
Abstract
Background:
Genomic deletions in medullary thyroid cancer (MTC) are rare. Selpercatinib is a highly selective RET inhibitor for treatment of metastatic RET-altered MTC. We report a 35-year-old male with an aggressive metastatic MTC harboring p.632_633del RET that was poorly responsive to RET kinase inhibitor selpercatinib.
Objective:
Our objective was to understand the clinical phenotype of p.632_633del RET in MTC in the context of novel RET kinase inhibitor treatment.
Methods:
Wild-type and p.632_633del RET sequences were modeled using a lighter version of the AlphaFold2 (AF2) software. Functional studies were performed on transfected HEK 293 cells (pCMV6-Entry, pCMV6-RET, or pCMV6-RET(p.632_633del) treated with inhibitors for 24 hours and analyzed on luciferase assays.
Results:
Structural modeling revealed a paucity of disulfide bridge between Cys630-Cys634 in p.632_633del RET sequences, apparent in wild-type, while forming an intermolecular disulfide bridge between two Cys656. Proximity of juxtamembrane segments of each dimer may impede Tyr687 phosphorylation and stable conformation of intracellular RET that hosts selpercatinib. In vitro experiments confirmed a reduction in efficacy of selpercatinib upon p.632_633del RET compared with wild-type RET control.
Conclusion:
Clinical presentation together with structural modeling and functional studies suggests that p.632_633del RET results in poor response to selpercatinib.
Introduction
Medullary thyroid cancer (MTC) is a neuroendocrine tumor derived from C cells in the thyroid gland and it accounts for 1–2% of all thyroid cancer cases. 1 MTCs may be sporadic or associated with multiple endocrine neoplasia type 2 (MEN2). 2 The REarranged during transfection (RET) proto-oncogene is located on chromosome 10q11.2 and encodes a single transmembrane receptor of the tyrosine kinase family. RET is a receptor for neurotrophic factors of the glial cell line derived from neurotrophic factor (GDNF) family. 3 Binding is facilitated by lipid-anchored proteins termed GDNF family receptor alpha (GFRα).
GDNF and GFRα stimulation activates the RET receptor by dimerization and subsequent autophosphorylation of intracellular tyrosine residues on the kinase domains. Phosphorylation within the kinase domains results in the formation of binding sites for adapter proteins that mediate the activation signal to cascade pathways. 4 Mutations identified in the extracellular domains cause activation of RET even in the absence of ligand stimulation. 5
Somatic RET mutations are seen in 50% of sporadic MTCs, and are more frequent in larger tumors. 6 –8 The commonest mutation RETM918T is associated with an aggressive clinical course and poor prognosis. 9 Genomic RET deletions are found in several malignancies including esophageal adenocarcinoma and ovarian cancer. 10 RET deletions are uncommon in MTC, seen in ∼9–17% of patients with sporadic RET-mutated MTC (Supplementary Table S1). 8,11 –15
The p.632_633del RET is produced from a 6 base pair (bp) in-frame deletion in exon 11 [NM_020975.4(RET):c.1894_1899delGAGCTG (
Calcitonin and carcinoembryonic antigen (CEA) are the current gold standard biochemical markers to monitor biochemical response in MTC. In recent years, the development of liquid biopsy, that is, sampling and analysis of blood or other bodily fluids for informative biomarkers, such as circulating cell-free DNA (cfDNA), microRNAs (miRNAs), proteins, and metabolites, 19 has been found to be a powerful clinical tool for several cancers. The role of ctDNA in thyroid cancer is evolving with increasing capability to detect RET mutations. 19 The ability to detect mutations using liquid biopsy allows for initiation of targeted therapies, monitoring treatment response, and detecting new mutations associated with emergent treatment resistance.
We describe the clinical phenotype and course of a patient whose MTC harbored somatic p.632_633del RET who progressed on selpercatinib. We tested the hypothesis that the mutation creates resistance by using structural modeling and functional studies to examine whether p.632_633del RET hinders binding of selpercatinib.
