
Abstract
Background:
Hypothyroidism causes ovarian dysfunction and infertility in women, in addition to being associated with hyperprolactinemia and reduced hypothalamic expression of kisspeptin (Kp). However, it remains unknown whether and how Kp is able to reverse the ovarian dysfunction caused by hypothyroidism.
Methods:
Hypothyroidism was induced in adult female Wistar rats using 6-propyl-2-thiouracil for 3 months. In the last month, half of the animals received Kp10. Blood samples were collected for dosage of free thyroxine, thyrotropin (TSH), luteinizing hormone (LH), prolactin (PRL), progesterone (P4), and estradiol (E2), and uteruses and ovaries were collected for histomorphometry. Body and ovarian weight and the number of corpora lutea were also evaluated. Half of the brains were evaluated by immunohistochemistry to Kp, and the other half had the arcuate nucleus of hypothalamus (ARC) and preoptic area microdissected for gene evaluation of Kiss1, Nkb, Pdyn, and Gnrh1. The pituitary gland and corpora lutea were also dissected for gene evaluation.
Results:
Hypothyroidism kept the animals predominantly acyclic and promoted a reduction in ovarian weight, number of corpora lutea, endometrial thickness, number of endometrial glands, and plasma LH, in addition to increasing the luteal messenger RNA (mRNA) expression of Star and Cyp11a1 and reducing 20αHsd. An increase in plasma PRL and P4 levels was also caused by hypothyroidism. Kp immunoreactivity and Kiss1 and Nkb mRNA levels in the ARC and Kiss1 in the anteroventral periventricular nucleus of hypothalamus were reduced in hypothyroid rats. Hypothyroid animals had lower pituitary gene expression of Gnrhr, Lhb, Prl, and Drd2, and an increase in Tshb. The treatment with Kp10 restored estrous cyclicality, plasma LH, ovarian and uterine morphology, and Cyp11a1, 3βHsd, and 20αHsd mRNA levels in the corpora lutea. Kp10 treatment did not alter gene expression for Kiss1 or Nkb in the ARC of hypothyroid rats. Nevertheless, Kp10 increased Lhb mRNA levels and reduced Tshb in the pituitary compared with the hypothyroid group.
Conclusions:
The present findings characterize the inhibitory effects of hypothyroidism on the hypothalamic–pituitary–gonadal axis in female rats and demonstrate that Kp10 is able to reverse the ovarian dysfunction caused by hypothyroidism, regardless of hyperprolactinemia.
Introduction
Hypothyroidism is associated with delayed puberty onset, anovulation, ovarian cysts, menstrual irregularity, 1,2 as well as failures in the occurrence of the preovulatory surge of luteinizing hormone (LH) and a reduction in gonadotropin-releasing hormone (GnRH) biosynthesis. 3 Hypothyroidism can be also accompanied by hyperprolactinemia, which is one of the main causes of infertility in women. 4,5
Dufourny et al 6 observed thyroid hormone receptors alpha (Thra) in the hypothalamus of sheep, suggesting that thyroid hormone deficiency may also directly affect the activity of Kiss1 neurons. The control of GnRH release during the ovarian cycle is performed by two groups of Kiss1-expressing neurons located in the arcuate nucleus of hypothalamus (ARC) and anteroventral periventricular nucleus (AVPV). 7,8 Moreover, kisspeptin (Kp) neurons in the ARC coexpress neurokinin-B (NKB) and dynorphin (KNDy neurons), which are neuropeptides that self-regulate the release of Kp and, consequently, the activity of GnRH neurons. 9 –13
Although the thyroid function is closely related to the control of gonadotropin and prolactin (PRL) secretion, evidence suggests that thyroid hormones can alter the central expression of Kp. In a study with male tilapia (Oreochromis niloticus), Ogawa et al 14 demonstrated that tilapia with hypothyroidism has reduced Kiss2 and Gnrh1, which were restored by triiodothyronine. Tomori et al 15 demonstrated that hypothyroidism in rats reduced the expression of Kp in the ARC. Furthermore, a recent study by our group showed that maternal hypothyroidism reduces the expression of Kiss1 and its receptor Kiss1r in the maternal–fetal interface of rats, suggesting another way in which hypothyroidism affects female fertility. 16
However, even with the observed hypothalamic reduction of Kp caused by thyroid hypofunction, it remains unknown whether and how Kp is able to revert the ovarian dysfunction caused by hypothyroidism. This is the first study to evaluate the use of Kp in gonadal dysfunction caused by hypothyroidism, and our results provide new information on the role of KNDy neurons in ovarian dysfunction caused by hypothyroidism.
Materials and Methods
Experimental design
A 3-month-old female Wistar rats (12–16 animals per group) (200–230 g) were housed under controlled temperature (22 ± 2°C), luminosity (1200 hours light/1200 hours dark), and airflow, with water and commercial chow ad libitum. Hypothyroidism was induced by daily oral administration of 6-propyl-2-thiouracil (PTU) (Sigma–Aldrich, St. Louis, MO) [4 mg/(kg·d)] for 3 months, 17 while the control group (12 animals) received water as a placebo. After 2 months of induction, half of the hypothyroid rats (16 animals) received intraperitoneal (i.p.) Kp10 (Cat. No. 4243; Tocris Bioscience, Bristol, UK) [8 μg/(kg·d), i.p.] for 30 days, while the others were treated with vehicle. Daily evaluations of the estrous cycle were conducted until the end of the 3-month experimental period, and then, on the day of diestrus, the animals were euthanized by decapitation 30 minutes after administration of Kp10 or placebo.
Blood samples were collected during the euthanasia, for dosage of free thyroxine (fT4), thyrotropin (TSH), LH, PRL, estradiol (E2), and progesterone (P4), and uteruses and ovaries were collected for histomorphometry according to Silva et al, 18 Sinha et al, 19 and Meng et al. 20 Body and ovarian weight and the number of corpora lutea were also evaluated. Half of the brains were evaluated by immunohistochemistry (IHC), and the other half had the ARC and preoptic area (POA) microdissected, stored in TRIzol, and frozen at −80°C for gene evaluation. The pituitary gland and three corpora lutea per rat were also dissected, stored in TRIzol, and frozen at −80°C for gene evaluation. All evaluations were performed blindly and without knowledge of the experimental groups. All the experiments were approved by the Ethics Committee on Animal Use (CEUA) of the Universidade Estadual de Santa Cruz (Protocol No. 036/16).
