
Abstract
Background:
Sex dimorphism strongly impacts tumor biology, with most cancers having a male predominance. Uniquely, thyroid cancer (TC) is the only nonreproductive cancer with striking female predominance with three- to four-fold higher incidence among females, although males generally have more aggressive disease. The molecular basis for this observation is not known, and current approaches in treatment and surveillance are not sex specific.
Summary:
Although TC has overall good prognosis, 6–20% of patients develop regional or distant metastasis, one third of whom are not responsive to conventional treatment approaches and suffer a 10-year survival rate of only 10%. More efficacious treatment strategies are needed for these aggressive TCs, as tyrosine kinase inhibitors and immunotherapy have major toxicities without demonstrable overall survival benefit. Emerging evidence indicates a role of sex hormones, genetics, and the immune system in modulation of both risk for TC and its progression in a sex-specific manner.
Conclusion:
Greater understanding of the molecular mechanisms underlying sex differences in TC pathogenesis could provide insights into the development of sex-specific, targeted, and effective strategies for prevention, diagnosis, and management. This review summarizes emerging evidence for the importance of sex in the pathogenesis, progression, and response to treatment in differentiated TC with emphasis on the role of sex hormones, genetics, and the immune system.
Introduction
Overlooking sex as an important biological determinant of health and disease can lead to mistaken concepts underlying the incidence, prevalence, diagnosis, and management of neoplasms. The belief that both sexes have identical biology can generate conclusions based on single-sex data neglecting sex dimorphism as a significant variable, for example, as in the male predominance in most cancers (1). Sex refers to the biological differences between males and females, including differences in chromosomal arrangements, reproductive organs, and hormone levels. Gender relates to human behaviors, attitudes, and expectations based on society or cultures.
Thyroid cancer (TC) is the most common endocrine neoplasm (2), of which 94% are well-differentiated thyroid carcinomas (DTC) of either papillary thyroid carcinomas (PTC) or follicular thyroid carcinomas (FTC) type, with PTC accounting for >85% diagnosed in the United States and FTC reflecting 7% (3,4). Although TC is the only nonreproductive based cancer with a strong female bias, with three- to four-fold higher incidence in women, males generally demonstrate a more aggressive disease course (1,5 –7).
Men often present at a more advanced disease stage and have a higher recurrence rate compared with women (8 –10). The molecular basis for this sex bias is enigmatic, and current evaluation and treatment approaches are not tailored to sex. Most patients with DTC are successfully treated with thyroidectomy followed selectively by radioiodine ablation or adjuvant therapy, according to their risk stratification (11).
Management of the ∼6–20% of patients who develop distant metastasis is problematic, especially in the 33% of patients with dedifferentiated tumors that have lost ability to concentrate iodine, becoming RAI refractory (RAIR). These patients exhibit higher mortality with a 10-year survival rate of only 10% from the time of detection of metastases (12 –15). While novel alternative treatment strategies for RAIR TC have emerged, for example, tyrosine kinase inhibitors, survival benefit has not been demonstrated, and use is associated with significant toxicities (16 –18). Although immunotherapy is emerging as a possible option for advanced TC, its rationale is based on research from non-TC trials associated with major toxicity and no demonstrably clear benefit (19 –21).
It is plausible that better understanding of molecular mechanisms underlying sex differences in TC could contribute insights into the development of sex-specific, targeted, and effective strategies for prevention, diagnosis, treatment, or surveillance.
Epidemiological and Clinical Studies
Epidemiological studies among nonreproductive cancers indicate a strong male sex predominance with a significantly higher incidence in men at 32 sites, with disparities >2-fold at 15 sites and >4-fold at 5 sites (1). The sex disparity in TC increases from childhood through puberty, with gradual disappearance after menopause and equalization of incidence in both sexes in older adults (5,22) suggesting a role for sex hormones.
Despite the higher TC incidence in females, several retrospective studies concluded that sex was not a significant prognostic feature (23 –25). In a cohort of 2479 TC patients (25), regression analysis indicated no significant sex differences in disease-specific survival (DSS) for any histological subtype of TC. As this analysis was performed on patients of all ages rather than subdividing patients into different reproductive age groups, their conclusion was not age-controlled, and it could reflect an averaging out of age-dependent, sex-specific differences identified in more recent studies.