Case
A 35-year-old man presented with a 9-month history of worsening right-sided flank pain. He had no significant medical history or noteworthy family history. Calcitonin was elevated at 36 pmol/L (reference range [RR] <4) and CEA was 183.4 μg/L (RR ≤3). 68GaDOTATATE PET imaging demonstrated mild uptake in the T12 vertebral body, right transverse process of T7, and moderate tracer uptake (SUVmax 12.1) on the seventh rib. No uptake was seen on an FDG PET scan. Total thyroidectomy with modified radical right neck dissection and right seventh rib resection was performed.
Histology confirmed a 21 × 21 × 15 mm right thyroid MTC without nodal involvement but with tumor necrosis and a Ki67 proliferative index of 8%, indicating high-grade disease (Fig. 1). 20,21 Pathological analysis of the resected seventh rib confirmed metastatic MTC. Stereotactic body radiation therapy was used to target the T7 and T12 vertebral lesions.
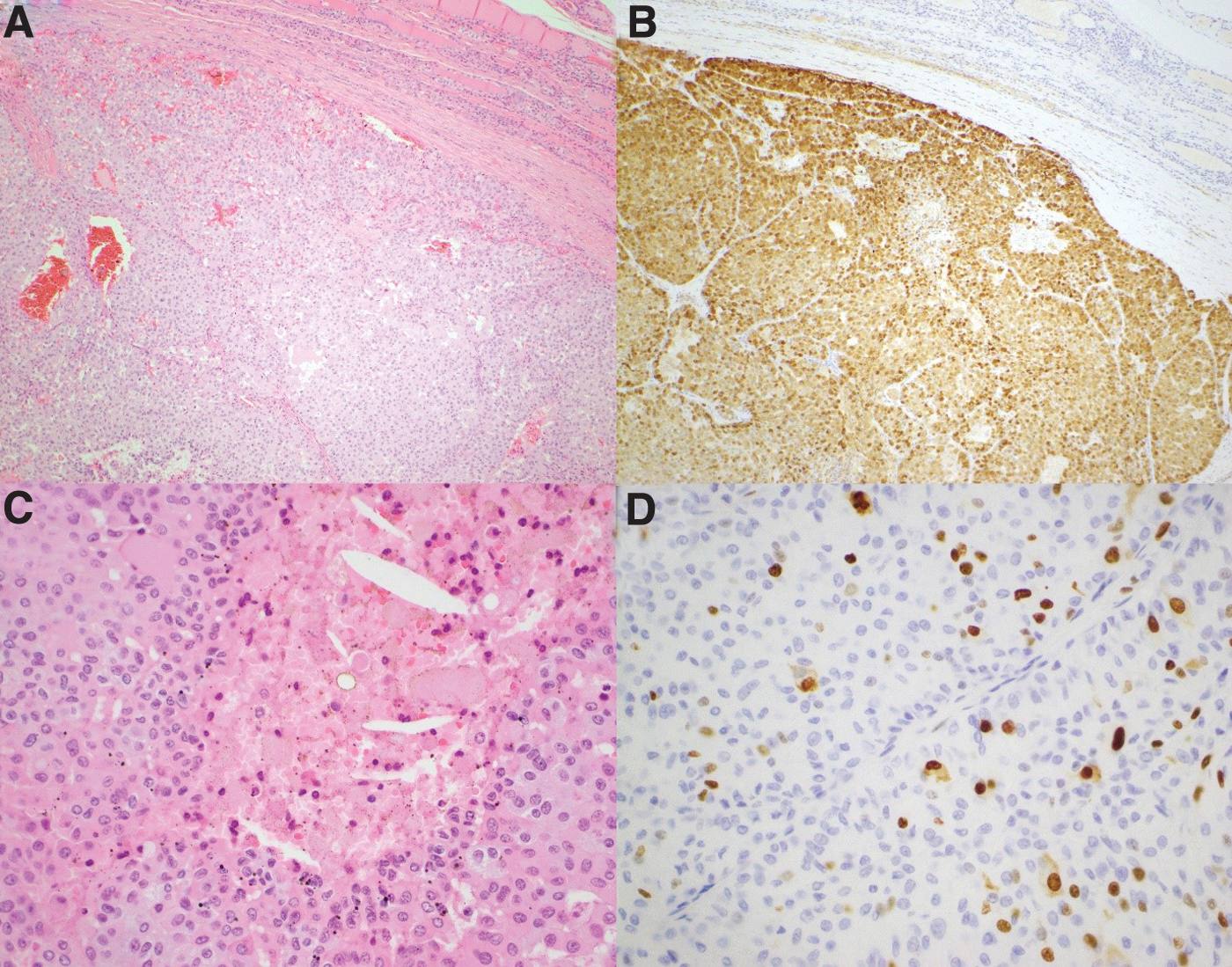
Serial H&E (
Somatic mutation testing was performed by next-generation sequencing (NGS) of the macrodissected tumor sample, using a custom gene panel on an Illumina MiSeq and detected a pathogenic heterozygous RET variant c.1894_1899delGAGCTG, (
Calcitonin and CEA were undetectable 2 months postoperatively. Progress imaging with CT chest, abdomen, and pelvis identified a 19 mm lytic lesion in T12 vertebral body posteriorly and superiorly and an increase in radiodensity of the 7th and 8th thoracic vertebral bodies. A new ill-defined hypoattenuating lesion of 10.7 mm was noted in the 4th liver segment. All bone lesions were 68Ga-DOTATATE avid. Calcitonin reached a peak of 55.6 ng/L (RR <9) and CEA 60.5 μg/L (RR <5) 18 months after diagnosis.