Hormone assay
Blood samples were collected in heparin tubes, centrifuged at 1008 g for 20 minutes at 4°C to obtain plasma, and stored at −20°C. Dosage of fT4 and TSH was performed by enzyme-linked immunosorbent assay (ELISA) following the manufacturer's instructions (fT4, IMMULITE; Siemens Medical Solutions Diagnostics, Malvern, PA; Sensitivity: 0.04 μg/dL; rat TSH, IMMULITE Third Generation TSH kit; Diagnostic Products, Los Angeles, CA; Sensitivity: 0.03 ng/mL). 21 –23 LH and PRL levels were evaluated by ultrasensitive ELISA, as previously described. 24,25 Plasma E2 and P4 levels were evaluated using the DRG kit (E2 ELISA, EIA-2693; DRG Instruments GmbH, Hamburg, Germany) and rat progesterone kit (P4 Elisa, RTC008R; Biovendor Research and Diagnostics Products, Brno, Czech Republic), respectively, following the manufacturer's instructions. For each ELISA, all samples were assayed in one assay.
IHC of Kp neurons
Brain processing and IHC were performed, as previously described. 26 The analysis of the images was blind using the ImageJ software (National Institutes of Health, USA). The number of Kp-immunoreactive (ir) neurons in the ARC was bilaterally quantified in four sections per animal, approximately between −2.4 and −3.96 mm of the bregma. The integrated optical density (IOD) of Kp-ir fibers was also quantified in the ARC. The IOD was calculated and subtracted from the background staining using the ImageJ software.
Quantitative real-time reverse transcriptase-polymerase chain reaction
Microdissections of the brains were performed using the punch technique, and quantitative real-time reverse transcriptase-polymerase chain reaction (qPCR) analyses of messenger RNA (mRNA) levels were performed as previously described. 25 The initiators were delineated upon Rattus norvegicus mRNA sequence, and the amplification efficiency was analyzed for each gene (Table 1; Supplementary Figs. S1 and S2). Gene expression was analyzed by the 2−ΔΔCT method; 27 the results obtained for each group being quantitatively compared after normalization based on the expression of glyceraldehyde 3-phosphate dehydrogenase (Gapdh) for hypothalamus and pituitary and ribosomal protein L19 (Rpl19) for corpus luteum.
List of Genes and Nucleotide Sequences for Quantitative Real-Time Reverse Transcriptase-Polymerase Chain Reaction Primers
F, forward primer; R, reverse primer.
Statistical analysis
Data are presented as mean ± standard error of the mean. The data were tested for normality (Shapiro–Wilk) and homoscedasticity (Brown–Forsythe) of the errors, and the differences between the three groups were determined by one-way analysis of variance, followed by the Student–Newman–Keuls test for multiple comparisons, using GraphPad Prism 8.0.2 software®, while the differences between the two groups in the IHC analysis were determined by the unpaired Student t-test. In all analyses, the differences were considered statistically significant if p < 0.05.
Results
Confirmation of hypothyroidism
Hypothyroidism was confirmed by the reduction of fT4 levels and increase of plasma TSH in rats of the hypothyroid group and the group treated with Kp10 compared with the control group (Fig. 1A, B; *p < 0.05, **p < 0.01), in addition to the reduction of body weight throughout the experiment (Fig. 1C; *p < 0.05).
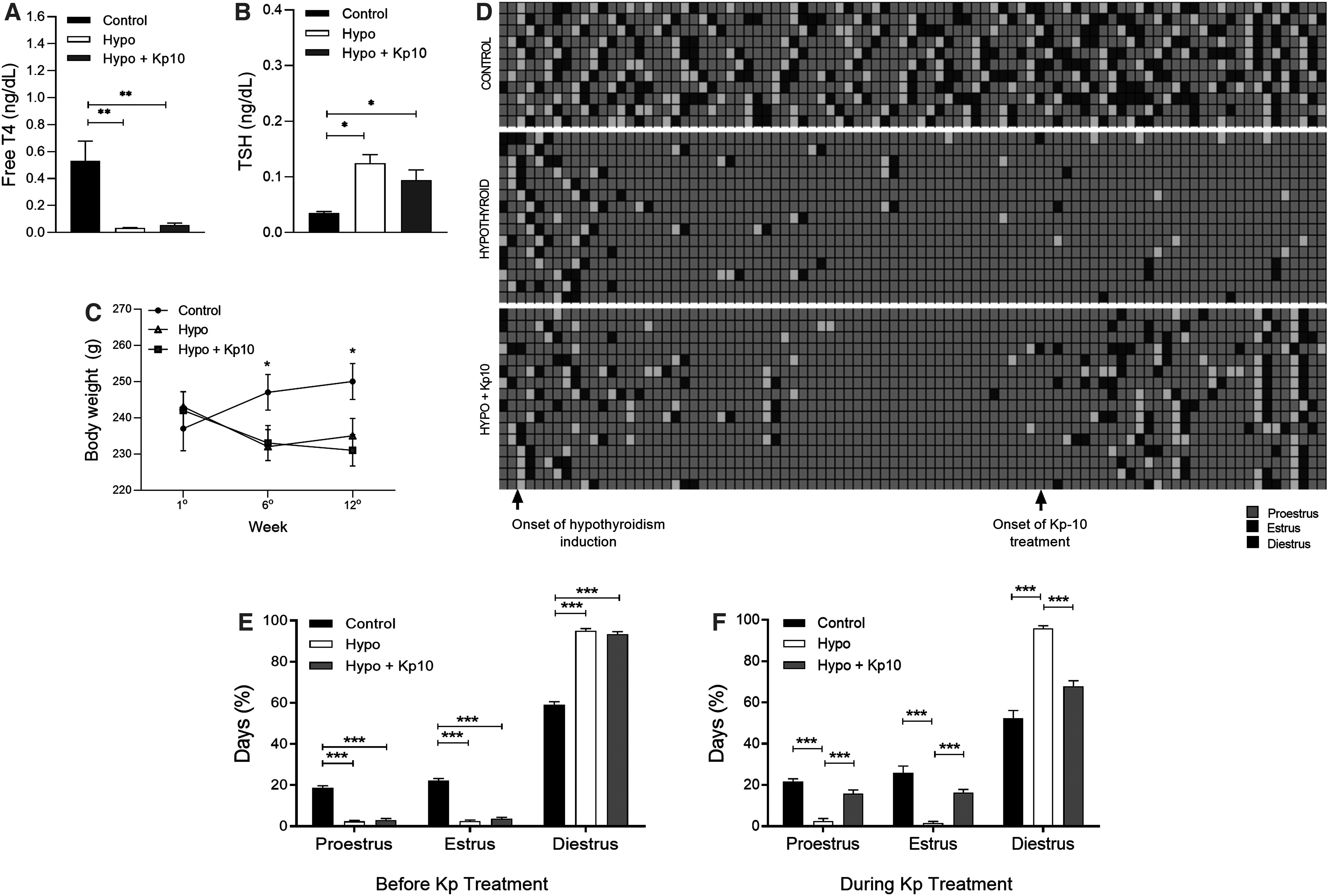
Confirmation of hypothyroidism and recovery of cyclicality in hypothyroid rats after treatment with Kp10. (
Kp10 treatment restored cyclicity in hypothyroid rats
After the onset of hypothyroidism, hypothyroid rats began to exhibit irregular cycles (Fig. 1D), with a lower percentage of days on proestrus and estrus compared with the control group, remaining predominantly acyclic and on diestrus until the end of the third month (Fig. 1D, E; ***p < 0.001). However, in the last month, following the start of Kp10 treatment, rats returned to regular cycles 8 to 9 days after initiation of treatment (Fig. 1D), as confirmed by the increased number of days on proestrus and estrus, and the reduced days on diestrus in relation to the hypothyroid group (Fig. 1D, F; ***p < 0.001), equaling the control (p > 0.05).