However, age, histological subtype, and tumor stage had a significant impact on patient survival. In another study of 241 PTC cases (23), patient age at presentation, tumor size, encapsulation, extrathyroidal extension, multicentricity, or distant metastasis were significant prognostic factors. Survival did not differ between the sexes either for all age groups or for patients >40 years. Outcomes in patients <40 years were not analyzed to determine whether they differed in women of child-bearing age from age-matched male controls.
Another study of 4187 patients followed over 35 years (1969–2004) examined the changing features of DTC over time (24). Poor outcomes were associated with age, histologic subtype, tumor size, extrathyroidal microinvasion, and lymph node metastases, but not by sex. Several studies generalizable to all cancers likewise concluded that sex is not a significant variable determining TC outcome. The EUROCARE-4 study indicated that age-adjusted relative survival was higher in women than in men at 15 sites, including TC (26).
Younger age at diagnosis was the main determinant predicting a female advantage becoming negligible in the elderly. A study of 412,471 cancer cases from 25 sites (27) indicated lower 5-year net survival for all cancers in men (47.1%; confidence interval [CI 46.9–47.6]) than women (52.0%, [CI 51.7–52.3]). A survival disadvantage for men was observed for 11 cancers, which included TC. Neither of the latter reports noted a sex difference in TC incidence and concluded that survival in men is lower only when age is considered as a co-variable.
A more recent study confirmed the higher incidence of small TCs (≤2 cm) in women with a ratio of 4.39:1 compared with men (28) using the Surveillance, Epidemiology, and End Results (SEER) database. No sex difference was found in the prevalence of undiagnosed TC at autopsy in available databases. The authors also compared sex difference in the incidence of medullary and anaplastic TC and noted that the female/male ratio approaches 1:1, especially in more aggressive tumors.
They concluded that overdiagnosis contributes to the higher prevalence of DTC in women. As in an earlier study (25), they analyzed patients of all ages rather than subdividing patients into different reproductive age groups, thereby possibly averaging out age-dependent, sex-specific differences.
Recent studies on the incidence, severity, and prognosis of DTC have been appropriately age and gender-stratified and allow greater insight into these important sex-specific variables. For the National Thyroid Cancer Treatment Cooperative Study Group Registry data (6), sex-specific mortality was normalized by using age-matched subjects and a multivariate Cox proportional hazard regression model was employed to assess the impact of age and sex. Although the outcome of PTC was similar between men and women, subgroup analysis showed women <55 years had better DSS than men (hazard ratio [HR] 0.33, [CI 0.13–0.81]).
Notably, the HR for DSS increased to become similar to men for women diagnosed at 55–69 years (HR 1.01, [CI 0.42–2.37]) and at ≥70 years (HR 1.17, [CI 0.48–2.85]). This observation parallels epidemiological data showing an equalization of TC incidence in both sexes in older adults. A retrospective cohort study of SEER data of 68,337 patients with DTC (27) showed that the DSS of follicular variant PTC (FVPTC) and FTC were similar to classical PTC (CPTC) among male patients (FVPTC vs. CPTC: HR = 0.947, p = 0.776; FTC vs. CPTC: HR = 1.512, p = 0.104). However, among premenopausal females (<55 year), FVPTC had better DSS than CPTC (HR = 0.321, p = 0.038) while FTC had worse DSS than CPTC (HR = 3.272, p = 0.013).
Further, in postmenopausal women (>55 years), FTC had a worse prognosis than CPTC (HR = 2.145, p = 0.002) and no statistically significant difference was found between CPTC and FVPTC among older women (HR = 0.724, p = 0.087). When the entire cohort was age-subdivided, among patients <55 years, women had significantly better DSS than men with CPTC (HR = 0.376, p < 0.001) or FVPTC (HR = 0.102, p < 0.001). No sex difference in DSS was observed among patients >55 years, and outcomes in FTC were not affected by sex in patients of all ages. Female sex was also an important prognostic feature in two additional large retrospectively studied cohorts (25,29).
Thus, epidemiological studies in DTC have shown inconsistent results regarding sex-difference in DSS (6,25,26,29 –31). Apparent discrepancies between studies may relate to study design, especially if different reproductive age groups, and statistical analyses were employed. As described earlier, sex disparities in TC are age-dependent, with the greatest sex dimorphism during reproductive age, and studies biased toward older patient cohorts would not be expected to find a sex-specific effect. Given the possible influence of sex hormones on tumor development, progression, and response to therapy, it seems imperative to analyze the interaction of sex and histological subtypes of DTC within specific reproductive ages in large cohorts.
