
Abstract
Background:
The human adrenal cortex undergoes several rapid remodeling steps during its lifetime. In rodents, similar remodeling occurs postnatally in the “X-zone” layer through unknown mechanisms. Furthermore, little is known regarding the impact of thyroid hormone (TH) on adrenal glands in humans.
Methods:
To investigate the impact of TH on adrenal pathophysiology, we created two genetic murine models mimicking human nonautoimmune hypothyroidism and hyperthyroidism. Moreover, we analyzed serum thyrotropin (TSH) and steroid hormone concentrations in patients diagnosed with congenital hypothyroidism and premature adrenarche (PA).
Results:
We found that TH receptor beta-mediated hypertrophy of the X-zone significantly elevated the adrenal weights of hyperthyroid women. In the hypothyroid model, the X-zone was poorly developed in both sexes. Moreover, large reciprocal changes in the expression levels of genes that regulate adrenal cortical function were observed with both models. Unexpectedly, up- and downregulation of several genes involved in catecholamine synthesis were detected in the adrenal glands of the hypothyroid and hyperthyroid models, respectively. Furthermore, TSH and adrenal steroid concentrations correlated positively in pediatric patients with congenital hypothyroidism and PA.
Conclusions:
Our results revealed that congenital hypothyroidism and hyperthyroidism functionally affect adrenal gland development and related steroidogenic activity, as well as the adrenal medulla.
Introduction
Thyroid hormones (THs) play an essential roles in the development, growth, and metabolism (1,2). Abnormal TH concentrations in early life may disturb bone, lipid, and energy metabolism, or neuronal and lung development (3 –7). Furthermore, a correlation between THs and adrenal steroids in patients with congenital hypothyroidism (CH) were recently reported (8); however, the pathophysiological role of THs in the adrenal glands are unclear.
To date, the relationship between the thyroid and adrenal glands has been mainly demonstrated by the pharmacological administration of THs or by investigating their roles in adult adrenals, but the data from more physiological models are lacking. In rodents, the administering THs increased adrenal weights (9,10), whereas adrenal weights decreased in hypothyroid mice (11). Furthermore, THs can also have an impact on the adrenal–pituitary axis, as thyroidectomized rats present with decreased adrenocorticotropic hormone (ACTH) concentrations (12). Recently, a direct adrenal effect of TH was reported to be mediated through thyroid hormone receptor-β1 (THRβ1) expressed in the murine X-zone (13). Furthermore, Thrb
In a recent study conducted on human neonates with severe CH, higher serum concentrations of 17-hydroxyprogesterone (17-OHP), dehydroepiandrosterone sulfate, Δ4-androstenedione, and testosterone were detected, compared with euthyroid controls (8). This finding suggests that subnormal TH concentrations at birth may decelerate the involution of the fetal zone. The responses of adrenal glands to ectopic thyrotropin (TSH) receptor expression or TSH-releasing hormone have rarely been reported (14,15), and their physiological impacts remain unknown.