Eighteen months after diagnosis, selpercatinib was commenced at 160 mg BD as part of the LIBRETTO-001 trial. 12 The dose was maintained despite a photosensitive rash (which resolved with reduced sun exposure), mild pleural effusions, and a one month drug interruption after a nasal septum perforation. Calcitonin decreased to 2 ng/L and CEA to 3.2 μg/L within 3 months of starting selpercatinib. In accordance with RECIST 1.1, a partial response was documented at cycle 9 (a single cycle duration being 4 weeks) of treatment, however, by cycle 13 there was progressive disease with increased size of liver lesions (Supplementary Table S2).
After cycle 28, a new left ilium lesion noted on 68Ga-DOTATATE PET imaging was treated with radiation therapy. After 12 months on selpercatinib, further progression was noted on CT chest, abdomen, and pelvis with new lymphadenopathy, liver lesions, and lytic lesions in T4, 8,9,10, 11, and 12 and L1. CEA was 9 μg/L and calcitonin 10 ng/L. ctDNA extracted at the time from plasma and analyzed on Thermo Fisher Ion Torrent NGS panel (Oncomine™ Pan-Cancer Cell-Free Assay) detected NM_020975.4(RET):c.1894_1899delGAGCTG (
No new mutation was detected on the serial sampling; however, the panel was unable to assess for fusion mutations. Selpercatinib was ceased with an intent to start cabozantinib; however, with worsening progressive metastatic disease complicated by pulmonary embolism, the patient died before commencing this treatment.
Materials and Methods
The protein sequence of human RET was retrieved from Uniprot database (Id P07949), an online resource for protein sequence and functional information. Deletion of residues Glu632 and Leu633 were found as FASTA file in the database. Wild-type and p.632_633del RET sequences were modeled using a lighter version of the AlphaFold2 (AF2) software 22 installed on a ColabFold notebook. 23 The option “Use templates” was used.
