
Abstract
Background:
Graves' orbitopathy (GO) is a disfiguring and sight-threatening autoimmune disease. Previous studies have shown the infiltration of macrophages in GO orbital connective tissues. However, the immunophenotypes of macrophages and their modulatory effects on orbital fibroblasts (OFs) have not been examined so far. In this study, we sought to determine the pathophysiology of macrophages in GO.
Methods:
In this case–control study, orbital connective tissues collected from 40 GO patients and 20 healthy controls were immunohistochemically stained for cytokines and macrophage cell surface antigens. The polarization of orbital-infiltrating macrophages was investigated by flow cytometry and immunofluorescence. Effects of interleukin (IL)-6 combined with soluble IL-6 receptor (sIL-6R) on the proliferation, differentiation, and inflammation of different OF subsets were examined by CCK-8, Western blotting, and Luminex assays, respectively. The antigen-presenting abilities of different OF subsets under IL-6/sIL-6R signaling were studied by proteomics. Finally, the differentiation of CD8+ IL-17A-producing T cells by sIL-6R was tested.
Results:
GO orbital connective tissues displayed increased IL-6, sIL-6R, STAT3, and IL-17A levels. CD86+ M1-like macrophages were predominant in active GO patients, while stable GO patients tended to have more CD163+ M2-like macrophages. The expression of IL-6 was higher in M1-like macrophages, and the expression of transforming growth factor-β was higher in M2-like macrophages both in GO orbital connective tissues in situ in vivo and in cell culture system in vitro. The IL-6/sIL-6R stimulation promoted the fibrosis of both CD34+ and CD34− OFs. Monocyte chemoattractant protein-1 expression was also induced by IL-6/sIL-6R stimulation in both OF subsets. IL-6/sIL-6R stimulation enhanced the antigen processing of CD34+ OFs through upregulating the intact major histocompatibility complex I and antigen transporters. However, the protein expressions of the thyrotropin receptor and insulin-like growth factor 1 receptor could not be directly increased by IL-6/sIL-6R stimulation in CD34+ OFs. Furthermore, sIL-6R was conducive to the differentiation of CD8+ IL-17A-producing T cells.
Conclusions:
Our study demonstrated the immunophenotypes of orbital-infiltrating macrophages that may activate OFs depending on the IL-6/sIL-6R signaling in GO. Our preclinical findings implicate, at least in part, the molecular rationale for blocking sIL-6R as a promising therapeutic agent for GO.
Introduction
Graves' orbitopathy (GO) that occurs predominantly in patients with Graves' disease (GD) 1,2 is an autoimmune condition provoked by the binding of circulating thyrotropin receptor (TSHR) autoantibodies produced by autoreactive B cells controlled and aided by autoreactive T cells to the TSHR expressed on orbital fibroblasts (OFs) as well as the interactions between various cytokines released by T cells and OFs. 3,4 The cross talk between TSHR and insulin-like growth factor 1 receptor (IGF-1R) on OFs results in overproduction of extracellular matrix (ECM) that causes the expansion of the orbital contents. 5 Orbital inflammation and tissue-remodeling are further exacerbated by pathological differentiation of OFs activated by different T cell subsets. T helper (Th) 1 cells secrete interferon (IFN)-γ and lead to early inflammatory responses in active GO, while Th2 cells secrete interleukin (IL)-4 and mediate late fibrotic responses in stable GO. 6
However, GO pathogenesis has not been fully elucidated. The recently identified CD34+ OFs originating from circulating bone marrow-derived fibrocytes seem to play a key role in GO autoimmunity. 7 Furthermore, increased proportions of Th17 cells in orbital connective tissues aggravate both GO inflammation and fibrosis. 8,9 The emerging role of macrophages has also been implicated in GO. A previous study demonstrated that GO orbital connective tissues were infiltrated by CD68+ macrophages positively correlated with monocyte chemoattractant protein (MCP)-1 expression. 10 Orbital macrophage infiltration was apparently downregulated in successfully treated stable GO patients. 11 Macrophages might activate OFs by producing platelet-derived growth factors. 12
A correlation between the expression of CD68 and CD34 indicated a potential interaction between macrophages and OFs. 13 Yet the plasticity of macrophages and how they contribute to the dysfunction of different OF subsets have not been investigated carefully. In this study, we aimed to explore the polarization status of orbital-infiltrating macrophages and their underlying regulation of both the CD34+ and CD34− OF subtypes, specifically to determine the contribution of the IL-6 and soluble IL-6 receptor (sIL-6R) pathway.
Materials and Methods
Sample collection
This was a case–control study. Fifty-three GO patients and 32 healthy controls were initially enrolled consecutively from the Department of Ophthalmology, Shanghai Ninth People's Hospital, Shanghai Jiao Tong University School of Medicine, from July 2021 to February 2022. Our study was one of the studies within the framework of the Sample Database Project of Shanghai Ninth People's Hospital (YBKB201901). Informed consent was obtained by two investigators (Y.L. and Yi Wang) from all participants for review of clinical data and collection of orbital connective tissue and blood samples as approved by the local Ethics Committee (SH9H-2019-T192-2) for the biobank. The study was independent and there was no patient or control overlap with prior studies from the biobank. Patients were eligible if they were 18 years of age or older, with active or stable GO and in need of orbital decompression surgery. Control subjects who required blepharoplasty were age- and gender- matched.
The exclusion criteria were as follows: (1) other autoimmune diseases such as systemic lupus erythematosus, IgG4-related disease, Sjogren's syndrome, rheumatoid arthritis, ankylosing spondylitis, psoriasis, or uveitis; (2) active infection; (3) chronic inflammation such as tuberculosis, (4) history of orbital trauma within 3 months; (5) current pregnancy or lactation; or (6) history of steroid or immunosuppressive agent therapy within 3 months.