Kp10 treatment restored ovarian function and uterine morphology in hypothyroid rats
In the macroscopic evaluation of the ovaries, hypothyroid rats displayed lower ovarian weight (Fig. 2A–G; ****p < 0.0001) and reduced number of corpora lutea (Fig. 2H; ***p < 0.001) compared with control rats, while treatment with Kp10 increased ovarian weight (Fig. 2G; ***p < 0.001) and restored the number of corpora lutea to the control levels (Fig. 2H; **p < 0.01).
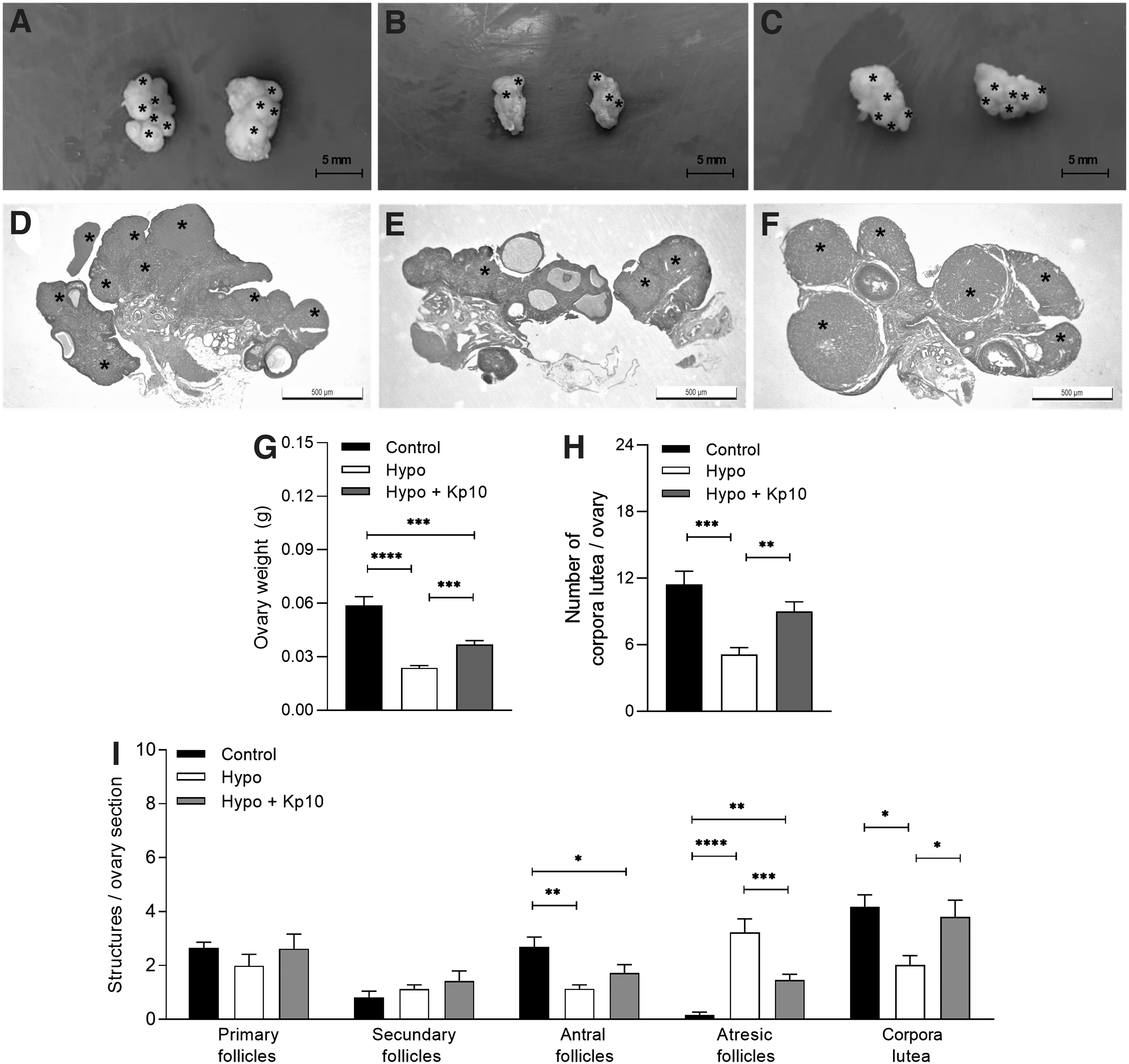
Kp10 treatment restores ovarian morphology in hypothyroid rats. (
Histomorphometry of the ovaries in serial cuts was performed and hypothyroidism reduced the number of antral follicles and increased follicular atresia (Fig. 2I; *p < 0.05, **p < 0.01, ****p < 0.0001), in addition to reducing the number of corpora lutea per ovarian section. Treatment with Kp10 caused a significant reduction in the number of atretic follicles and restored the number of corpora lutea the control levels (Fig. 2I; *p < 0.05, ***p < 0.001). However, Kp10 did not restore the number of antral follicles. Regarding the primary and secondary follicles, no differences were observed between the groups (Fig. 2I; p > 0.05).
Hypothyroidism did not affect the Kiss1 luteal expression (Fig. 3A; p > 0.05); however, it increased the mRNA expression of Cyp11a1 and reduced 20aHsd mRNA compared with the control group (Fig. 3C, F; ***p < 0.001, *p < 0.05), while a trend toward increase was observed for Star mRNA (Fig. 3B; p = 0.05). Interestingly, the treatment with Kp10 reduced the mRNA levels of Cyp11a1 and 3bHsd in the corpora lutea of hypothyroid rats, while it increased 20aHsd mRNA expression (Fig. 3C, E, F; p < 0.05), showing no significant differences in relation to the control group (p > 0.05). Regarding Nr5a1 mRNA, no differences were observed among the experimental groups (Fig. 3D; p > 0.05).
