
Abstract
Background:
Hashimoto's thyroiditis is a common autoimmune thyroid disorder characterized by thyroid lymphocytic infiltrates and autoreactive antibodies against thyroglobulin (TgAbs) and thyroperoxidase. Final evolution of the disease can lead to hypothyroidism with destruction of the thyroid architecture. Interleukin-4 (IL-4) is involved in the humoral immune response and B cell activation required in autoimmune thyroiditis (AT) progression. We used our mouse model overexpressing IL-4 by thyrocytes (Thyr-IL4) to study the impact of a local IL-4 expression in AT using transgenic nonobese diabetic (NOD.H2h4) derived animals treated with iodide-supplemented water to increase the incidence of spontaneous AT (SAT).
Methods:
Thyr-IL4 NOD.H2h4 and nonpathogenic C57BL/6 animals aged 8 weeks were exposed to 0.05% sodium iodide (NaI) in their drinking water for 8 and 16 weeks. Circulating TgAbs and expression of intrathyroidal cytokines were quantified. Thyroid inflammation was assessed by classical histological analyses, including identification of some immune cell populations. The most sensitive parameter to evaluate the thyroid function, serum thyrotropin (TSH), was also measured at the end of the treatment.
Results:
Relative to wild-type (WT) animals, Thyr-IL4 NOD.H2h4 mice developed severe accelerated SAT with elevated serum TgAbs and numerous thyroid infiltrates mainly composed of CD4+/CD8+ T cells, B lymphocytes, and monocytes/macrophages. Thyroid expression of T helper (Th) Th1/Th2 cytokines was also enhanced, as well as IL-17. In contrast, excessive iodide supply did not induce TgAbs in WT and Thyr-IL4 SAT-resistant C57BL/6 animals. However, moderate leukocyte infiltrations in transgenic thyroids were evident compared to WT, but associated with a limited number of T and B cells and a different cytokine profile from Thyr-IL4 NOD.H2h4 mice. Finally, and despite their diverse immune responses, both transgenic strains presented marked thyroid enlargement and elevated serum TSH at the end of the treatment in contrast to their WT littermates.
Conclusions:
These findings demonstrated that ectopic expression of IL-4 from thyrocytes enhanced the severity of accelerated SAT in disease-prone Thyr-IL4 NOD.H2h4 animals and promoted thyroid leukocyte infiltration in SAT-resistant transgenic C57BL/6 mice. Moreover, impaired thyroid function emerged in both transgenic strains during the progression of the disease.
Introduction
Iodine is essential for the synthesis of triiodothyronine/thyroxine (T3/T4) thyroid hormones (TH) and is actively transported by thyroid follicular cells (TFCs) through the basal Na+/I− symporter NIS. 1 In susceptible individuals, iodine overload can cause autoimmune thyroiditis (AT). 2 Increased incidence of AT was clearly reported in multiple countries with iodized salt supplementation programs to correct iodine deficiency. 3 Hashimoto's thyroiditis is the most frequent organ-specific autoimmune disease affecting mostly the female population. 4 This autoimmune disorder is characterized by thyroid lymphocytic infiltrates and circulating autoantibodies against the main thyroid antigens thyroperoxydase (TPO) and thyroglobulin (TG). 5
Different mouse models have been developed to study the etiology of this human disease (reviewed in Quaratino 6 ). Experimental AT (EAT) can be induced in C57BL/6 mice after immunization with recombinant mouse Tpo. 7 Despite a clear destruction of TFCs in immunized mice, serum T4 levels remain normal. In contrast to C57BL/6, nonobese diabetic (NOD.H2h4) mice develop spontaneous AT (SAT), which is accelerated after iodized water supplementation. Excessive iodination of Tg facilitates the generation of new pathogenic epitopes and is believed to promote SAT in NOD.H2h4 animals. 8 The SAT severity is well correlated with serum titers of autoantibodies against murine Tg (mTg; antibodies against thyroglobulin [TgAbs]). 9,10 While almost 100% of the mice develop thyroid lesions, serum T4 levels are again not significantly altered by the excessive iodide ingestion. 11,12
Increased serum levels of the T helper (Th) Th1 cytokine IFNγ with accumulation of peripheral Th1 cells are frequently observed in patients suffering of severe Hashimoto's thyroiditis. 13,14 Many reports demonstrate that AT is mainly a Th1-mediated autoimmune disease with a critical role played by IFNγ. 5,15,16 In contrast, interleukin-4 (IL-4) positively regulates the Th2 immune response, including B cell activation and the development of thyroiditis, as well as autoantibody production rely on B lymphocytes. 17 IL-4 is known to repress the Th1 immune response, but its implication in autoimmune diseases remains unclear. 18 IL-4 can suppress autoimmune diabetes in transgenic NOD mice with pancreatic IL-4 expression, 19 whereas IL-4 treatment worsens the disease progression in models of colitis or uveoretinitis. 20,21 In an adoptive model of EAT, IL-4 incubation of spleen cells during mTg priming results in lower levels of TgAbs. 22 Conversely, IL-4 has been shown to increase the severity of granulomatous EAT. 23 Nevertheless, genetic invalidation of IFNγ but not IL-4 protects the animals from SAT, demonstrating that IFNγ is required for disease development. 24
Recently, we have shown that thyroid IL-4 expression could mitigate hyperthyroidism in a mouse model of Graves' disease. 25 Knowing the antagonistic effect of IL-4 on the Th1 immune response, we have evaluated if the thyroid expression of IL-4 by thyrocytes (Thyr-IL4) could protect NOD.H2h4 animals from accelerated SAT.
Materials and Methods
Animals
Thyr-IL4 C57BL/6J mice overexpressing the murine IL-4 have been described elsewhere. 26 NOD.H2h4 mice 27 were kindly provided by Profs. Sandra McLachlan and Basil Rapoport. To generate Thyr-IL4 NOD.H2h4 mice, transgenic animals were backcrossed to wild-type (WT) NOD.H2h4 for at least six generations. Only N6 and N7 animals were used in the subsequent experiments. The majority of experiments were performed on the Thyr-IL4 NOD.H2h4 line 52, and some results were reproduced in the independent transgenic line 30.
Both males and females were exposed to 0.05% sodium iodide (NaI; 217638; Sigma-Aldrich) in drinking water or regular water (control group) from age 8 weeks to accelerate SAT. Blood was collected from the retro-orbital plexus after 8 weeks and at the sacrifice of the mice (16 weeks). Animals were polymerase chain reaction (PCR) genotyped from mouse tail DNA using primers targeting the transgenic construct. 26 All animal experiments were conducted in accordance with the guidelines and approval of the Institutional Animal Care and Use Committee at the “Université libre de Bruxelles” (CEBEA permits 505N/649N). A conventional animal housing facility was used to breed the animals in regulated humidity, temperature, and light/dark room conditions. Ad libitum access to food and water was also provided.
Enzyme-linked immunosorbent assay for autoantibodies to mouse Tg (TgAbs)
mTg was isolated from 30 to 35 thyroid lobes after homogenization in Dulbecco's phosphate-buffered saline (DPBS; supplemented with protease cocktail inhibitors (cOmplete Mini; Roche), filtrated through a 0.22 μm filter (VWR) and purified on a size exclusion column HiLoad 16/600 (GE HealthCare) as previously described. 28 Enzyme-linked immunosorbent assay for TgAbs was performed on 96-well plates (3355; Thermo Scientific) coated with 1.5 μg/mL mTg using sera (1:100 dilution) in duplicate. 29 Negative controls were sera from untreated young 8-week-old NOD.H2h4 and C57BL/6 animals. Values are presented as optical densities (OD) at 490 nm of the tested sera subtracted from the blank value (without mouse serum).
Serum thyrotropin assay
Mouse serum thyrotropin (TSH) levels were measured with a TSH bioassay using stable JP26 CHO cells expressing the human TSH receptor. 30 cAMP was measured by radioimmunoassay, and data were expressed as picomoles of cAMP/50,000 cells.