Based on these recent observations regarding the role of THs in neonatal adrenal development, we explored the molecular mechanisms underlying adrenal gland development and function in two mouse models of thyroid disease (neonatal hypothyroidism and hyperthyroidism). Furthermore, to enhance the translational relevance of this study, postnatal TSH and adrenal steroid concentrations were assessed in pediatric patients with premature adrenarche (PA), CH, or hyperthyroidism.
Results
Female mice with hyperthyroidism had enlarged adrenal glands
Homozygous (HOM) female TSHR knock-in (KI) mice (TSHRD633HKI), carrying an activating TSHR mutation, had significantly elevated serum TH concentrations at one to four months of age (Fig. 1A, upper panel). Furthermore, the serum TSH concentrations in HOM TSHRD633HKI females were lower than those in WT mice before the age of six months. Thereafter, serum TSH levels remained low in HOM mice over the entire 12-month study period (Supplementary Fig. S1). Heterozygous (HET) KI females showed subclinical hyperthyroidism, with decreased serum TSH, and TH concentrations that were still in the normal range (Supplementary Fig. S1). In agreement with a previous study, HOM males only presented with mild overt hyperthyroidism at approximately two months of age (16).
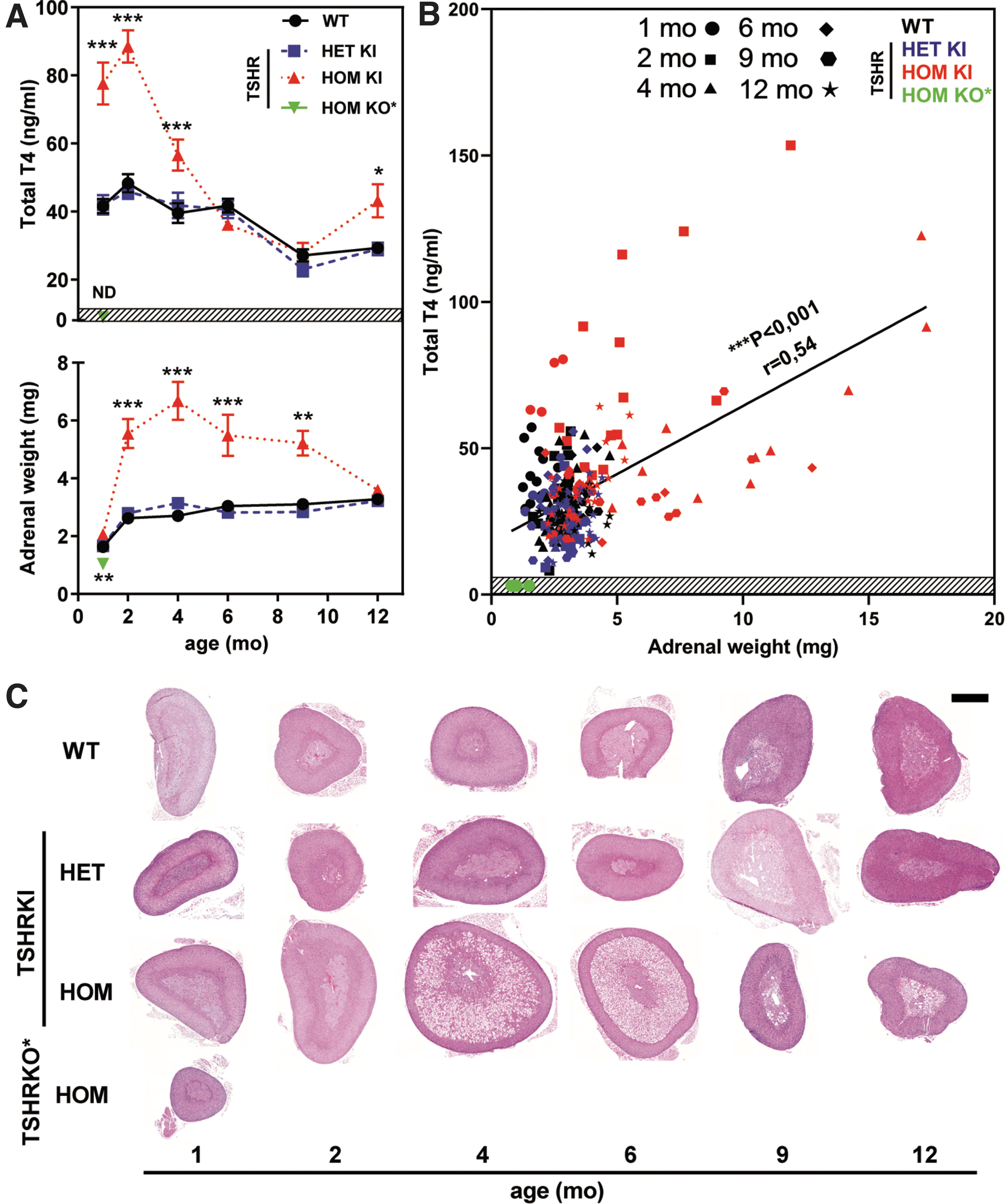
Altered TH concentrations at early age induce large changes in adrenal gland weight and histology. Serum TH concentration, adrenal weight, and size in the hyperthyroid TSHRD633HKI (TSHRKI mice carry an activating D633H mutation) and hypothyroid TSHRKO* (TSHRKO* mice carry an inactivating TSHR V448Afs31 mutant) nulliparous female mouse models at different ages. (
In hyperthyroid TSHRD633HKI females, adrenal weights were significantly elevated (Fig. 1A, lower panel) between two and nine months of age, as compared with those in WT mice. However, no changes in adrenal weight were observed in HET-KI females or mutant males (Supplementary Fig. S2A–F). Adrenal weights in hyperthyroid female TSHRD633HKI mice (Fig. 1A, lower panel) correlated positively (r = 0.54; ***p < 0.001) with serum thyroxine concentrations (Fig. 1B).
The adrenals became slightly enlarged already at 1 month of age, peaked at 4 months of age (HOM: mean adrenal weight 6.68 mg, SEM ±0.66: vs. WT 2.71 mg, SEM ±0.10), and then became smaller with age (Figs. 1 and 2).
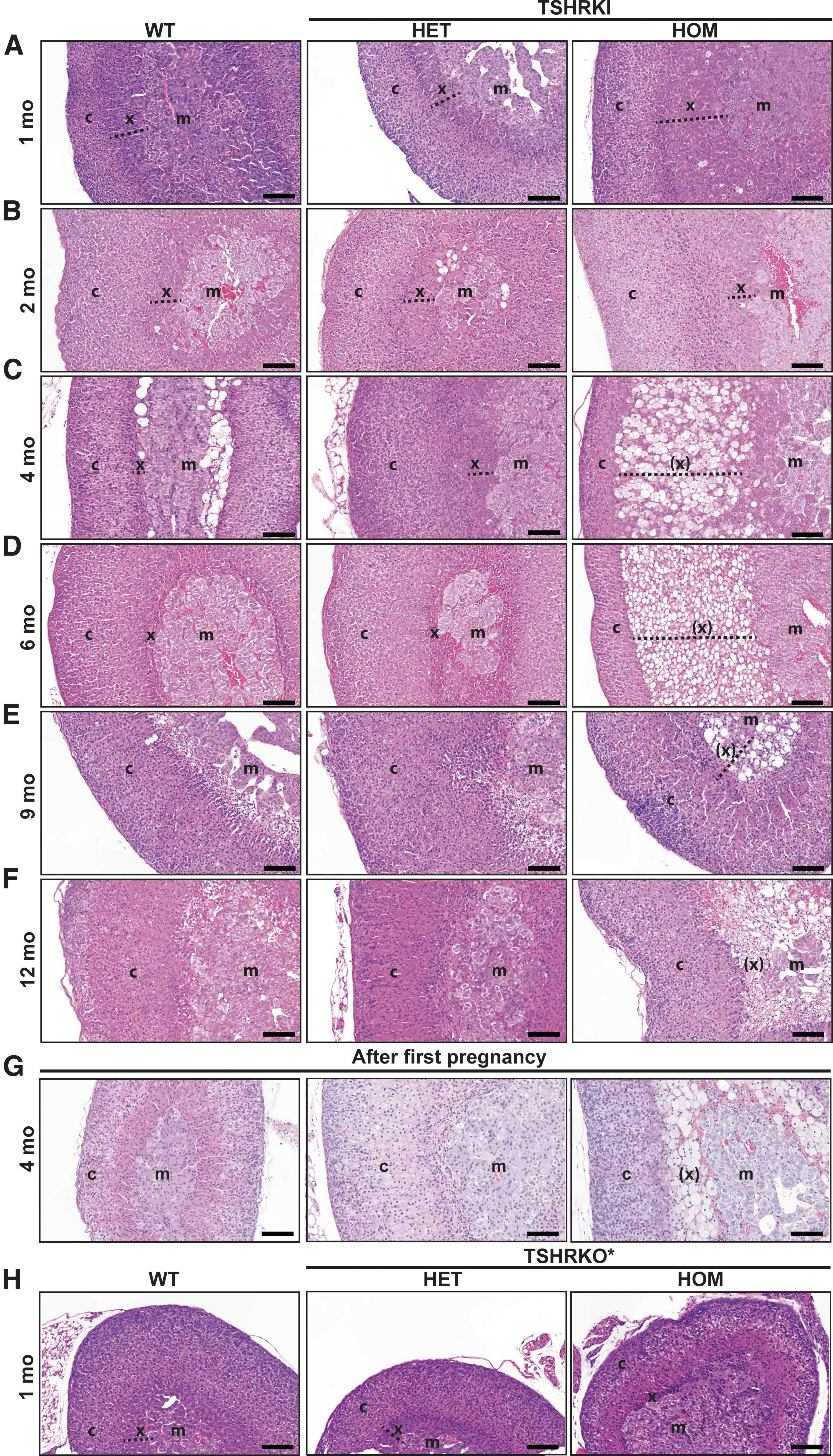
Postnatal hyperthyroidism leads to hypertrophy, extended vacuolization, and delayed regression of X-zone in hyperthyroid TSHRD633HKI female mice. Adrenal histology was evaluated in hyperthyroid (TSHRD633HKI) and hypothyroid (TSHRKO*) models. Furthermore, impact of first pregnancy on the regression of X-zone was visualized in TSHRD633HKI mice. Representative HE-stained images of adrenals of control (WT), HET, and HOM TSHRD633HKI virgin female mice at different ages (1–12 months) (
Postnatal hyperthyroidism led to hypertrophy, extended vacuolization, and delayed involution of the X-zone in female mice, and the lack of TH at birth impaired X-zone development
Histological analysis indicated that the increase in adrenal weights was associated with X-zone hypertrophy (Figs. 1C and 2A–F). Thereafter, extended areas of vacuolated cells appeared between the medulla and cortex at two months of age (Figs. 1C and 2A–F). No obvious alterations were observed in these zones between the adrenals from TSHRD633HKI and WT mice (Fig. 2). The X-zone normally disappears after the first pregnancy in mice (17). However, in contrast to the WT mice, relatively large areas of vacuolated cells persisted in the HOM TSHRD633HKI mice after pregnancy (Fig. 2G).