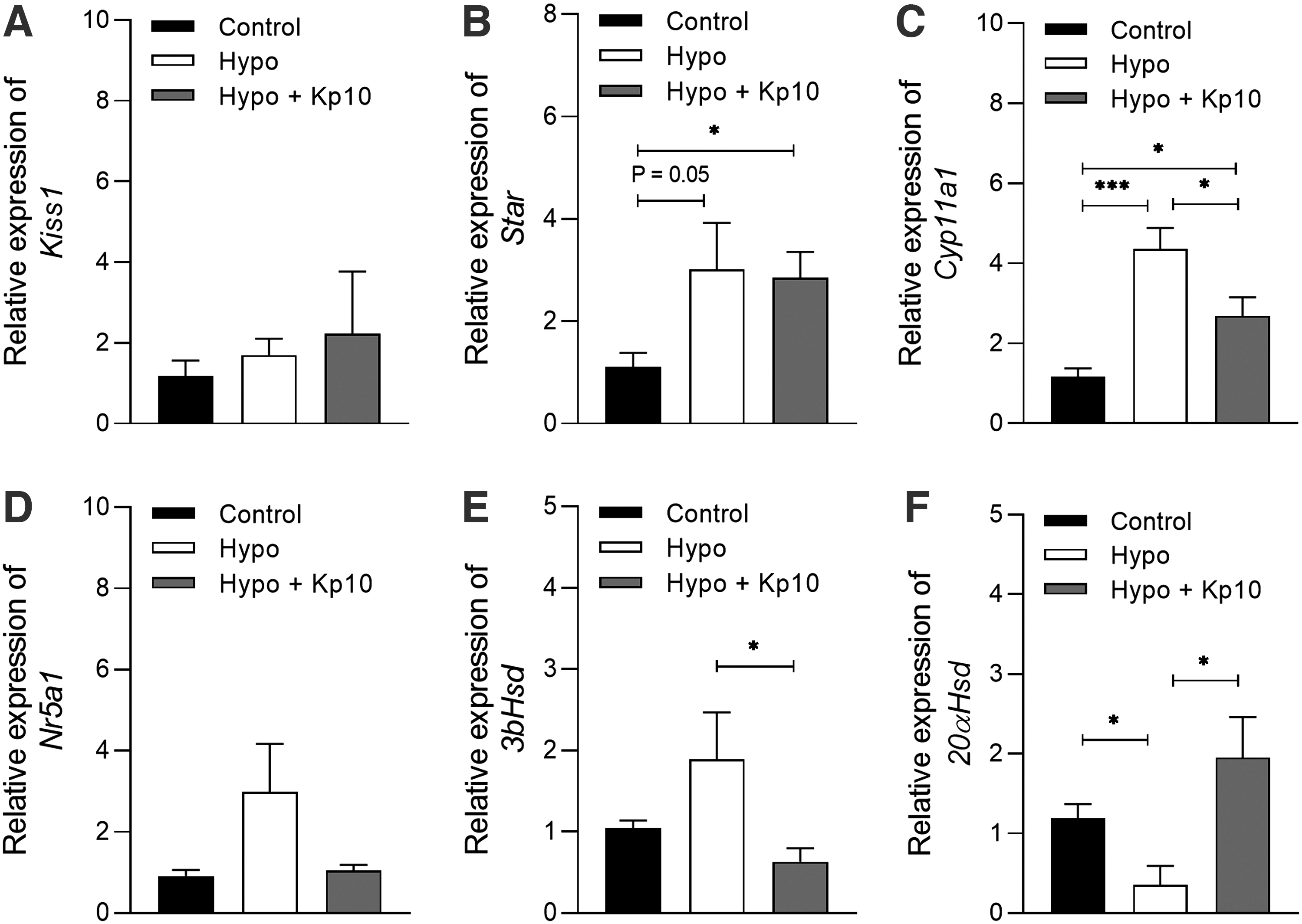
Kp10 treatment partially restores luteal gene expression of steroidogenic and transport proteins in hypothyroid rats. (
In the uterine evaluation, the hypothyroid group presented lower endometrial thickness and reduced number of endometrial glands compared with the control (Fig. 4A–E; **p < 0.01, ****p < 0.0001), while treatment with Kp10 restored endometrial thickness and the number endometrial glands (Fig. 4A–E; *p < 0.05, ****p < 0.0001), equaling those of control animals (p > 0.05). Regarding myometrium, no differences in thickness between groups were observed (Fig. 4F; p > 0.05).
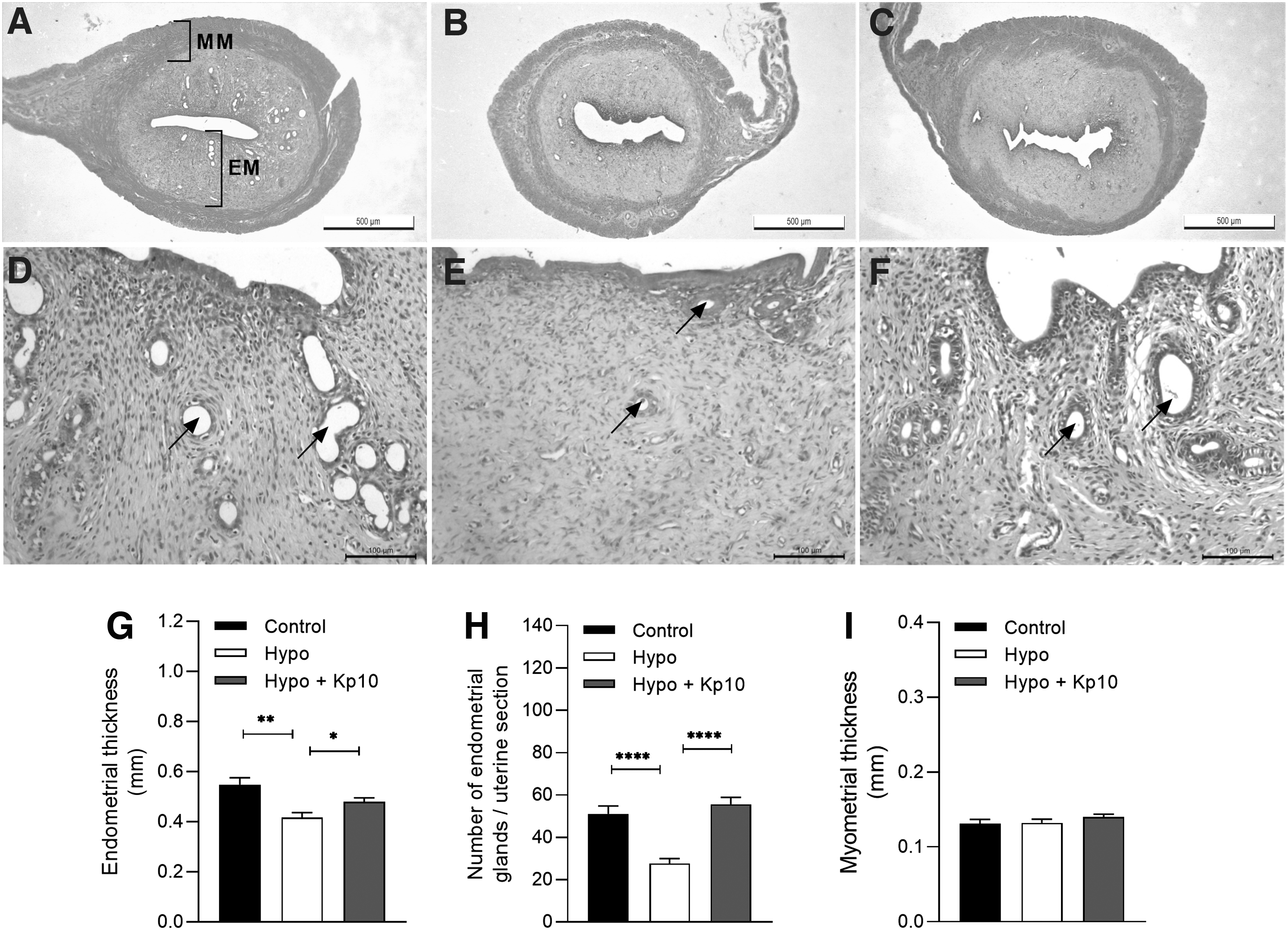
Kp10 treatment restores uterine morphology in hypothyroid rats. (
Kp treatment restored plasma LH but did not reverse the hyperprolactinemia and hyperprogesteronism caused by hypothyroidism
Both the hypothyroid group and the group treated with Kp10 presented higher plasma PRL and P4 levels when compared with the control group (Fig. 5A, D; *p < 0.05, ****p < 0.0001). Moreover, the hypothyroidism reduced the plasma LH level relative