All experiments were strictly compliant with the tenets of the Declaration of Helsinki. Orbital connective tissues and peripheral blood were taken from all participants during orbital decompression surgery for patients or blepharoplasty for age- and gender-matched healthy controls. The orbital connective tissues were immunohistochemically stained for cytokines and compared between GO patients and healthy controls in average optical density (AOD) measured by the Fiji/ImageJ open-source software. The polarization of orbital-infiltrating macrophages was investigated by immunohistochemistry, flow cytometry, and immunofluorescence and compared among the active GO, stable GO, and control groups. Subjects who failed complete experimental procedures due to insufficient tissue volumes were not included in the analyses.
Statistical analyses
Statistical analyses were performed by using GraphPad Prism (8.0) software. For continuous variables that conform to a normal distribution, Student's t test or one-way analysis of variance was carried out for comparison of two or more groups, respectively. Mann–Whitney U test or Kruskal–Wallis test was used to analyze the differences between two groups or among more than two groups, respectively, which was suitable when continuous variables failed normal distribution. Chi-square test was used for statistical analyses of categorical variables. Spearman rank correlation was used for the correlation analysis. Results were presented as mean ± standard deviation. p-Values are marked as follows: *<0.05; ** <0.01; *** <0.001; **** <0.0001.
A detailed experimental scheme is provided in Supplementary Materials.
Results
A total of 40 eligible patients (20 active and 20 stable GO patients) and 20 healthy controls were finally analyzed. The detailed recruitment process of participants is shown in Supplementary Figure S1. The demographic and clinical characteristics of GO patients and controls are described in Table 1. There were no significant differences in gender, age, or smoking habit among the active GO, stable GO, and control groups. There were no significant differences in thyroid function status, antithyroid therapy, or disease duration between the active GO and stable GO groups. Active GO patients had a significantly higher clinical activity score (CAS) and TSHR autoantibody level compared with stable GO patients.
Baseline Parameters of the Studied Population
Continuous variables were presented as mean ± standard deviation.
CAS, clinical activity score; GO, Graves' orbitopathy; hyper, hyperthyroidism; hypo, hypothyroidism; NA, not applicable; normal, normal thyroid function.
Macrophages infiltrated GO orbital connective tissues