Immunohistochemical analysis of the X-zone marker 20αHSD revealed abundant staining of the inner cortical area of the hyperthyroid females (Fig. 3). In contrast, the X-zone layer was very thin and showed a weak 20αHSD staining in mice with CH carrying an inactivating TSHR mutant (namely TSHR V488fs31, hereafter called TSHRKO* mice; Fig. 3C and Supplementary Fig. S3A, B). The zone containing vacuolated cells in the adrenals of female TSHRD633HKI mice derived from the 20αHSD-positive X-zone cells, and 20αHSD staining diminished during involution (Fig. 2). No difference was found in the appearance of the medulla or staining with the chromogranin A (CgA) marker in TSHRD633HKI mice, when compared with WT mice (Fig. 3). However, in TSHRKO* mice, the cortex structure was partly disorganized, and CgA-positive areas were frequently observed within the cortex (Fig. 3). Oil Red O staining showed intense lipid staining in the altered zone of the HOM TSHRD633HKI females, whereas the cortex of the hypothyroid TSHRKOs* presented diminished staining (Supplementary Fig. S4).
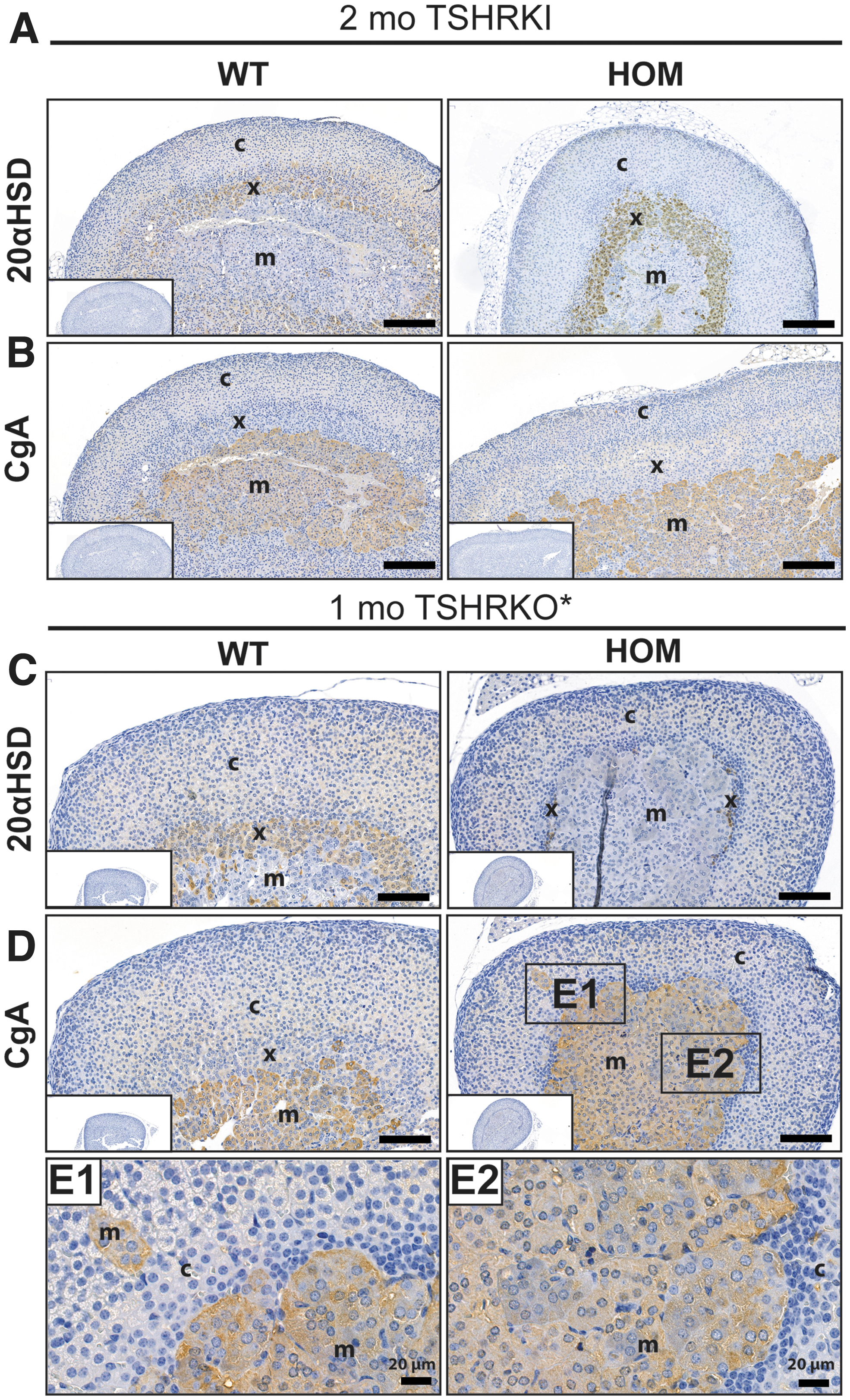
Immunohistochemical analysis of the adrenal X-zone 20αHSD and medulla CgA markers in the two-month-old TSHRKI and one-month old TSHRKO* female mice. IHC staining with antibodies against (
It is worth noting that both the TSHRD633HKI and TSHRKO* lines were fed soya-free RM3 E pellets containing ∼1.4 mg/kg of iodide, which exceeds the minimum (0.15 mg/kg), but is less than the standard amount (6 mg/kg) present in rodent feed. The very high iodine content of standard commercial rodent diets can play a role in the thyroid phenotypes in some models (18).
