
Abstract
Background:
Multinodular goiter (MNG) is the most common disorder of the thyroid gland. Aging and genetic mutations that impair thyroid hormone (TH) production have been implicated in the development of MNG. XB130 is an adaptor/scaffold protein predominantly expressed in the thyroid gland. XB130 deficiency leads to transient postnatal growth retardation in mice due to congenital hypothyroidism. We studied the formation of MNG and possible mechanisms in elderly mice.
Methods:
Thyroid glands of male and female Xb130-knockout (Xb130−/− ), heterozygous (Xb130+/− ), and wild-type (Xb130+/+ ) mice at the ages of 12–20 months were harvested for visual examination, histopathological, and immunohistological analyses. Blood and thyroid samples were collected after feeding elderly mice with a low iodine diet for 125I uptake and perchlorate discharge assay. The activity of thyroperoxidase (Tpo) was examined by spectrophotometric evaluation of iodide oxidation.
Results:
While moderate MNG was seen in Xb130+/+ and Xb130+/− mice, severe MNG, characterized by multiple nodules intermixed with dilated colloid-rich macrofollicles, was found only in Xb130−/− mice at 18 months. Thyrocyte cytoskeletal structure and cell adhesion molecules were disorganized, and TH production was significantly reduced. Reduced iodide organification was seen in elderly Xb130+/+ mice and further enhanced in Xb130−/− mice. In Xb130+/+ mice, Tpo shows high affinity with hydrogen peroxide (H2O2) throughout aging, but reduced affinity with iodide in an age-dependent manner. By contrast, in elderly Xb130−/− mice, the affinity of Tpo for iodide remained high, but the affinity of Tpo for H2O2 was reduced.
Conclusions:
The pathophysiological features in the thyroid glands of aged Xb130−/− mice closely resemble the features of MNG in humans. Moderate MNG in elderly mice was dramatically aggravated by XB130 deficiency. Reduced affinity of Tpo for H2O2 may contribute to MNG development via oxidative stress. This could be specific to XB130 deficiency but also could be a common mechanism in MNG. Its clinical relevance should be further investigated.
Introduction
Multinodular goiter (MNG) is the most frequent disorder of the thyroid gland (1). Low production of thyroid hormone (TH) induces thyrotropin (TSH) secretion and subsequently global and focal follicular cell proliferation, which causes diffuse thyroid enlargement; prolonged hyperplasia ultimately undergoes focal degeneration and results in nodule formation (1,2). Thyroid dyshormonogenesis due to genetic defects in the proteins regulating TH biosynthesis can cause hypothyroidism leading to thyroid hyperplasia (3). Disruption of genes related to cellular and molecular functions of thyrocytes can also result in MNG. For example, overexpression of a thyroid-specific transcription factor Foxe1 in the thyroid leads to the development of MNG in mice (4).
XB130 (actin filament-associated protein 1-like 2, AFAP1L2) is an adaptor protein predominantly expressed in the thyroid gland (5 –7). Adult Xb130-knockout (Xb130−/− ) mice exhibit normal physiology in general, and their life span is comparable to that of wild-type (Xb130+/+ ) mice (8). However, we serendipitously discovered transient growth retardation of Xb130−/− mice at the early postnatal stage due to congenital hypothyroidism. Enlarged thyroid glands with delayed folliculogenesis and reduced thyroglobulin (Tg) iodination and release into the follicle were found in Xb130−/− mice (9).
Nontoxic and toxic MNG are the main etiologies of goiter in patients aged 55 years and older (10). Mice have been widely used for aging studies, as it is cost-effective, serving 99% of human genes and physiologically resembling humans (11). It is estimated that 1-year-old mice are about 58 years of human age, and 2-year-old mice are about 70 years of human age (12). We found enlarged thyroid glands with multiple nodules in 18-month-old Xb130−/− mice.
Thyroperoxidase (Tpo) is a thyroid-specific glycosylated hemoprotein that, using hydrogen peroxide (H2O2) as oxidative equivalent, catalyzes the oxidation of iodide and coupling of iodotyrosine residues in Tg (13). We report that the kinetic characteristics of Tpo in iodide oxidation reaction are altered with aging, which may contribute to the pathogenesis of MNG in Xb130−/− mice.
Materials and Methods
Please see detailed methods in Supplementary Data.
Animals
Xb130−/− mice were generated as previously described (8). Animal procedures were approved by the Animal Care and Use Committee of the University Health Network (Toronto, Canada).
Histopathological and immunohistochemical analyses
Harvested mouse thyroid tissues were fixed in 10% neutral buffered formalin for 48 hours at room temperature. Hematoxylin and eosin (H&E) staining, Masson's trichrome staining, immunohistochemistry staining, and immunofluorescence staining were performed following standard protocols.
Thyroid function tests
Whole blood was collected from isoflurane-anesthetized mice by cardiac puncture. Serum total thyroxine (T4), triiodothyronine (T3), and TSH concentrations were measured by radioimmunoassay (14). T4 and T3 bound to Tg (Tg-T3 and Tg-T4) and non-Tg-bound T3 and T4 (non-Tg-T3 and non-Tg-T4) were measured as previously described (14).
125I uptake and perchlorate discharge test
Mice fed with a low iodine diet (LID; <0.05 ppm iodine; Harlan Teklad Co., Madison, WI) for 2 weeks were injected intraperitoneally with 125I (8 μCi/mouse) (15). 125I uptake and perchlorate discharge test was performed as described (9).
Quantitative RT-qPCR
The protocol for quantitative reverse transcriptase PCR (RT-qPCR) has been described (9). The expression levels of messenger RNA (mRNA) were normalized with a housekeeping gene, glyceraldehyde 3-phosphate dehydrogenase (GAPDH). Primer sequences are available in Supplementary Table S1.