Thyroid histology procedures
Thyroid lobes were dissected and weighed before being frozen in Tissue-Tek optimal cutting temperature (4583 compound; Sakura). Thyroid blocks were sectioned at 5 μm, mounted on glass slides, and sections were stained with hematoxylin and eosin (Sigma-Aldrich) before being scanned with the NanoZoomer-SQ Digital Slide scanner (Hamamatsu).
For immunofluorescence staining, cryosections fixed in 4% paraformaldehyde were incubated in blocking solution (phosphate buffered saline containing 3% bovine serum albumin [Sigma-Aldrich], 5% horse serum [Invitrogen], and 0.3% Triton X-100 [Bio-Rad]) before incubation with primary antibodies: anti-CD106 (Vascular Cell Adhesion Molecule-1 [VCAM-1];550547; BD Pharmingen), anti-CD62E (E-selectin; 553749; BD Pharmingen), anti-CD4 (GK1.5 14-0041-82; Invitrogen), anti-CD8 (GK1.5 14-0081-82; Invitrogen), anti-B220 (FP10422050; MBL), and anti-CD45 (30F11; BioLegend) with anti-NKX2.1 antibody (ab76013; Abcam). After washing, the tissues were incubated with two secondary antibodies: anti-rabbit Alexa Fluor 488 (A21206; Invitrogen) and anti-rat Rhodamine Red-X (712-295-150; Jackson) and nuclei stained with DAPI (D9564; Sigma-Aldrich). The images were acquired on a Zeiss Axio Imager Zoom V16 microscope (Carl Zeiss) using a × 20 magnification and analyzed with Zen 3.3 (Blue edition) Software. Percentage of positive cells was calculated in multiple samples with QuPath software on whole thyroid cryosections. 31
Real-time quantitative PCR
Total RNA was extracted from one thyroid lobe using the RNeasy Mini Kit (HB-0435; Qiagen). After DNAse treatment, 1 μg of total RNA was reverse-transcribed with Superscript II reverse-transcriptase (Invitrogen). Real-time quantitative PCR was performed using KAPA Fast SYBR Green (Kapa Biosystems) on a C1000 Touch Thermal Cycler CFX96 Real-Time System (Bio-Rad) with gene-specific, intron-spanning primers (Supplementary Table S1). Relative RNA expressions were analyzed with the Pfaffl method after normalization to Retention Endoplasmic Reticulum 1 homolog (RER1) and TATA-box Binding Protein (TBP) genes. 32,33 Primer sequences were selected to target the 5′ untranslated region of the IL-4 transcript to amplify selectively the endogenous IL-4 isoform.
Flow cytometry
Immediately after dissection, single-cell suspensions were prepared from individual mouse thyroid, immunostained, and analyzed on a FACS BD LSRFortessa™ (BD Biosciences) (Supplementary Methods S1).
Statistical analyses
Statistical analyses were achieved using the Mann–Whitney U test (GraphPad Prism 6.0 software; GraphPad Software, Inc., San Diego, CA). Values were considered significant at *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, and the data of individual mice were presented in the graphs with the median and interquartile range.
Results
Severe accelerated SAT in Thyr-IL4 NOD.H2h4 animals
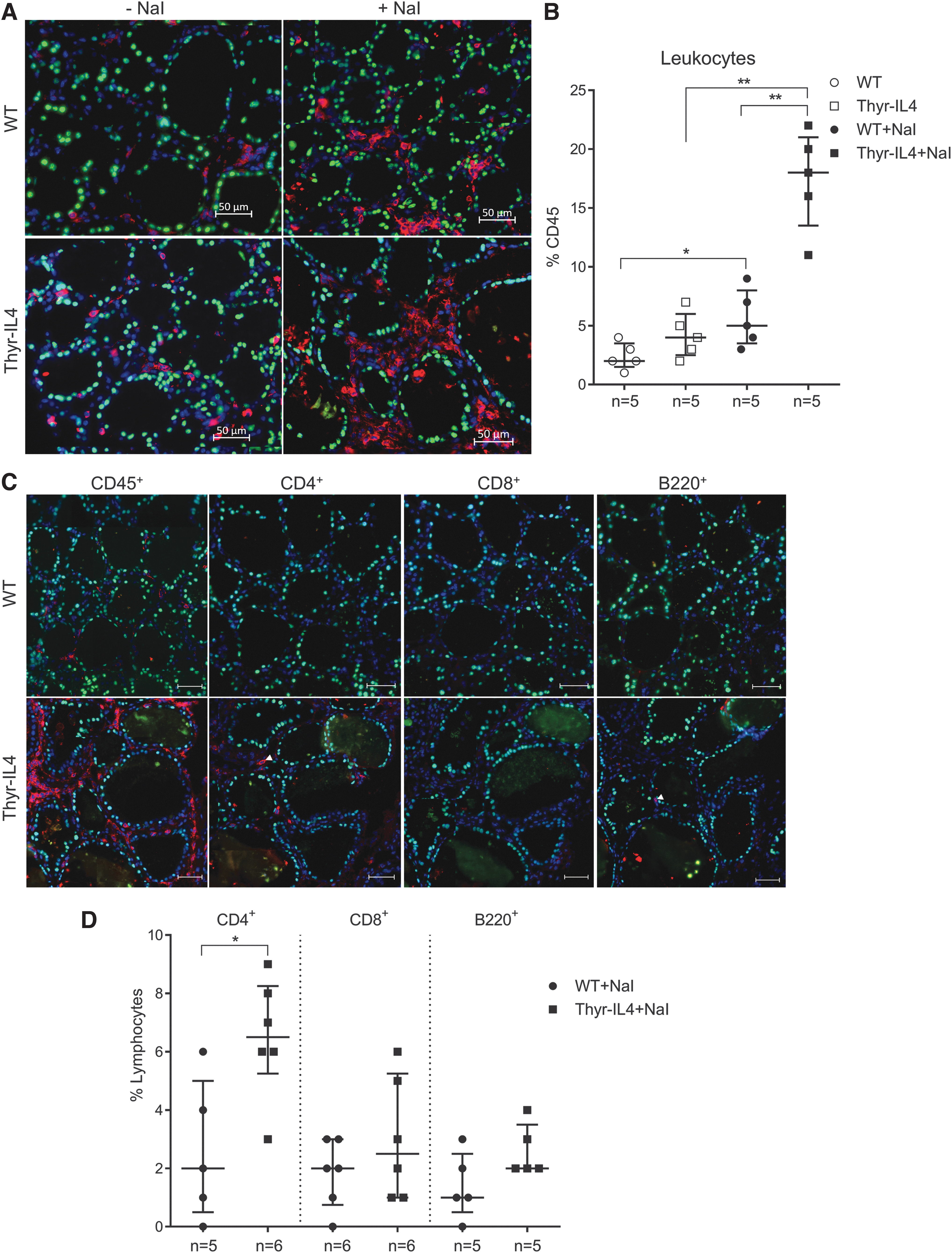
WT and Thyr-IL4 NOD.H2h4 animals were treated with sodium-iodized water (0.05% NaI) during 8 and 16 weeks. Circulating TgAbs were constantly more elevated in Thyr-IL4 animals relative to WT after exposure to NaI. A higher number of transgenic animals presented this increase, although the incidence did not reach statistical significance (Fig. 1A). Consistent with TgAb titers, the severity of accelerated SAT was markedly enhanced in Thyr-IL4 with multiple foci of mononuclear cell infiltrates throughout the thyroid parenchyma, replacing the normal thyroid architecture in some transgenic animals (Fig. 1B). Anti-CD45 immunostaining confirmed the presence of numerous leukocytes in the thyroid of WT and Thyr-IL4 animals after 16 weeks of NaI treatment (Fig. 2A). Quantitatively, the percentage of CD45+ cells in treated mice was drastically increased in transgenic (38%) relative to WT (18%) animals (Fig. 2B). Moreover, age matched Thyr-IL4 animals under regular water displayed also more frequent intrathyroidal leukocytes relative to WT littermates.