Altered TH concentrations modified adrenal gland gene expression at two months of age
To compare the impact of congenital hyperthyroidism and CH on adrenal gene expression, RNA sequencing (RNA-seq) was performed using adrenal RNA from both models and controls. Differential expression analyses of thyroid-responsive genes in the adrenals showed 186 upregulated and 26 downregulated genes in the hyperthyroid model (Fig. 4A–C). Of interest, in the TSHRKO* adrenals, 327 genes were upregulated and 34 were downregulated. Comparing the gene expression profiles between the hyperthyroid and hypothyroid groups revealed 277 upregulated and 130 downregulated genes (Fig. 4D–E). An overall evaluation of genes linked to a direct response to TH showed 134 and 283 genes differentially expressed genes in the hyperthyroid and hypothyroid models, respectively (Fig. 4F).
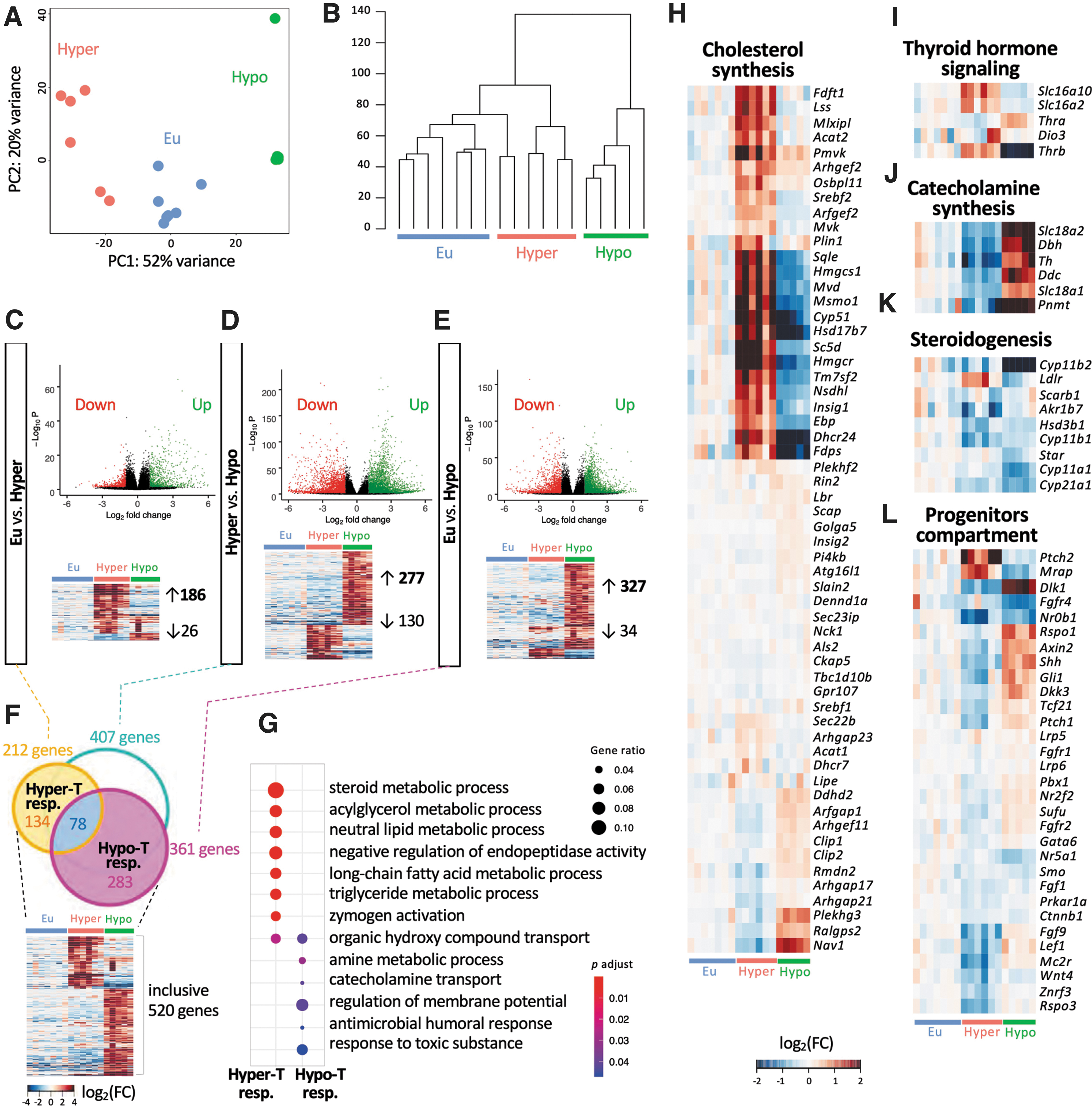
Altered TH concentrations modify adrenal gland gene expression at the age of two months. Adrenal transcriptome (RNA sequencing) analysis of the hyper-(TSHRD633HKI) and hypothyroid (TSHRKO*) female mice. (
Gene ontology (
Genes involved in steroidogenesis, specifically Star, Cyp11a1, Hsd3b1, Cyp21a1, Cyp11b1, Cyp11b2, Akr1b7, and Scarb1 were slightly downregulated in general in both mouse models (Fig. 4K). The Ldlr gene was upregulated in KI mice in response to higher TH concentrations. Genes required for TH signaling, including Slc16a10, Slc16a2, and Thrb were upregulated and downregulated in TSHRD633HKI and TSHRKO* mice, respectively. Of interest, Thra was significantly upregulated (1.5-fold increase) in TSHRKO* adrenals (Fig. 4I). The gene expression profile of adrenal progenitors revealed that Ptch2 and Mrap were clearly upregulated in hyperthyroidism and that Fgf9, Lef1, Mc2r, Wnt4, and Rspo3 were downregulated (Fig. 4L). In line with our RNA-seq analysis, similar gene expression changes were observed by quantitative real-time polymerase chain reaction analysis, and Cyp51 showed notable immunohistochemical staining in the altered X-zone (Supplementary Fig. S5 and Fig. 5).