Western blot
Standard Western blotting protocol was followed (8,16). Mouse antibodies against Tg (1:5000), iodinated (I)-Tg (1:5000) (15), and Tpo (1:1000; Santa Cruz Biotechnology), as well as the anti-mouse secondary antibody (1:10,000; Abcam, Cambridge, MA), were used.
Tpo activity assay
Iodide oxidation activity of Tpo was measured (17). Data were analyzed using the Michaelis–Menten equation and SigmaPlot 10 (Systat Software, Inc., San Jose, CA).
Statistical analysis
All values are expressed as mean ± standard error of the mean. Most data were analyzed through two-way analysis of variance with Bonferroni test as post hoc analysis. The expression of genes related to TH synthesis was analyzed by unpaired t-test. p-Values <0.05 are considered statistically significant.
Results
Enlarged thyroid gland of aged Xb130−/− mice with multiple nodules
Full-body necropsy of 18-month-old mice (M18) showed distinct variability of the size of the thyroid glands (Fig. 1A). The average weight of thyroid glands of Xb130−/− mice was ∼11-fold greater than that of Xb130+/+ mice and 5-fold greater than that of Xb130+/− mice (p < 0.001, Fig. 1B), despite their similarity in body weight (Fig. 1C) and similar incidence of overall malignancy (Supplementary Fig. S1).

Enlarged thyroid glands of aged Xb130−/−
mice. (
The enlarged thyroid glands showed pathological features resembling those of human MNG. Normal thyroid glands with regular-sized follicles (Fig. 1D.i, ii) were observed in 91.1% of Xb130+/+ mice and 62.9% of Xb130+/− mice (Fig. 1E). A moderate form of MNG (diffusely enlarged goiter containing a few hyperplastic cellular nodules with large colloid-filled follicles, as shown in Fig. 1D.iii, iv) was found in 37.1% of Xb130+/− mice and 36.4% of Xb130−/− mice (Fig. 1E). A severe form of MNG with multiple nodules, severely deformed follicular architecture with enlarged complex colloid-rich nodules admixed with hyperplastic cellular areas (Fig. 1D.v, vi), was found in 45.5% of Xb130−/− mice (Fig. 1E). The incidence of MNG in thyroid glands of Xb130−/− mice was significantly higher than that in age-matched Xb130+/+ or Xb130+/− mice (Fig. 1E). The severe form of MNG in Xb130−/− mice were seen in both sexes.
Pathological characteristics of MNG in the thyroid glands of Xb130−/− mice