The inherent recall bias in retrospective studies and lack of standard methodology to obtain clinical and pathological data may contribute to discrepancies among epidemiological studies evaluating DTC prognosis.
The Role of Sex Hormones
Based on observations that sex disparities in TC increase from childhood toward adulthood and gradually disappear in older adults (1,5,6,22,32), a key focus on the sex dimorphism in DTC has been on exploring the role of differences in the sex hormones.
Estrogen receptor (ER) is expressed in both normal and malignant thyroid tissues of both sexes (33 –36). In a cohort of 351 patients, ER expression was observed among 24% of benign goiters, 31% of PTC, and 22% of follicular adenomas. No differences in ER expression were found between females and males of different age groups, nor did ER expression correlate with lymph node status and vascular/capsular invasion (33). Some studies have found a higher expression level of ER-alpha subtype (ERα) in thyroid tumors versus normal thyroid tissue.
Expression levels of ERα and androgen receptor were associated with a more aggressive phenotype demonstrating higher levels of postsurgical serum thyroglobulin and capsular invasion. Expression levels of ER beta-subtype (ERβ) were associated with less aggressive phenotypes with less vascular invasion (37). Other studies have found higher levels of ER expression in DTC versus undifferentiated or anaplastic cancers (34,38). Inconsistencies in the outcomes between studies of ER expression may be due to different experimental techniques (e.g., PCR vs. immunohistochemistry).
In addition, other possible confounding factors include the subject's age and cancer histological sub-type. In addition to the presence or absence of ER, studies have investigated ER subtype expression profiles in normal versus malignant thyroid, with findings pointing to a stimulatory effect of ERα on PTC growth and progression and an inhibitory role for ERβ in TC development (35,36,38 –41). The ERα binding activates the signal transduction pathways (ERK/MAPK and PI3K/AKT) related to cell cycle progression and prevention of apoptosis, whereas activation of ERβ induces phosphorylation of p38/MAPK, which is involved in apoptosis (42 –44). Consequently, increased ratios of ERα/ERβ expression are associated with TC development while low ERβ expression is associated with poor survival in TC (40,45,46).
Studies of estrogen effects on both normal and malignant thyroid cell behavior indicate increased DNA synthesis and proliferation, likely through a classic genomic pathway (35,38,47). Estradiol led to 50–150% enhancement of proliferation, enhanced adhesion, migration, and invasion of thyroid cells in vitro (48). That exposure to estrogen modulates levels of adhesion molecules, metalloproteinases, and epithelia to mesenchymal transition molecules indicates its potential role in metastatic processes and the dissemination of cancer cells (49 –53).
Gonadectomy reduced TC incidence only in female rats with irradiation-induced thyroid follicular tumors. Estrogen treatment of castrated mice increased TC incidence in both sexes (54,55). Employing a transgenic FTC model, castration led to lower rates of cancer in females (56). In male rats, orchiectomy led to smaller cancers and testosterone exposure to orchiectomized rats led to a progressively higher tumor incidence (57). Means serum thyrotropin (TSH) was noted to be higher among irradiated rates compared with nonirradiated animals. Further, testosterone treatment of orchiectomized rates resulted in increased mean serum TSH level via an unknown mechanism and may have contributed to tumorigenesis via TSH-mediated growth-promoting actions (57). Testosterone exposure modulated the expression level of tumor suppressor and immune regulatory genes in orchiectomized mice (56).
The underlying molecular mechanism for the observed sex bias in the studies described remains unknown. Inconsistent experimental conditions, divergent study protocols, and use of a variety of experimental tools have contributed to conflicting results.
Genetic and Epigenetic Contributions
Failure to address sex as an important variable in cancer may result from a tendency to assume that men and women have similar biology and genetic code with respect to cancer development, progression, and responses to treatment. A few studies have challenged this dogma and provided evidence that sex differences in cancers go beyond their morphological and behavioral characteristics and that sex-specific molecular signatures may influence outcomes. In this section, we present emerging evidence on sex-specific molecular signatures of cancers with a focus on TC.
Studies interrogating existing large data sets minimizing possible bias inherent in small sample sizes have concluded that sex influences various cancers (58 –61). A computational analysis of The Cancer Genome Atlas (TCGA) indicated significant sex differences at the molecular level for certain cancers (58). Cancers were divided into a strong sex-effect group with a greater number of sex-biased molecular signatures, and a weak sex-effect group with a small number of genes with a sex-biased effect.