to the control group (Fig. 5B; **p < 0.01), while the treatment with Kp10 increased LH levels (Fig. 5B; *p < 0.05), equaling to those of the control group (p > 0.05). No significant differences were observed between the groups regarding the plasma concentration of E2 (Fig. 5C; p > 0.05).
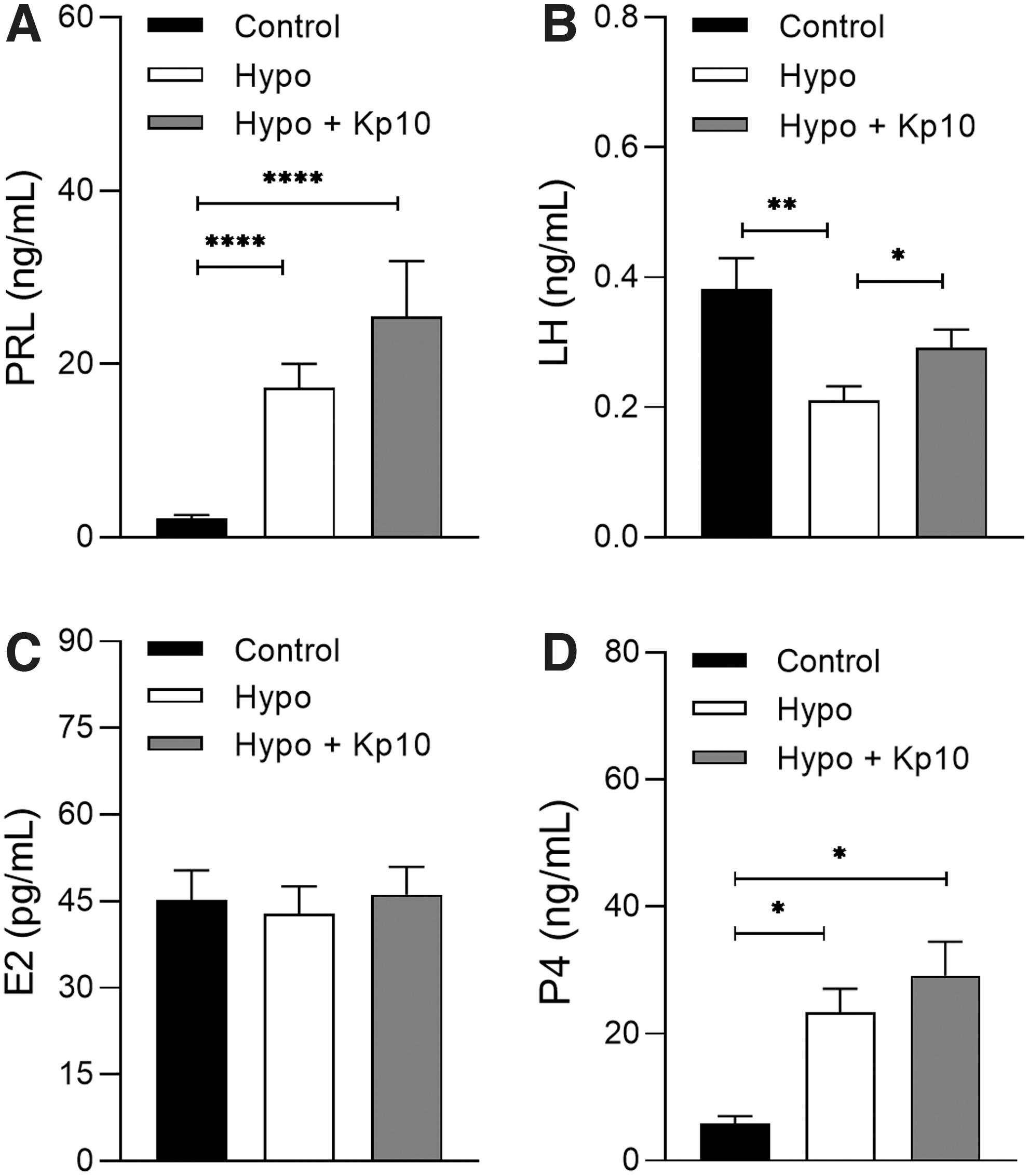
Plasma PRL, LH, E2, and P4 levels in control, hypothyroid, and hypothyroid rats treated with Kp10. (
Kp10 treatment restored the pituitary expression of Lhb and Tshb in hypothyroid rats
Hypothyroid rats showed a reduction in the pituitary expression of mRNA for Lhb compared with the control group (Fig. 6A; **p < 0.01), while the Kp10 treatment not only restored pituitary expression of Lhb (Fig. 6A; ****p < 0.0001) but also resulted in a higher level of transcripts compared with the control group (*p < 0.05). Regarding mRNA expression for Fshb, there was no difference between the groups (Fig. 6B; p > 0.05), while both the hypothyroid group and the Kp10-treated group showed significant reduction in Gnrhr transcripts compared with the control group (Fig. 6C; **p < 0.01).