The severe form of MNG in Xb130−/− mice displayed a variety of pathological characteristics. In contrast to normal thyroid glands containing uniform-sized follicles lined by a single layer of cuboidal epithelial cells (Fig. 2A.i), MNG found in Xb130−/− mice was characterized by heterogeneous follicles of variable sizes (Fig. 2A.ii). Large colloid-rich macrofollicles were frequently lined by thin flattened epithelium (Fig. 2A.iii), whereas tall columnar epithelial cells were observed in nodules composed of microfollicles with pseudopapillary hyperplasia (Fig. 2A.iv).
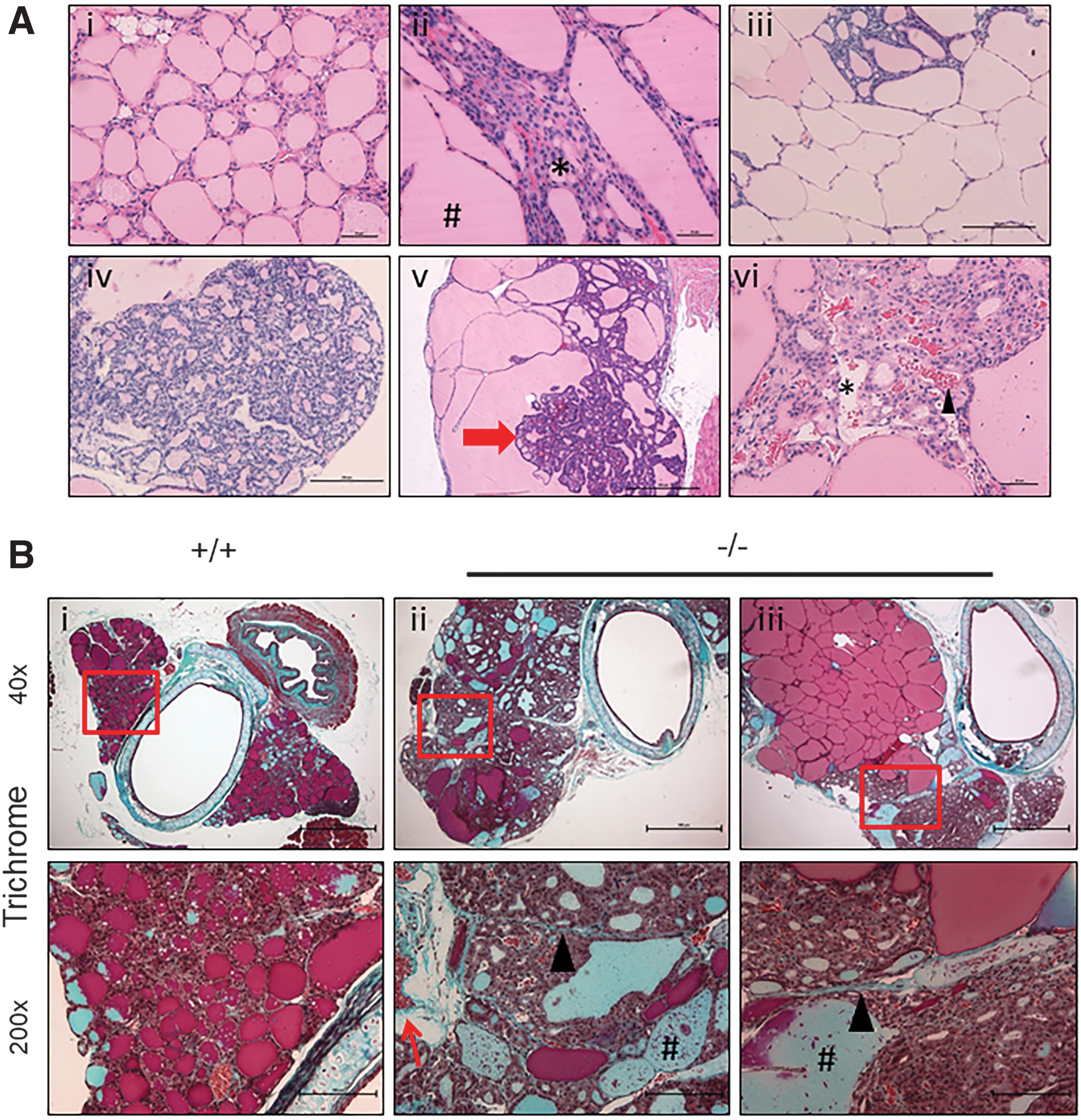
Pathological characteristics of the thyroid glands of aged Xb130−/−
mice. (
In many cases, markedly enlarged thyroid glands of Xb130−/− mice contained large colloid-filled nodules with small papillary infoldings protruding into the lumen. Small subfollicles, known as Sanderson's pollsters, protruded into the colloid of larger follicles. Throughout these thyroid glands, there was no evidence of nuclear features of papillary carcinoma (Fig. 2A.v). The thyroid glands of Xb130−/− mice showed focal hemorrhage in more cellular areas of the MNG (Fig. 2A.vi).
Masson's trichrome staining showed little or no fibrosis with acidophilic dense colloid in normal thyroid glands of Xb130+/+ mice (Fig. 2B.i). The enlarged thyroid glands of Xb130−/− mice had areas of fibrosis and hyperplastic papillary nodules with basophilic diluted colloid (Fig. 2B.ii, iii). Acidophilic dense follicles were found in colloid-rich areas of Xb130−/− thyroid glands (Fig. 2B.iii), and many of the follicles contained dead cell debris within the colloid (Fig. 2B.ii, iii). The variable colloid storage and heterogeneity of follicle size are typical features of MNG, in which fibrosis is often associated with hemorrhage and degeneration (18).
Disrupted cytoskeleton organization in the thyroid glands of aged Xb130−/− mice
XB130 was mainly localized at the apical membrane of thyrocytes in thyroid glands of Xb130+/+ mice, forming a ring-like structure along the colloid lumen (Fig. 3A). Proper thyroid follicle structure is essential for TH synthesis (19), and follicular architecture is maintained by the integrity of cytoskeleton (20). Actin filaments in the normal thyroid glands of Xb130+/+ mice, as shown by phalloidin staining, were predominantly near the apical membrane of thyrocytes, displaying ring-like staining, similar to that of XB130 (Fig. 3B).