The TC fell within the strong sex-effect group, with 78 genes showing clear sex-biased expression signatures. Of these, 40 genes (51%) were located on sex chromosomes. The study also showed that sex dimorphism in the gene expression signature of TC extended to genes with epigenetic function, including genes involved in DNA methylation (KDM5C, KDM5D, KDM6A), and histone-lysine-specific demethylase genes that modulate cancer risk, progression, and response to treatment in other cancers (62 –68).
That normal tissue samples had less sex-biased genes than malignant samples suggested that sex bias might be amplified during tumorigenesis, at least based on mRNA expression data. This study also examined a number of clinically actionable genes, including targets of FDA-approved drugs, and identified a relationship with sex-biased gene expression (69,70). Another study based on existing large cancer databases identified sex dimorphism in somatic mutation profiles, including differences in mutation density and frequency and a correlation to biomarkers of patient outcome (61).
Despite the important insights these studies provide, there are limitations associated with using large databases not designed to study the sex effect, as male and female patient groups for a specific cancer type may have different demographic and tumor characteristics. Another limitation may apply to the analysis of female genes when menopausal status has not been considered. Finally, fully comprehensive insights from the TGCA database are limited by the inclusion of only well-differentiated low- to intermediate risk TC. It is important to incorporate high-risk TC cases to fully understand the basis for sex bias along the severity spectrum.
Because TC has a more aggressive course in males, the inclusion of only low- to moderate risk cases eliminates the opportunity to interrogate the specific role of sex bias in the aggressive nature of the disease in men.
X-inactivation may be a mechanism underlying sex bias in cancers
Sex-biased genes located on the X-chromosome show enrichment for genes that escape X-inactivation, a process postulated to underlie the mechanism for observed sex bias in some cancers (60). X-chromosome inactivation is a newly identified mechanism underlying a possible female advantage in oncological diseases. Theoretically, X-chromosome inactivation in females leads to the presence of a single X-chromosome gene copy and thereby equilibrates cancer risk among males and females with mutations on any X-chromosome-linked genes, including tumor suppressor genes. However, up to 15% of X-linked genes escape X-chromosome inactivation, leading to the presence of a second copy of the gene located on the X-chromosome (71 –73). In addition, about 10% of X-linked genes show a variable pattern of inactivation or a tissue-specific X-chromosome inactivation profile (72,74,75).
X-chromosome inactivation has been implicated in the mechanisms underlying the higher propensity of women to develop autoimmune disorders, including autoimmune thyroid disease (AITD) (76). Skewed X-chromosome inactivation contributes to a strong female predominance in AITD (77 –80) via unequal inactivation of the same X chromosome from either maternal or paternal origin. Consequently, self-antigens on one X chromosome are not expressed at sufficiently high quantities in the thymus or at peripheral immune sites involved in tolerance induction. This loss of immunological tolerance to X-linked antigens is considered to lead to higher prevalence of AITD in females (78 –80).
Further, there is evidence for the contribution of several genes that escape X-inactivation, including CD40L, CD99, IRAK-1, TLR7, and USP27X that are key regulators of cellular and adaptive immune response to female bias in autoimmunity (77,78). Given the known association between AITD and risk of TC, higher female prevalence in TC may be related to underlying causes of female predominance in AITD (80 –82).
In a study of sex bias in >4100 cancers across 21 tumor types, six regions (ATRX, CNKSR2, DDX3X, KDM5C, KDM6A, and MAGEC3) on the X-chromosome were identified with frequent loss-of-function mutations in males. These regions account for tumor suppressors, which, when mutated, lead to a higher incidence of cancers in males. On this basis, it has been suggested that a second functional copy of a tumor suppressor gene in any of these regions might protect females from the negative effects of the mutated copy (60) (Fig. 1).