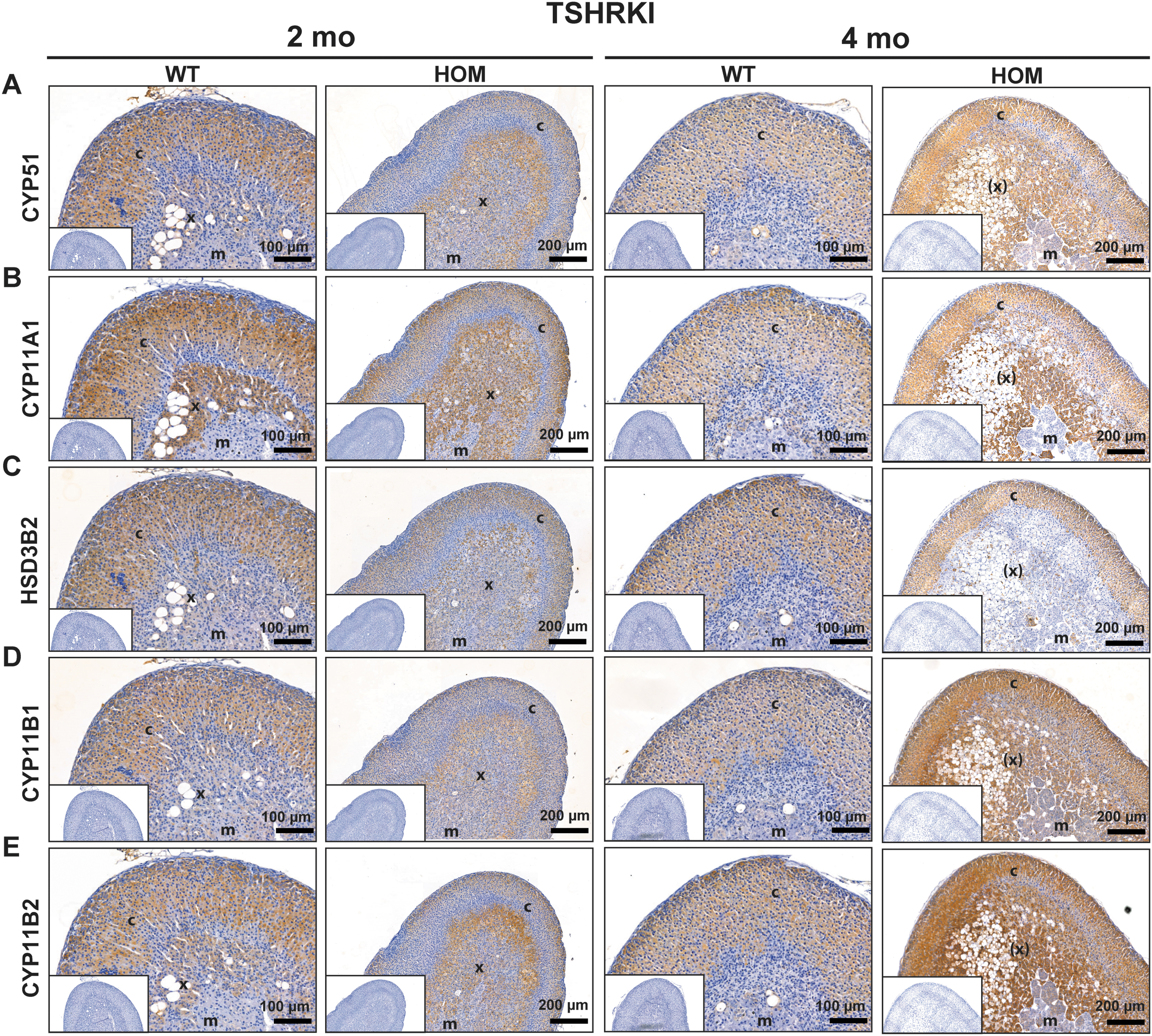
Altered adrenal X-zone area of the hyperthyroid TSHRD633HKI females shows abundant staining of CYP51, CYP11A1, HSD3B2, CYP11B1, and CYP11B2 steroidogenic enzymes. Adrenal IHC detection of the markers involved in adrenal steroidogenesis in two- and four-month-old TSHRD633HKI female mice. IHC staining with antibodies against (
Thrb but not Tshr, was abundantly expressed in the inner cortex of adrenals in female, hyperthyroid TSHRD633HKI mice
RNAScope analysis showed that Thrb was abundantly expressed in the inner cortex of the adrenal glands of female TSHRD633HKI mice, whereas only weak staining was observed with the controls (Supplementary Fig. S6B). Conversely, in the adrenals of TSHRKO* mice, Thrb transcripts were almost absent (Supplementary Fig. S6E). No apparent TSHR staining was observed in the cortices of the TSHRD633HKI or TSHRKO* mouse adrenal glands (Supplementary Fig. S6C, F). These findings agreed with our RNA-seq data showing very low Tshr expression in the adrenals (fragments per kilobase million <1), and that primary adrenal cells from four-month-old WT female mice did not respond to recombinant human TSH by producing cAMP (Supplementary Fig. S7). Cyclic adenosine monophosphate (cAMP) production was only detectable after stimulation with forskolin and bovine pituitary TSH extract (which is known to be contaminated with other pituitary hormones including ACTH).
Serum pituitary hormones, aldosterone, corticosterone, sodium concentrations, and intra-adrenal steroid hormones in hypothyroidism and hyperthyroidism
Serum pituitary hormone levels were measured to study the potential impact of TH on the hypothalamus–pituitary–adrenal axis (Supplementary Fig. S8). No significant changes in the serum ACTH, aldosterone, or corticosterone concentrations were observed between hyperthyroid and WT female mice (Supplementary Fig. S8). The serum sodium concentrations were slightly elevated in the hyperthyroid female mice, but such elevations were not seen in the hypothyroid model (Fig. 6). Mass spectrometric analysis of steroids in adrenal homogenates from four-month-old hyperthyroid mice showed no significant changes in the estrogen, estradiol, androstenedione, dihydrotestosterone, or testosterone concentrations, although the progesterone concentrations were significantly lower in the hyperthyroid HOM KI females than in control mice (Fig. 6).