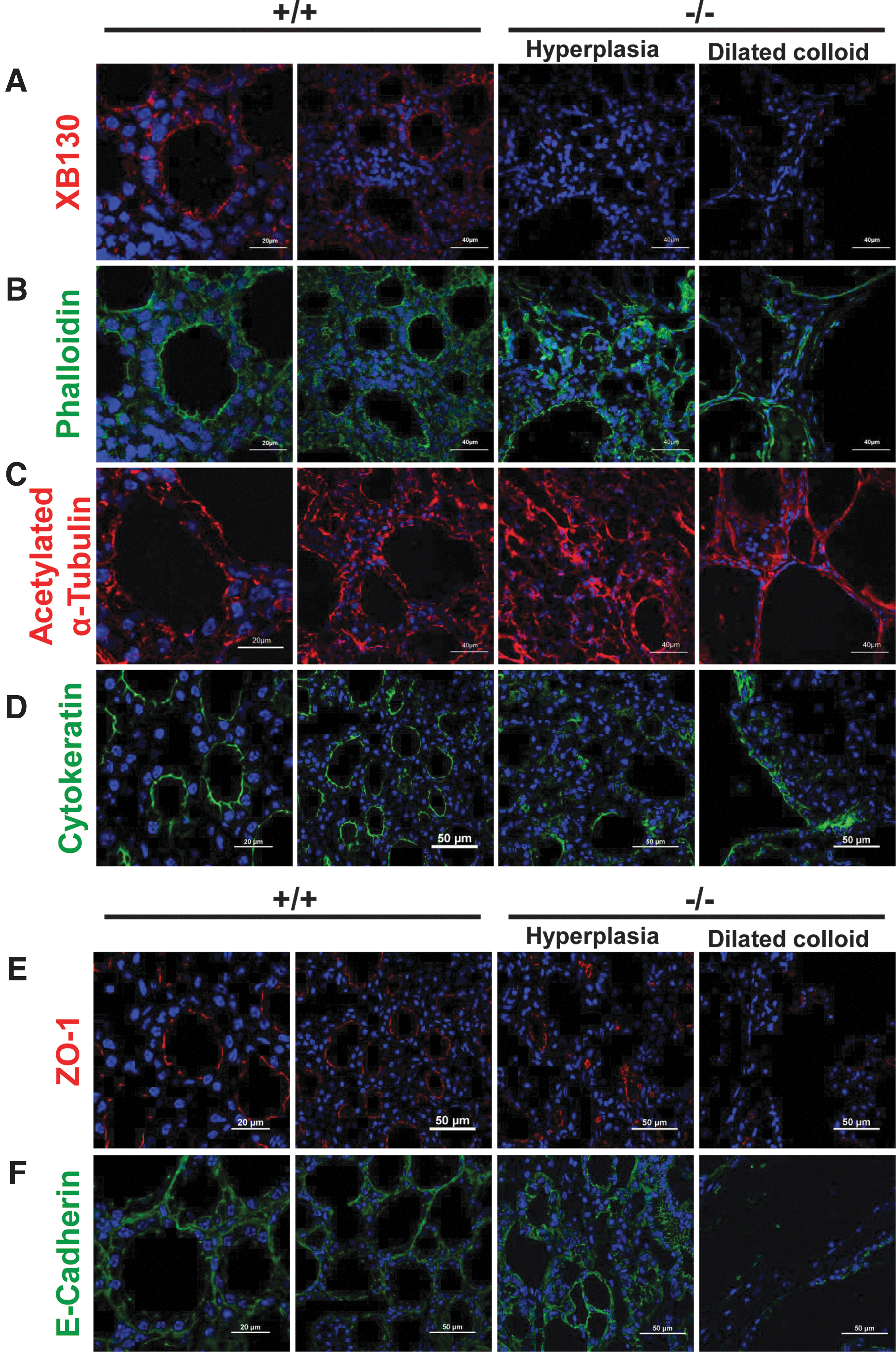
Disrupted cytoskeletal organization and epithelial junctions in thyroid follicles of Xb130−/−
mice at M18. Immunofluorescent staining of (
In contrast, phalloidin staining in the thyrocytes of Xb130−/− mice was dispersed in the cytoplasm of hyperplastic epithelium, lacking the ring-like structure around the apical membrane. In the flattened epithelium of dilated follicles, deformed architecture was evident (Fig. 3B). Similarly, microtubules and intermediate filaments, shown as acetylated α-tubulin and cytokeratin, respectively, were irregularly distributed in the thyrocytes of Xb130−/− mice, in contrast to their uniform expression along the apical membrane of the thyrocytes of Xb130+/+ mice (Fig. 3C, D).
Tight junctions and adherens junctions are critical to maintain thyroid epithelial cell polarization (21). In Xb130+/+ mice, zonula occludens-1 (ZO-1), a tight junction-associated protein, was expressed apicolaterally in thyrocytes (Fig. 3E). The adherens junction protein E-cadherin was predominantly expressed basolaterally in the thyrocytes of Xb130+/+ mice (Fig. 3F). In Xb130−/− mice, hyperplastic follicles exhibit an inconsistent distribution of both ZO-1 and E-cadherin, and they were nearly absent in dilated colloid-filled follicles (Fig. 3E, F).
Chronic hypothyroidism in Xb130−/− mice
To examine the effects of the observed MNG pathology on TH production, we assessed serum levels of TSH, T4, and T3 by radioimmunoassay. Xb130−/− mice at month 12 (M12) had significantly elevated circulating TSH with lower T4 levels compared with those in Xb130+/+ mice (Fig. 4A, B). The serum T4 levels in M18 Xb130+/+ mice were lower than those at M12 (Fig. 4B), associated with significantly higher TSH levels (Fig. 4A), indicating aging-related hypothyroidism. Serum TSH and TH levels in M18 Xb130−/− mice were comparable to those of Xb130+/+ mice (Fig. 4A, B).
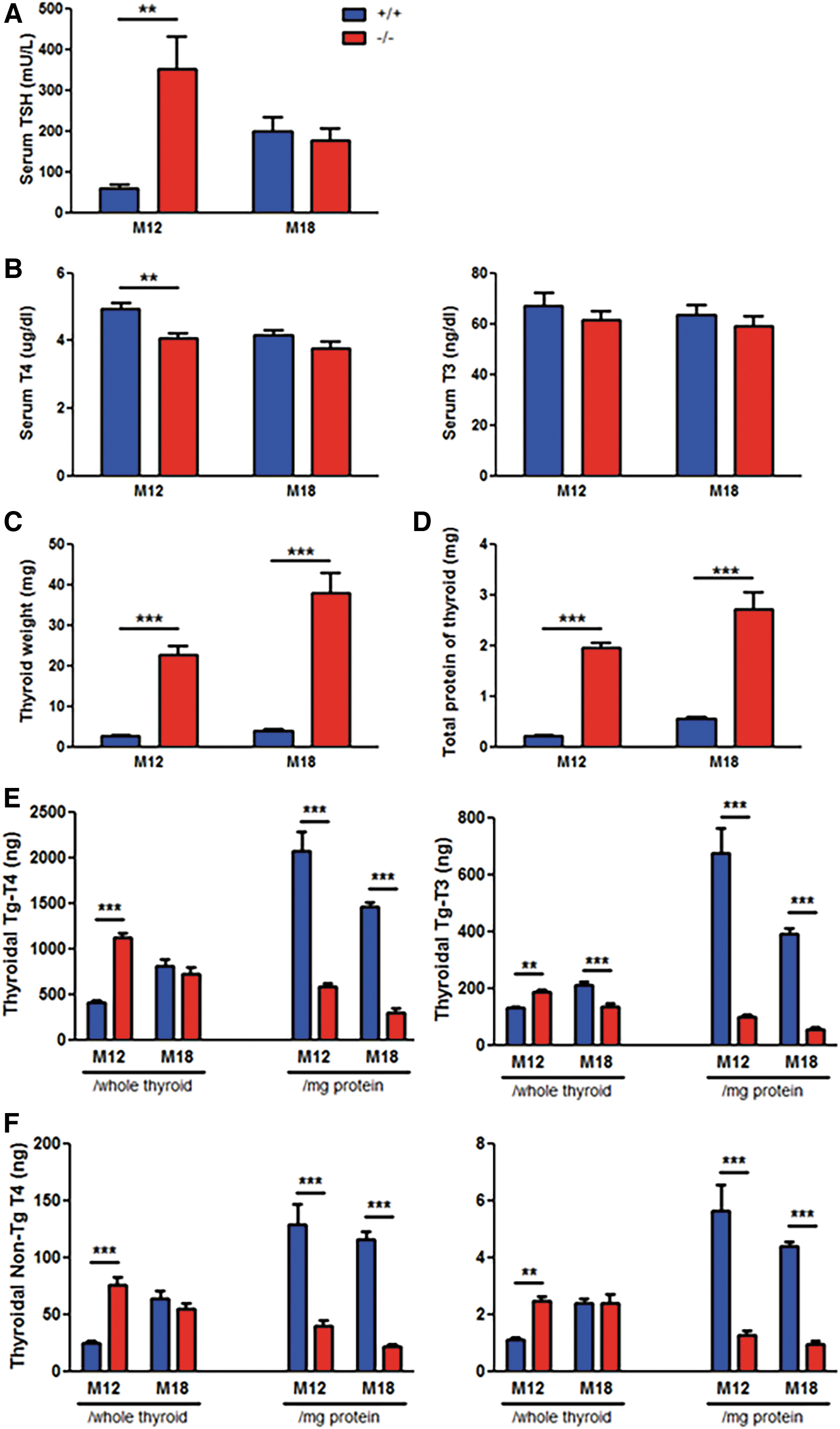
Xb130−/−
mice showed hypothyroidism at month 12 (M12) and euthyroid at M18. (