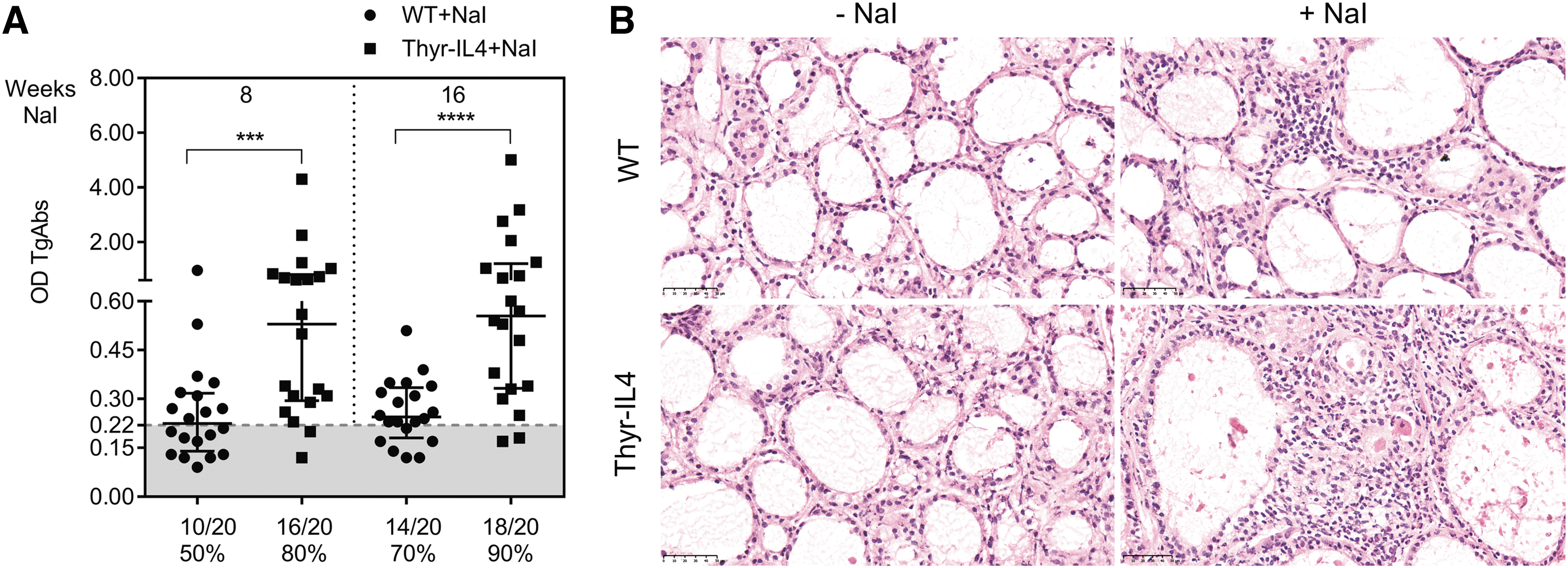
Thyr-IL4 NOD.H2h4 mice developed severe SAT after treatment with iodized drinking water (0.05% NaI). (
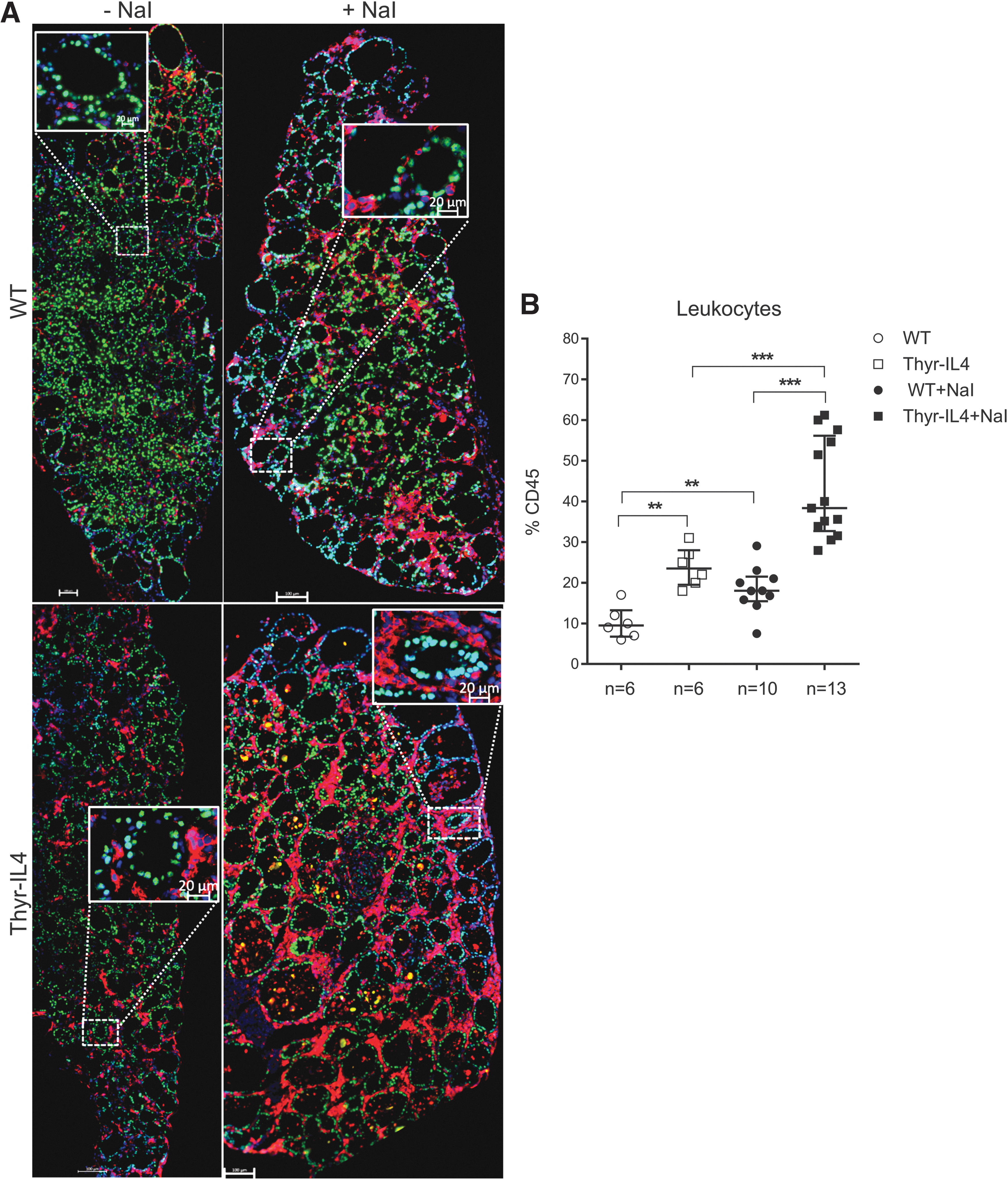
Extensive leukocyte infiltrations are observed in the thyroids of Thyr-IL4 NOD.H2h4 mice. (
Upregulation of the endothelial activation markers VCAM-1 and E-selectin was also enhanced in the thyroids of NaI treated Thyr-IL4 mice (Supplementary Fig. S1). As previously reported in WT NOD.H2h4, 34 we found that the majority of CD45+ cells encompassed CD4+ T lymphocytes and B220+ B cells forming lymphoid clusters, while CD8+ T cells were more scattered in the thyroid tissue (Fig. 3A). This inflammatory cell distribution was conserved in transgenic treated animals, although the respective cellular populations were all significantly enhanced (Fig. 3B), reflecting the intense thyroid inflammation. After antibody staining and gating on intact cells, the different immune cell populations were identified by flow cytometry as a percentage of the total number of CD45+ cells (Supplementary Fig. S2). At the end of the NaI treatment, the leukocyte population infiltrating the thyroid of Thyr-IL4 mice was significantly enriched in CD8+ T and B220+ B lymphocytes (Fig. 4).
