
Abstract
Background:
In contrast to the thyroid hormones (THs) 3,3′,5-triiodothyronine (T3) and 3,3′,5,5′-tetraiodothyronine (thyroxine or T4), the binding characteristics of the thyroid hormone distributor proteins (THDP), thyroxine-binding globulin (TBG), albumin, and transthyretin in relation to TH metabolites are mostly lacking. In this study, we determined the distribution and binding affinity of TH metabolites to THDP, which is important for adequate interpretation of TH metabolite concentrations.
Methods:
Distribution of 125I-3,3′-diiodothyronine (3,3′-T2), -T3, -3,3′,5′-triiodothyronine (rT3), -3,3′,5-triiodothyroacetic acid (TA3), and -3,3′,5,5′-tetraiodothyroacetic acid (TA4) to TBG, transthyretin, and albumin was determined by agar gel electrophoresis. The rank order of affinity (IC50) of TBG and transthyretin to thyronine (T0), 3-monoiodothyronine (3-T1), 3,5-diiodothyronine (3,5-T2), 3,3′-T2, T3, rT3, T4, TA3, and TA4 was determined with a radioligand, competitive binding assay. In healthy subjects, associations of serum TBG, transthyretin, and albumin with TH and its metabolites were analyzed using multiple linear regression models, adjusted for sex and age.
Results:
While T3 and T4 are predominantly bound to TBG, we demonstrated that the predominant THDP of 3,3′-T2 and rT3 is albumin, of TA3 is transthyretin and albumin, and of TA4 is transthyretin. With the radioligand binding assay, we showed that the rank order of affinity was T4>TA4 = rT3>T3>TA3 = 3,3′-T2 > 3-T1 = 3,5-T2>T0 for TBG (IC50-range: 0.36 nM to >100 μM) and TA4>T4 = TA3>rT3>T3 > 3,3′-T2 > 3-T1 > 3,5-T2>T0 for transthyretin (IC50-range: 0.94 nM to >100 μM). TBG, transthyretin, and albumin were not associated with T0, 3-T1, 3,3′-T2, rT3, and TA4.
Conclusions:
Differences in serum TBG, transthyretin, and albumin concentrations within the reference interval do not influence serum concentrations of T0, 3-T1, 3,3′-T2, rT3, and TA4. Distribution of TH metabolites between THDP differs from T4 and T3, which predominantly bind to TBG. The results from our study have potential clinical importance for adequate interpretation of TH metabolism in (patho)physiology.
Introduction
Thyroid hormones (THs) are important for the regulation of growth, development, and metabolism (1). According to the established concepts, the thyroid gland in human mainly produces the prohormone 3,3′,5,5′-tetraiodothyronine (thyroxine or T4). Approximately 20% of the circulating active hormone 3,3′,5-triiodothyronine (T3) is derived from thyroidal secretion, with the remainder being produced from T4 in other tissues such as the liver and skeletal muscle. THs are predominantly metabolized via sequential extrathyroidal deiodination by deiodinases yielding the group of iodothyronines (2,3).
Alternative metabolic pathways include alanine side-chain modification yielding iodothyronamines and iodothyroacetic acids, conjugation of the phenolic hydroxyl group with sulfonic acid or glucuronic acid yielding iodothyro sulfates and iodothyro glucuronides, respectively, and oxidative cleavage of the diphenyl ether ring yielding iodotyrosines (2,3). Side-chain modifications may contribute significantly to TH metabolism, which is underscored by the fact that a substantial proportion of THs are excreted as thyroacetic acid (TA0) in urine (4), although others found only 14% of T3 and 1–2% of T4 undergoing oxidative deamination (5,6).
The free hormone hypothesis suggests that the “unbound” or “free” TH is available for uptake by target tissues and is therefore physiologically most important for TH action (7,8). In the human circulation, only 0.03% of T4 and 0.3% of T3 are in “unbound”/“free” form, and the majority of T4 and T3 is bound to thyroxine-binding globulin (TBG), transthyretin (TTR; also known as [thyroxine-binding] prealbumin), and albumin (9). These three thyroid hormone distributor proteins (THDP) enable a buffer system to maintain an extrathyroidal TH pool and distribute THs across target tissues (10,11).
In human serum, albumin is the main protein (35–50 g/L), while TTR and TBG concentrations are 100- and 1000-fold lower, respectively. Human TBG has the highest affinity for T3 and T4 (nanomolar range) followed by TTR and albumin (both micromolar range) resulting in 75% of T3 and T4 that is bound to TBG and a small fraction that is bound to human TTR (T3: <5%; T4: 20%) and human albumin (T3: 20%; T4: 5%) (10,12). Most large, omni- or herbivorous mammals including rodents express TBG, but the binding affinity of TBG for T4 varies between species (12).