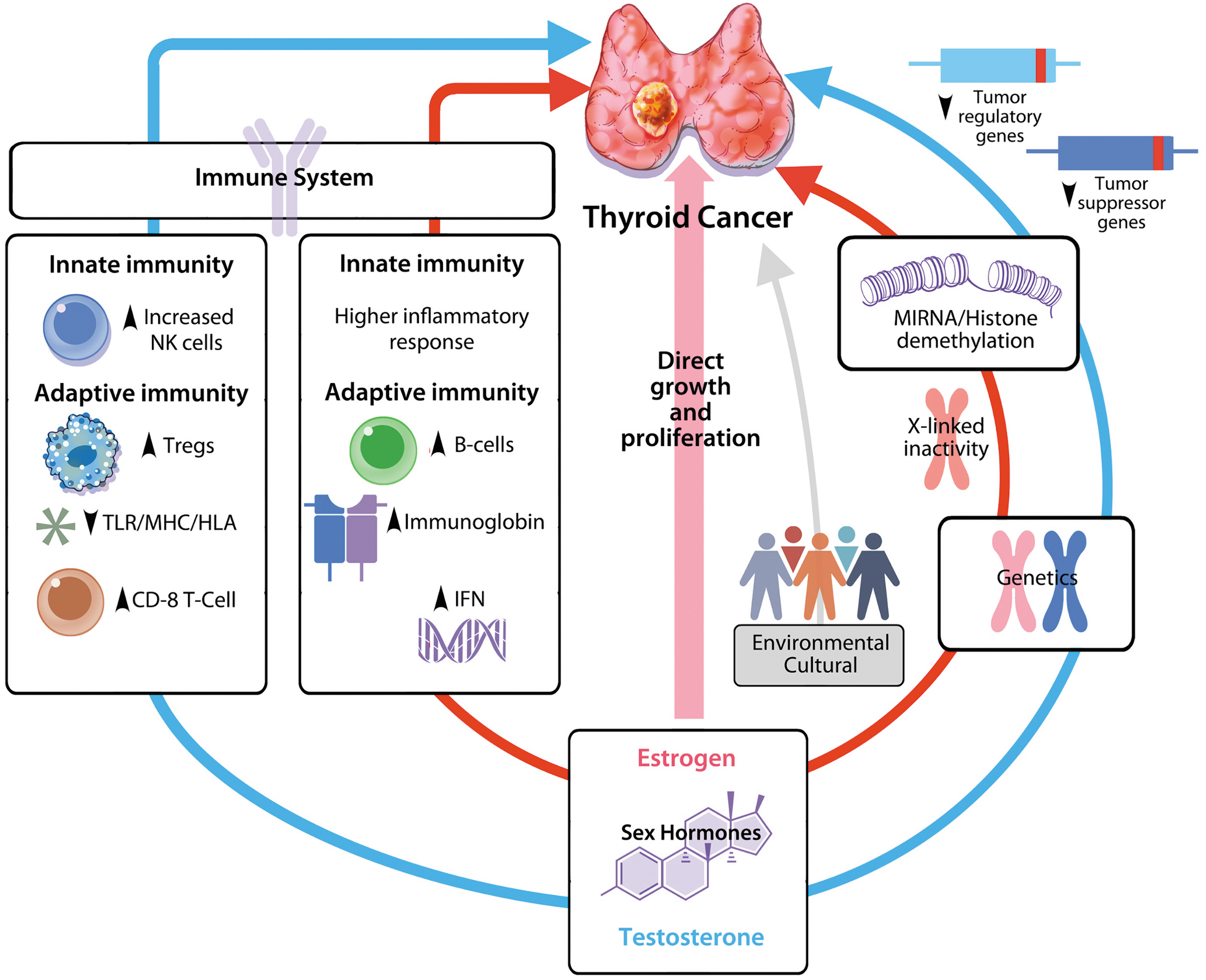
The Figures depict how the risk and progression of TC is likely modulated by an interplay between sex hormones, immune system, genetics, and environmental factors. Sex hormones may modulate the immune response to TC, with estrogen directly promoting growth and proliferation as well as indirectly orchestrating immune response by promoting inflammation and priming of effective immune response against cancer cells in the tumor micro-environment. Testosterone has a less inflammatory and a more immunosuppressive effect likely by modulating the expression level of tumor suppressor and immune regulatory genes that ultimately impair the development of an effective anti-cancer immune response, resulting in larger tumor burden and advanced disease. X-Chromosome inactivation may be an important mechanism underlying genetic basis for sex bias in TC via an extra copy of certain genes (such as genes for tumor regulators), which provide protection in about 10% X-lined genes that escape X-chromosome inactivation in women. Epigenetic regulation may also influence TC development, progression, and response to treatment in a sex-specific manner via miRNAs or histone demethylation, possibly regulating immune response to TC. The roles of environmental and nutritional factors are not well established and include cultural and behavioral components. NK, natural killer; TC, Thyroid cancer; TLR/MHC/HLA, toll-like receptor/major histocompatibility complex/human leukocyte antigen.
