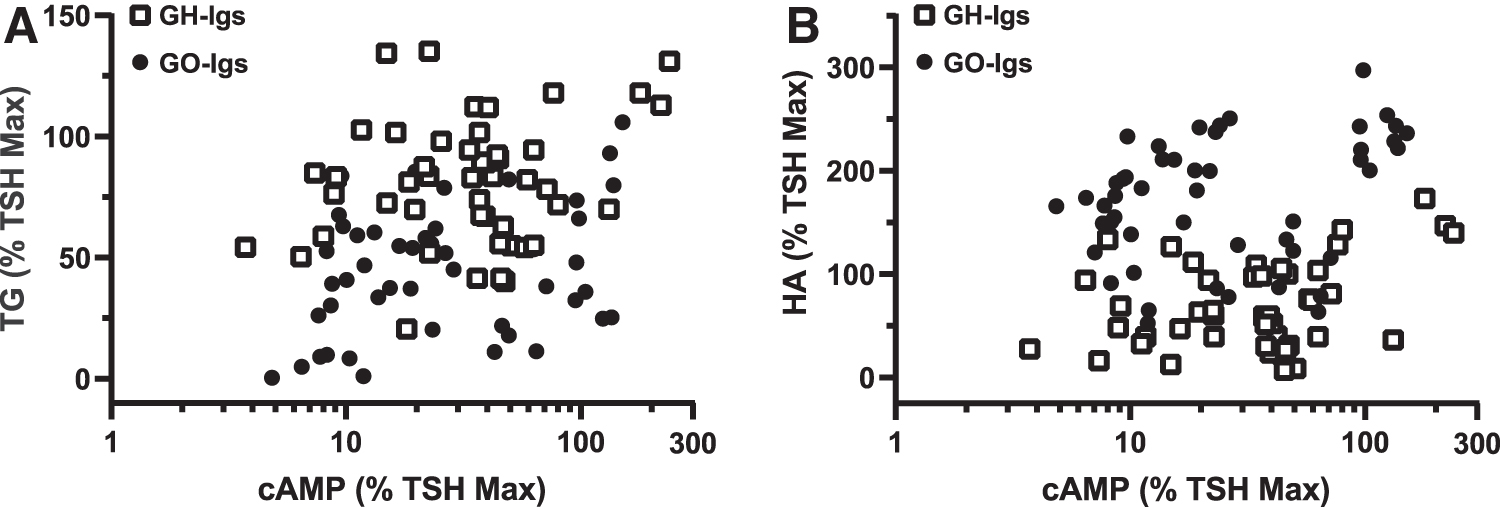
Correlation between GH-Ig and GO-Ig stimulation of cAMP production by HEK-TSHR cells, TG secretion by thyrocytes, and HA secretion by orbital fibroblasts.
The slopes of the TG versus cAMP lines were not significantly different between the GH-Ig and GO-Ig groups, nor were the slopes of the lines comparing HA versus cAMP. When GH-Igs and GO-Igs are combined into the same group, TG and HA positively correlate with cAMP, but HA versus cAMP significantly deviates from linearity according to a runs test. Taken together, GH-Igs are stronger stimulators of TG, and GO-Igs are stronger stimulators of HA. However, the strength of the correlation with cAMP cannot be used to distinguish between the two groups of antibodies.
Discussion
In this study, we sought to answer the following questions: (1) do in vitro stimulation assays with human thyrocytes/orbital fibroblasts correlate with current clinical assays in assessing the activities of TSAbs in GH and GO patients; and (2) do TSAbs from patients with GH alone differ in their activities to stimulate thyrocytes and orbital fibroblasts from TSAbs from patients with GO with accompanying GH? TSAbs are involved in GH and GO pathogenesis by stimulating TSHRs on thyrocytes and orbital fibroblasts, respectively. It is important to note that TSAbs obtained from Graves' patients are not monoclonal but comprised multiple antibodies that may have stimulatory or inhibitory activities, and that the results measured in all assays are the average effects of these different responses. We performed head-to-head comparisons of the TSAb-stimulating activities on nonthyroidal, nonorbital cells engineered to overexpress TSHRs and on human thyrocytes and human orbital fibroblasts in primary cultures using antibodies isolated from distinct GH-only and GO patient populations. The differences we observed when comparing autoantibodies from these two groups reveal the importance of cell context to TSHR signaling and the intrinsic differences between GH and GO autoantibodies.
This study demonstrates the potential limitations of using nonthyroidal, nonfibroblast cells overexpressing TSHR to determine TSAb activity, as this is currently done via a single intracellular pathway. Indeed, it is likely that activation of TSHRs in thyrocytes and orbital fibroblasts involves activation of multiple pathways in addition to that initiated by stimulation of cAMP production (22 –24). Different TSAbs in the populations of autoantibodies obtained from Graves' patients may signal via different pathways in addition to those initiated by cAMP and may exhibit biased signaling.