In recent years, several physiological functions have been suggested for TH metabolites, including rapid noncanonical effects on energy metabolism by 3,5-diiodothyronine (3,5-T2) and 3,3′-diiodothyronine (3,3′-T2) (13 –16), on neuronal function and cancer cell proliferation by 3,3′,5′-triiodothyronine (rT3) (17 –19), and on thyrotropin (TSH) secretion, the liver, bone, and heart by 3,3′,5-triiodothyroacetic acid (TA3) and 3,3′,5,5′-tetraiodothyroacetic acid (TA4) (20 –22). Free concentrations of TH and its metabolites (THM) are dependent on serum THDP concentrations and their binding affinity, which could affect clearance, tissue uptake, and metabolism. Literature on the binding characteristics of THDP to TH metabolites is inconclusive due to the use of nonselective antibodies, nonstandardized methods, or use of human serum as a source of THDP instead of purified THDP (23 –25).
To date, robust data on serum THDP characteristics in relation to the different metabolites are therefore lacking. At present, purified THDP, purified metabolites, and selective analytical methods to quantify TBG, TTR, and albumin are available as well as highly selective liquid chromatography-tandem mass spectrometry (LC-MS/MS) methods to quantify total concentrations of multiple THM in serum in a single run (26 –32). These developments provide the opportunity to adequately study the binding characteristics of THDP in relation to THM as well as the association with in vivo concentrations. This knowledge is important for adequate interpretation of THM concentrations.
The purpose of this study is to evaluate if the THM reference interval is influenced by differences in serum THDP concentrations. In addition, we directly estimated the distribution of 3,3′-T2, T3, rT3, TA3, and TA4 among serum TBG, TTR, and albumin with electrophoresis and measured the binding affinity of TBG and TTR to THM with a competitive binding assay.
Materials and Methods
Chemicals
Glycine, sodium azide, acetic acid, and bromophenol blue were obtained from Merck (Darmstadt, Germany). Agar Difco was obtained from Becton, Dickinson and Company (New Jersey, USA). Sterile water was obtained from Baxter (Illinois, USA). 125I-3,3′-T2, -T3, -T4, -rT3, -TA3, and -TA4 were produced according to Mol and Visser (33). L-thyronine (T0) (purity >98.0%), T3 (purity ≥95%), and T4 (purity ≥98%) were obtained from Sigma-Aldrich, rT3 (purity ≥98.0%) from Calbiochem, and 3-monoiodothyronine (3-T1), 3,5-T2, 3,3′-T2, TA3, and TA4 from Toronto Research Chemicals (all purity >95.0%). TBG (purity ≥99%) and TTR (purity >95%) were obtained from Bio-Rad. HEN-buffer contained 0.1 M
Agarose gel electrophoresis
Electrophoresis was used to determine the distribution of 3,3′-T2, T3, rT3, TA3, and TA4 among TBG, TTR, and albumin in serum from healthy donors (three men and three women, previously collected to determine reference intervals (26,34,35). We did not succeed to determine the distribution of T0, 3-T1, and 3,5-T2 because T0 lacks iodine atoms and the inner ring cannot be labeled with radioactive iodine using standard methods. We did not determine the distribution of T4 as this is well-known (10). 125I-3,3′-T2, -T3, -rT3, -TA3, and -TA4 (800,000 CPM/sample) were evaporated to dryness under a stream of nitrogen at 37°C (Dri-Block DB-3D; Techne, Staffordshire, United Kingdom) and incubated (two hours, room temperature) with serum (50 μL).
Serum (5 μL) was mixed with a visual marker (1 μL 0.25% bromophenol blue) and was separated by agarose gel electrophoresis, 9 g/L agarose, pH 8.6, with a glycine buffer containing 15.1 g/L glycine, 0.77% (v/v) acetic acid, and 0.2 g/L sodium azide; the applied current was 70 mA/slide (∼150 V) for approximately one hour (computer-controlled electrophoresis power supply model 3000Xi; Bio-Rad, Hercules, USA). The gel was sliced into 2-mm fragments and each fragment was transferred to a polypropylene tube (5 mL; Greiner bio-one, Alphen aan den Rijn, The Netherlands). Radioactivity representing bound THM was measured (2470 WIZARD2; PerkinElmer, Groningen, The Netherlands).