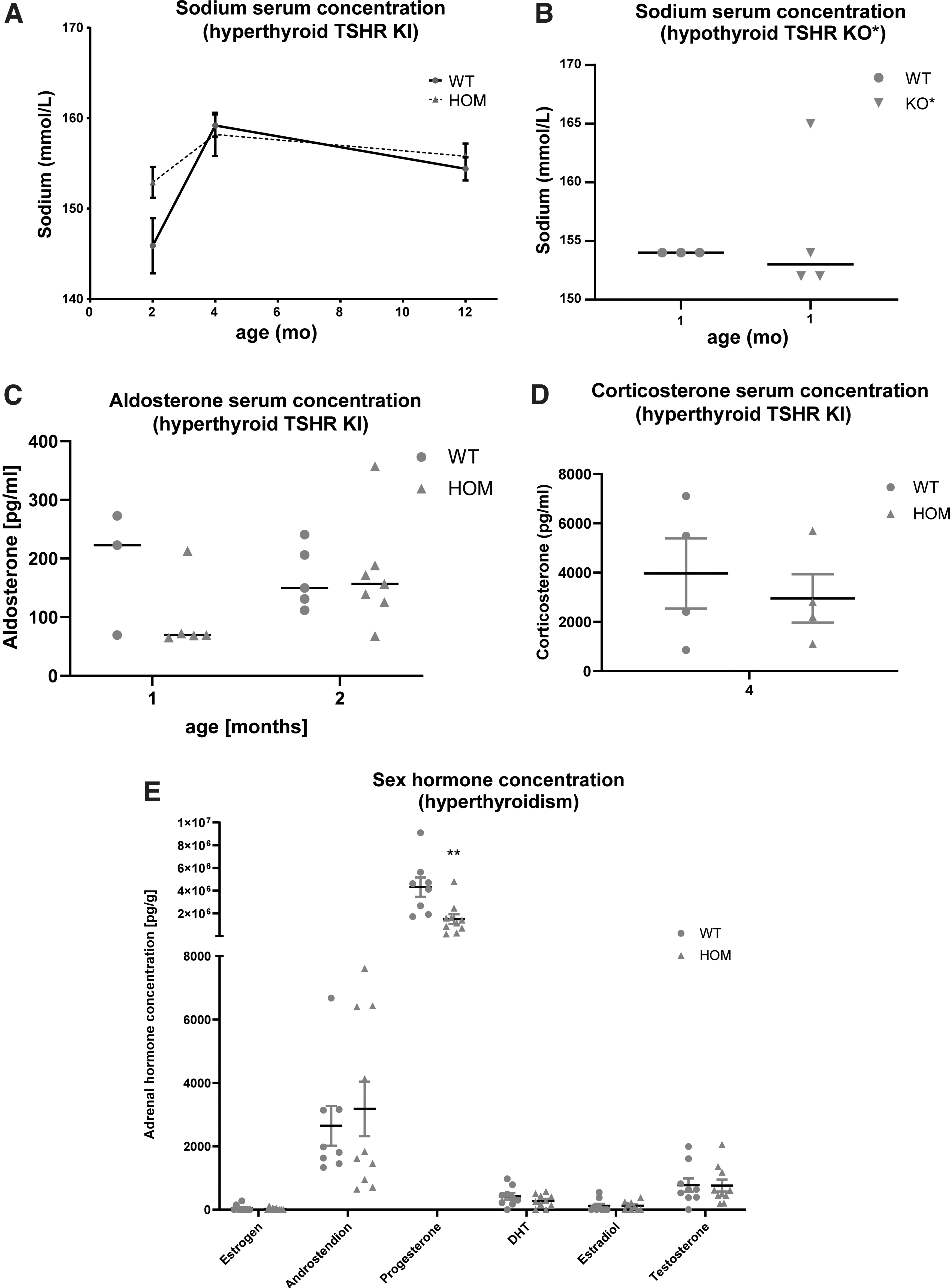
Serum sodium, aldosterone, corticosterone, and adrenal sex hormone concentrations in the TSHRD633HKI and TSHRKO* mice. Sodium serum concentrations in the (
Correlations between TSH concentrations and adrenal steroids in humans at birth and during PA
To test whether altered thyroid function is associated with the adrenal steroidogenesis in humans at birth, we studied a group of term newborns with CH and compared their 17-OHP concentrations with those of healthy term newborns of the same sex. The CH group showed higher 17-OHP concentrations (9.2 nmol/L; SD = 4.1; *p = 0.02) than the controls (5.5 nmol/L, SD = 2.8; Fig. 7). Moreover, we observed a weak correlation between umbilical serum TSH and 17-OHP concentrations was observed (***p < 0.001, r = 0.51). Overall, 2 of 33 patients with CH also had PA and 2 of 300 mothers had a history of taking any thyroid medication. In contrast, none of the 160 healthy controls had PA. As expected, owing to a low incidence of congenital hyperthyroidism, we could only find three newborns with congenital hyperthyroidism diagnosis at birth in our registry. To study whether serum TSH concentrations could affect adrenarche, we retrospectively evaluated the serum TSH concentrations in a previously described follow-up cohort of 98 children diagnosed of PA and 73 age-matched control children (19). In the group of children with PA, serum TSH concentrations showed a weak positive correlation with the concentrations of the main steroids produced by the adrenal zona reticularis (dehydroepiandrosterone [DHEA] and DHEA-S), as given in Table 1.

Serum TSH and adrenal steroid concentrations in controls and patients with congenital hypothyroidism. (
Correlations Between Serum Thyrotropin and Adrenal Androgen Concentrations in Patients Diagnosed with Premature Adrenarche and Controls at the Time of Diagnosis
Values in bold are statistically significant. Significance levels: ** p < 0.01; *** p < 0.001.
DHEA, dehydroepiandrosterone; r, Pearson's correlation; N, number of cases.
Discussion
In this study, we investigated the role of TH in adrenal development and function, using two murine models that mimic human CH and hyperthyroidism. Furthermore, we evaluated the potential impact of the thyroid on adrenal steroidogenesis in children with CH and PA. Female mice with a patient-derived constitutively activating TSHR mutation developed mild transient hyperthyroidism after birth, concomitant hypertrophy of the adrenal X-zone, and greatly enlarged adrenals. These findings support previous experimental data demonstrating the role of TH administration on X-zone development in rodents (9,20,21) and recent data suggesting that TH directly regulates the mouse X-zone, particularly in females, through THRβ1 (13).
