
Abstract
Many members of the American Thyroid Association played prominent roles in discovering the various aspects of the hypothalamic–pituitary–thyroid axis. This axis is fundamental for maintaining the normal serum levels of circulating thyroid hormones (THs) and thus the euthyroid state. The pituitary glycoprotein hormone, thyrotropin (TSH), controls the activity of the thyroid gland. Thyrotropin-releasing hormone and the negative feedback mechanism of circulating TH regulate the synthesis and the secretion of TSH. The dynamic interplay of these two dominant mechanisms has essential effects on TSH release. Therefore, the finding of abnormal serum levels of TSH often indicates the presence of a disorder of thyroid gland function. A summary of key historical discoveries in the understanding of the hypothalamic–pituitary axis is presented.
Introduction
The normal secretion of thyroid hormones (THs) is essential for the regulation of their physiological effects on brain development and function, growth, basal metabolic rate, thermogenesis, energy expenditure, muscular contraction, heart rate stimulation and the expression, and activities of numerous proteins involved in lipid and carbohydrate metabolism. The hypothalamic–pituitary–thyroid (HPT) axis synchronizes the secretion of normal levels of TH (thyroxine [T4] and triiodothyronine [T3]) and there is a negative feedback mechanism between circulating TH, pituitary thyrotropin (TSH), and hypothalamic thyrotropin-releasing hormone (TRH) (Fig. 1).
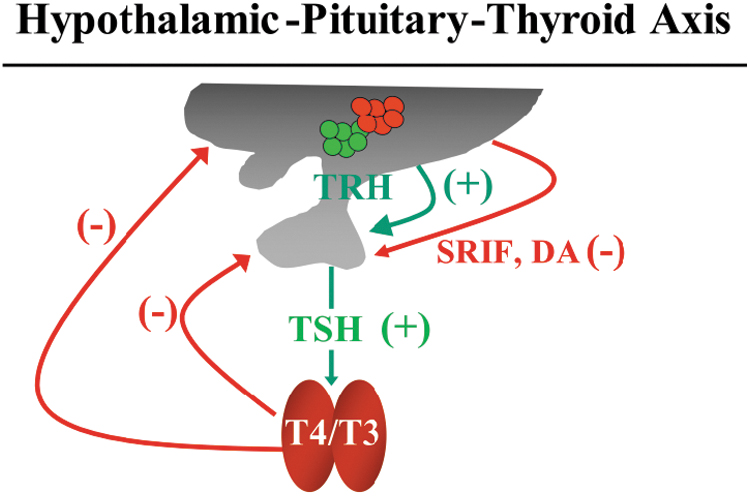
The HPT axis. TRH stimulates (+) pituitary thyrotropes to secrete TSH, which, in turn, stimulate the thyroid gland to produce THs (T4 and T3). THs control the secretion of TRH and TSH through a negative feedback mechanism (−). TSH secretion from the pituitary is inhibited (−) by the hypothalamic hormone somatostatin (SRIF) and dopamine (DA). HPT, hypothalamic–pituitary–thyroid; T4, thyroxine; T3, triiodothyronine; TH, thyroid hormone; TRH, thyrotropin-releasing hormone; TSH, thyrotropin.
The association between the pituitary gland and endocrine disease was clearly recognized in humans by Harvey Cushing who, performing complete hypophysectomy, reported a characteristic series of symptoms including bradycardia, ataxia, hypothermia, loss of consciousness, hypogonadism, and reduced growth rate. Studies in animal models showed that lesions of the pituitary and/or the hypothalamus resulted in hypothyroidism. 1 In contrast, injection of pituitary and/or hypothalamic extracts resulted in increased circulating TH. 2 In humans, gene mutations or polymorphisms, as well as pituitary or hypothalamic lesions of one or more components of the HPT axis, may cause thyroid disorders.