Limitation of calculation time on ColabFold notebook required to cut RET sequence in 3 subsequences, in accordance with the RET domains (Supplementary Fig. S2A): 4 Cadherine-like Domains (CDL), Cysteine Rich Domain (CRD), Transmembrane Domain (TMD), Juxtamembrane domain (JM), Tyrosine Kinase Domain 1 (TK1), and Tyrosine Kinase Domain 2 (TK2). The 3 subsequences are (1) the extended extracellular RET that includes 4 CDL, CRD, and TMD corresponding to residues from 1 to 663; (2) the extracellular RET that includes CRD and TMD corresponding to residues from 508 to 663; and (3) the intracellular RET that includes TMD, JM, TK1, and TK2 domains corresponding to residues from 621 to 1015. For each subsequence, both the RET monomer and the dimer were built.
Functional studies were performed on HEK293 cells as follows. The day before treatment, HEK293 cells were plated in Opti-MEM (Thermo Fisher Scientific, Waltham, MA) containing 0.1% FCS, in a white 96-well microplate at a density of 3 × 104 cells per well. These cells were transfected with 40 ng pFR-Luc; 2 ng pFA2-Elk1 (Agilent Technologies, Santa Clara, CA); 2 ng pRL-CMV (Promega, Madison, WI); and 40 ng of either pCMV6-Entry, pCMV6-RET, or pCMV6-RET(p.632_633del) (Origene, Rockville, MD) per well with XtremeGene HP transfection reagent (Roche, Basel, Switzerland).
The cells were treated with inhibitors (Sellekchem, Houston, TX) or DMSO (Sigma-Aldrich, St Louis, MO) for 24 hours and then luciferase assays were performed using Promega's Dual-Glo Luciferase regents as per the manufacturer's instructions.
The study was approved by the human research ethics committee at Northern Sydney Local Health District (ETH00672/STE001486) and Royal North Shore Hospital Institutional Biosafety Committee (IBC/22/002). Approval from trial sponsor (Eli Lily and Company) was obtained to publish the case details.
Results
Validation of modeling quality
Structure prediction of the extended extracellular domains (4CDL, CRD, and TMD) with AF2 provided in high agreement with the available crystal structures in the Protein Data bank (PDB code 4UX8 and 6GL7). Modeling of the intracellular domain (TM, JM, TK1, and TK2) with AF2 also led to models in high agreement with available crystal structures (7JU6 and 7JU5). TMD and JM were modeled without using templates as crystal structures were currently available for these domains (Supplementary Fig. S2).
Modeling of the extracellular domain RET monomer
In the wild type, the two cysteine residues Cys630 and Cys634 that surround the Glu632 and Leu633 residues make an intramolecular disulfide bridge (Supplementary Fig. S3A). These predictions are in accordance with Santoro et al. 24 The intramolecular covalent bond is extremely rigid and leads to a hinge bending of the CRD with respect to TMD. Contrarily to the wild type, the two cysteines Cys630 and Cys636 surrounding the p.632_633del region cannot continue to form a disulfide bridge as the geometry between these two cysteines is no longer optimal (Fig. 2A). The CRD is flexible with respect to TMD.