Competitive protein binding assay
Competitive protein-binding assays displacing 125I-T4 bound to TBG or TTR were used to determine quantitative data on binding of T0, 3-T1, 3,5-T2, 3,3′-T2, T3, rT3, T4, TA3, and TA4 to TBG or TTR. An initial protein binding of ∼15% between 125I-T4 and TBG or TTR was preferred to prevent ligand depletion. The concentration responses of TBG, TTR, and albumin were determined by comparing several concentrations of TBG (range: 0.0056–1.85 nM), TTR (range: 0.75–7.5 nM), and albumin (range: 30–1000 nM). A concentration of 0.093 nM TBG and 3 nM TTR was considered optimal (Supplementary Fig. S1).
For albumin, a linear relationship between the THDP and 125I-T4 binding could not be established, impeding assay development. For TBG, increasing concentrations of nonradioactive THM (0–30 μM) were mixed with ∼0.02 nM 125I-T4 (40,000 CPM/sample) and 0.093 nM TBG in 400 μL HEN-buffer and incubated (2 hours, 30°C, pH 7.4). For TTR, increasing concentrations of nonradioactive THM (0–100 μM) were mixed with ∼0.05 nM 125I-T4 (50,000 CPM/sample) and 3 nM TTR in 200 μL HEN-buffer and incubated (overnight, 4°C, pH 7.4). Subsequently, samples were centrifuged (1 minute, 1500 rpm, 4°C) on polyacrylamide columns to separate unbound 125I-T4. The column was washed (400 or 200 μL of HEN-buffer for TBG and TTR, respectively), and radioactivity in the combined eluates was measured (Riastar C5420; PerkinElmer).
TBG, TTR, and albumin measurement
We have used fresh residual material from daily routine analysis of healthy blood donors. In agreement with the federation of Dutch medical scientific societies code of conduct for responsible use (METC Erasmus M and Sanquin) no permission form the Ethical Committee for the use of anonymous residual material is required. Donors do have the option to declare a no-cooperation statement for this procedure. Exclusion criteria were donors with heart and vascular disease, epilepsy, autoimmune disease, malignant disease, a history of thyroid disease, and use of any medication.
From all participants, a written informed consent was obtained, and age and gender were noted. TBG was measured in serum by an immunometric assay (Immulite 2000 XPi; Siemens, Los Angeles, USA), and TTR and albumin were measured in serum by an immunoturbidimetric and colorimetric assay, respectively (Cobas 6000; Roche, Mannheim, Germany).
Statistical analysis
GraphPad Prism (version 5.03) was used to calculate the half-maximal inhibitory concentration (IC50) values using a nonlinear fit to determine the binding affinity of TBG and TTR for THM. R statistical software (version 3.5.2; packages dplyr, ggplot2, ggpubr) was used to study the associations between TBG, TTR, and albumin, and the associations of TBG, TTR, and albumin with THM using multiple linear regression models adjusted for sex and age. We added product interaction terms to study effect modification by sex or by age, with a p-value <0.15 threshold for subsequent stratification. Relevant error and/or model-fit outliers were detected based on visual inspection of the data (histograms and XY plots for associations tested with regression analysis) and a quantitative approach (top/bottom 2.5th percent for error outliers and a change in the effect estimate of interest or the R 2 of at least 10%) (36).
A maximum of two outliers for TBG were excluded for regression analyses, which led only to reduced effect estimates. In case of relevant effect modification by age continuously, stratified analyses aimed at quantifying differences were performed by quartiles. For all other analyses, a p-value <0.05 was considered statistically significant (37).
Results
Population characteristics
In total, 253 healthy individuals (132 men, 121 women) were eligible for the study, of which two individuals were excluded due to an insufficient amount of serum. Median (interquartile ranges) age of men was 54 (43–63) years and of women was 37 (26–55) years.
Serum protein distribution of THM
T3 was predominantly bound to TBG (74.6%) in contrast to 3,3′-T2 and rT3, which are predominantly bound to albumin (58.4% and 62.6%, respectively; Table 1 and Fig. 1 and Supplementary Figs. S2 and S3). For the iodothyroacetic acids, TA3 is equally distributed between albumin (46.9%) and TTR (48.7%), whereas TA4 is predominantly bound to TTR (84.1%) and only a small fraction (13.1%) to albumin (Table 1 and Fig. 1 and Supplementary Figs. S2 and S3).