Our immunohistochemistry data showed higher expressions (AOD) of IL-6 (0.36 ± 0.10 vs. 0.27 ± 0.15, p = 0.0400), IL-6R (0.30 ± 0.08 vs. 0.24 ± 0.08, p = 0.0078), STAT3 (0.13 ± 0.07 vs. 0.07 ± 0.04, p = 0.0015), and IL-17A (0.23 ± 0.10 vs. 0.16 ± 0.08, p = 0.0058) in GO orbital connective tissues compared with controls, indicating that the IL-6/STAT3 pathway may involve in GO pathogenesis (Fig. 1A). CD86+ M1-like macrophages were slightly enriched in the active GO group (27.40 ± 7.44/high power field [HPF]) compared with the stable GO group (22.15 ± 6.71/HPF; p = 0.3261), and were significantly increased compared with control group (8.75 ± 4.92/HPF; p < 0.0001).
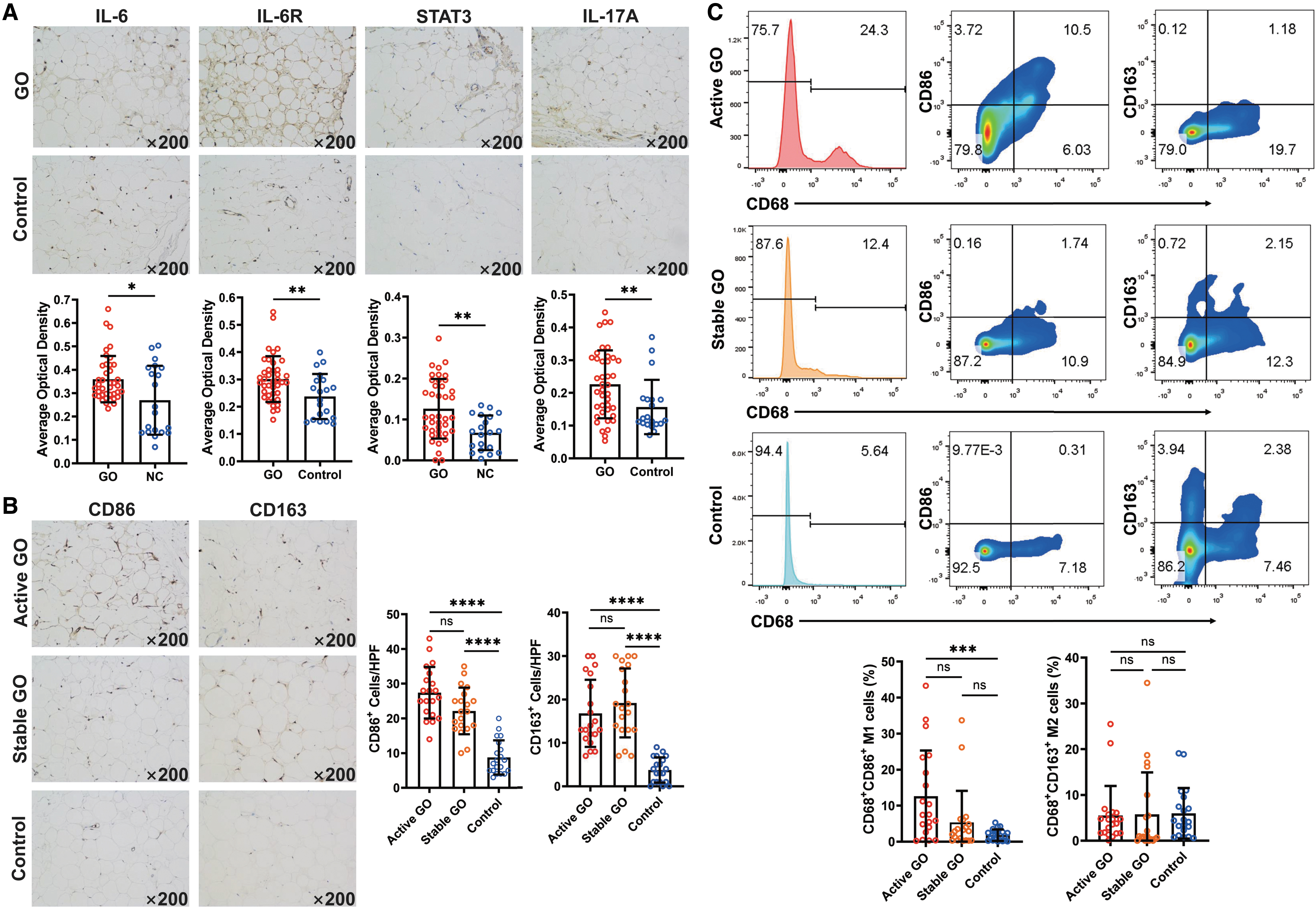
Macrophage polarization in orbital connective tissues from active and stable GO patients and healthy controls. (
In the chronic phase, with the alleviation of orbital inflammation, CD163+ M2-like macrophages tended to increase in the stable GO group, but without significant difference compared with the active GO group. Those cells were significantly recruited in both active GO (16.80 ± 7.74/HPF; p < 0.0001) and stable GO (19.20 ± 7.94/HPF; p < 0.0001) groups compared with control group (3.75 ± 2.90/HPF) (Fig. 1B). In line with in situ immunohistochemical staining, flow cytometry further confirmed that the proportion of CD68+CD86+ M1-like macrophages in the active GO group (13.26% ± 12.70%) was higher than those in the stable GO (5.34% ± 8.78%; p = 0.0344) and control (1.79% ± 1.60%; p = 0.0002) groups. However, the proportions of CD68+CD163+ M2-like macrophages displayed no significant difference among the active GO (5.49% ± 6.47%), stable GO (5.75% ± 9.18%), and control (5.98% ± 5.53%) groups (Fig. 1C).
Orbital-infiltrating M1-like macrophages secreted higher IL-6 in active GO patients
Flow cytometry also revealed that the proportion of CD68+CD86+ IL-6-producing M1-like macrophages in active GO group was the highest (19.20% ± 13.59%), compared with stable GO (6.59% ± 4.79%; p = 0.0123) and control (4.45% ± 1.73%; p = 0.0301) groups (Fig. 2A). Yet, the proportion of CD68+CD163+ TGF-β-producing M2-like macrophages was much lower in the active GO group (6.85 ± 5.00%) than in both the stable (15.67 ± 7.62%; p = 0.0119) and control (14.15 ± 5.92%; p = 0.0740) groups (Fig. 2A). In situ immunofluorescence further verified the presence of IL-6-producing M1-like macrophages in active GO and TGF-β-producing M2-like macrophages in stable GO groups, respectively (Fig. 2B). Moreover, IL-6 and TGF-β levels were detected by enzyme-linked immunosorbent assay (ELISA) in the supernatants from in vitro cultured M0, M1, or M2-like macrophages derived from human primary CD14+ monocytes (Fig. 2C).
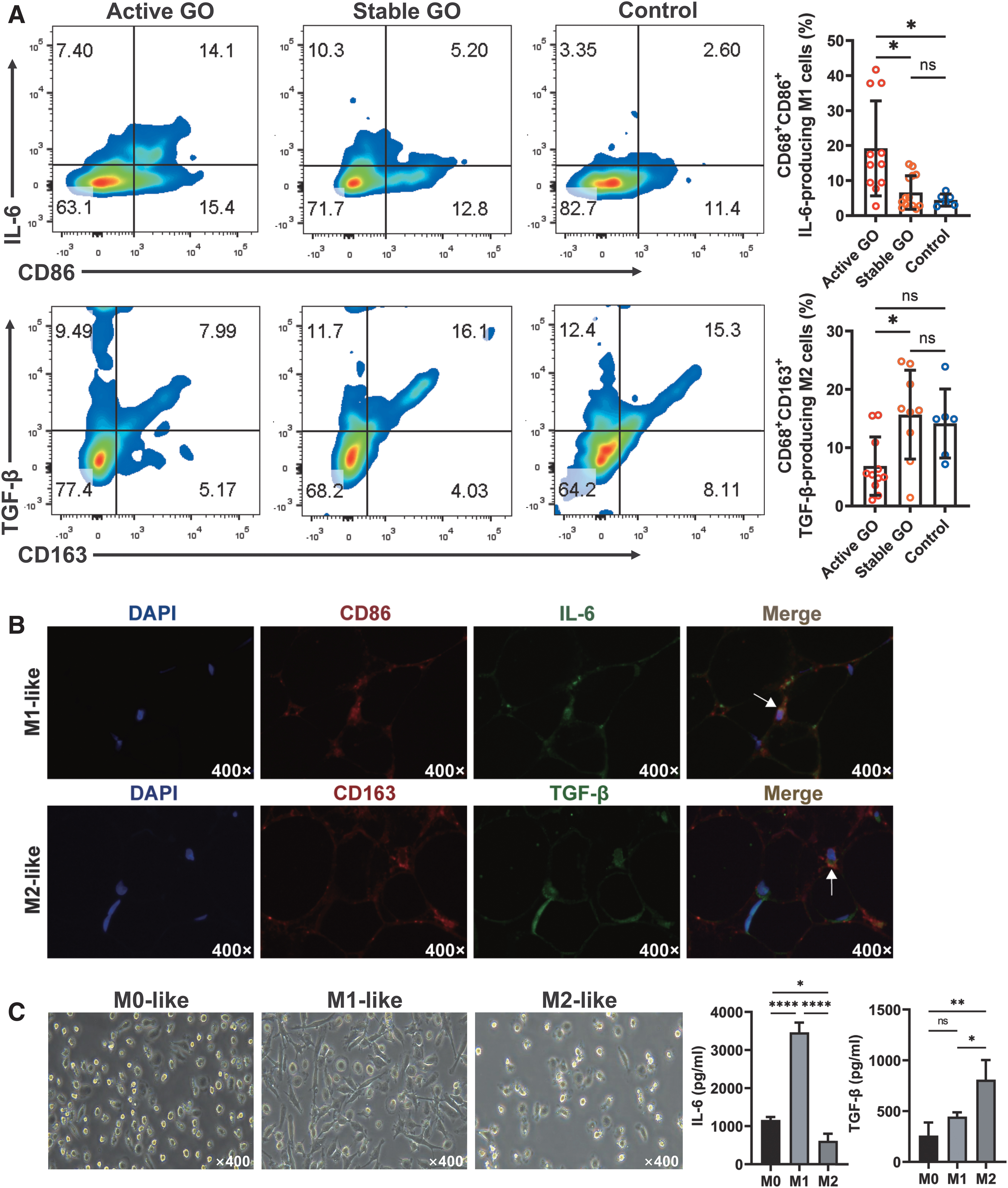
IL-6 secretion by M1-like macrophages and TGF-β secretion by M2-like macrophages in orbital connective tissues from GO patients. (
Our data demonstrated that the concentration of IL-6 (3463.0 ± 259.3 pg/mL) was the highest in M1 media, while TGF-β (808.9 ± 193.7 pg/mL) was the highest in M2 media (Fig. 2C).