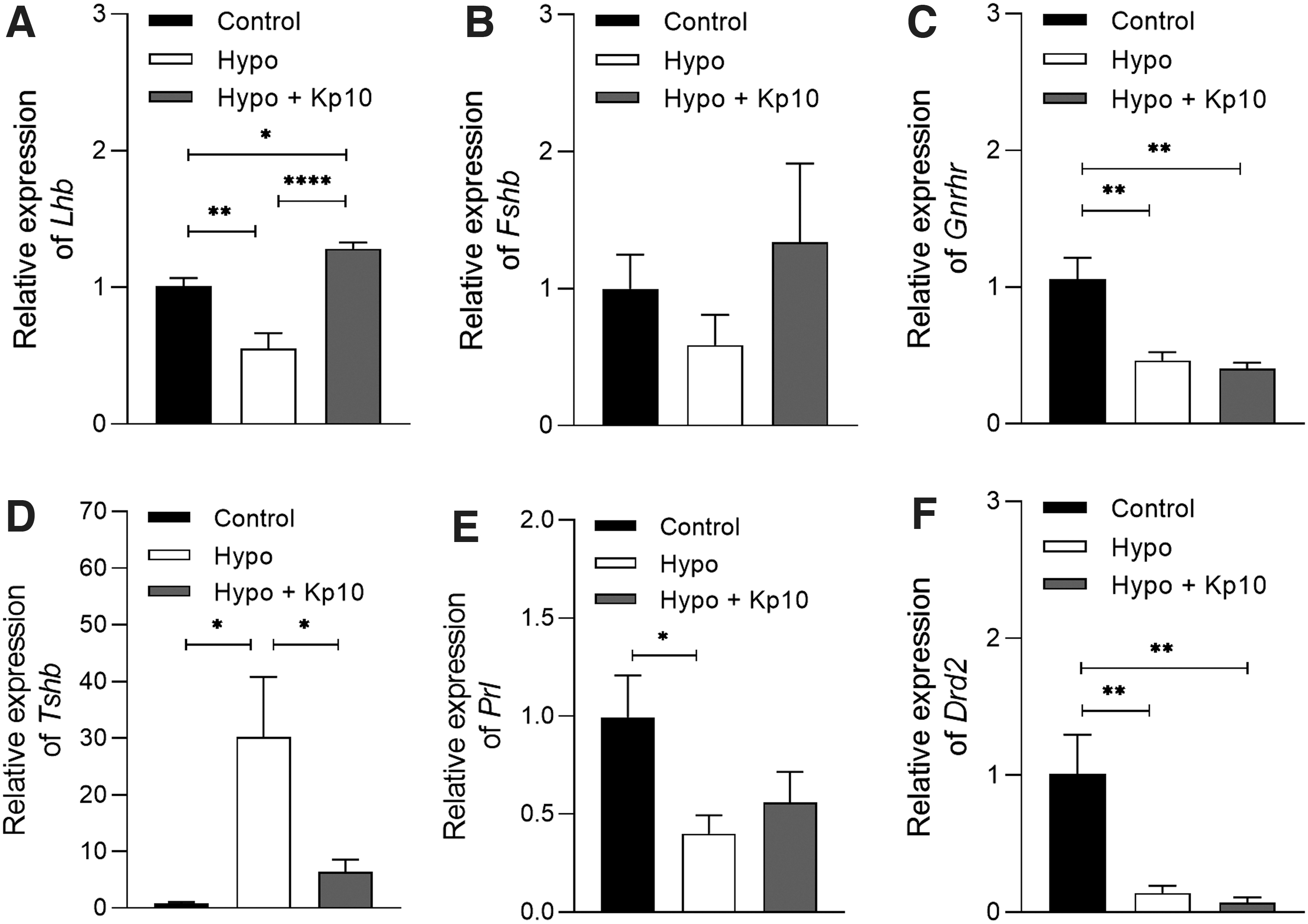
Effect of Kp10 on the pituitary gene expression of Lhb, Fshb, Gnrhr, Tshb, Prl, and Drd2 in hypothyroid rats. (
Regarding Tshb, as expected, hypothyroidism increased mRNA expression compared with the control (Fig. 6D; *p < 0.05), while the Kp10 treatment reduced the increase in gene expression caused by hypothyroidism (Fig. 6D; *p < 0.05), toward control levels (p > 0.05). Hypothyroid rats also showed a reduction in the pituitary expression of Prl mRNA compared with the control (Fig. 6E; *p < 0.05), while the treatment with Kp10 showed no significant differences to the control or hypothyroid groups. Interestingly, regarding transcripts expression for Drd2, both the hypothyroid group and the Kp10-treated group showed significant reduction compared with the control group (Fig. 6F; **p < 0.01).
Hypothyroidism reduced the hypothalamic expression of Kp and NKB
Hypothyroidism reduced the number of Kp-ir neurons in the ARC compared with the control group (Fig. 7A–C; ***p < 0.001) as well as the IOD of Kp immunoreactivity (Fig. 7D; **p < 0.01). Through the analysis of qPCR, we also verified that hypothyroidism reduced the gene expression of Kiss1 (Fig. 7E; ***p < 0.001) and Nkb (Fig. 7F; ***p < 0.001) in the ARC compared with the control group, while no effect was observed in Prodyn expression (Fig. 7G; p > 0.05). Treatment with Kp10 did not alter the gene expression of Kiss1, Nkb, or Prodyn in the ARC of hypothyroid rats (Fig. 7E–G; p > 0.05).