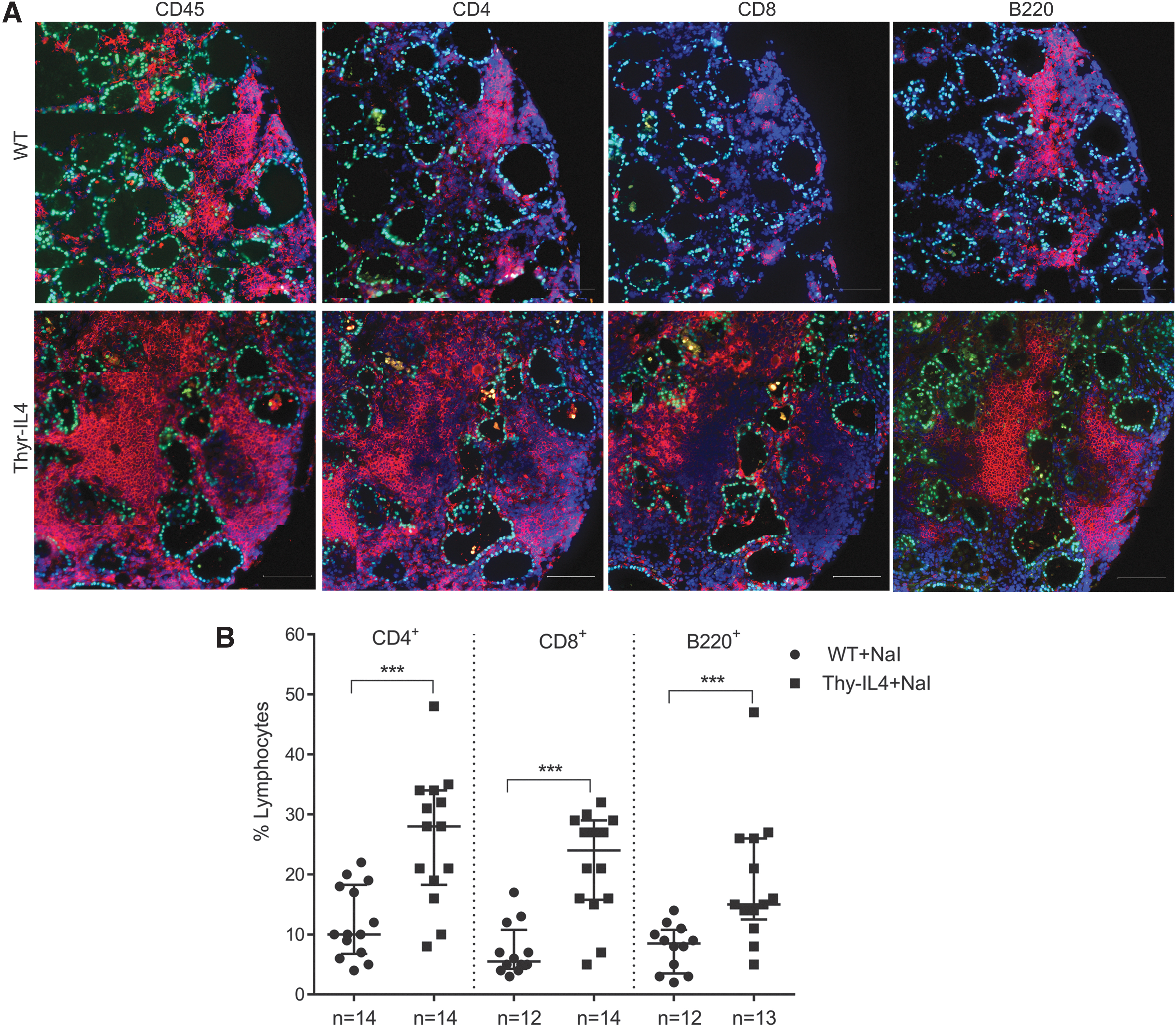
Characterization of T and B cell populations in the thyroids of WT and Thyr-IL4 NOD.H2h4 mice exposed 16 weeks to iodized water. (
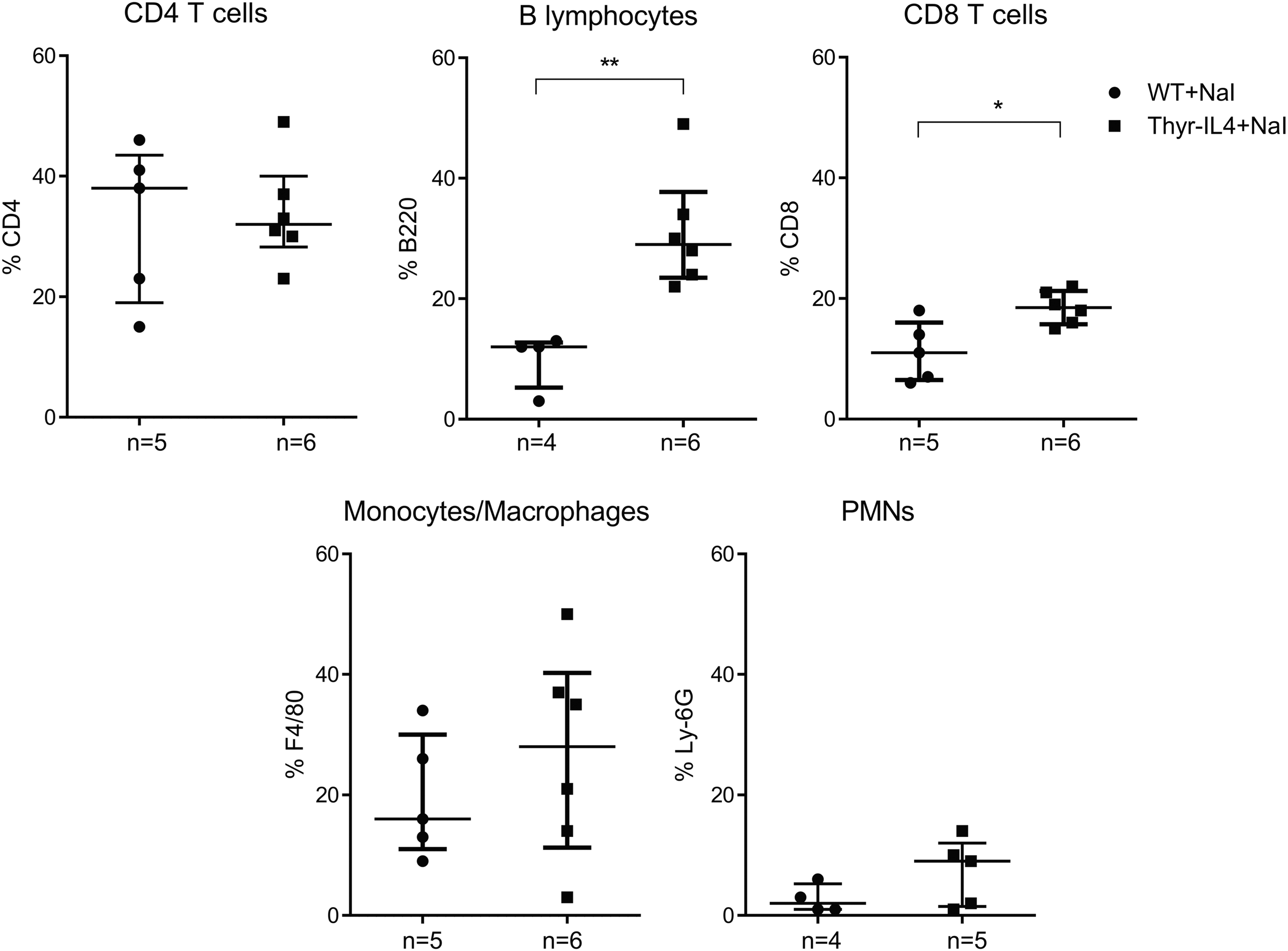
CD45+ immune cell distribution in thyroid infiltrates by flow cytometry. Thyroid single cell suspensions from 16 weeks of NaI treated WT and Thyr-IL4 NOD.H2h4 animals were stained with the respective fluorescent-labeled antibodies. CD45+ vs. immune cells (CD4, CD8, F4/80, Ly-6G, and B220) were plotted after gating on the intact cell population (DAPI− cells). The CD4, CD8, F4/80, LY-6G, and B220 immune cell populations are presented as a percentage of the total number of CD45+ cells. The numbers of mice studied in each group are indicated. *p < 0.05, **p < 0.01.