sIL-6R was increased in the sera of active GO patients
We next examined serum levels of IL-6 and sIL-6R in the three groups by ELISA. Our data showed that there was no significant difference of serum IL-6 levels among active GO (15.79 ± 6.77 pg/mL), stable GO (15.13 ± 4.96 pg/mL), and control (16.13 ± 6.06 pg/mL) groups (Fig. 3A). No significant correlation between CAS and serum IL-6 level (R 2 = 0.069, p = 0.101) was observed (Fig. 3B). Intriguingly, the serum sIL-6R level in active GO group (27.62 ± 5.85 ng/mL) was significantly elevated compared with those of stable GO (21.46 ± 5.75 ng/mL, p = 0.0036) and control (19.05 ± 5.66 ng/mL, p < 0.0001) groups, but there was no significant difference between stable GO and control (p = 0.3884) groups (Fig. 3C). In addition, a positive correlation was observed between serum sIL-6R level and CAS (R 2 = 0.250, p = 0.001) (Fig. 3D).
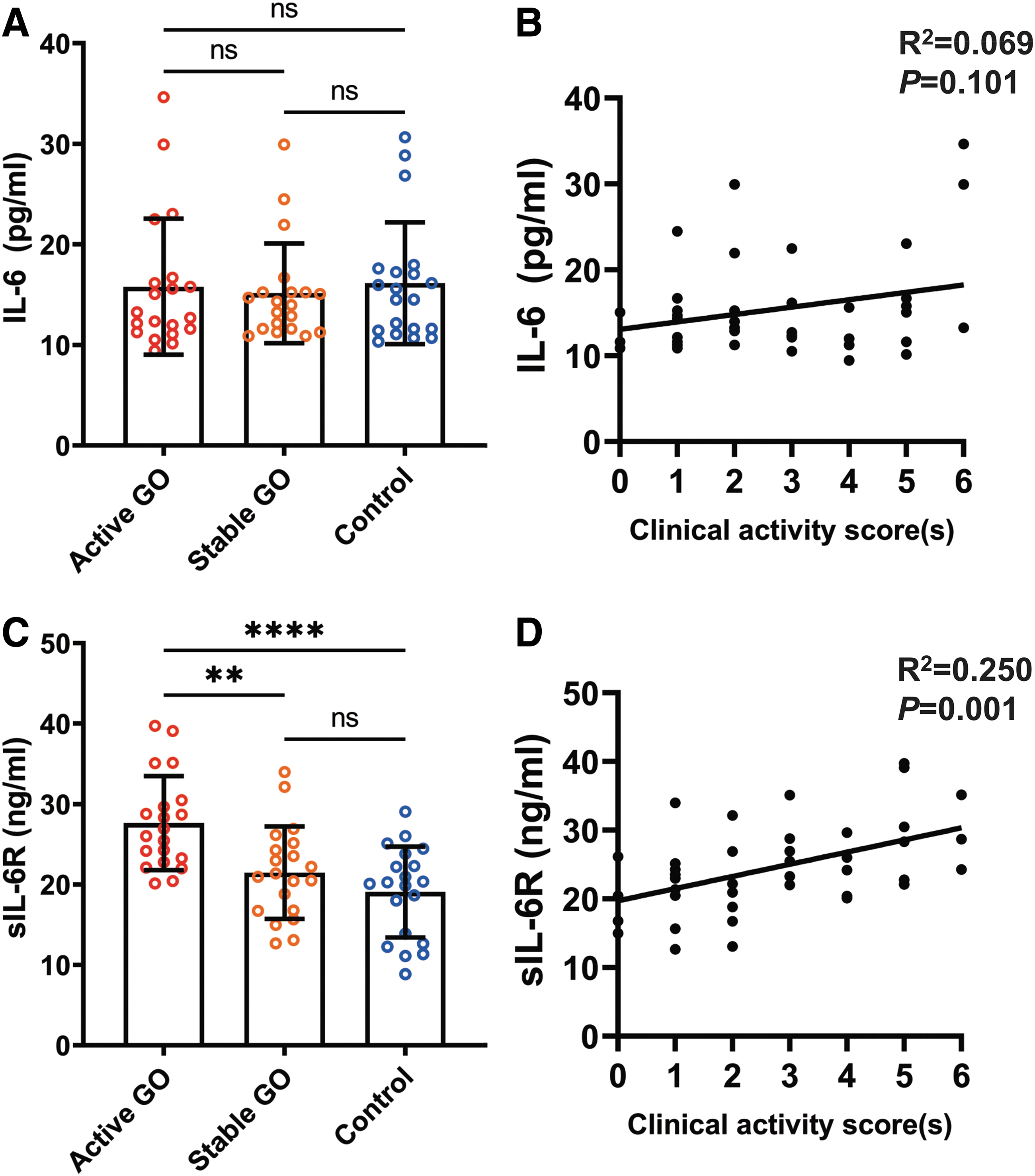
Serum levels of IL-6 and sIL-6R in GO patients and their correlations with CAS. (
IL-6 plus sIL-6R promoted the fibrosis of OFs
Neither IL-6 nor IL-6 plus sIL-6R significantly affected the proliferation of CD34+ or CD34− OFs (Fig. 4A). IL-6 plus sIL-6R enhanced the expression of ECM proteins including fibronectin, collagen I, α-SMA, and TIMP-1 in both CD34+ and CD34− OFs, which was more pronounced compared with IL-6 or sIL-6R alone (Fig. 4B). The profibrotic effect of IL-6 plus sIL-6R on OF subsets could be reversed by tocilizumab (Fig. 4B). A signal transduction examination revealed that IL-6 alone could not upregulate the phosphorylation of STAT3 in both CD34+ and CD34− OFs, whereas IL-6 plus sIL-6R synergistically activated STAT3 (Fig. 4C). Simultaneously, IL-6 plus sIL-6R upregulated the phosphorylation of ERK in both CD34+ and CD34− OFs, with a more potent effect on the CD34+ OFs (Fig. 4C). The expression of Akt phosphorylation was also upregulated in both CD34+ and CD34− OFs with IL-6 plus sIL-6R treatment (Fig. 4C).