The average thyroid weight (Fig. 4C) and total protein per thyroid (Fig. 4D) were significantly higher in Xb130−/− mice at both M12 and M18. Therefore, thyroidal TH levels were expressed as either per thyroid or per mg protein. At M12, free TH (non-Tg-T4 and non-Tg-T3) and TH bound to Tg (Tg-T4 and Tg-T3), when expressed as per thyroid, were all significantly higher in thyroid glands of Xb130−/− mice (Fig. 4E, F), yet their serum T4 level was significantly lower (Fig. 4B), suggesting a defect in secretion of TH. When the amounts of thyroidal TH were expressed as per mg protein, these levels were markedly less in the Xb130−/− group at both M12 and M18 (Fig. 4E, F), indicating reduced TH production in the thyroid glands. Moreover, these levels at M18 were lower than those at M12 in Xb130+/+ mice (Fig. 4E, F), further supporting an age-related decrease in TH production.
Compensatory responses of Xb130−/− mice to restore thyroid homeostasis
We assessed some of the genes regulated by TSH through the cAMP pathway (22) with RT-qPCR. Expression of Tshr, Nis, Pendrin, DuoxA1, and Tpo in the thyroid glands of Xb130−/− mice was significantly upregulated (Fig. 5). Expression of Tg, Duox1, Duox2, and DuoxA2 was similar in Xb130+/+ and XB130−/− mice (Fig. 5). This profile shows compensatory responses to the defect of TH production.
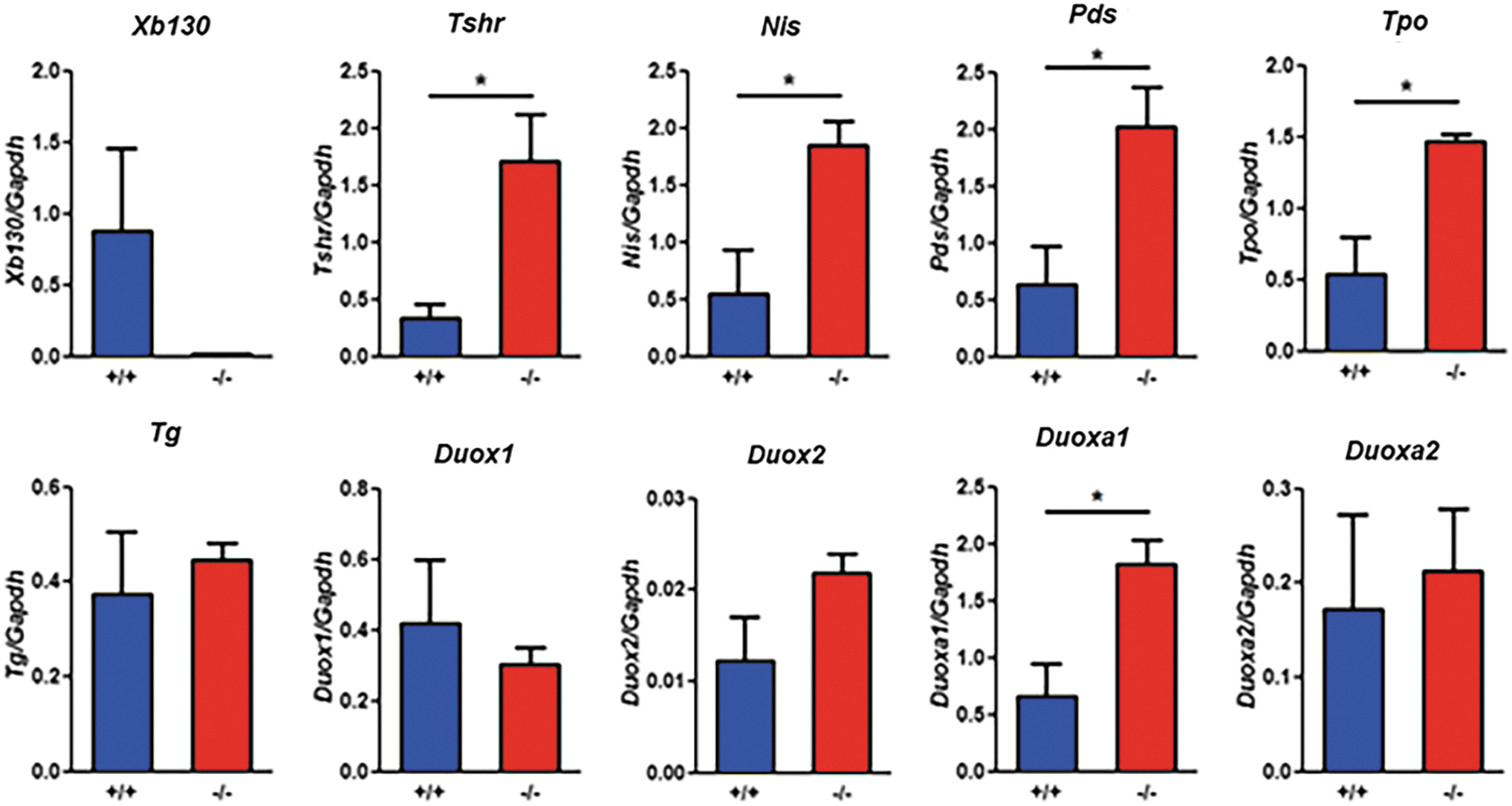
Compensatory responses in gene expression in the thyroid glands of mice at M18. Thyroidal expression of selected thyroid hormone synthesis-related genes in Xb130+/+ and Xb130−/− mice at M18, determined by RT-qPCR and normalized with GAPDH. Expression of Xb130, Tshr, Nis, Pds, Tpo, and Duoxa1 genes was significantly elevated in the Xb130−/− group. *p < 0.05, n = 3 per group. GAPDH, glyceraldehyde 3-phosphate dehydrogenase; RT-qPCR, quantitative reverse transcriptase PCR; Tpo, thyroperoxidase. Color images are available online.
Impaired iodide organification in aged Xb130−/− mice
To determine whether impaired TH production is due to defective iodide uptake and/or organification, we performed the 125I uptake and perchlorate discharge test in mice at M18 (Fig. 6A). Two weeks of LID reduced serum T4 and T3 levels in both genotypes (Fig. 6B.i, ii), with elevated TSH, especially in Xb130−/− mice (Fig. 6B.iii). High amounts of 125I per thyroid were found in Xb130−/− mice (Fig. 6C.i), but the 125I per mg of thyroid weight was comparable between the two genotypes (Fig. 6C.ii).

Defect in iodide organification in aged Xb130−/−
mice. (