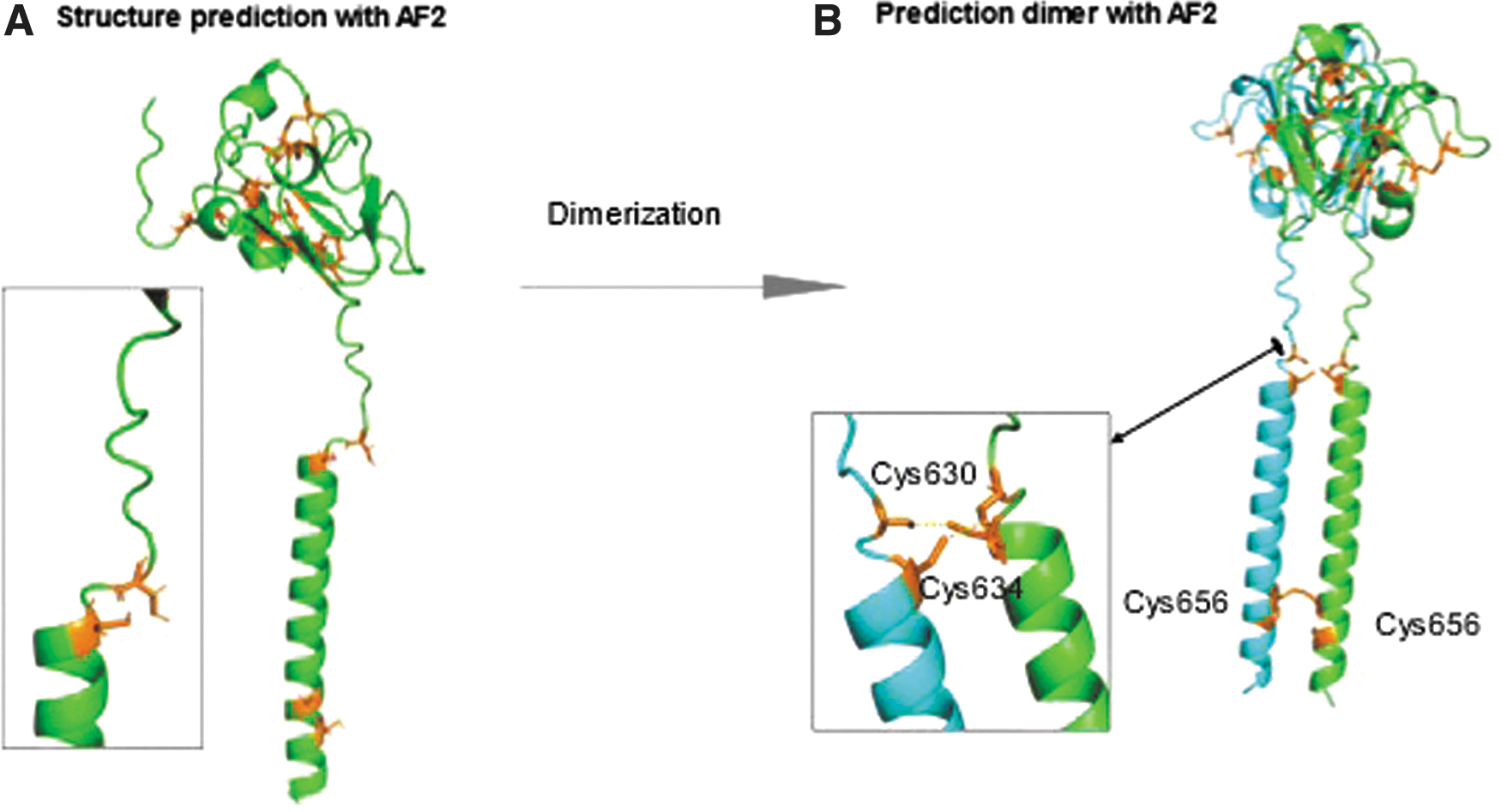
(
Modeling of the extracellular domain RET dimer
AF2 was not able to model the RET wild-type dimer. The only model provided shows low confidence where Cys630 and Cys634 are no longer bonded. As the disulfide bridge is a strong covalent bond, it was not broken during the assembly with another subunit. In contrast, model of the p.632_633del RET dimer with AF2 gave high confidence models where Cys656 from each monomer was forming a disulfide bridge. A dimerization of RET is observed (Fig. 2B).