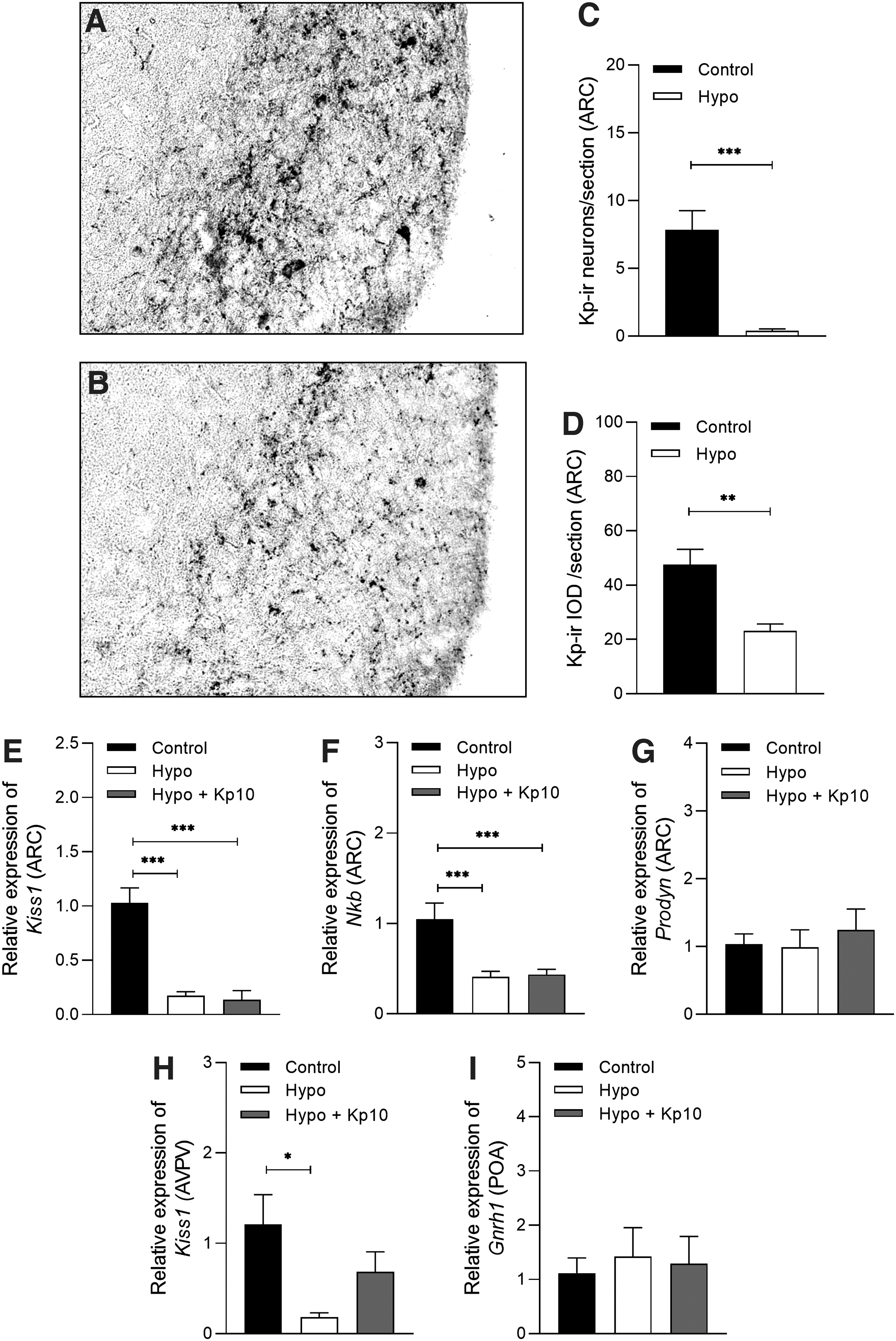
Effect of hypothyroidism on Kp immunoreactivity and gene expression in KNDy and GnRH neurons. (
Although the animals were euthanized on diestrus, we also evaluated the mRNA expression for Kiss1 in the AVPV, as the other major population of Kp neurons. Likewise to the ARC, hypothyroidism reduced the expression of Kiss1 transcripts in AVPV compared with the control group (Fig. 7H; *p < 0.05). Interestingly, the treatment with Kp10 partially increased Kiss1 levels in the AVPV of hypothyroid rats, which were not statistically different from either the control or hypothyroid groups (Fig. 7H; p > 0.05). The levels of mRNA for Gnrh1 in the POA did not differ between the groups (Fig. 7I; p > 0.05).
Discussion
This study demonstrated that treatment with Kp10 restores ovarian function of rats with chronic hypothyroidism, as it was able to restore cyclicity, ovulatory rate, luteal steroidogenesis, uterine morphology, and plasma LH levels, independent of hyperprolactinemia. In addition, the inhibitory effects of hypothyroidism on the gonadotropic axis occur not only by hypothalamic suppression of Kiss1 mRNA and peptide but also by suppression of Nkb gene transcripts in KNDy neurons, suggesting that the suppression of ovarian function caused by hypothyroidism is associated with a failure in both Kp and neurokinin expression at hypothalamic level.
Hypothyroidism kept the rats predominantly acyclic. Furthermore, it reduced endometrial thickness, the number of endometrial glands and ovarian weight, as well as the number of corpora lutea and antral follicles, while increased follicular atresia, even without altering plasma E2 levels. All these results are in agreement with previous studies that also demonstrated irregular estrous cycle, 15,28 reduced endometrial thickness, 29,30 reduced folliculogenesis and ovulation, 30 –32 and increased follicular atresia. 30,32 The plasma E2 results also corroborate other studies on hypothyroidism that, similar to this, carried out evaluations with rats on the day of diestrus. 33
Although hypothyroidism did not affect Kiss1 luteal gene expression, it increased the expression of transcripts for Star and Cyp11a1, key transport and steroidogenic proteins in the corpus luteum, while decreasing 20αHsd, the enzyme responsible for P4 catabolism. 34 In addition, it increased plasma levels of P4. These results are in agreement with previous studies that demonstrated increased P4 and steroidogenic activity, decreased 20aHSD mRNA and protein expression, and longer life span of the corpus luteum in hypothyroid rats. 17,18,33,35
The treatment with Kp10 was able to restore the uterine morphology and the number of corpora lutea and reduce follicular atresia caused by hypothyroidism, even in the presence of hyperprolactinemia. Fernandois et al 36 showed that Kp intraovarian administration increased the number of preovulatory follicles and corpora lutea in aging rats. Furthermore, the Kp10 administration was able to reduce the luteal gene expression of Cyp11a1 and 3βHsd in hypothyroid rats and restore the expression of 20aHsd, favoring the adequate steroidogenesis of the corpus luteum and return of the reproductive cycle 35 as evidenced by the regular estrous cycles in hypothyroid animals concomitantly with Kp10 treatment.