Despite evidence for their role in cancer sex bias, X-chromosome inactivation has been frequently either omitted from genome wide association studies (GWAS) or there has been insufficient power to test for the sex-specific impact of genetic variation (83 –85). Among the reasons offered for omitting X-chromosomes in GWAS are the technical difficulties in data analysis using standard array genotyping platforms. For example, given variability in X-chromosome inactivation, it is challenging to discern which genetic variants are on the silenced version of the X-chromosome using standard sequencing technologies given that X-inactivation can be tissue specific (83 –85). Failure to include X-chromosomes in GWAS analysis likely encumbers insights into the molecular basis of sex bias in cancer development, progression, and management.
Epigenetic regulation may sex specifically influence cancer development, progression, and response to treatment
Growing evidence indicates that epigenetic factors play an important role in the risk of cancer development, progression, and response to treatment. MicroRNAs (miRs) are short noncoding RNAs (about 20–22 nucleotides) that influence cellular processes, including proliferation, cell cycle regulation, differentiation, and apoptosis, by their ability to regulate gene expression post-transcriptionally. Dysregulation of miRs-associated cellular signaling circuits can lead to abnormal cell fate decisions and cancer development (86 –88). MiRs can have an important oncogenic function in the development of TC and have been also studied as biomarkers in TC with strong prognostic utility (89 –94).
A larger number of miRs exist on X-chromosomes relative to Y-chromosomes, a difference contributing to greater epigenetic regulation in females (88,95). In cancer, evidence indicates the importance of miRs in the development of immune response in the tumor microenvironment (TME) through PD-L1 receptor-mediated pathways with a role as potential therapeutic targets and prognostic markers (96 –99) (Fig. 1).
Insufficient understanding of the molecular basis of sex dimorphism in cancer incidence and progression underpins the need for high-throughput approaches using GWAS, mRNA-Seq, scRNASeq, ATAC-Seq, and polysome profiling to quantify differences in genetic and epigenetic regulation of sex-specific pathways impacting TC pathogenesis. The four-core mouse model can be used as a potential tool to investigate the sex difference in the pathogenesis, progression, and response to treatment in DTC independent of sex chromosome complement (100 –102).
Contributions of the Immune System
Growing interest and efforts aimed at understanding the contribution of the immune system in TC may relate to recognition of the role of the immune system in tumorigenesis and to discovery of immune-based cancer treatments such as with immune checkpoint blockade (ICB). ICB technologies have revolutionized treatment options for several difficult-to-treat cancers, including nonsmall-cell lung cancer and melanoma. The lack of effective alternative treatment options for advanced, RAIR TC has spurred the search for other therapeutic options such as immune-based strategies.
Thyroid tissue is immunogenic, particularly in the setting of chronic AITD, for example, chronic lymphocytic thyroiditis (Hashimoto's thyroiditis [HT]), a common condition affecting ∼2% of the population (103). In HT, the thyroid is infiltrated by large numbers of immune cells and soluble mediators, including chemokines, cytokines, and growth factors (103 –105). HT has a strong female predominance, with a mean incidence of 3.5–5/1000 in women and 0.6–1.0/1000 in men (106). HT is associated with TC and is found in 20–50% of PTC (107 –109), but no clear causal relationship between the two has been established.
Profiling immune cells immediately surrounding a cancer provide information about the type of immune response induced against cancer. TME represents a complex network of cancer cells, host immune cells, blood vessels, extracellular matrix, and various signaling molecules. The TME has been described as a “battlefield” in which the winner is the side that manages to evade the “barrage” from a complex network of physiological or pathological “artilleries.” Understanding TME biology provides insights into how to successfully intervene in tumorigenesis and/or develop treatment targeting processes and cells within the TME.
Several studies (110 –113) have compared the frequency and distribution of immune cells infiltrating the TME in TC with normal thyroid tissue and described the tumor-driven changes in TC versus normal thyroid. The development of immune responses is strongly sex biased, which adds to the complexity of the relationship between immune responses and TC (114,115). Males are more susceptible to many infectious diseases and cancers of nonreproductive organs, whereas the prevalence and extent of many autoimmune diseases have a striking female bias (114 –117).
Both cellular and humoral immune responses are generally stronger in women, who mount higher antibody responses, generate more effector CD4 T cells, more robust cytokine production and inflammatory responses against infection, as well as enhanced rejection of tumors and allografts (114,118 –121) (Fig. 1). The sex difference in immune response becomes most pronounced in women during reproductive age, with a waning in later life likely reflecting an altered contribution of sex hormones (114).