Modeling of the intracellular domain RET dimer
AF2 provided a model of the p.632_633del RET dimer where relative arrangement of TK domains of each monomer differs from the X-ray structures of the wild-type RET (Supplementary Fig. S4). Particularly, the loops containing residues 732–736 that border the binding pocket of selpercatinib are separated from each other in p.632_633del RET dimer (Supplementary Fig. S4A) while being close in wild-type RET complexed with selpercatinib (Supplementary Fig. S4B). 25
In vitro studies
In vitro experiments confirmed a significant reduction in efficacy of selpercatinib upon cells containing the p.632_633del RET as compared with control cells with the wild-type RET. Pralsetinib was similarly affected, whereas the inhibitory action of cabozantanib and vandetanib was not significantly impacted by presence of mutated p.632_633del RET (Fig. 3).
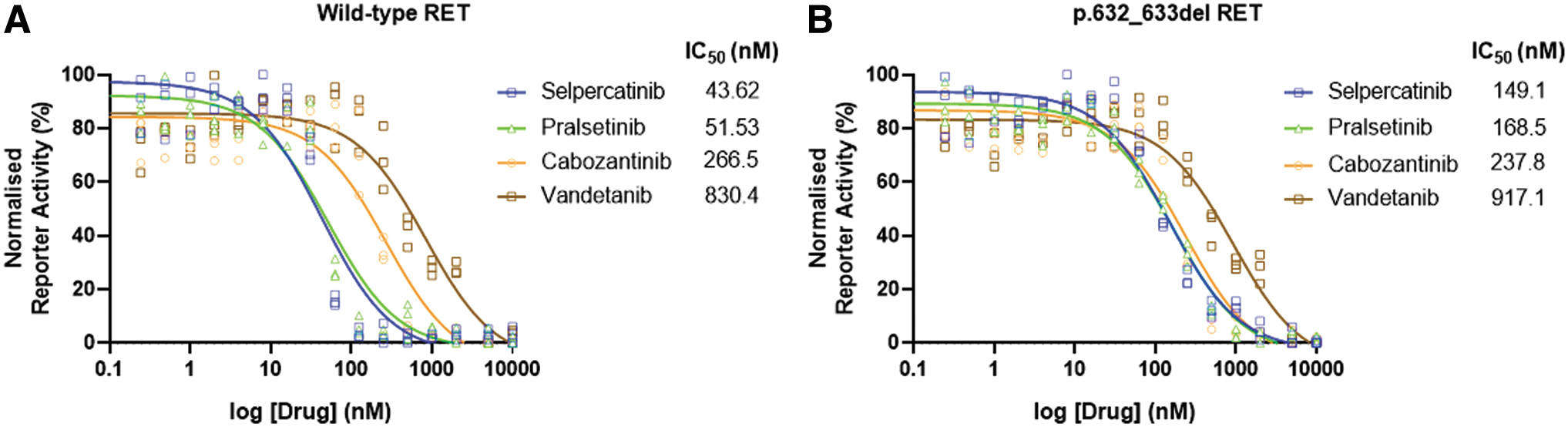
MAPK reporter responses to selective and multikinase RET inhibitors. IC50 values for selpercatinib, pralsetinib, cabozantinib, and vandetanib in HEK293 cell-based assays of (
Discussion
This is the first description of the clinical course and poor response to selpercatinib in MTC associated with p.632_633del RET. RET oncogene mutations are typically heterozygous single nucleotide variants causing autoactivation of this tyrosine kinase receptor. 26 Interstitial deletions occur in 9–17% of sporadic RET mutated MTC cases. 8,11 –15,27 The presence of deletion has been associated with aggressive disease. 15
Only a handful of case reports have described the clinical course of deletions in the CRD of the RET protein amino acid residues 630 to 634.
14
–17,28,29
Ceccherini et al described a heterozygous somatic 6bp in-frame RET deletion variant producing
Functional studies have found that deletion of the residues Glu632 to Leu633 induces a stable receptor dimer formation that results in constitutive dimerization and autophosphorylation of the receptor.