The concept of the HPT axis was established in the middle of the 20th century. 3 –6 The discovery of the various components of the HPT axis took place in an inverted order: first TH T4, 7,8 second the TSH, 9,10 and lastly, TRH. 11,12
In all these steps, the thyroidologist members of the American Thyroid Association (ATA) played a crucial role in the discovery of the various hormones, their chemistry, their action, and the mechanisms involved in the negative feedback regulation.
Thyrotropin-Releasing Hormone
In 1948, Geoffrey W. Harris conducted a series of experiments and concluded: “the hypothalamus served as the key link between the endocrine and nervous systems in reacting to the surrounding environment.” 1 In 1951, Monte Greer examined the inhibition of thyroxine synthesis and TSH overproduction in response to the antithyroid drug thiouracil and clearly showed that a hypothalamic lesion prevented these responses. 3 In 1955, Saffran et al suggested that an unknown substance from the hypothalamus was able to regulate the secretion of the pituitary hormones and they coined the term “releasing factors.” 13
TRH is synthetized and secreted from neuron belonging to the hypothalamic paraventricular nucleus (PVN), which projects axons to the median eminence (ME). In the ME, TRH is processed from nerve terminals located close to the portal vessels and to the tanycytes. 14
The chemical structure of TRH was discovered in 1970 in the laboratories of two Nobel Prize winners, Andrew V. Schally and Roger Guillemin (Fig. 2). TRH is a tripeptide [

Nobel Prize winners Roger Guillemin and Andrew V. Schally who discovered the chemical structure of TRH in 1970.
A TRH-degrading ectoenzyme ([TRH-DE] pyroglutamyl peptidase II) present in the membrane of the tanycytes can inactivate TRH before it enters the portal vessels. 17 Therefore, TRH-DE modulates the amount of TRH that enters in the pituitary gland. A fast degradation of circulating TRH, which has a half-life of about five minutes, is catalyzed by various enzymes. These include pyroglutamyl-aminopeptidase 1 and 2, thyroliberinase, and proline endopeptidase, which are partially controlled by TH, and have a direct action on the expression of the TRH gene. 18 John P. Wilber and Bruce D. Weintraub discovered that the gene encoding TRH was located on chromosome 3. 19
The control of TRH secretion is mainly due to the feedback mechanism of circulating levels of T4 and T320 and the hypothalamic peptide somatostatin, which inhibits TRH secretion. 21 TRH stimulates both in vivo and in vitro the secretion of TSH and controls the correct glycosylation of TSH, which is fundamental for the normal biological activity of TSH. 22 Moreover, TRH is effective in stimulating the secretion of prolactin from pituitary lactotrope cells.
The mentioned actions are due to the binding of TRH to its type 1 receptor (TRH-R1) that belong to the family of G-protein coupled receptors (GPCR), which activate the pathway of Ca2+-dependent protein kinases. 23,24 Biallelic inactivating mutations in the 5′-part of the TRH receptor gene are responsible for congenital central hypothyroidism. 25
Finally, the clinical use of synthetic TRH represented a novel and useful test for the investigation of thyroid diseases. Most of the first data were published by American thyroidologists, 26,27 who demonstrated that the TRH-stimulation test is of special clinical value in discriminating between primary and central hypothyroidism and in assessing the functional pituitary TSH reserve in the presence of pituitary and hypothalamic lesions. Furthermore, the findings in patients with hyperthyroidism suggested a possible clinical utility of the TRH test in the differentiation of compensated and decompensated autonomous adenomas, particularly for borderline cases.
Furthermore, in patients scintigraphically suspected to have autonomous thyroid nodules, a normal response to the TRH test helped to exclude this diagnosis and avoid further diagnostic procedures, such as the T3 suppression test. Nonetheless, the TRH test is currently obsolete due to the ultrasensitive immunometric measurement of TSH and it is only used in the research setting for the phenotyping of patients with unexplained alterations in the HPT axis.