Estrogen has a proliferative effect on both innate and adaptive immune response and causes increased production of IFNγ and overall cytotoxicity in natural killer (NK) cells (122,123). Estrogen also enhances the expression of pattern recognition receptors, including TLR4, and facilitates the differentiation of bone marrow precursor cells into functional dendritic cells (124 –126). Low estrogen levels promote Th1 type immune responses and cell-mediated immunity, whereas high levels are associated with Th2 type immune responses and humoral immunity across diverse species and cell culture systems (114,127).
Testosterone has general immunosuppressive activity and reduces NK cell activity, expression of TLR4 on macrophages (128,129), and synthesis of TNF, inducible nitric oxide synthase, and nitric oxide production by macrophages (130 –132). Androgens promote differentiation of neutrophils from myeloid progenitor cells, and testosterone deficiency has been associated with neutropenia (133 –135). Testosterone exposure has been associated with downregulation of surface expression of major histocompatibility complex/human leukocyte antigen and costimulatory molecules with impaired inflammatory cytokine production (136,137). In general, males produce lower antibody levels in response to infection and vaccination (138,139) (Fig. 1).
Based on the available evidence for the sex difference in immune response, cancers in women must evade more efficient mechanisms of immune surveillance and undergo a more intensive immune-editing process to overcome host immune response. Cancers that evade a stronger immune surveillance in women are likely less immunogenic, as they must be enriched with stronger mechanisms of immune escape than similar cancers in men. Consequently, cancers in women may be less responsive to immunotherapy compared with comparable cancers in men (140,141).
Recent experimental studies indicate that sex hormones modulate multiple regulatory elements involving PD-1/PDL-1 pathways (142 –146). Meta-analysis of clinical trials involving immune check point inhibitors (ICIs) shows conflicting findings with regard to sex differences and response to immunotherapy. A systematic review of 20 randomized controlled trials of ICIs involving 11,351 patients with advanced cancers indicated that overall survival was significantly better in men (HR 0.72 vs. 0.86, p = 0.0019) (140), findings not supported by other meta-analyses (147 –149).
No sex differences were found in overall survival in another meta-analysis of 23 clinical trials that compared immunotherapy with standard care for advanced solid-organ cancers. And no sex-differences were found (even in subgroup analysis) based on disease site, line of therapy, or class of immunotherapy (147). In another study based on TCGA data, sex difference in overall survival was evaluated with a focus on molecular biomarkers and molecular fingerprints of immune components employing an ICB treatment data set. Significant differences were found in overall survival between sexes for a number of cancers, but the impact of the findings is weakened by the small sample size for individual cancer types (150).
Despite the well-established sex dimorphism in the immune system, sex-specific interrogation of immune response across different reproductive ages in TC has not been performed, although studies describing various immune cell infiltration within TC TME are emerging (110,113,151). Given the central role of the TME in cancer progression and response to treatment, there is a clear need for detailed, sex-specific description of immune landscape of TC TME across different reproductive age groups and cancer histological subtypes.
This could be a critical step toward better understanding the underlying molecular mechanism of sex bias in TC tumorigenesis and to developing sex-specific, targeted, and, by design, more effective treatments. Available studies of immune response in TC mainly are based on immunohistochemistry and RNA expression signatures. Most studies are based on analysis of immune cell distribution in archival tissues, that is, the primary TC, but provide limited evaluation of the physiological immune compartments.
Although this approach provides insight into aspects of the immune response in TC TME, it may not capture all aspects of differences in the complex immune response within other important immune compartments such as normal thyroid, metastatic sites, blood, and lymph nodes. In addition, most current RNA profiling TC research strategies require tissue dissociation, which leads to a loss of morphological and spatial information that provides critical insight into biological activity during tumorigenesis and immune response in the TME.
Cellular proportions, heterogeneity, and deeper spatial distribution of immune cells within the TME provide valuable information about the specific immunological response and serve to identify biomarkers that may predict the success of immune-based therapy (152). By preserving tissue architecture, more recent technologies of multiplex tissue imaging and spatial molecular profiling would provide insight into genetic, transcriptional, and proteomic events, and the spatial organization and interaction of cells.