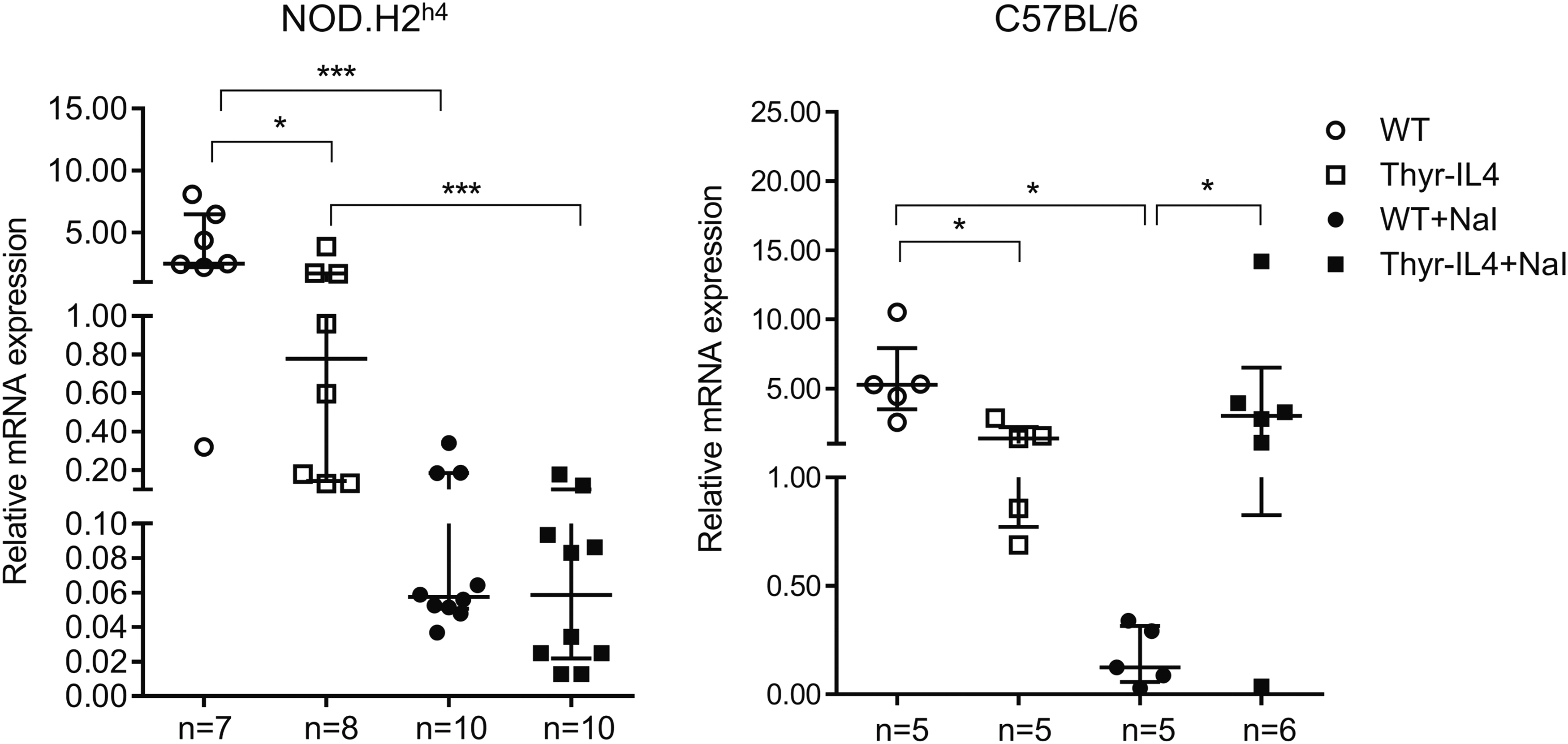
Finally, the messenger RNA (mRNA) expression of the main Th1 and Th2 cytokines was quantified in the thyroid lobes of animals 16 weeks after receiving regular or iodide-supplemented water (Fig. 5). As expected, excessive iodide supply of WT NOD.H2h4 moderately induced the expression of IFNγ, TNFα, IL-4, and IL-10. 10 Consistent with the substantial thyroid infiltrate, the expression of TNFα, IL-4, and IL-10 was considerably augmented in NaI treated Thyr-IL4 animals, as well as TGFβ, IL-13, and IL-17. Accumulation of Th17 cells was reported in the thyroid of NOD.H2h4 animals, and IL-17 −/− animals develop reduced iodide-induced SAT lesions. 35 Surprisingly, thyroid expression of IFNγ was already elevated in transgenic mice receiving regular water and was not statistically augmented after accelerated SAT. These data were reproduced in the second independent Thyr-IL4 NOD.H2h4 line 30 (Supplementary Fig. S3).
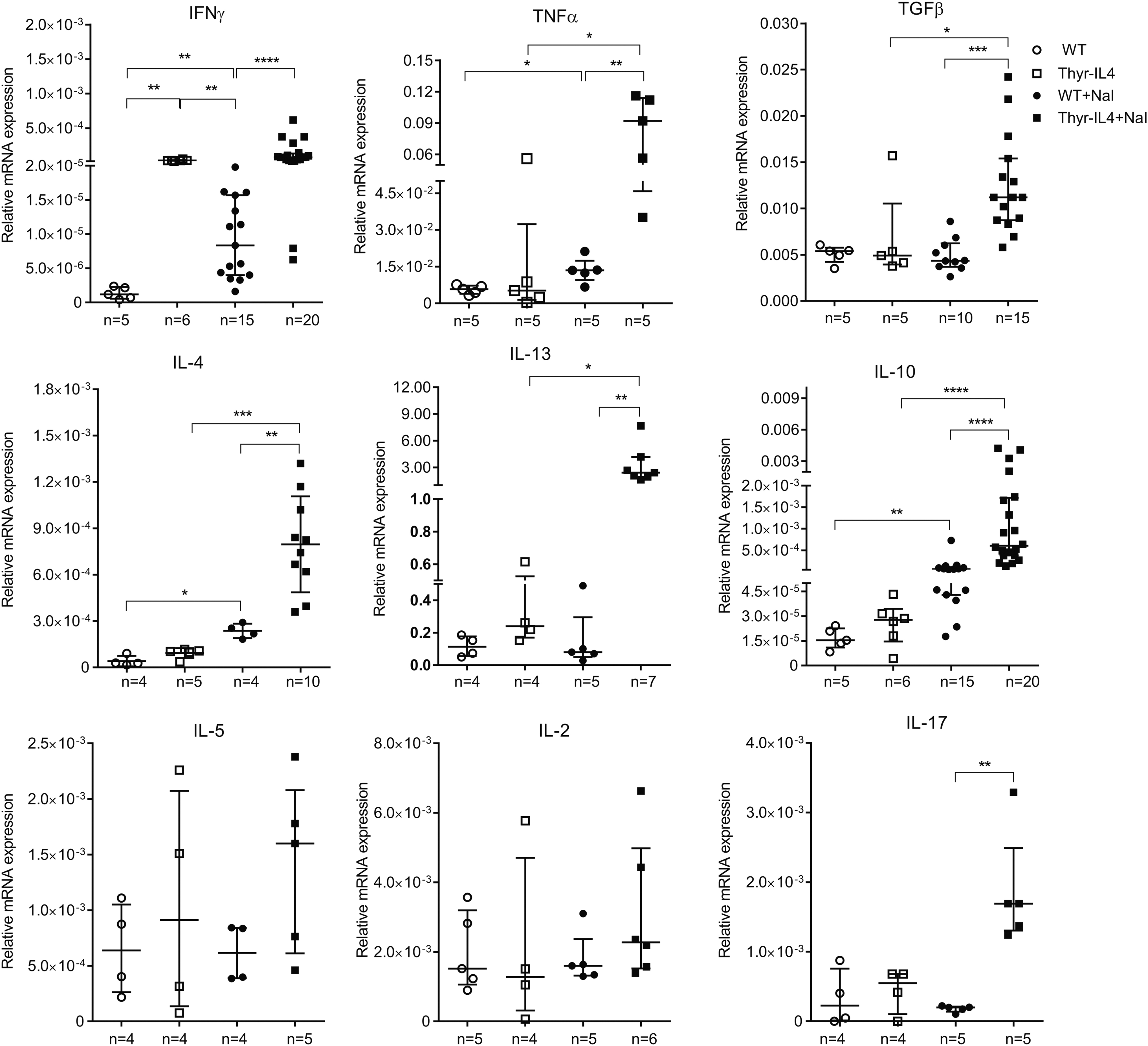
Intrathyroidal cytokine mRNA expression in NOD.H2h4 mice. Relative mRNA expression of cytokines was determined by RT-qPCR in thyroids collected after 16 weeks of NaI treatment. The data are presented by median with interquartile range. IL-4 corresponds to the endogenous IL-4 isoform mRNA. The numbers of mice studied in each group are indicated. IL-4, interleukin-4; mRNA, messenger RNA; RT-qPCR, real-time quantitative polymerase chain reaction. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001.