Surprisingly, we observed significant reciprocal downregulation and upregulation of genes involved in catecholamine synthesis in the adrenals of the hyperthyroid and hypothyroid models, respectively. These observations suggest an interaction between TH and the adrenal medulla. In addition, TSH levels positively correlated with adrenal steroid concentrations in patients with CH or PA. In summary, our findings revealed that congenital hyperthyroidism or CH modifies development of the adrenal cortex and its steroidogenic activity, and crucially impact on medullary gene expression (Fig. 8).
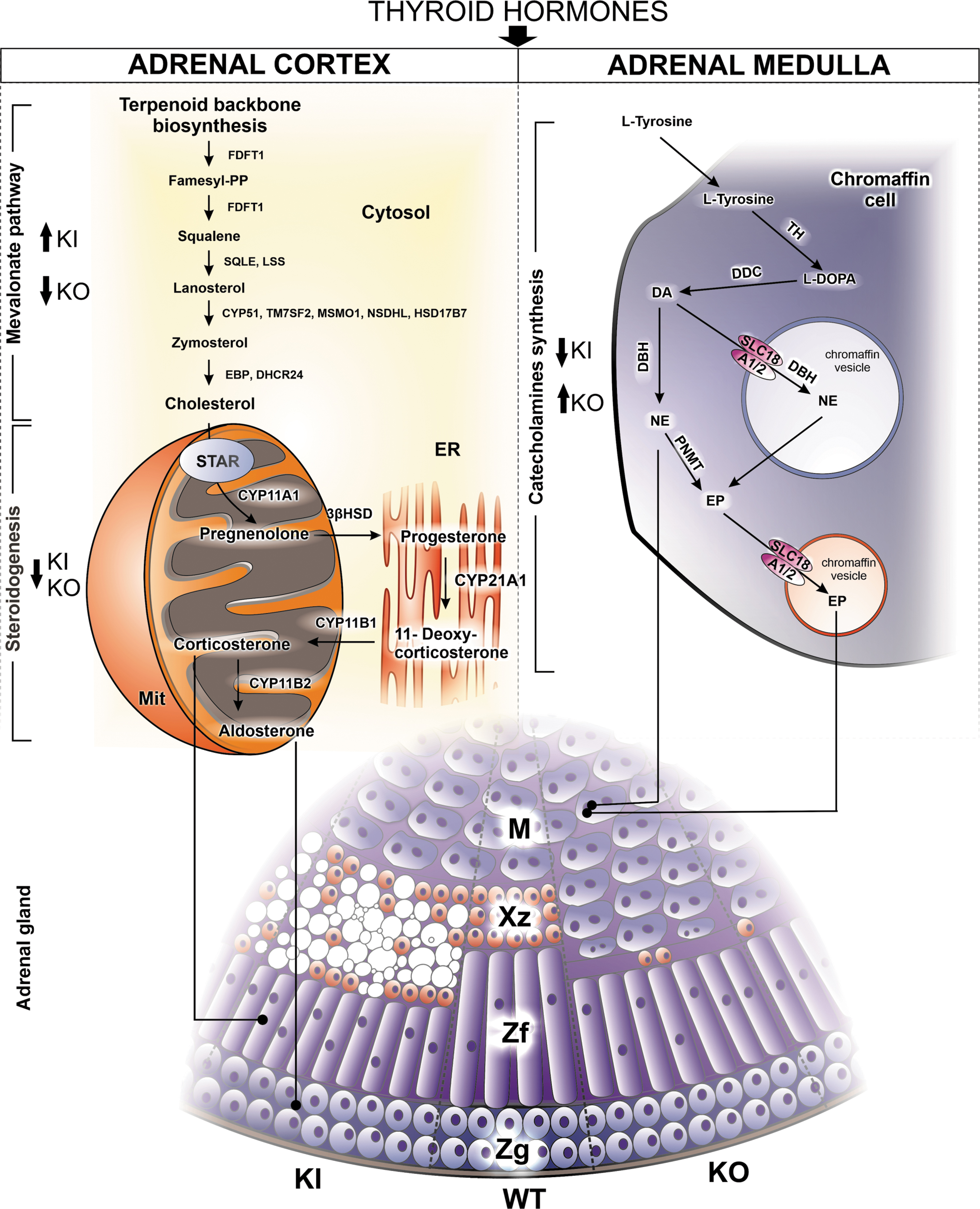
Summary of adrenal phenotypes in the hyper- and hypothyroid TSHR D633H KI and TSHRKO* mice. TSHRD633HKI and TSHRKO* mice carry an activating TSHR D633H or inactivating V448Afs31 mutations, respectively. The arrows indicate the up- or downregulations in KI and KO* mice. Color images are available online.
Enlarged adrenals were only observed in hyperthyroid female mice carrying an activating TSHR mutation. The adrenal size in these mice correlated with the serum TH concentrations, and no alterations in adrenal size or histology were observed in HET females or in HET or HOM males. In contrast, mice of both sexes carrying an inactivating TSHR mutation (and thus lacking TH at birth) presented with a poorly developed or almost absent X-zone in the adrenal glands. The sex differences can be explained by sexually dimorphic regression of the X-zone, which regresses after approximately four weeks of age in males, but persists in nulliparous females (17). HOM TSHRD633HKI males also presented elevated TH concentrations after birth, but had a normal adrenal size and histology. Thus, TH may not prevent a relatively rapid regression of the X-zone in males, mediated through testosterone (22).
Similarly, in hyperthyroid females after pregnancy, the altered X-zone area was partially diminished, indicating that TH did not completely prevent the physiological pregnancy-induced apoptosis in the X-zone with our model. In addition, adrenal responses may be mediated by TSHR (15,21) or TSH (23). As our models were based on activating and inactivating TSHR mutations, we evaluated a possible direct effect of TSHR on the adrenal phenotype. However, a direct role of TSHR in adrenals was excluded based on low TSHR expression found by RNA-seq and in situ hybridization, as well as the lack of TSH-mediated cAMP responses in adrenal primary cell culture. Pharmacological T3 treatment at an early age can promote sexually dimorphic gene expression in adrenals and affect genes that regulate adrenal cholesterol synthesis and steroidogenesis (23). Extensive TH-mediated lipid accumulation in hyperthyroid mouse adrenals has been proposed to directly result from TH activity on the adrenal glands through THRβ1 in the X-zone area (13).