Effects of IL-6 and sIL-6R on the proliferation and fibrosis of OFs. (
IL-6 plus sIL-6R enhanced the proinflammatory and autoimmune functions of OFs
Both CD34+ and CD34− OFs mainly expressed MCP-1, IL-8, and GRO-α at rest (Fig. 5A). When treated with IL-6 plus sIL-6R, increased levels of MCP-1 were observed in both subsets (Fig. 5A). To further unravel the regulatory effects of IL-6 plus sIL-6R on OFs, we performed a proteomic analysis by liquid chromatograph-mass spectrometry/mass spectrometry and found elevated expressions of major histocompatibility complex (MHC) I peptide including MHC I α and β chains and antigen transporter (TAP) 1 and 2 (Fig. 5B). Higher expressions of STAT3 and NF-κB were also observed in CD34+ OFs at 48 hours after treatment of IL-6 plus sIL-6R (Fig. 5B). In CD34− OFs, STAT3, NF-κB, MHC I β chain, and TAP1 were not found to be increased at long-term treatment (Fig. 5B). Yet, the expressions of CD40 and intercellular cell adhesion molecule (ICAM)-1 were upregulated in both CD34+ and CD34− OFs at 48 hours after treatment of IL-6 plus sIL-6R (Fig. 5B).
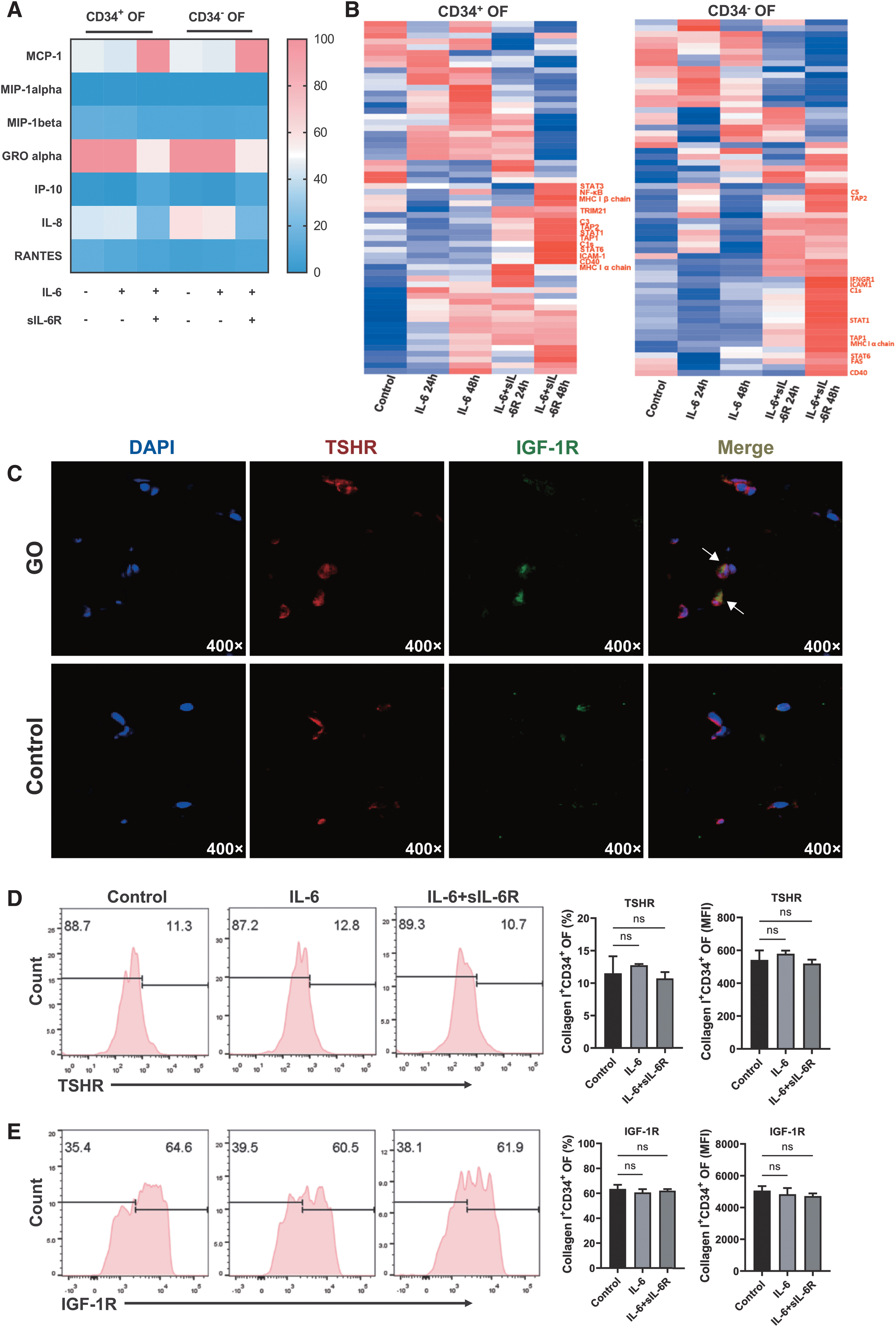
Effects of IL-6 and sIL-6R on chemokine secretion and antigen processing function of OFs. (
In addition, immunofluorescence of orbital connective tissues from GO patients showed enriched expression of THSR and IGF-1R and colocalization of these two receptors around the nuclei of orbital fat cells (Fig. 5C). Only a slight expression of THSR and IGF-1R was observed in the control group (Fig. 5C). To explore the effect of IL-6 plus sIL-6R on the expression of these two receptors in GO, we performed flow cytometry and found that neither TSHR nor IGF-1R changed significantly upon IL-6 plus sIL-6R treatment in CD34+ OFs (Fig. 5D, E).