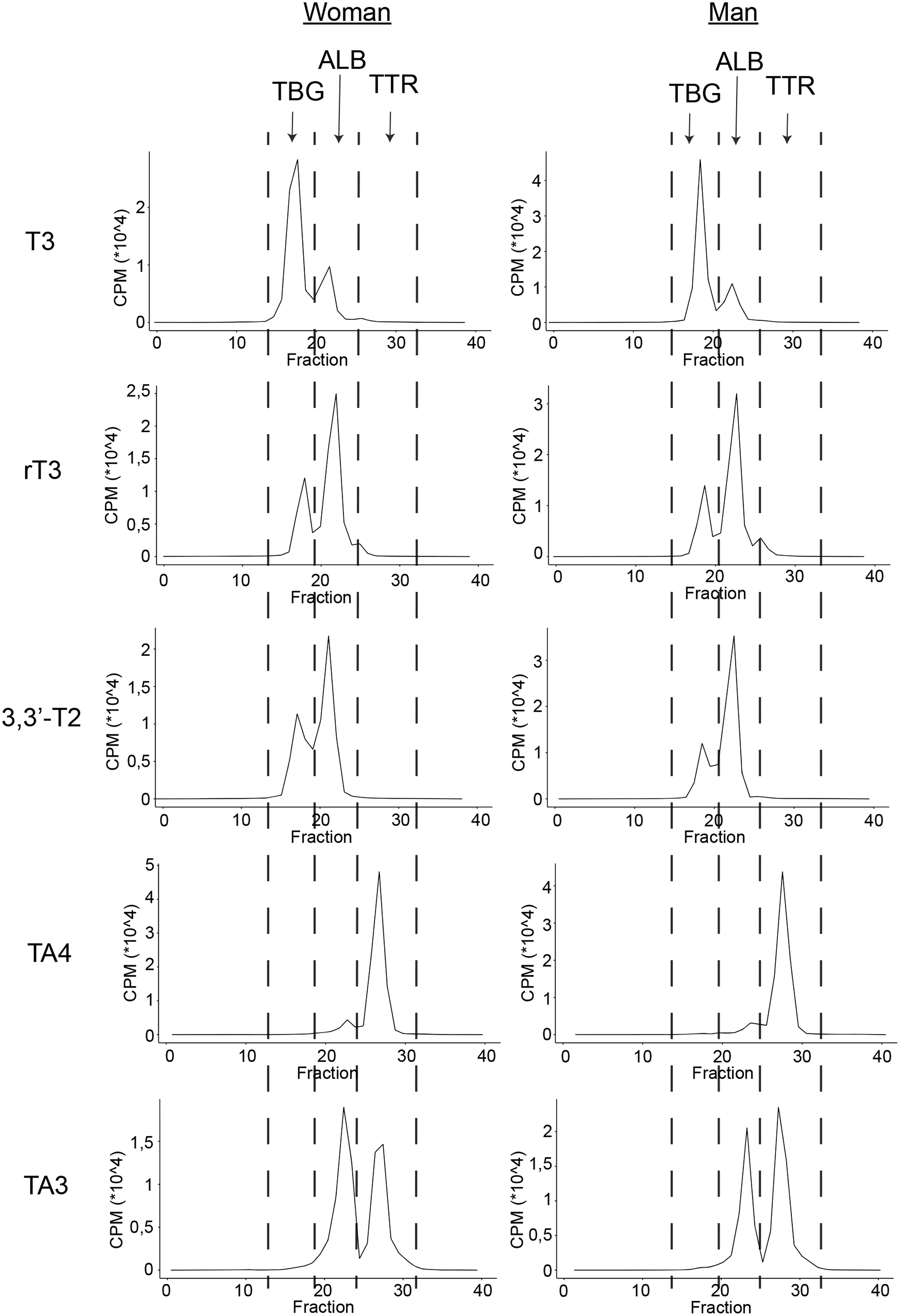
Serum electrophoresis pattern of T3, rT3, 3,3′-T2, TA4, and TA3. Representative electrophoresis pattern of T3, rT3, 3,3′-T2, TA4, and TA3. Electrophoresis patterns of other two women and men are added in Supplementary Figs. S2 and S3. 3,3′-T2, 3,3′-diiodothyronine; ALB, albumin; TA3, 3,3′,5-triiodothyroacetic acid; T3, 3,3’,5-triiodothyronine; rT3, 3,3′,5′-triiodothyronine; TA4, 3,3′,5,5′-tetraiodothyroacetic acid; TBG, thyroxine-binding globulin; TTR, transthyretin.