Thyrotropin
In 1916, two American scientists, Smith and Allen, identified a thyrotropic substance in the anterior pituitary of tadpoles. 9,10,28 A few years later, Leo Loeb and Max Aron independently showed that injection of a pituitary extract induced thyroid hyperplasia in guinea pigs. 29 Thereafter, successful research was conducted to separate and isolate the various hormones of the anterior pituitary. The thyrotropic substance was purified and characterized by Anderson and Collip in 1935. 30
In 1973, TSH was found to be a glycoprotein and was assigned a molecular weight between 26 and 30 kDa by Pierce et al. 31 A few years later, Pierce and Cornell showed that TSH is a heterodimeric glycoprotein hormone constituted by two subunits, α and β. 32 The α-subunit is common to the other glycoprotein hormones from the pituitary (luteinizing hormone [LH] and follicle stimulating hormone [FSH]) and from the placenta (chorionic gonadotropin), while the β-subunit confers functional and immunological specificity to the TSH molecule.
Recombinant DNA technology was then used to understand the tissue-specific developmentally regulated expression of the four different glycoprotein hormones: CG, LH, FSH, and TSH. The isolation of a full-length cDNA encoding the common α-subunit demonstrates that there is a single human gene for this protein, expressed in the pituitary and placenta, for the production of the various glycoprotein hormones. From this, it is concluded that the control for the expression of glycoprotein hormones probably resides in the β-subunit genes.
In the following years, many laboratories in the United States were able to clone the cDNA encoding TSH subunits of various species and to characterize their structure and expression. 33 –35 Moreover, it was demonstrated that the hypothalamic hormones, dopamine and TRH, can act directly on pituitary cells to modulate the transcription rate of the TSH subunit genes 36 and that the carbohydrate moiety of TSH is essential for full bioactivity of the hormone. 37 Finally, biallelic mutations of the TSH β-subunit gene can lead to central hypothyroidism characterized by a hyperplastic pituitary gland, high serum glycoprotein hormone α-subunit, and variable circulating TSH levels. 38
TSH secretion is pulsatile 39 and shows a circadian rhythm both in rat and in humans. 40 Interestingly, Ridgway and collaborators demonstrated that TSH pulsatility is significantly synchronized with prolactin (PRL) pulsatility 41 (Fig. 3). Hershman and colleagues were able to show that, during the afternoon, TSH concentrations are at the lowest levels and reach a zenith between 10 PM and 2 AM. They also documented that circadian variations are under a dopaminergic control. 42 Moreover, the same authors showed a sleep-related inhibition of TSH release. In fact, after the onset of sleep, TSH levels decrease, which is prevented by keeping the subject awake. 43,44 Finally, circannual variation in TSH secretion has been recently documented. 45

E. Chester Ridgway (1942–2014) who left us too soon.
TRH stimulates the secretion of TSH, while somatostatin and dopamine have inhibitory effects on TSH secretion. 21,42 TSH response to TRH is partially inhibited by corticosteroids, whereas estrogens strengthen the response of TSH to TRH. 46,47 Nevertheless, the most important mechanism for the regulation of TSH secretion is the negative feedback by TH, in particular T3, mediated by the β 2 isoform of TH receptors. 48 TH needs a series of transporters to enter in and to exit from the various cells of human body. Among them, two are the most important transporters: the membrane monocarboxylate transporters and organic anionic transport proteins. 49
TH tissue concentrations are regulated by the activity of 2 deiodinases (Dio2 and 3) in a tissue-specific manner. 50 In the hypothalamus, pituitary, and target tissues such as skeletal muscle, white and brown adipose tissue, Dio2 is responsible for T4 conversion to T3, while Dio3 converts T4 to reverse T3, and T3 to 3,5-diiodothyronine. In the thyrotropes and in the paraventricular hypothalamic nuclei, Silva and Larsen showed that T3 generated from T4 inhibits the transcription of the TSH β-subunit and the TRH gene. 51
The physiological TSH actions occur by binding to its specific cognate GPCR, the TSH receptor, localized on the basal membrane of thyroid cells. This binding results in the activation of cyclic adenosine monophosphate (cAMP)-dependent mechanisms throughout coupling to the α-subunit of the stimulatory G protein, which triggers the synthesis and secretion of TH. Nonetheless, other intracellular effectors, such as calcium and diacylglycerol, may promote some of the biological effects of TSH.