Correspondingly, with our hyperthyroid model, when the lipid-laden extension of the adrenal inner cortex was prominent, the strongest upregulation was observed in genes associated with cholesterol metabolism. Cyp51 was also shown to be highly expressed in the altered X-zone area, suggesting that hyperthyroidism might directly trigger its expression in the altered adrenal cells of TSHRD633HKI females. Consistently, T3 has been shown to stimulate Cyp51 expression in primary mouse granulosa cells through THRβ1 and/or activation of the PI3K-Akt-pathway (24). Overall, apart from the gene expression changes and slightly decreased intra-adrenal progesterone concentrations, no obvious alterations occurred in adrenal steroidogenesis. Similar concentrations of serum ACTH, corticosterone, and aldosterone concentrations were found in both hyperthyroid females and controls. However, our analysis was limited to a small number of different steroids.
In our hyperthyroid model, the adrenal expression of Slc16a10 and Slc16a2 (MCT8), and Thrb were upregulated, whereas in the KOs adrenals Thrb was downregulated. A recent finding suggests that TH uptake in the adrenal gland is mainly mediated by MCT8, as TH activity in adrenals was partially blunted in Mct8KO mice (25). Furthermore, Thra expression was unexpectedly increased in the hypothyroid model, suggesting a compensation for TH signaling potentially because of the lack of X-zone in TSHRKO*.
Abundant upregulation and downregulation of key genes involved in catecholamine synthesis observed in our hypothyroid and hyperthyroid models, respectively, suggest a direct function of TH in the adrenal medulla. In addition to altered medullary gene expression, histological alterations and CgA-positive staining in the cortex were observed in our hypothyroid model. These findings are supported by previous data showing that experimentally modifying the TH concentrations can alter adrenal medullary function (26,27). However, further studies are warranted to elucidate whether altered medullary gene expression reflects changes in the adrenal cortex or compensation of the medulla to altered body temperature with our models (28).
An important question is whether TH plays a role in human adrenal function and development. To our knowledge, adrenal function in patients with thyroid hormone resistance, activating or inactivating TSHR mutations has not been studied (29). Recent results suggested a role for TH in the fetal adrenal cortex remodeling (8). Consistently, the 17-OHP concentrations at birth in patients with CH positively correlated with the serum TSH concentrations in our study. However, both observations were hampered by the small number of cases. Congenital hyperthyroidism is a very rare condition: thus, a larger cohort is warranted to evaluate the potential impact of congenital hyperthyroidism on the adrenals. A positive correlation between TSH and adrenal steroid concentrations in patients with CH seems to contradict with the reduced or enlarged adrenal X-zones seen in our hypothyroid or hyperthyroid murine models, respectively.
Although the human fetal adrenal zone and mouse X-zone share similarities, they might show differences in their responses to TH. To date, a knowledge gap exists regarding the initiation mechanism of adrenarche (the rise in adrenal androgen synthesis in humans before puberty). During adrenarche, growth of the zona reticularis has been observed, together with increased expression of key steroidogenic enzymes (30). However, recent evidence indicates that adrenarche does not present a rapid rise of adrenal androgens, but rather that adrenarchal programming occurs during early infancy (31,32).
Similarly, a positive correlation between serum TSH and adrenal steroids was demonstrated in children with PA, in this study. This correlation could not be explained by an increased body weight, which is also associated with adrenarche and elevated TSH concentrations (33).The potential impact of TSH on the adrenal cortex in these patients probably occurred during early infancy, as there is no clinical evidence of significantly altered adrenal steroidogenesis in hypothyroidism or hyperthyroidism at the time of diagnosis. Moreover, no adrenal phenotypes were observed with our previous models of adult-onset hypothyroidism (28).
Pregnancies of CH fetuses are often complicated by a prolonged gestational length. Recent data generated by Camm et al. with sheep fetuses, suggested that TH deficiency in utero suppresses the development of the fetal hypothalamic–pituitary–adrenal axis and contributes to the delay of fetal maturation and delivery (34). In agreement with our study, they also demonstrated decreased total adrenal gland masses, an increased relative mass of the adrenal medulla, and decreased expression of several key genes involved in steroidogenesis in CH fetuses compared with controls.
In conclusion, we report that altered thyroid function in early life plays an essential role in adrenal development in mice. Furthermore, we demonstrated that thyroid function impacts on adrenal function in PA and CH in humans.
Footnotes
Acknowledgments
The authors thank Prof. Johanna T. Arola for commenting on the adrenal histology, and the technicians Taina Kirjonen, Anna Kostiander, Nina Messner, Heli Niittymäki, Katri Hovirinta, Marja-Riitta Kajaala and Erica Nyman for technical assistance. The authors also thank Joonas Khabbal and Emrah Yatkin for laboratory measurements.
Study Approval
All experiments were performed in accordance with the institutional guidelines approved by the National Animal Experiment Board (license number 10266). For the human study, the register-based evaluation was approved by the Ethics Committee of the Hospital District of Southwest Finland (approval number 108/180/2010). The study protocol was approved previously by the Research Ethics Committee of the Kuopio University Hospital, and an informed written consent was obtained from the parents and their children.
Authors' Contributions
K.P., H.J., C.L., H.U. and J.K. conceptualized the study. C.L., F.-P.Z., R.P., and M.P. participated in generation of the models. K.P., C.L., S.T., H.J., H.U., K.P., M.C., H.S.Z., C.J.H., C.O., J.J., and M.D. performed research. B.-M.L. and R.K. investigated clinical data. K.P., C.L., H.J., J.J., and J.K. analyzed the data. I.H., N.R., M.P., J.T., A.K., H.Z., and C.J.H. supervised the work. K.P. and J.K. wrote the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
J.K. received grants from Finnish Pediatric and Medical Foundations, Turku University Hospital and Sigrid Juselius, and K.P. from TuDMM, Finnish Cultural (Grant no: 00200833 and 85211866), and Finnish Cancer Foundations. C.J.H. is supported by the United States National Institutes of Health, Grant No. R00HD082686.
Supplementary Material
Supplementary Data
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Supplementary Figure S7
Supplementary Figure S8
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3