Binding Characteristics of Thyroid Hormone and Its Metabolites with Human Thyroid Hormone Distributor Proteins
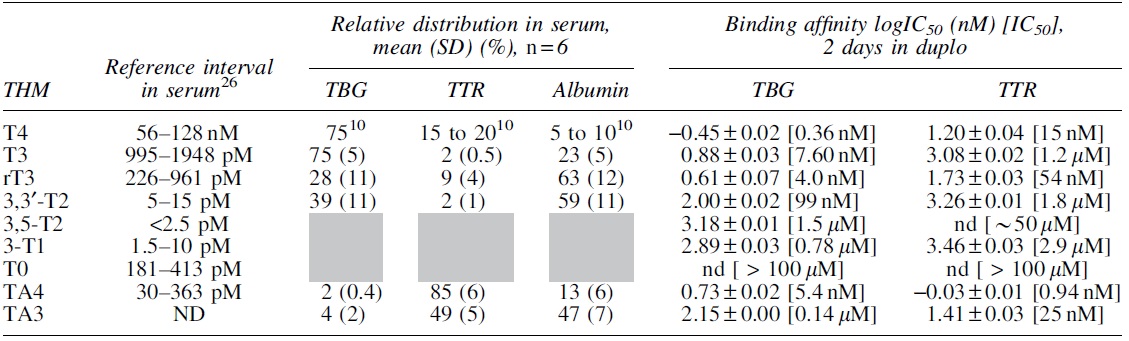
Reference interval is previously determined with LC-MS/MS (26). The relative distribution in serum for T4 was not determined as this is reported frequently in literature (10). Binding affinity was not determined for albumin. The relative distribution in serum was not determined for 3,5-T2, 3-T1, and T0 (represented in gray).
3-T1, 3-monoiodothyronine; 3,3′-T2, 3,3′-diiodothyronine; 3,5-T2, 3,5-diiodothyronine; T0, thyronine; T3, 3,3’,5-triiodothyronine; rT3, 3,3′,5′-triiodothyronine; T4, thyroxine; TA3, 3,3′,5-triiodothyroacetic acid; TA4, 3,3′,5,5′-tetraiodothyroacetic acid; TBG, thyroxine-binding globulin; THDP, thyroid hormone distributor protein; THM, thyroid hormone and its metabolites; TTR, transthyretin; SD, standard deviation; n = total number of individuals; ND, not detected; nd, not determined.
Binding affinity of TBG and TTR for THM
TBG had the highest affinity for T4 and was followed in descending order by TA4 = rT3>T3>TA3 = 3,3′-T2 > 3,5-T2 = 3-T1>T0 (Table 1 and Supplementary Fig. S4). TTR had the highest affinity for TA4 followed in descending order by T4 = TA3>rT3>T3 > 3,3′-T2 > 3-T1 > 3,5-T2>T0 (Table 1 and Supplementary Fig. S4). We could only estimate an IC50 value for TTR-3,5-T2 since we were unable to fit a curve for the competitive binding.
Association of TBG, TTR, and albumin with THM
The majority of 3,5-T2 (>90%) measurements were below the lower limit of detection and TA3 was not detected, which prohibited analyses. There was no association between TBG, TTR, and/or albumin, and therefore, the linear model was not adjusted for the concentration of other THDP (Supplementary Fig. S5). There was a positive association of TBG with T3 and T4 (respectively, p = 8.1 × 10−5 and p = 0.0029; Fig. 2), but not for TTR or albumin with T3 and T4 (Figs. 3 and 4). There was no association of TBG, TTR, or albumin with T0, 3-T1, 3,3′-T2, rT3, or TA4 (Figs. 2–4).