IL-6 plus sIL-6R promoted the differentiation of naive CD8+ T cells into Tc17 cells
In the Tc17 cell differentiation system, additional supplement of sIL-6R significantly increased the frequency of IL-17A+ T cells (3.85 ± 1.31% vs. 1.13 ± 0.72, p = 0.0265), which could be attenuated by tocilizumab treatment (1.92 ± 0.57%) (Fig. 6A). Although the frequency of IFN-γ+ T cells was slightly elevated by an additional supplement of sIL-6R, there was no significant difference (2.87 ± 1.68% vs. 1.32 ± 0.89%, p = 0.31) (Fig. 6A). Neither RORγt nor RORγtTbet showed significant difference upon sIL-6R stimulation (Fig. 6B). However, sIL-6R enhanced the expression of CXCR3 (4.50 ± 1.75% vs. 0.99 ± 0.34%, p = 0.0206), not CCR6 (3.43 ± 1.00% vs. 1.33 ± 0.68%, p = 0.0633), on in vitro differentiated Tc17 cells, which could be downregulated by tocilizumab treatment (Fig. 6C).
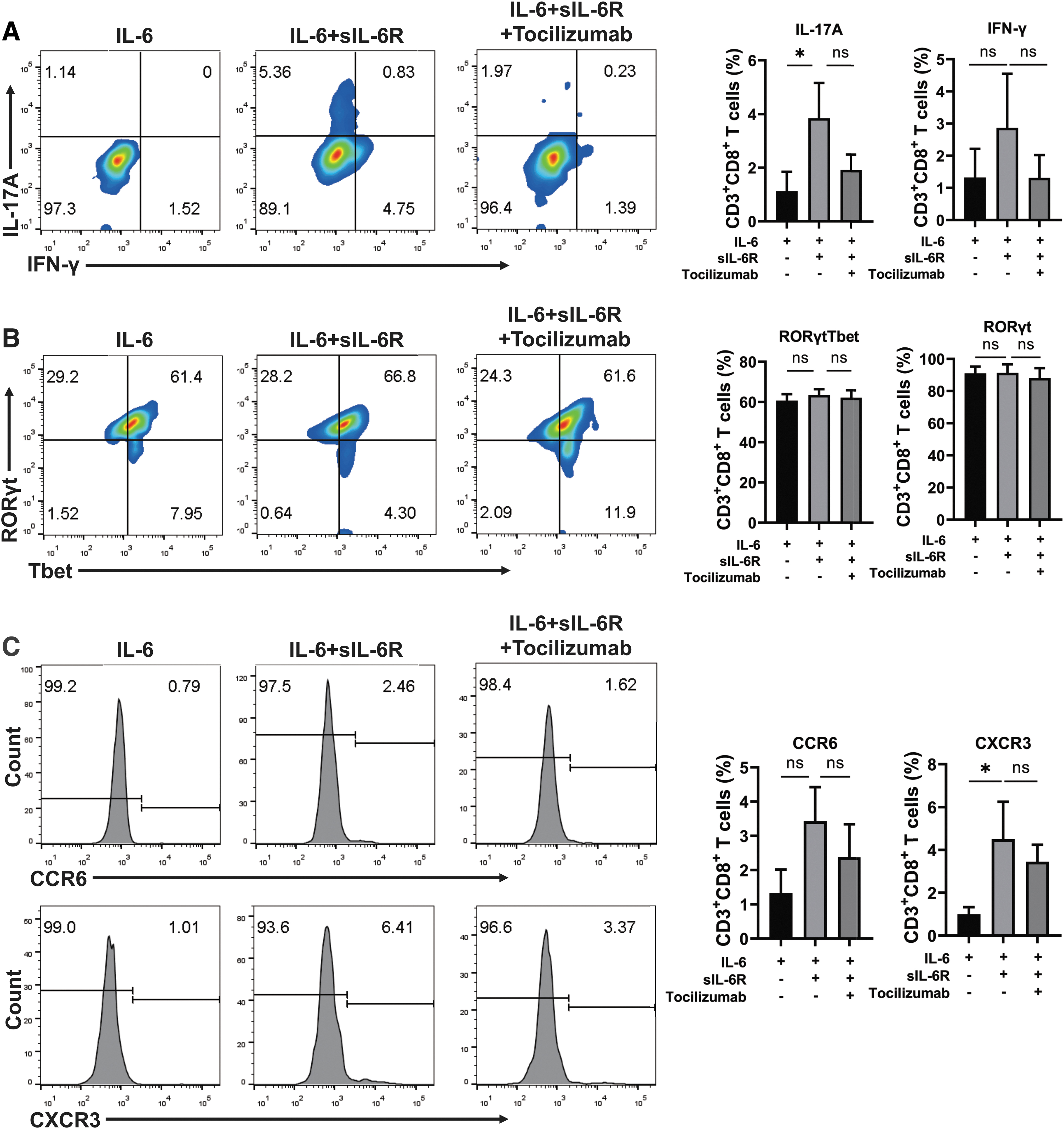
Effect of sIL-6R on the differentiation of CD8+ IL-17A-producing T cells. (
Discussion
Human autoimmune diseases are mainly driven by adaptive immunity, namely, by autoreactive B and T cell overactivity. In GO, it has been suggested that both CD4+ T cells and CD20+ B cells infiltrating orbital connective tissues were correlated with CAS. 14,15 In addition, the infiltrating CD8+ T cells were found to have a significant effect on diplopia of GO patients, while CD19+ B cells were found to be associated with exophthalmos. 15 However, early innate immunity involving macrophages is the starter of all adaptive immunity, both in normal immunity and autoimmunity. 16,17 Previous studies have reported the appearance of macrophages in GO orbital connective tissues. 10 –13 We recently revealed that CD14+CD68+ macrophages and CD141+ dendritic cells were the main cell components of antigen-presenting cells in the local immune microenvironment in GO orbits, 18 indicating that macrophages might regulate the activation of GO autoimmunity.