After animals were treated with ClO4 − , the thyroidal 125I levels in Xb130−/− mice were significantly lower than those in Xb130+/+ mice (Fig. 6C.i, ii). Blood levels of 125I were significantly increased in both genotypes (Fig. 6D.i), and levels in Xb130−/− mice were significantly higher than those in Xb130+/+ mice (Fig. 6D.ii). The impairment of iodide organification in thyroid glands of Xb130−/− mice was further supported by reduced iodinated Tg in the thyroid glands, despite a higher Tg content revealed by Western blot (Fig. 6E).
Altered Tpo activity and binding affinity with its substrates in Xb130−/− mice
To determine if the impaired iodide organification is due to the change of Tpo activity, we measured iodide oxidation reaction with tissue lysates from thyroid glands. Iodide oxidation reaction catalyzed by Tpo in tissue lysates reaches an Abs353 peak, followed by a gradual decline (Fig. 7A). The rates of reaction were determined from the initial linear phase of the reaction curve. Surprisingly, they were markedly higher in Xb130−/− mice, especially in aged ones, than those in age-matched Xb130+/+ mice (Fig. 7B). This increase in total Tpo peroxidase activity in Xb130−/− mice was associated with higher Tpo protein expression (Fig. 7C) and significantly higher Tpo mRNA levels (Fig. 5).
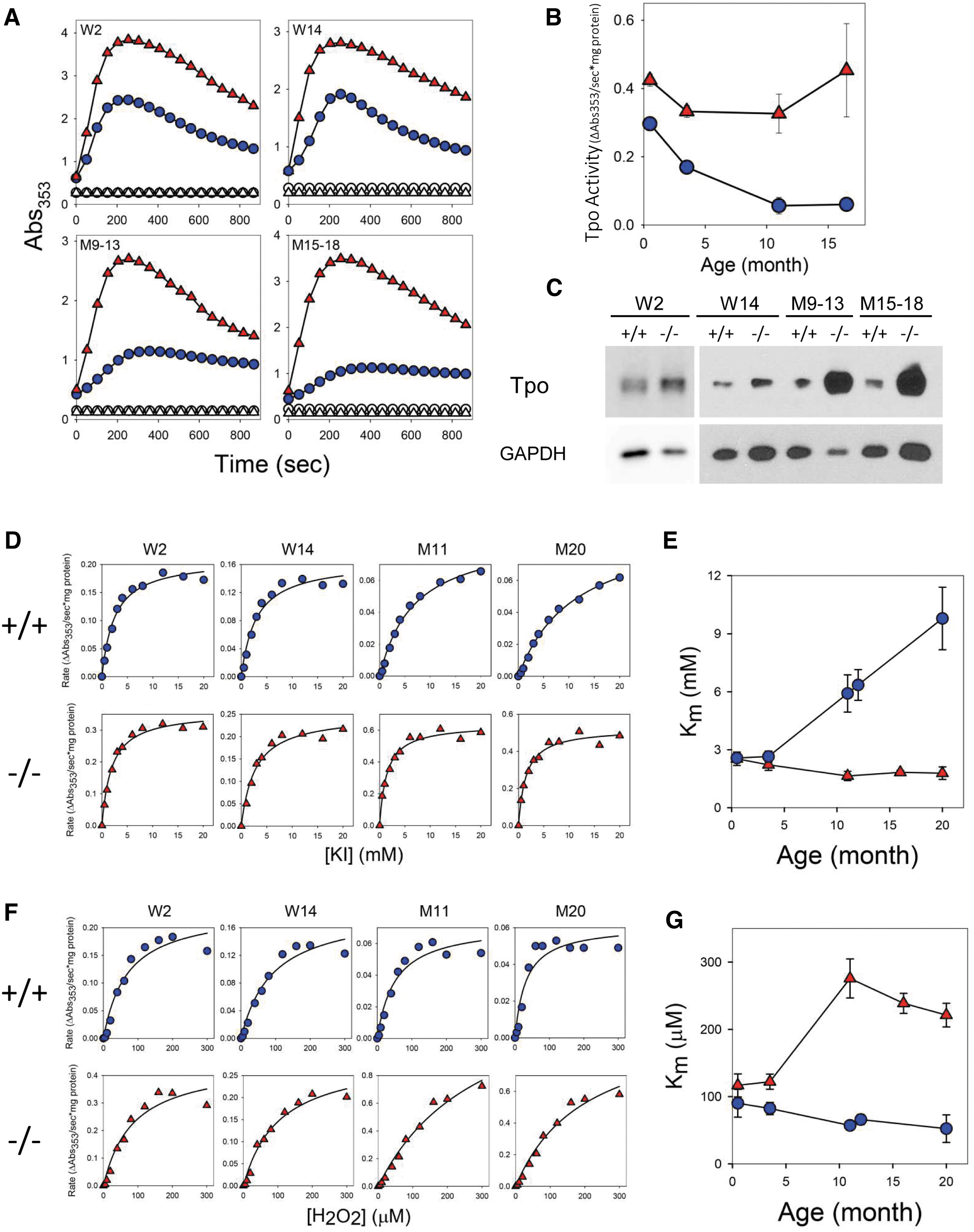
Altered Tpo activity in Xb130−/−
mice. (
We further studied the affinity of Tpo with its substrates, iodide and H2O2. Initial rates of the iodide oxidation reaction increase in hyperbolic manner as a function of iodide concentration in both genotypes (Fig. 7D). Tpo in Xb130+/+ mice at week 2 (W2) and week 14 (W14) exhibited a Km of ∼3.2 mM with a Vmax of ∼0.18 seconds −1 (mg of protein)−1 (Fig. 7E). The value of Km markedly increased with age, ranging to ∼12.0 mM at month 20 (M20), showing that in Xb130+/+ mice Tpo exhibited a reduction in iodide affinity with aging. The Km of Tpo for iodide in Xb130−/− mice at W2 and W14 was comparable to that in age-matched Xb130+/+ mice, however, it remained at similar levels through aging (Fig. 7E), demonstrating that the affinity of Tpo for iodide remains high in Xb130−/− mice.
By contrast, the affinity of Tpo for H2O2 remains relatively high in Xb130+/+ mice from W2 to M20, with a Km of ∼110 μM and a Vmax of ∼0.20 seconds −1 (mg of protein)−1 (Fig. 7F, G), indicating their efficient use of H2O2 as an oxidative equivalent. Although Tpo in Xb130−/− mice at W2 and W14 also exhibited relatively high affinity for H2O2, an elevation in Km to ∼300 μM at month 11 (M11) to M20 indicated a reduction in its affinity for H2O2 with aging (Fig. 7G). The lower binding affinity between Tpo and H2O2 may affect clearance of H2O2 in the thyroid glands.