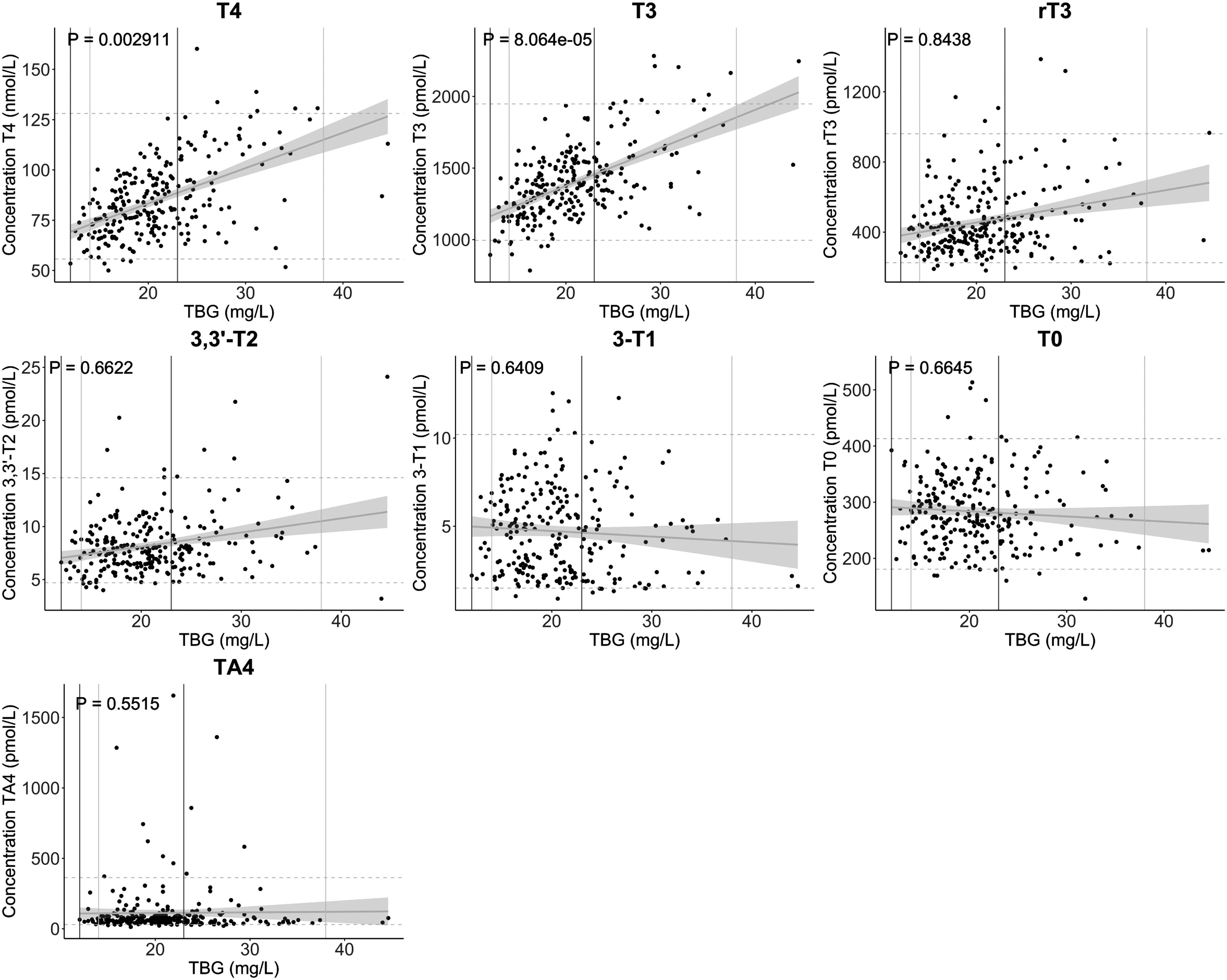
Association of human TBG with THM in healthy individuals. The regression line and confidence interval are presented in gray for all healthy individuals. Lines represent reference intervals for the THM (dashed, gray) and human TBG (men = solid, dark gray, women = solid, light gray). A linear model adjusted for sex and age was used (p-value <0.05 was considered statistically significant). THM, thyroid hormone and its metabolites.

Association of human TTR with THM in healthy individuals. The regression line and confidence interval are presented in gray for all healthy individuals. Lines represent reference intervals for the THM (dashed, gray) and human TTR (solid, gray). A linear model adjusted for sex and age was used (p-value <0.05 was considered statistically significant).

Association of human albumin with THM in healthy individuals. The regression line and confidence interval are presented in gray for all healthy individuals. Lines represent reference intervals for the THM (dashed, gray) and human albumin (solid, gray). A linear model adjusted for sex and age was used (p-value <0.05 was considered statistically significant).
Screening for sex-specific differences in the association of TBG, TTR, or albumin with THM suggested that the association of TBG with T0, 3,3′-T2, T3, rT3, and T4 and the association of TTR with 3,3′-T2, T3, rT3, T4, and TA4 could differ between men and women (Supplementary Table S1). Subsequent stratification was only performed with these associations. After stratification, the results implicated that there is a positive association of TTR with T4 and T3 in women (represented in light gray), but not in men (represented in dark gray). No potential clinically relevant patterns could be extracted from the other association plots (Supplementary Figs. S6 and S7).