Loss-of-function mutations in the gene encoding the TSH receptor produce the so-called resistance to TSH action, which is characterized by elevated levels of biologically active serum TSH, absence of goiter, and mild-to-severe hypothyroidism. 52 In contrast, monoallelic gain-of-function somatic mutations of the TSH receptor are a major cause of toxic thyroid adenomas, whereas patients with sporadic or familial nonautoimmune hyperthyroidism have de novo or inherited monoallelic germline mutations of the TSH receptor. 53
It is noteworthy that TSH stimulates every step involved in TH synthesis. In fact, TSH controls the synthesis of the Na+/I symporter (NIS), which is fundamental for the iodide uptake. The stimulation of colloid endocytosis and thyroglobulin proteolysis occurs even earlier than the induction of NIS synthesis. 54 Moreover, TSH stimulates H2O2 production, iodide incorporation into tyrosine residues of thyroglobulin, and its exocytosis in the follicular lumen. Finally, chronic stimulation of the thyroid by TSH increases the transcriptional and translational activities of thyroid cells inducing hyperplasia and goiter.
To attain normal bioactivity, TSH needs proper glycosylation, a process that requires the interaction of TRH with its receptor on thyrotropes. 38 In patients with central hypothyroidism, normal or even slightly elevated serum levels of TSH may be observed, despite below normal serum levels of free T4. Studies performed in Weintraub's laboratory showed that partially purified TSH from patients with central hypothyroidism has reduced biological activity. Moreover, chronic administration of oral TRH to these patients normalized the glycosylation process, enhancing both its TSH receptor binding affinity and its capacity to activate adenyl cyclase. 55
In contrast, enhanced TSH bioactivity is invariably found in sera from patients with TH resistance. Moreover, variations of TSH bioactivity (mostly related to different TSH glycosylation) have been observed in normal subjects during the nocturnal TSH surge, in normal fetuses during the last trimester of pregnancy, in primary hypothyroidism, in patients with TSH-secreting pituitary adenomas, and in nonthyroidal illnesses. 56
Studies on TSH molecular structure and biological activity have led to the synthesis of small molecules with either antagonist or superagonist activity on the TSH receptor. 57 As reported from Gershengorn's laboratory, a nonpeptidic antagonist, therefore, devoid of intrinsic immunogenicity, would be particularly useful in the treatment of different forms of hyperthyroidism, such as Graves' disease, Graves' orbitopathy, TSH-secreting pituitary adenomas, and activating mutations of the TSH receptor. As far as the application of a super agonist is concerned, it may be useful in patients with well-differentiated thyroid cancer who may undergo radioiodine ablation of residual cancer or thyroid remnant, thus preventing prolonged periods of symptomatic hypothyroidism due to TH withdrawal.
Nonetheless, it is worth noting that today in these patients, TSH stimulation is elicited by the administration of recombinant human TSH, a current standard of care that was initially investigated more than 30 years ago.
Wondisford et al showed the significant role of the pituitary-specific transcription factor Pit-1 in the regulation of human TSH β-subunit expression. 58 Indeed, expression of an inactive mutant of Pit-1 in a clonal rat pituitary-cell line (GH3) decreases TRH stimulation of TSH, whereas transfection of Pit-1 in cell lines lacking this factor restores cAMP induction of the TSH β gene. Taken together these results strongly support a significant role of Pit-1 in the regulation of TSH gene expression.