These findings demonstrated that ectopic expression of IL-4 by TFCs increased the severity of iodide-induced SAT lesions with large lymphocytic infiltrations and could influence the composition and/or the activation of the immune cells infiltrating the inflamed tissue.
Thyroid inflammation in SAT-resistant Thyr-IL4 C57BL/6 animals on iodized water
Compared to disease-prone NOD.H2h4 animals, virtually none of the parental C57BL/6 presented elevated TgAbs after 8 or 16 weeks on iodized-supplemented water (Fig. 6A). Unexpectedly, moderate mononuclear infiltrates were evident in the thyroids of transgenic mice at the end of the treatment (Fig. 6B). Immunostaining confirmed the leukocytic nature of the thyroid infiltrate, with quantitatively more CD45+ cells in Thyr-IL4 animals (18%) relative to WT littermates (5%) on NaI (Fig. 7A, B). Notably, these infiltrating cells could penetrate into the colloidal lumen of transgenic thyroids. Contrary to NOD.H2h4 mice, immunophenotyping of these immune cells on serial sections revealed that a very limited number of leukocytes were positives for CD4, CD8, or B220 (Fig. 7C). Only the frequency of CD4+ T cells among total intrathyroidal cells (DAPI+) was slightly but significantly augmented in Thyr-IL4 thyroids after the treatment (Fig. 7D).
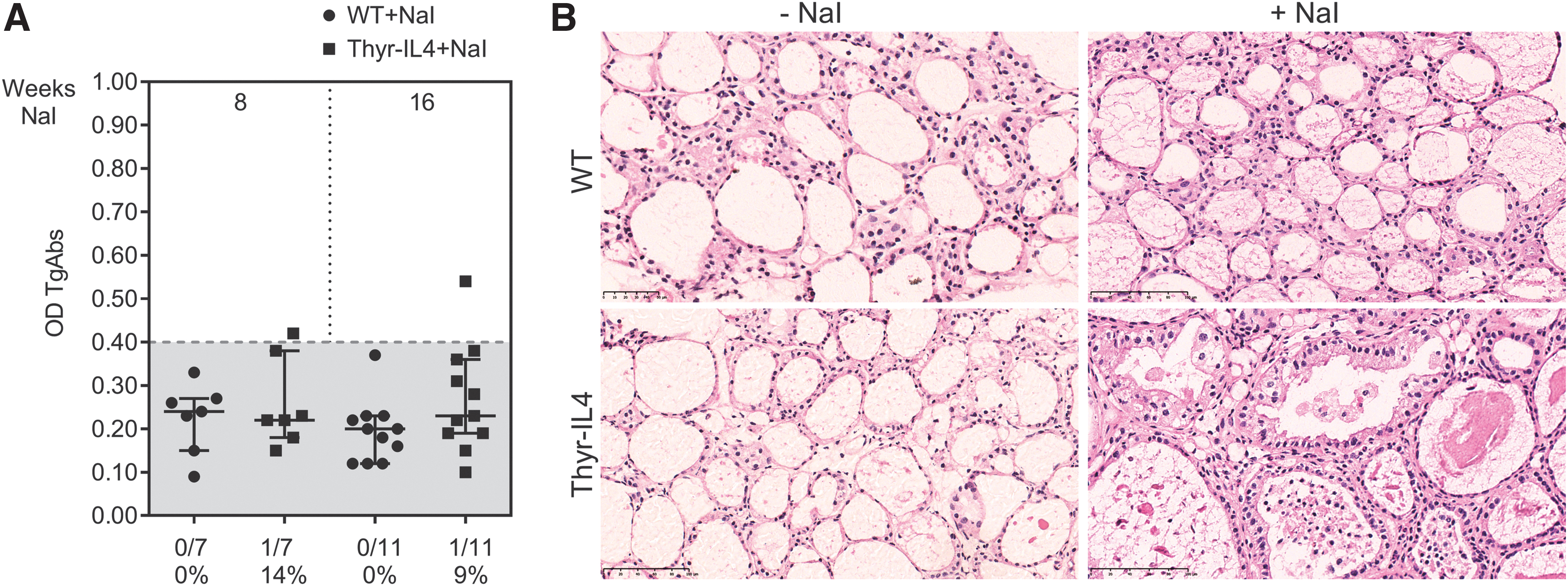
Thyr-IL4 C57BL/6 mice developed large thyroid leukocyte infiltrations in absence of elevated TgAbs after 16 weeks on iodized water (NaI). (
Intrathyroidal immune cell populations observed in treated Thyr-IL4 C57BL/6 mice were different from NOD.H2h4. (
While the induction of intrathyroidal TNFα, TGFβ, IL-4, and IL-13 was reproduced in Thyr-IL4 C57BL/6 relative to WT mice after NaI exposure, no significant induction of IFNγ or IL-10 could be measured (Fig. 8). Transgenic animals on regular water presented also elevated expression of IL-13 and IFNγ compared to WT. The major difference between the two mouse strains concerned IL-5, which was not modulated in the SAT-prone NOD.H2h4 model, but clearly induced in the thyroids of NaI treated WT and transgenic C57BL/6 mice. We quantified the levels of some circulating cytokines in the serum of NOD.H2h4 and C57BL/6 treated animals (Supplementary Fig. S4). Despite some significant modulations observed, no definitive conclusion could be drawn due to the limited sensitivity of the method. Nevertheless, overexpression of circulating IL-4 was noticeably detected in the serum of all Thyr-IL4 animals.