Screening for age-specific differences in the association of TBG, TTR, or albumin with THM indicated that the association of TBG with T0, the association of TTR with T0, 3,3′-T2, and rT3, and the association of albumin with 3-T1, 3,3′-T2, and T3 could differ with age (Supplementary Table S2). After stratification, no potential clinically relevant patterns could be extracted from the association plots (Supplementary Fig. S8).
Discussion
In this study, we demonstrate that in contrast to the association of TBG with T3 and T4, differences in THDP concentrations are not associated with total serum TH metabolite concentrations within the reference interval in a linear model adjusted for sex and age. We found a positive association of TTR with T4 and T3 in women, but not in men, which will require further replication. We also showed that the binding characteristics of 3,3′-T2, rT3, TA3, and TA4 differ from T3 and T4.
Knowledge about the influence of THDP on TH metabolite concentrations, distribution of metabolites among THDP, and the binding affinity of THDP for metabolites is potentially important for adequate interpretation of serum concentrations. If fluctuations in THDP affect the pool of serum TH metabolites, this could be falsely interpreted as changes in the activity of metabolic pathways. In addition, since biological functions for TH metabolites have been proposed, changes in the concentration of THDP could also affect their biological availability and therefore action.
The association of T3 and T4 with TBG was expected, since TBG is the main THDP for T3 and T4. At higher total T4 and T3 concentrations, an increase in serum
TBG concentration decreases serum free T4 (fT4) concentrations. Although TTR and albumin account for 25% of serum T4 and T3 binding, TTR and albumin are not associated with T4 and T3 concentrations, at least within the normal range. Since the free hormone hypothesis states that TH uptake and metabolism, and therefore TH action, are dependent on the free and not the total fraction, downstream metabolites would in this case be altered via changes in HPT-axis activity when concentrations of serum THDP change because the negative feedback mechanism corrects fT4 concentrations. This, however, ignores the possible uptake of THM-bound THDP, as seen for TTR-T4 in, for example, the blood–cerebrospinal fluid barrier and placenta (38). The physiological relevance of these potential other mechanisms is not fully elucidated.
An association could be expected for TA4 and its metabolite TA3, which can effectively lower TSH production due to the high-affinity binding of TA3 to the pituitary TH receptor (39,40). TA4 is predominantly bound by TTR. Theoretically, decreases in serum TTR could therefore increase free-TA4 concentrations resulting in lower TSH secretion. Consequently, this will result in lower TH production, lower fT4, and lower downstream metabolite production, including TA4. However, we found no TTR-TA4 association, even though TA4 is present in subnanomolar concentrations and efficiently deiodinated to TA3. For TA3, an association could not be established, because TA3 could not be detected in healthy individuals.
For other metabolites, we speculated that high-affinity binding of TH metabolites to THDP buffers metabolites to protect them from clearance and restrict their cellular uptake and subsequent metabolism via deiodination or alternative pathways. However, we found no association with any THDP, for which there could be several reasons. First, metabolites may not bind to THDPs. However, apart from T0 this is likely not the case since binding assays showed binding of all metabolites, apart from T0, to TBG and TTR and as electrophoresis showed full distribution of tracer amounts of metabolites over THDP in serum containing endogenous THDP and THM. However, technical limitations impeded to test the distribution of T0, 3-T1, and 3,5-T2, the metabolites with the lowest binding affinities, among the THDP. Therefore, it cannot be excluded that for these metabolites, a considerable amount is present in free form.
Second, THM concentrations are determined by the rate of production and degradation/clearance. THs are metabolized via multiple pathways in several tissues, including inner- or outer ring deiodination, alanine side-chain metabolism, conjugation of the phenolic hydroxyl group, and oxidative cleavage of the diphenyl ether ring. Individual differences in rate-limiting steps of these pathways could be dominant over individual differences in protein binding to determine the amount of circulating THM. Finally, our study was conducted with serum samples from healthy individuals. It cannot be excluded that under (patho)physiological conditions leading to substantial changes in serum concentrations of THDP, such as pregnancy or genetic disorders, an association would be present.
In agreement with published data, our competitive binding data show that deiodination of THs results in a major loss of TBG- and TTR affinity supporting the importance of iodine atoms, in particular in the outer ring, for binding to TBG and TTR (25,41 –43). Perhaps this major loss in affinity also decreases the effect of fluctuations on TH binding protein on serum THM, although we were unable to determine the distribution of metabolites with fully deiodinated outer rings among the THDP. As previously reported by others (43), there is a major increase of affinity for TTR after oxidative deamination into iodothyroacetic acids. Nevertheless, iodothyroacetic acids are less distributed among TBG and albumin despite an affinity of TBG for TA3 and TA4 in the same order of magnitude as 3,3′-T2 and rT3, respectively.