In fact, mutations of PIT-1 in humans cause clinical syndromes of combined pituitary hormone deficiency, where TSH, GH, and PRL secretions are compromised due to the loss of the pituitary cells that produce these hormones. The significant role of Pit-1 in the control of TSH synthesis and secretion has been demonstrated by the “anti-Pit-1 antibody syndrome,” in which circulating Pit-1 antibodies are associated with combined deficiencies of GH, prolactin, and TSH. 59
Finally, the use of ultrasensitive immunometric assays for TSH measurement makes this key diagnostic test for thyroid dysfunction more precise. Nonetheless, even in the presence of a normal function of the HPT axis, artifacts in TSH measurement methods may be present and should be considered to prevent an incorrect diagnosis. Artificially elevated levels of TSH may result from the presence of endogenous antimouse γ globulin antibodies, as well as presence of large molecular-sized TSH that is mostly a complex of TSH and IgG (“macro TSH”).
Moreover, many hormone immunoassays use the biotin streptavidin interaction to immobilize immune complexes. The intake of high dose biotin can interfere with immunoassays using the biotin streptavidin interaction, thus generating falsely low or falsely high measurements of hormones, depending on the type of immunoassay used.
The presence of assay interference should be investigated especially in patients with normal levels of circulating free THs, lack of signs and symptoms of hypothyroidism, absence of antithyroid autoantibodies, and/or a normal thyroid ultrasound examination.
The HPT Axis in Pathological Conditions
Serum levels, pulse amplitude, and nocturnal peak of TSH are blunted in many nonthyroidal conditions, including starvation, moderate and severe illnesses, neuropsychiatric disorders, effects of drugs, and many others.
In fasted obese men, Hershman and coworkers showed that serum TSH, T4, and T3 are lower than in normal subjects and a blunted response of TSH to TRH is present. 60 These findings suggest that the thyrotropes remain responsive during short-term fasting and that the decrease in TSH is due to decreased hypothalamic TRH synthesis and release. 61 It is interesting to note that serum leptin concentrations are low in fasted persons. Since leptin has a direct action to modulate HPT axis by regulating TRH gene expression, the fall in circulating leptin levels probably resets the set point for TH negative feedback on TRH biosynthesis, thereby allowing adaptation to starvation. 62
The variations in circulating serum TSH that are present during fasting are much more critical during severe illness. Serum TSH is usually reduced, as are serum THs, particularly circulating T3, although in rare cases elevated serum TSH levels may be found. 63 Decrease of nocturnal TSH secretion and pulse amplitude, as well as impaired TSH response to TRH, may explain these results. Indeed, recent studies showed low levels of TRH mRNA in the PVN of patients who died of nonthyroidal disease. 64
Moreover, changes in neuroendocrine pathways have been suggested, but the activation of proinflammatory cytokines may have a critical role in the suppression of TSH secretion in nonthyroidal illness of acutely ill patients. Cytokines, such as interleukin (IL)-1β, tumor necrosis factor-α, and IL-6, are elevated in serum of patients with nonthyroidal illnesses, and they may exert a marked inhibitory activity on TRH–TSH synthesis and secretion. However, the patients do not have clinical signs and symptoms of hypothyroidism. Interestingly, during recovery from illness, an increase in serum TSH may be observed, even if circulating THs are still reduced.
Abnormal sialylation of TSH molecules may reduce the hormone bioactivity, thus explaining this apparent thyroid cell resistance to TSH. Lastly, the HPT axis almost always returns to normal after complete recovery, indicating the transient nature of these changes.
Similar alterations of the HPT axis found in nonthyroidal illness syndrome can be present in some neuropsychiatric disorders. In patients with anorexia nervosa or depressive illness, both serum TSH and its response to TRH may be blunted. The etiology of these changes is not known, though it has been speculated that they are a consequence of abnormal TRH secretion. Moreover, the same situation may be found in patients with chronic alcoholism and in those with opioid addiction.
In conclusion, many members of the ATA played important roles in discovering the various aspects of the HPT axis. These discoveries have formed the foundation of our understanding of the HPT axis as well as the pathogenesis of thyroid diseases. Furthermore, these discoveries have informed current strategies for diagnosis and treatment of thyroid diseases and future research directions.
Footnotes
Authors' Contributions
J.M.H. and P.B.-P. conceived, wrote, and edited this article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.