Discussion
MNG is associated with chronic iodine deficiency and other exogenous factors such as natural goitrogens, smoking, and lack of selenium and iron (23). However, intrinsic glandular defects by altered genetic factors also result in MNG (23 –26). XB130 is an adaptor/scaffold protein enriched in thyrocytes and involved in maintaining polarity and apical membrane structure and function of thyrocytes and thus involved in TH production and secretion (9). Here, we show that XB130 deficiency enhances the susceptibility to MNG in elderly mice, and the pathophysiological characteristics of severe MNG are comparable to those in humans. We found opposite changes of Tpo affinity with iodide and H2O2 between Xb130+/+ and Xb130−/− mice, which may contribute to the pathogenesis of MNG through oxidative stress.
Chronic hypothyroidism causes MNG in Xb130−/− mice
We have recently shown that Xb130−/− mice exhibit transient postnatal growth retardation due to congenital hypothyroidism (9). With the enlargement of thyroid glands, these animals showed fully compensated physiological function (8). In young adulthood, XB130 deficiency enhances lipopolysaccharide-induced septic response and acute lung injury (27), or carcinogen-induced skin tumorigenesis (28). These defects may reflect the local role of XB130 in the lung or skin. In the present study, we found that the thyroid glands of elderly Xb130−/− mice develop pathological changes similar to MNG in humans. Since moderate MNG pathology was also noted in wild-type and heterozygous mice, the lack of XB130 may enhance aging-related MNG formation.
XB130 and cytoskeletal structure in thyrocytes
The major function of XB130 in the thyroid glands is to maintain the polarity of thyrocytes and its apical membrane structure and function, through mediating interactions between microfilament and microtubule systems in thyrocytes (29). Lack of XB130 delayed folliculogenesis, reduced microvilli, and inhibited recruitment of Tpo to the apical membrane (9). In the present study, we also found disorganized actin filaments, acetylated α-tubulin, and cytokeratin in the thyrocytes of Xb130 −/− mice, as well as disorganized cell adhesion molecules (ZO-1 and E-cadherin).