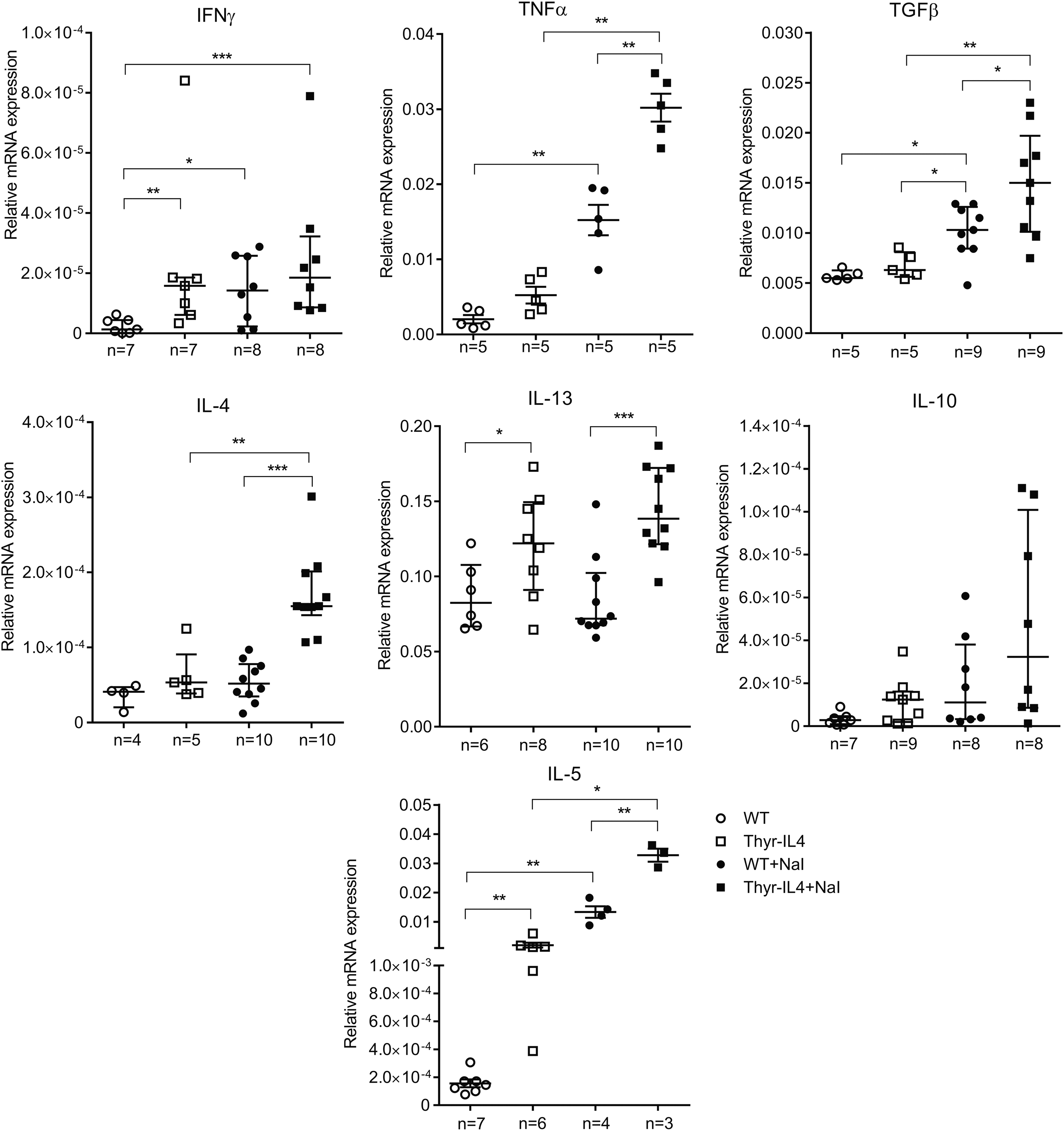
Intrathyroidal cytokine mRNA expression in C57BL/6 mice. Relative mRNA expression of cytokines was determined by RT-qPCR on thyroids collected after 16 weeks on iodized water. The data are presented by median with interquartile range. IL-4 corresponds to the endogenous IL-4 isoform mRNA. The numbers of mice studied in each group are indicated. *p < 0.05, **p < 0.01, ***p < 0.001.
All these data demonstrated that a constitutive expression of IL-4 by the thyrocytes could induce non-AT after high iodide intake in SAT-resistant C57BL/6 animals.
Altered thyroid function in Thyr-IL4 animals on iodized-supplemented water
In contrast to WT animals, both NaI-treated Thyr-IL4 strains developed marked thyroid enlargement associated with elevated circulating TSH levels and lower serum T4 (Fig. 9, Supplementary Fig. S5 and data not shown). Nontransgenic animals showed normal thyroidal Nis downregulation after 16 weeks of NaI supply, demonstrating their capacity to escape the Wolff–Chaikoff block to resume TH synthesis 12,36 (Fig. 10). This degree of Nis repression was similar in Thyr-IL4 NOD.H2h4 despite their severe hypothyroid phenotype. In contrast, Nis expression was inappropriately preserved in Thyr-IL4 C57BL/6 animals compared to WT littermates, suggesting a failure in limiting iodide uptake in the thyroid.
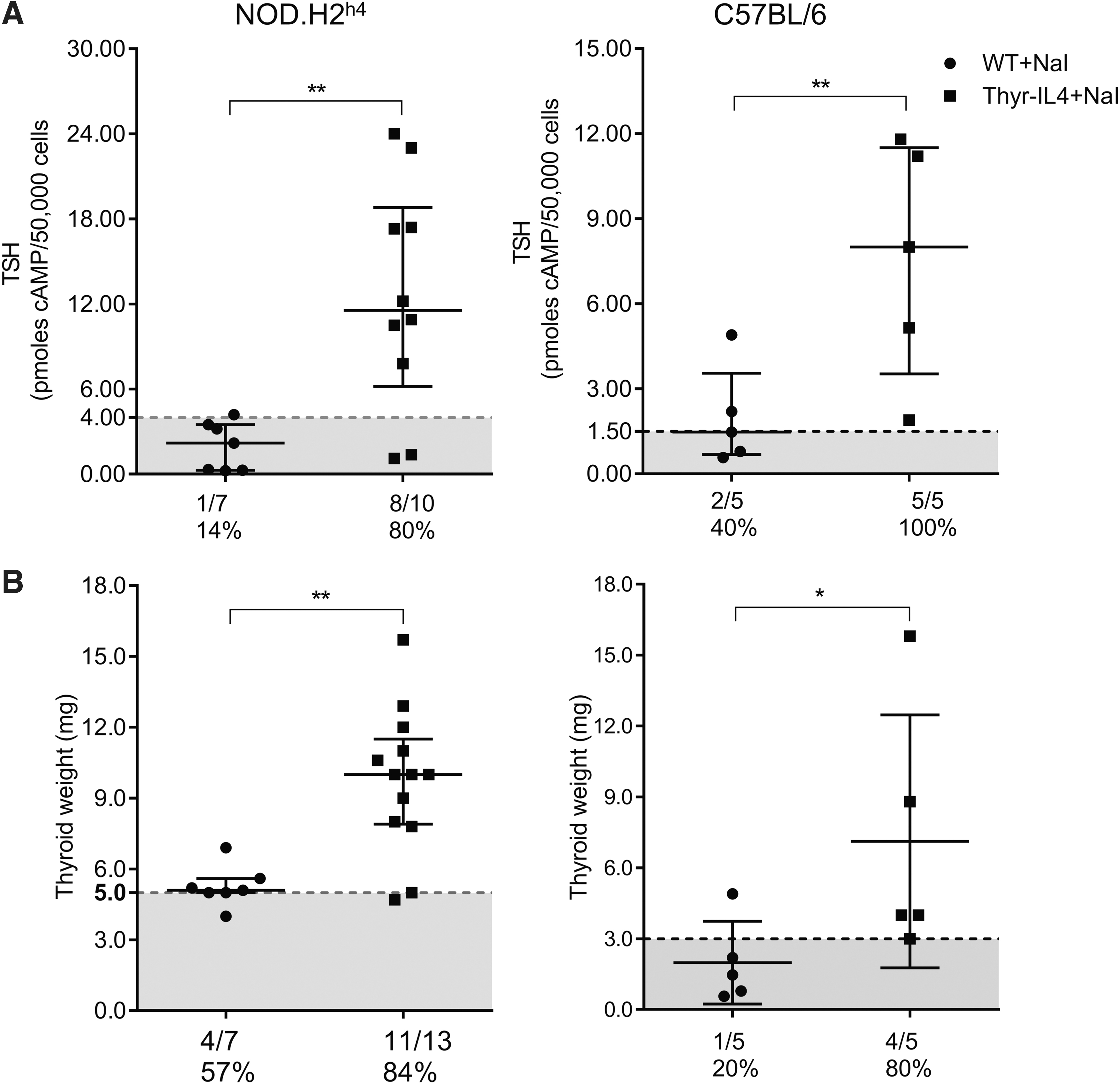
Altered thyroid function in Thyr-IL4 males exposed to iodized water. (
Relative expression of Nis mRNA in NOD.H2h4 and C57BL/6 mice treated for 16 weeks (+NaI) or not with iodized water. The number of mice is presented under each group. The values of individual mice that are presented in the graphs and median with interquartile range are shown. *p < 0.05, ***p < 0.001.