30
More recently Ma et al reported that
Our case is notable for progressive disease that occurred without increased serum calcitonin. Nonsecretory MTC occurs in only 0.83% of all cases. 31,32 Furthermore, calcitonin may be unreliable in patients on kinase inhibitors such as selpercatinib, as the drug directly inhibits calcitonin gene expression resulting in calcitonin levels disproportionately low to the effects on tumor burden. 33 We were able to identify the oncogenic RET mutation in ctDNA and demonstrate an increase in variant allele frequency with structural progression, which may be a more reliable means of monitoring disease response in this setting. 34
Selpercatinib is a highly selective RET inhibitor, which binds to the ATP binding site of the RET kinase. Acquired solvent front resistant mutations have been described at RET G810C/S and RETY806C/N, which hinder binding of selpercatinib. 25,35 Our modeling suggests that p.632_633del RET may be resistant to treatment with selpercatinib. In p.632_633del RET, the disulfide bridge between Cys630 and Cys634 seen in the wild type is no longer possible. Consequently, CRD of each RET monomer is no longer correctly orientated, increasing its proximity compared with wild type.
Indeed, the hinge bending observed between CRD and TM domain for each monomer makes bonding between TM domains impossible in the wild type. Even with giving internal flexibly in the TM domain, Cys 566 and Cys659 are not orientated toward each other, making it impossible for the formation of a salt bridge in the wild type. In p632_633del RET, an intermolecular disulfide bridge is made between the two Cys656, at the end of the TMD near the juxtamembrane segment, leading to autophosphorylation of RET, even without the interaction of the ligand GDNF-GFRα (Supplementary Fig. S5).
Selpercatinib binding is located on the TK1 domain and is surrounded by loop residues 732–736 from each monomer; the modeling suggests that the intracellular p.632_633del RET and subsequent Cys656-Cys656 disulfide bridge bring the two JM segments to proximity and impede the formation of fully functional TK1-TK2 domains. Indeed, JM is likely to stabilize an active form of RET in a manner dependent on Tyr687. 36 Proximity of JM segments to each dimer may impede Tyr687 phosphorylation and thus inhibit stable conformation of the intracellular RET that hosts selpercatinib.
In vitro studies confirmed our modeling findings, demonstrating reduced selpercatinib efficacy for p.632_633del RET compared with wild-type RET, with approximately threefold increase in IC50. In contrast, broad tyrosine kinase inhibitors, which inhibit VEGF (i.e., cabozantinib or vandetinib), were able to maintain its inhibitor effects. Recently pralsetinib, a selective RET inhibitor, was found to be efficacious in inhibiting the gain of function p.c630del RET mutation. 29 In contrast, our data showed that pralsetinib was not as efficacious when compared with more broad spectrum tyrosine kinase inhibitors in the presence of a p.632_633del RET.
Our patient, with a p.632_633del RET, had an initial partial response to selpercatinib followed by significant progression, reflecting a poor overall response to treatment. The patient's initial partial response and presence of the mutation before commencement of therapy exclude both a primary and secondary resistance mechanism, respectively, and is consistent with the recent reporting of two other patients in the LIBRETTO-001 trial who had a partial response to selpercatinib. 15 Nevertheless, our findings suggest that patients with p.632_633del RET are at risk of poorly responding to selpercatinib and those who progress on therapy may potentially benefit from an early change in management.
In conclusion, we demonstrate a single case showing MTC associated with somatic
Footnotes
Authors' Contributions
A.W. contributed to ethics submission, ctDNA sample collection, ctDNA extraction and quantification, data collection, and prepared and edited the article. K.B. made structural models and reviewed and edited article. B.W. was involved in ctDNA testing. M.G. reviewed and edited the article. C.L. carried out genetic and somatic mutation testing of RET and reviewed and edited the article. A.G. was involved in review of histology and reviewed and edited the article. B.R. took charge of patient management and reviewed and edited the article. M.B. developed assays, in vitro experiments, ctDNA analysis, and reviewed and edited the article. R.C.-B. reviewed and edited the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was involved in this study.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Table S1
Supplementary Table S2