This indicates that the binding capacity of TTR is high enough to bind the majority of TA3 and TA4. The rank order of affinity of TBG to THM is in accordance with previous literature, except for TA4, for which we found a higher binding affinity, and for 3,3′-T2 and 3,5-T2 for which we found a lower binding affinity (25,44,45). The binding affinity of TTR to 3,3′-T2, rT3, T4, and TA4 was in the same order of magnitude with published data, but lower for TA3 and 3-T1 (∼6-fold and ∼100-fold, respectively) and higher for 3,5-T2 (∼5-fold) (41,46 –49). We found an IC50-value of 1.2 μM for T3-TTR, which is within the wide range (80 nM–5 μM) reported (47,49).
Unexpectedly, we noticed an initial increased binding of radiolabeled T4 with increasing concentrations of T3 and 3,5-T2 before competition at higher doses, which likely results in an overestimation of the IC50 values. TTR contains two ligand binding sites that display negative cooperativity (50,51). Our results could mean that binding of T3 and 3,5-T2 to the second binding site may increase the affinity of the first site for the radioligand at concentrations where T3 and 3,5-T2 do not yet fully compete for binding.
The strengths of the current study are that we used human purified THDP (TBG (purity ≥99%) and TTR (purity >95%), in contrast to prior studies using human (diluted) serum or purified TBG with an impurity of albumin (23 –25). We also used THMs with high purity. In addition, we used selective analytical methods to quantify TBG, TTR, albumin, and THMs.
This study also has some limitations. First, the electrophoresis is performed at a nonphysiological pH of 8.6. Although the incubation of the samples was at physiological pH, performing the electrophoresis at nonphysiological pH may cause dissociation of the hormone and the THDP during electrophoresis. The conditions used for electrophoresis with a pH of 8.6 are consistent with earlier literature (52) and required for optimal separation of the THDP. This is likely due to the comparative isoelectric point of THDP [e.g., the theoretical isoelectric point of TBG is 5.87, TTR is 5.49, and albumin is 5.67 (53)] that would result in suboptimal resolution at pH 7.4. Tata suggested that both the number and the affinity of binding sites are not affected by a pH shift between physiological and 8.6 for T4 (54).
Second, the distribution of T0, 3-T1 and 3,5-T2 among the THDP could not be tested due to unavailability of inner ring labeled metabolites. Third, our LC-MS/MS method measures only total THM concentrations and not the free form. Fourth, 3,5-T2 and TA3 could not be measured with the current panel impeding the analyses of the association of TBG, TTR, and albumin with these metabolites. Because we measure multiple THs and metabolites in a single run, this restricts the possibilities to optimize the sample preparation, the liquid chromatography and the mass spectrometry settings for individual compounds. Fifth, serum samples from patients with (patho)physiological conditions leading to substantial changes in serum concentration of THDP, such as pregnancy or genetic disorders, are not included. Sixth, we did not perform adjustment for multiple statistical testing, so some of the results of this study may be subject to type 1 error and should be considered exploratory, requiring future replication.
In conclusion, differences in serum THDP concentrations do not influence total THM concentrations within the reference interval. Sex and age differences will require further replication. For the binding characteristics, we demonstrated that the most important THDP for 3,3′-T2 and rT3 is albumin, for TA4 is transthyretin, and for TA3 is equally distributed between transthyretin and albumin in contrast to T3 and T4, which are predominantly bound to TBG. The findings from our study have potential clinical importance for adequate interpretation of TH metabolism in (patho)physiology.
Footnotes
Authors' Contributions
R.M.S.J., M.E.M., R.P., and Y.B.d.R. designed the current study. R.M.S.J. and M.E.M. conducted the experiments. R.M.S.J. conducted the statistical analyses with the help of T.I.M.K. R.M.S.J. and M.E.M. wrote the article. All authors interpreted the experiments, and reviewed and revised the article to improve its intellectual and technical content.
Acknowledgment
We thank Selmar Leeuwenburgh for the 125I-labeling of TH (metabolites) used in the electrophoresis and competitive protein binding experiments.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Supplementary Figure S7
Supplementary Figure S8
Supplementary Table S1
Supplementary Table S2