Discussion
Meta-analyses have revealed some significant associations between IL-4 genetic variants and the risk to develop autoimmune thyroid diseases. 37,38 Based on the ability of IL-4 to counteract the Th1 immune response, it has been proposed that it could be used as a therapeutic agent to treat autoimmune pathologies. 39 Using Thyr-IL4 animals, we explored whether a thyroid expression of IL-4 could fulfill this assumption in the context of AT. However, we have clearly demonstrated that Thyr-IL4 NOD.H2h4 developed severe SAT lesions after excessive iodide ingestion. Thyroid inflammation in NOD.H2h4 animals has been well documented. 40 The infiltrate is mainly composed of CD4+, CD8+, and B220+ lymphocytes associated with elevation of thyroid Th1/Th2 cytokine expression, 10,34 and all these SAT hallmarks were exacerbated in treated Thyr-IL4 NOD.H2h4 animals. The leukocyte enrichment in CD8+ T and B lymphocytes could explain the massive thyroid destruction with high TgAbs observed in Thyr-IL4 mice. However, we cannot exclude that the artificial thyroid expression of the IL-4 transgene would be responsible for this unexpected phenotype.
The major limitation of this study is the absence of mechanisms by which IL-4 enhances thyroiditis. The present data might seem somewhat inconsistent with previous studies showing that IL-4 knockout (KO) NOD.H2h4 normally developed accelerated SAT. 24 However, the published data did not exclude a role of IL-4 during the disease progression. Similar unreliable findings were reported in NOD.H2h4 mice with thyroid expression of IFNγ. Actually, Thyr-IFNγ animals were protected against SAT, which was unexpected considering the decisive role of this cytokine in AT. 41,42 In Thyr-IL4 mice, local accumulation of IL-4 could improve the capacity of autoreactive B cells to respond to thyroid autoantigens. Moreover during SAT progression in WT NOD.H2h4, elevation of thyroidal IL-4 is only detected late in the disease 10 with endothelial induction of VCAM-1 43 known to be regulated by IL-4. 44 Therefore, the precocious thyroid expression of IL-4 in Thyr-IL4 mice could also facilitate the leukocyte infiltration by early VCAM-1 induction (Supplementary Fig. S1).
Expression of the Intracellular Adhesion Molecule-1 (ICAM-1) was shown on thyrocytes from Hashimoto's patients and positively regulated by IL-4 in bronchial and colonic epithelial cells. 45 –47 The corresponding IL-4 receptor being expressed in TFCs, 48 it is possible that IL-4 could also mediate ICAM-1 expression in Thyr-IL4 mice promoting thyroid inflammation. Furthermore, the systemic humoral immune response in transgenic animals could be improved by the elevated IL-4 serum levels.
A second consequence of the ectopic IL-4 overexpression in absence of NaI treatment was the Th1 polarization of leukocytes infiltrating the transgenic thyroid tissue illustrated by the intrathyroidal increase of IFNγ. IL-4 is able to accelerate Th1-mediated colitis with upregulation of IFNγ in the intestine. 20 Thyrocytes from Hashimoto's patients present IFNγ-induced major histocompatibility complex (MHC) II that is repressed in IFNγ KO NOD.H2h4. 24,49 In addition, IFNγ acts synergistically with iodine to increase the expression of ICAM-1 on thyrocytes of NOD.H2h4, suggesting that ICAM-1 could be a key disease susceptibility factor. 42 Further experiments with blocking antibodies would help to discriminate the respective role of IL-4 and IFNγ in Thyr-IL4 NOD.H2h4 accelerated SAT.
In contrast to NOD.H2h4 presenting the MHC haplotype H2k, excessive iodide intake does not induce AT in C57BL/6 having the MHC haplotype H.2b,12,50 Surprisingly, a significant number of treated Thyr-IL4 C57BL/6 developed substantial mononuclear thyroid infiltrates. However, this was not associated with elevated TgAbs, suggesting that the thyroid or systemic expression of IL-4 was not sufficient to trigger autoimmunity in C57BL/6. A similar phenotype was observed in C57BL/6 with thyroid expression of the chemokine CCL21 that present large lymphocytic thyroid lesions without TgAbs. 51
Nevertheless, our findings support a role of IL-4 in NaI-induced thyroid inflammation. Similar to Thyr-IL4 NOD.H2h4, thyroid elevation of Th1 and Th2 cytokines was measured. However, the limited number of CD4+, CD8+, and B lymphocytes found in the leukocyte infiltrates and the marked IL-5 increase suggested a different disease etiology in transgenic C57BL/6 with a SAT-resistant genetic background. IL-5 implication in AT is still unclear with only few studies reporting an elevation of serum IL-5 in Hashimoto's patients. 52,53 Further experiments should be conducted to better characterize the immune cell population in Thyr-IL4 C57BL/6 mice.
Patients with subclinical thyroid disorders, like Hashimoto's thyroiditis, could develop hypothyroidism after excessive iodine ingestion. 3 However, the various mouse models generated to study AT do not frequently develop hypothyroidism. 7,11,29,51 In contrast, our transgenic animals present iodide-induced thyroid inflammation concomitant with primary hypothyroidism. Hence, they constitute a new valuable tool to better characterize the pathophysiology of this autoimmune thyroid disease.
It is commonly accepted that inflammatory cytokines negatively affect the thyroid function, including NIS inhibition. 54 In addition, impairment to reduce the excessive iodide intake through the Wolff–Chaikoff escape phenomenon could also result in hypothyroidism. 55 The thyroid dysfunction in transgenic C57BL/6 mice could be caused by the failure to reduce iodide excess associated with inflammatory cytokines produced locally in the inflamed tissue. The inappropriate preservation of Nis expression could be linked to the moderate inflammation preserving the TFC response to elevated TSH levels that may counterbalance Nis downregulation. In contrast, the large thyroid lesions in Thyr-IL4 NOD.H2h4 could most probably provoke TH synthesis defect as observed in the final evolution of Hashimoto's thyroiditis characterized by the destruction and fibrous replacement of TFCs.
In conclusion, our findings demonstrated that expression of IL-4 by thyrocytes enhanced the severity of accelerated SAT in Thyr-IL4 NOD.H2h4 animals and promoted leukocyte infiltrations in the thyroid of SAT-resistant transgenic C57BL/6 mice. Moreover, disease development in both transgenic strains was associated with the emergence of a substantial hypothyroidism.
Footnotes
Acknowledgments
The authors gratefully acknowledge Profs. Basil Rapoport and Sandra McLachlan (Thyroid Autoimmune Disease Unit, Cedars-Sinai Research Institute, Los Angeles, CA) for providing the NOD.H2h4 mice and their continuous interest and judicious advice in the present study. The authors thank Jean-Marie Vanderwinden for the use of the microscope facility. Special thanks to Sang Van Tran for its excellent technical assistance.
Authors' Contributions
K.M., F.M., and X.D.D. designed the project, and K.M. performed all the experiments. S.D. participated in some part of the project involving the manipulation of the mice. X.D.D. wrote the article. K.M., J.-E.D., and F.M. helped and edited the article. X.D.D. provided conceptual advice. All coauthors have approved the article for submission.
Author Disclosure Statement
All the authors have no conflicts of interests to disclose.
Funding Information
This work was supported by the “Fonds de la Recherche Scientifique” (FRS-FNRS; Grant Nos. J002114F–J005918F) and the Fund Doctor J.P. Naets, managed by the King Baudouin Foundation.
Supplementary Material
Supplementary Methods S1
Supplementary Table S1
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5