Macrophages process and present self-antigens to activate autoreactive T and B cells, which in turn recruit more macrophages into the sites of inflammatory orbit and amplify autoimmunity through the production of chemokines such as MCP-1. 10 On the contrary, macrophages could promote fibrosis of OFs, 19 which participates in the tissue-remodeling process mediated by cellular and humoral immunity in GO. In a recent study of GO murine models immunized by TSHR plasmid, orbital inflammation was found to start with an initial wave of macrophages followed by infiltration of CD3+ T cells and production of autoantibodies. 20 Although present in the early phase of GO, macrophage infiltration seemed persistent and did not attenuate in long duration of the disease. 10 Moreover, macrophage infiltration in orbital connective tissues was found to be strongly correlated with disease activity and severity of GO, 11,21 suggesting macrophages to be an important pathological factor of GO apart from T and B cells.
Activated macrophages have two distinct phenotypes: the classical (M1) and alternative (M2) subsets. 22 M1-like macrophages produce inflammatory cytokines such as IL-6, IL-23, and IL-1β, while M2-like macrophages act as major part in anti-inflammatory and immunomodulatory functions by producing TGF-β and IL-10. 23 In many human autoimmune diseases such as systemic lupus erythematosus, 24 inflammatory bowel disease, 25 autoimmune neuritis, 26 allergic asthma, 27 and rheumatoid arthritis, 28 the imbalance between the M1/M2 ratio is implicated in inflammatory reaction as well as the reparatory phase, which could be also detected in GO in our study. The frequency of CD68+CD86+ IL-6-producing M1-like macrophages was augmented in orbital connective tissues of active GO.
A stable inactive GO phase was characterized by the trend of a shift from those M1-like macrophages to CD68+CD163+ TGF-β-producing M2-like macrophages, similar to the results of an interesting study showing a significant infiltration of CD68+CD163+ M2-like macrophages and a depletion of CD3+ and CD20+ lymphocytes in orbital connective tissues from patients with stable GO after low-dose rituximab treatment. 29 Collectively, macrophage polarization could reflect the biphasic nature of GO to a certain extent.
IL-6 is an important cytokine in GO autoimmunity, which could be produced by OFs on various stimulations. 30 –33 We have reported that OFs secreted very low levels of IL-6 at rest, 30 which proposed a possibility of another source of IL-6 other than OFs at GO onset. Our study provided a clue to the overexpressed IL-6 level in GO orbital connective tissues, suggesting a new way of blocking IL-6 through modulating macrophage polarization. It was surprising that there were no significant differences of serum IL-6 concentrations among the active and stable GO patients and controls. One possible reason was that GO is an organ-specific autoimmune disease. The orbital microenvironment has a unique immunological constitution as we reported previously. 18 It is different from the systemic condition and might be less affected by systemic factors such as thyroid function.
This was proved by the fact that IL-6 expression in local orbital connective tissues from GO patients was higher than those from controls in our study. The cellular basis is that OFs and macrophages secrete IL-6, but it is not known whether serum IL-6 comes from the local production or other circulating cell types.
In our study, IL-6 alone had little effect on both CD34+ and CD34− OFs. In fact, it has been shown early that the combination of sIL-6R and IL-6 instead of IL-6 alone could exert obvious effect on the expansion of human primitive hematopoietic progenitor cells. 34 Likewise, we found that serum sIL-6R was the highest in active GO patients and positively correlated with CAS, implying an essential role of sIL-6R in the response of OF to IL-6. This was consistent with a previous study from Salvi et al who reported that active GO patients had a significantly higher serum sIL-6R but not IL-6 level than stable GO patients. 35
It has been recognized that for low to medium levels of serum IL-6 that are characteristic for health or acute and chronic inflammations, most IL-6 should be trapped in IL-6/sIL-6R complexes. 36 This may account for the unchanged circulating IL-6 level shown in both Salvi et al's and our studies. Thus, it seems reasonable to assume that active GO is associated with robust sIL-6R concentration, which requires a further and larger scale investigation to verify. Besides, serum sIL-6R in GD patients could be influenced by thyroid function and antithyroid therapy. 35,37 In our study, most GO patients involved had euthyroidism, which could explain the relative low correlation between the serum sIL-6R level and CAS. Similar to increased orbital IL-6 expression, we also observed highly expressed IL-6R and the corresponding STAT3 levels in GO orbital connective tissues.