However, hyperplasia, fibrosis, and accumulation of colloid in the follicles may aggravate the already altered thyrocyte architecture and polarity. Therefore, these cytoskeletal structural changes could be both the causes and consequences of MNG. Kelch-like ECH-associated protein 1 (Keap1) is a negative regulator of the nuclear factor erythroid 2-related factor 2 (Nrf2) antioxidant response pathway. Knockdown of Keap1 induced age-dependent diffuse goiter in mice (30). Keap1 is associated with the actin cytoskeleton, and disruption of the actin cytoskeleton is seen in thyrocytes of Xb130−/− mice. It would be interesting to determine if the Keap1/Nrf2 antioxidant response pathway is activated in the thyroid glands of Xb130−/− mice.
Aging-related hypothyroidism in mice
We noted that thyroidal TH decreased with aging in both genotypes, when expressed as per mg of protein (Fig. 4E, F). In thyroid function tests, after 2 weeks of LID, the serum T4 and T3 levels decreased in both genotypes but remained in measurable ranges (Fig. 6B), while in young adult mice, the LID protocol reduced serum levels to nearly undetectable in Xb130+/+ mice (9). Moreover, in the present study, the leakage of 125I from thyroid glands to blood was seen in both genotypes after ClO4 − treatment, indicating reduced iodide organification in both genotypes, even though it was more severe in Xb130−/− mice (Fig. 6D). These observations suggest that reduced TH production and reduced iodide organification are the consequence of aging, seen in the elderly, and XB130 deficiency enhances or accelerates these processes.
Altered kinetic properties of Tpo in MNG of elderly Xb130−/− mice
Tpo is a membrane-bound glycoprotein located at the apical membrane of the thyrocytes. Using H2O2 as an oxidative equivalent, the protein catalyzes oxidation of iodide and coupling of tyrosyl residues, which are crucial steps in the TH biosynthesis. To compensate for their hypothyroidism, Xb130−/− mice exhibit upregulated expression of Tpo at both gene and protein levels (Figs. 5 and 7C). However, the iodide organification (Fig. 6) and the production of TH are significantly lower (Fig. 4).
Notably, the affinity of Tpo for iodide is high in Xb130+/+ mice at younger ages, denoted by its low Km, which is likely because mice require more TH for physical growth. With aging, the affinity of Tpo for iodide is reduced, which likely allows the thyroid follicles to accumulate additional iodide for storage and also prevents the overproduction of TH. By contrast, Tpo in Xb130−/− mice retains a high affinity for iodide throughout life, continuously promoting TH production. This suggests that Xb130−/− mice, and possibly humans with MNG, are predisposed to hyperfunction of Tpo in thyrocytes.
Interestingly, while the Tpo affinity for H2O2 in Xb130+/+ mice remains relatively high throughout life, it decreases with age in Xb130−/− mice. Tpo activity decreases with increasing H2O2 level (29); thus, an adequate, possibly very low level of H2O2 is needed for optimal Tpo activity. A decrease in the affinity of Tpo for H2O2 in aged Xb130−/− mice may be an adaptive response to promote TH production by maximizing Tpo activity. This reduced affinity also implies inefficient utilization of H2O2 in the thyroid gland. H2O2 is not only a substrate of Tpo but also a major source of reactive oxygen species.
Accumulated H2O2 likely contributes to inflammation, cytoskeletal changes, fibrosis, damage in genomic DNA, and increase in spontaneous mutation, which may result in nodular transformation in the thyroid gland (30). Therefore, efficient clearance of H2O2 by Tpo is crucial in maintaining thyroid homeostasis (31 –33). The expression of oxidative stress-related genes and proteins and their functions should be further studied. The changes of affinity of Tpo with its substrates are only seen in elderly mice; therefore, we speculated that this could be a common mechanism for the pathogenesis of MNG caused by chronic hypothyroidism during aging, which should be further studied clinically.
This is the first report on the role of XB130 in MNG. More mechanistic studies are required, and the clinical relevance of XB130 in humans, especially, in the formation of MNG should be further studied.
Footnotes
Acknowledgments
We would like to thank the Animal Resource Centre facility staffs for their help on mice work. We would like to also thank Zehong Guan for her precious advice on immunostaining and Dr. Susan Newbigging for histological studies. Finally, we would like to thank Deborah Scollard and Napoleon Law in the STTARR (Spatio-Temporal Targeting and Amplification of Radiation Response) program for their help on radioisotope studies and tissue processing.
Authors' Contributions
H.-R.C. and M.L. designed the project. J.S. designed and performed Tpo-related experiments. H.-R.C. participated in most of the experiments. H.-R.C. and H.S. designed and performed thyroid functional studies. Y.-Y.X. participated in immunofluorescent studies. X.B. performed RT-qPCR. Y.W. participated in some of the experiments. X.-H.L. performed serum and thyroid gland T4 and T3 determinations and serum TSH measurements. H.-R.C., J.S., and M.L. wrote the article. S.L.A. and S.R. provided conceptual advice, planned some of the experiments, and edited the article. All co-authors have approved the article for submission.
Author Disclosure Statement
No potential competing interests are disclosed.
Funding Information
Canadian Institutes of Health Research Operating Grants (MOP-13270, MOP-42546, and MOP-119514) supported the work performed in this study. H.-R.C. is supported by scholarships from the Department of Physiology in the University of Toronto: Dr. Albert and Doris Fields Graduate Scholarship in Cardiovascular Physiology and Doctoral Completion Award. J.S. is supported by Toronto General Hospital Research Institute postdoctoral fellowship. X.-H.L. and S.R. are supported in part by grant DK15070 from the National Institutes of Health.
Supplementary Material
Supplementary Data
Supplementary Figure S1
Supplementary Table S1