Recently, Chiang et al 37 demonstrated that the in vitro administration of Kp10 in caprine luteal cells reduced the protein and mRNA expression of StAR, Cyp11a1, and 3βHSD, corroborating the results of the present study. However, although the treatment with Kp restored the gene expression of 3βHsd and 20aHsd in the corpus luteum of hypothyroid rats to control levels, it was not able to reduce P4 secretion. This was an expected result since Kp has been shown to stimulate the in vitro production of P4 in luteal cells of rats. 38
The treatment with Kp10 also increased plasma LH to values similar to that of the control group, including the pituitary mRNA for Lhb, which was higher than that of the control group. This result suggests a central action of systemic administration of Kp10 for the return of ovarian function of hypothyroid rats, even though no difference in Fshb expression was observed between the groups. A similar result was observed in a hyperprolactinemic mouse model, which recovered the ovarian function and cyclicity after systemic treatment with Kp10. 39 Studies suggest that this increased Lhb is a reflection of the hypothalamic action of Kp10 stimulating the release of GnRH 40,41 and especially its direct action on the pituitary, as in vitro incubation of Nile tilapia (O. niloticus) pituitary cells with Kp10 increased mRNA levels for Lhb, 42 as also observed in primary culture of mouse pituitary cells and in Lßt2 gonadotropic cells. 41
However, the observed increase in the transcripts to Lhb after treatment with Kp10 was not associated with an increase in Gnrhr expression. This result was not expected since Jonak et al 43 demonstrated that GnRH induces the expression of the GnRHr in the gonadotropes, and Sukhbaatar et al 44 have shown that Kp is able to stimulate GnRHr expression in GnRH-producing GT1-7 cells. However, although we did not observe alterations in the Gnrhr gene transcription after treatment with Kp10, alterations in protein translation cannot be ruled out, as it was not possible to be analyzed in the present study.
Noteworthy, the return of plasma LH levels and cyclicity in hypothyroid rats treated with Kp10 occurred in the presence of hyperprolactinemia. This was actually the result expected because previous studies had shown that the administration of Kp10 increases PRL secretion in female rats and mice. 45,46 Moreover, the reduction in Prl mRNA at the pituitary observed in hypothyroid rats can be a consequence of hyperprolactinemia since prolactin (PRL) exerts negative feedback effect on its own secretion at the transcriptional level. 44 Furthermore, we suggest that the hyperprolactinemia observed in both hypothyroid and Kp10-treated rats may be due not only to the increase in the TSH-releasing hormone release observed in hypothyroidism, which has a stimulatory effect on pituitary lactotrophs 47,48 but also to the lower pituitary gene expression of Drd2 found in these animals, given that hypothalamic dopamine tonically suppresses the synthesis and secretion of PRL acting through this receptor. 49,50
Interestingly, Kp10 treatment reduced the higher pituitary Tshb expression caused by hypothyroidism, demonstrating that although Kp did not change plasma levels of fT4 and TSH levels, it can influence the Tshb expression at the anterior pituitary. The increase in Tshb transcription in hypothyroid rats was already expected since the inhibition triggered by thyroid hormones on these transcriptional and post-transcriptional steps is reduced when the levels of thyroid hormones are lower. In addition, thyroid hormones also have nongenomic actions by altering the arrangement of the cytoskeleton of anterior pituitary cells, while in hypothyroidism the poly(A) tail length of Tshb mRNA is increased, followed by a rise in the translational rate of Tshb. 51 The length of a poly(A) tail correlates with mRNA translational efficiency. 52
We also demonstrated that the suppression of ovarian function in the hypothyroid rats was accompanied by a reduction of the immunoreactive neurons for Kiss1 in the ARC and in mRNA expression for Kiss1 in the ARC and AVPV in comparison to the control group, although this reduction was not altered by treatment with Kp10. Similar results have been found in female mice treated with triclosan, an antibiotic that affects thyroid function, which showed reduced Kiss1 expression in ARC and AVPV, 5 as also observed by Tomori et al 15 in the ARC of hypothyroid rats.
This effect of hypothyroidism on hypothalamic Kp suppression is possibly related to increased PRL, as observed in the present study, since PRL suppresses Kp expression in the ARC of ovariectomized rats. 25,26 However, it is not possible to rule out the direct effect at hypothalamic level of thyroid hormone deficiency, as Dufourny et al 6 demonstrated that Kiss1 neurons in the hypothalamus of sheep have thyroid hormone receptors alpha (Thra). Moreover, it has already been shown that GnRH neurons of hamsters express mRNA for Thra. 53
We also showed that hypothyroidism significantly reduced the gene expression of Nkb in KNDy neurons, even without altering the expression of Prodyn, while treatment with Kp10 did not affect the expression of these genes. Kp and NKB have emerged as the primary regulators of GnRH and LH secretion. 7,13,53,54 Patients with loss of function mutations in Kp, NKB, and their respective receptors (KISS1R and NK3R) have a delay in reaching puberty. 47 –49,55 Inhibition of LH pulse and/or secretion using NKB receptor antagonists were also observed in ovariectomized sheep and nonhuman primates. 56,57 It is plausible that the dysregulation in mRNA expression for Nkb observed in rats with thyroid hypofunction of the present study may also be involved in the pathogenesis of ovarian dysfunction observed in hypothyroidism.
Interestingly, the levels of mRNA for Gnrh1 in POA were not affected by thyroid hypofunction, as also observed in previous studies, 58,59 or by treatment with Kp10, suggesting that GnRH suppression occurred at the level of protein translation or in the release by afferent fibers, although more studies are needed to elucidate this hypothesis.
As a limitation of the present study, we have not compared the use of Kp10 with thyroxine replacement, which is the main clinical treatment for patients with hypothyroidism. In addition, further studies are needed to evaluate the dopaminergic axis of hypothyroid rats treated with Kp10 to elucidate the effect of Kp on the hyperprolactinemic state caused by thyroid hypofunction.
The findings of the present study characterize the inhibitory effects of hypothyroidism induced by PTU treatment on hypothalamic control of the gonadal axis in rats, mainly in the expression of Kp and neurokinin in KNDy neurons, and demonstrate that treatment with Kp10 is able to reverse ovarian dysfunction caused by hypothyroidism, regardless of hyperprolactinemia.
Footnotes
Acknowledgment
The authors thank Ivo Arouca (Universidade Estadual de Santa Cruz, Ilheus, Brazil) for the technical support provided.
Authors' Contributions
L.S.O., T.Q.M.S., E.M.B., and J.F.S. conceived and designed the study. L.C.S., T.Q.M.S., E.M.B., J.M.A.C., B.R.S., and I.O.M. conducted sample collection. L.C.S., T.Q.M.S., and E.M.B. conducted histopathological preparation and analysis. P.C.H., D.O.G., and R.E.S. conducted immunohistochemical preparation and analysis. P.C.H., D.O.G., and R.E.S. performed ELISA. L.S.O., L.C.S., J.M.A.C., and J.F.S. performed qRT-PCR. L.S.O. and J.F.S. analyzed the data. J.F.S. and R.E.S. provided reagents. L.S.O., L.C.S., R.E.S., and J.F.S. wrote the article.
Data Availability
The data underlying this article will be shared on reasonable request to the corresponding author.
Author Disclosure Statement
The authors declare no conflicts of interest.
Funding Information
This work was supported by Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq; grant number: 408144/2018-1) and the Universidade Estadual de Santa Cruz (UESC).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2