Despite established evidence for sex bias in immune response, detailed understanding of the contribution of patient sex in mounting an effective immune response in cancer is lacking. Available evidence from preclinical and limited clinical studies points to a pro-inflammatory effect of estrogen and an immunosuppressive nature of testosterone. Women may better mount a more effective immune response against TC and better curb the process of tumorigenesis. Overall, the immunosuppressive effect of testosterone, in particular its effect on reducing NK cell activity and decreasing sensitivity to triggers of inflammation, may promote tumor “escape” from immune surveillance and killing, resulting in larger, more aggressive tumors.
More recent high-resolution cutting-edge technologies may provide sex-specific multi-dimensional characterization of genetic, transcriptional, and proteonomic cellular events while maintaining information about tissue heterogeneity and cell spatial organization within the TME. This will allow the kind of detailed sex-specific immune mapping of TC needed to develop targeted, sex-specific, and, by design, more effective immune therapies for advanced TC. Also needed are prospective, sex-specific immune studies and clinical trials of immunotherapies against the various types of TC that ensure equal sex representation in their preclinical study design. Analysis of data generated from these studies should consider the patient's sex, age, and individual characteristics, including personal, medical, and social influences.
Environmental and Social Factors
Several studies have examined environmental factors, such as nutritional status and exposure to ionizing radiation as potential contributors to TC risk. Effects of exposure to ionizing radiation have not shown sex differences (153 –156). In an updated, pooled analysis of 12 studies evaluating the risk of childhood exposure to external radiation, no significant difference in radiation exposure was found between sexes (p = 0.35) (156). In a subsequent pooled analysis of nine studies, no sex differences were found regarding the consequences of low-dose radiation exposure during childhood although this analysis was not part of the original study design (157). Thyroid nodules have four to five times higher prevalence in females (11).
This sex bias has likely contributed to higher rates of TC in women either through an inherent high risk of TC development in thyroid nodules, or via more rigorous clinical follow-up and imaging in women diagnosed with thyroid nodules, or more likely a combination of both (11,28,158).
Studies of dietary and nutritional factors as potential risk factors for developing TC have not shown sex differences (159 –161). Iodine deficiency has been consistently identified as a risk factor for FTC development, likely through an increased risk of goiter formation and/or thyrotropin stimulation (162 –164). Given the impact of iodine deficiency on reproductive outcomes, there has been a disproportionately higher number of studies in women. Iodine deficiency as a possible risk for TC development has not been vigorously studied in males (159).
Other explanations postulated for sex differences in cancer epidemiology include cultural and behavioral components such that women are more health conscious than men and attend more health appointments, leading to earlier discovery of disease/disease progression and better survival outcomes (165 –168) (Fig. 1). Additional factors influencing individual risk for developing cancer include smoking and obesity, both with higher prevalence in men compared with women (7).
Conclusions and Future Directions
Despite known sex bias in the incidence and progression of TC, its molecular basis remains enigmatic. Fundamental changes in our approach in epidemiology, basic science, and clinical studies are required to effectively interrogate the importance of sex in TC.
An appropriate study design that includes equal representation of sexes, and statistical analysis specific to sex and within appropriate reproductive age groups are critical first steps to generate the epidemiological data needed. Unifying study protocols and standardizing experimental approaches will enable a comparison of results generated by different studies. It will be imperative to equally include both sexes across the spectrum of experimental animal and human studies including in vitro cell models, with reporting of the sex of cell lines/animals. Standards should be created and observed for both the initial funding and subsequent publication of relevant studies to ensure an equal representation of sex of animals or human subjects and that data analysis examines the role of sex.
We need high-throughput approaches, using GWAS, mRNA-Seq, and polysome profiling to better profile and quantify differences in genetic and epigenetic regulation of sex-specific pathways involved in TC. Although recent genetic studies have investigated sex bias using existing large databases, reliance on only these databases may be misleading as these were not specifically designed to study the sex effect. Consequently, the distribution of either male or female patients for specific cancer types may be insufficiently powered.
Hence, there is a need for large-scale prospective studies designed specifically to investigate sex bias in TC. Lastly, sex-specific description of immune cell frequency, distribution, and proportions using cutting-edge spatial molecular techniques holds promise to generate detailed immune mapping needed to better understand the molecular basis for sex bias in TC and promote the design of targeted, sex-specific, and potentially more effective immune-based treatment for advanced TC.
Footnotes
Authors' Contributions
All authors have approved the attached version of the article and have agreed to the submission policies of THYROID. All authors listed have contributed sufficiently to the article to be included as authors.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.