These data also indicate that measurement of cAMP appears not to quantitatively assess stimulation of HA secretion and appears not to be an appropriate proxy for TSHR activation of orbital fibroblasts. Although generally HA and TG secretion positively correlated with cAMP production for both GO-Igs and GH-Igs, each group displayed a clear selectivity in downstream TSHR responses. Although at least a partial dependence of TG production on cAMP production is well established (25), it is not definitively known whether HA production is a downstream response of cAMP generation.
Indeed, a previous report suggested that activation of orbital fibroblasts by TSAbs was independent of cAMP signaling (26). Had TSAb activity from patients with GO been primarily dependent on cAMP, then GH-Igs would be expected to stimulate HA as well as GO-Igs, and combined analysis of both GH- and GO-Igs would have remained linear. Instead, the opposite was the case, suggesting that TSHR stimulates HA secretion through a pathway(s) that is, at least in part, independent of cAMP. This idea is supported by our previous observation that the TSHR/IGF-1R cross talk, which does not stimulate cAMP production (16), is a major pathway for TSAb-stimulated HA secretion by fibroblasts from patients with GO.
An important observation made herein is that autoantibodies generated by GH and GO patients may act as biased agonists for TSHR and that they have different effects on thyrocytes and orbital fibroblasts. This idea that TSAbs may be biased to signal via different pathways has been previously reported for monoclonal TSAbs in FRTL-5 rat thyroid cells (27).
The differences we report in stimulation by TSAb populations found in GH-Ig and GO-Ig pools may be characterized as biased signaling and may explain, in part, why patients exhibit different levels of stimulation of thyrocytes, hyperthyroidism, and of orbital fibroblasts. Of course, other differences may be involved, including cell autonomous signaling dissimilarities.
What has been clearly shown is that assessing the magnitude of the cAMP-PKA pathway stimulation by TSAbs is a good indicator of stimulation of thyrocytes, but may be a poorer measurement of stimulatory activity at orbital fibroblasts. Nevertheless, this type of assay, including the ones in use in the clinic currently, does appear to correlate with the severity of GO (2) and to have predictive value for the future development of GO (3). Notably, dilution analysis to measure TSAb-titers showed that GO patients had higher TSAb titers than GH patients (2).
The current study suggests that patients with GO have antibodies that stimulate TSHR pathways in addition to cAMP-PKA, and these cAMP-independent pathways may be better predictors of GO.
In conclusion, although in general, measurement of circulating TSHR autoantibodies by assays used in the clinic currently have been found to correlate with the status of GH and GO (2 –4), we found that they correlate better in vitro with stimulation of thyrocytes than orbital fibroblasts. This is so because the clinical assays currently only measure TSAb stimulation of the cAMP-PKA pathway, which appears more important in TG secretion by thyrocytes than in HA secretion by orbital fibroblasts. Potentially, exploring additional intracellular pathways and their readouts may help differentiate between thyroidal and extrathyroidal pathogenesis and eventually between both phenotypes (GH only vs. GH + GO).
Footnotes
Acknowledgments
We thank Tanja Diana, PhD, MSc (Molecular Thyroid Lab), Heike Elflein, MD, and Katharina Ponto, MD (Department of Ophthalmology), JGU Medical Center, Mainz, Germany, who have generously prepared and provided GO patient tissue and serum samples from GD patients for our experiments. We are also grateful for the contributions of Brianna Daley and Andrew Tobey of the Metabolic Disease Branch, National Institutes of Diabetes and Digestive and Kidney Diseases, National Institutes of Health, Bethesda, Maryland, who provided thyroid tissue.
Authors' Contributions
C.C.K., G.J.K., J.K.G., S.N., and M.C.G. made substantial contributions to the conception and design of the work. C.C.K., G.J.K., A.A., S.N., and M.C.G. made substantial contributions to the acquisition, analysis, or interpretation of data for the work. C.C.K., G.J.K., A.A., J.K.G., S.N., and M.C.G. made substantial contributions to drafting the work or revising it critically for important intellectual content. C.C.K., G.J.K., A.A., J.K.G., S.N., and M.C.G. gave final approval of the version to be published and agreed to be accountable for all aspects of the work.
Author Disclosure Statement
The authors at the NIH have nothing to disclose. The JGU Medical Center, Mainz, Germany, has received research-associated funding from the JGU Medical Faculty, AdvanceCor (Germany), Apitope (United Kingdom), Berlin-Chemie (Germany), Byondis (the Netherlands), Horizon Therapeutics (USA), Immunovant (USA), ISAR (Germany), Mediomics (USA), Merck (Germany), Novartis (USA), Quidel (USA), River Vision (USA), and Roche (Switzerland). G.J.K. consults for GlycoEra (Switzerland), Immunovant, Mediomics, Merck, Novartis, Roivant (USA), Quidel, and VASARAGEN (USA).
Funding Information
This work was supported by the Intramural Research Program of the National Institute of Diabetes and Digestive and Kidney Diseases, National Institutes of Health (Z01 DK011006).