Furthermore, only combined with sIL-6R could IL-6 exert potent effect on ECM synthesis of OFs in our in vitro models, which was blocked by the anti-IL-6R monoclonal antibody tocilizumab. IL-6 plus sIL-6R activated STAT3 and Akt pathways in OFs, consistent with the findings in IL-6 plus sIL-6R-induced fibrogenesis in Takayasu arteritis 38 as well as pulmonary and liver fibrosis. 39,40 Taken together, with the help of sufficient sIL-6R, IL-6 was a profibrotic factor stimulating myofibroblast differentiation of OFs, leading to GO tissue remodeling.
There were no visible differences in IL-6-triggered fibrosis or MCP-1, CD40, and ICAM-1 expression in both CD34+ and CD34− OFs in our study. However, those two OF subsets have distinct faces in GO pathogenesis under inflammatory conditions. 41 Notably, we found that the CD34+ OFs reacted more strongly than the CD34− OFs with regard to antigen processing. IL-6 plus sIL-6R could induce the integral MHC I heterodimer and the TAP1 and TAP2 42 in CD34+ OFs, some of which were not upregulated in CD34− OFs. Increased MHC I and TAP molecules on CD34+ OFs not only indicated the pathogenic nature of those cells inherited from their parental fibrocytes, but also were crucial for the initiation and regulation of adaptive immune responses. 43 Intriguingly, the protein levels of TSHR and IGF-1R were not affected by IL-6 signaling in CD34+ OFs, although IL-6 could increase TSHR messenger RNA (mRNA) expression. 44
Owing to the fact that GO orbital inflammation could be associated with early infiltration of macrophage, 20 it can be safely speculated that the presence of elevated IL-6 levels by macrophages could be necessary for GO autoimmunity through stimulating TSHR expression within the orbit. Hence, the unchanged TSHR protein levels by IL-6 treatment, here reported, might be due to the requirement of other signal pathways apart from IL-6 to enhance the translational expression of these autoantigens, which are worthy of further study. Altogether, IL-6 signaling strengthened the antigen-presenting function of CD34+ OFs through coordination of MHC I macromolecular assembly, but not upregulation of GO autoantigens.
Early studies delineated a CD8+ T cell-dependent GO orbital inflammation. 45,46 This was confirmed by the evidence of the influx of CD8+ T cells into the GO murine orbital region following macrophage infiltration. 20 We have displayed increased CD8+ IL-17A-producing T cells, namely Tc17 cells, in GO pathogenesis, 30 which have also been documented in other human autoimmune diseases. 47 We found that sIL-6R assisted IL-6 in the differentiation of naive CD8+ T cells into Tc17 cells. It can be safely assumed that orbital-infiltrating CD8+ T cells are likely to differentiate into Tc17 cells in the presence of IL-6-producing M1-like macrophages and that those Tc17 cells could be further activated through increased MHC I complex on the CD34+ OFs.
There are several limitations in our study. First, although the participants were consecutively enrolled, the insufficient available tissue volumes of some cases prevented us from including all the original surgical samples for analyses. Hence, the potential for selection bias still exists. Second, according to the procedures of our biological experiments, we evaluated the immunological features of GO patients at a single time point. Our results may not be suitable for prediction of the temporal fluctuation in the disease duration of GO. The available GO murine models could be helpful for the immunological research across time. Third, our study was exploratory and we did not have a clear definition of the primary outcome. Further confirmatory researches are required to validate our current results.
Conclusions
In conclusion, we unraveled that M1-like macrophages regulated the fibrosis, chemotaxis, and antigen presentation of OFs and played a potential supporting role in the differentiation of CD8+ Tc17 cells in GO, which is displayed as a schematic diagram in Supplementary Figure S2. Although IL-6 has been implicated in GO pathogenesis, our current study reported the priority of sIL-6R in IL-6 signaling of OFs. Our results shed new light on the immunopathological mechanisms of IL-6 in GO autoimmunity and further strengthen the therapeutic rationale for the efficacy of sIL-6R blockade with tocilizumab.
Footnotes
Authors' Contributions
Working design: S.F. and H.Z. Writing: Yi Lu and H.Z. Review and editing: B.L. and S.F. Investigation: Yi Lu, Yi Wang, Yang Wang, Y.H., and Y.W. Sample collection: J.S., Yinwei Li, X.L., and S.Z. Data curation and analysis: J.S., Y.L., Yi Wang, and Yang Wang.
Author Disclosure Statement
The authors have nothing to disclose.
Funding Information
This work was funded by the National Natural Science Foundation of China (81930024, 82071003, 82000879, 82271072 and 82271122), the National Key R&D Program of China (2018YFC1106100 and 2018YFC1106101), the Research Grant of the Shanghai Science and Technology Committee (20DZ2270800, 17DZ2260100, 19410761100, and 19DZ2331400), Shanghai Key Clinical Specialty, Shanghai Eye Disease Research Center (2022ZZ01003), the Innovative research team of high-level local universities in Shanghai, the Clinical Research Plan of SHDC (SHDC2020CR3051B), the Project of Medical Robots (IMR-NPH202002) from the Clinical Joint Research Center of the Institute of Medical Robots, the Shanghai Municipal Education Commission—Gaofeng Clinical Medicine Grant Support (20152228), the Shanghai JiaoTong University Translational Medicine Crossed Research Grant (ZH2018ZDA12, ZH2018QNA07), the Sample Database Project of Shanghai Ninth People's Hospital (YBKB201901), and the Joint Innovation Team for Young Physicians of Shanghai Ninth People's Hospital (QC202002).
Supplementary Material
Supplementary Materials
Supplementary Figure S1
Supplementary Figure S2