
Abstract
Background:
Thyroid hormone (triiodothyronine [T3]) is essential for development and organ metabolism in all vertebrates. T3 has both genomic and nongenomic effects on target cells. While much has been learnt on its genomic effects via T3 receptors (TRs) in vertebrate development, mostly through TR-knockout and TR-knockin studies, little is known about the effects of T3 on gene expression in animals in the absence of TR. We have been studying Xenopus metamorphosis as a model for mammalian postembryonic development, a period around birth when plasma T3 level peaks and many organs/tissues mature into their adult forms. We have recently generated TR double knockout (TRDKO) Xenopus tropicalis animals. This offers an opportunity to compare the effects of T3 on global gene expression in tadpole tissues in the presence or absence of TR.
Methods:
We analyzed the effects of T3 on gene expression in tadpole tail and intestine by using RNA-seq analysis on wild-type and TRDKO tadpoles with or without T3 treatment.
Results:
We observed that removing TRs reduced the number of genes regulated by T3 in both organs. Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analyses revealed that T3 affected distinct biological processes and pathways in wild-type and TRDKO tadpoles. Many GO terms and KEGG pathways that were enriched among genes regulated in wild-type tissues are likely involved in mediating the effects of T3 on metamorphosis, for example, those related to development, stem cells, apoptosis, and cell cycle/cell proliferation. However, such GO terms and pathways were not enriched among T3-regulated genes in TRDKO tadpoles. Instead, in TRDKO tadpoles, GO terms and pathways related to “metabolism” and “immune response” were highly enriched among T3-regulated genes. We further observed strong divergence in the TR-independent nongenomic effects of T3 in the intestine and tail.
Conclusions:
Our data suggest that T3 has distinct and organ-dependent effects on gene expression in developing tadpoles. The TR-mediated effects are consistent with the metamorphic changes, in agreement with the fact that TR is necessary and sufficient to mediate the effects of T3 on metamorphosis. T3 appears to have a major effect on metabolism and immune response via TR-independent nongenomic processes.
Introduction
Thyroid hormone (triiodothyronine [T3]) is essential for normal development as well as adult organ metabolism in vertebrates. 1,2 In human, plasma T3 level peaks in the several months around birth when many organs/tissues mature into their adult forms, a period referred to as postembryonic development. 3 Abnormal T3 level during postembryonic development causes serious health problems, including intellectual deficits and abnormal metabolic rate. 4,5 Anuran metamorphosis is the most dramatic process regulated by T3 and mimics mammalian postembryonic development. 6 –8
T3 regulates transcription via T3 receptors (TRs). 2,9 Studies on Xenopus laevis have suggested a dual function model for TR during anuran development. That is, unliganded TR recruits corepressors to T3-inducible genes in premetamorphic tadpoles to repress their expression and prevent premature metamorphosis, while liganded TR recruits coactivators to activate these genes to induce metamorphosis. 10 There are two TR genes, TRα and TRβ, in vertebrates. 9,11 Knocking out TRα or TRβ has distinct effects on different organs during metamorphosis in diploid Xenopus tropicalis. 12 –19
Somewhat surprisingly, tadpoles lacking both TRα and TRβ can develop to metamorphic climax with many adult organs formed, although they are then stalled at stage 61 for about 2 weeks before death. 13,20 This contrasts with tadpoles that cannot synthesize T3, which are stalled before the onset of metamorphosis (stage 54). 21,22 While it is possible that the depression of target genes upon TR knockout allows adult organ development, the finding may also raise the question whether T3 can regulate organ development through TR-independent nongenomic actions.
T3 has various nongenomic effects that involve binding to plasma membrane. 2,23 –26 For example, thyroxine (T4), the precursor for T3, can bind to integrin αvβ3 to affect angiogenesis and cell proliferation. 26 T3 can modulate transport systems such as ATP-binding cassette transporters and affect the immune system and the phosphorylation in PI3K/Akt/mTOR pathway. 27 –30 However, little is known about the global gene expression changes induced by nongenomic action of T3 in the absence of TR, especially in developing animals.
In this study, we have carried out RNA-seq analyses on the intestine and tail of wild-type and TR double knockout (TRDKO, i.e., lacking both TRα and TRβ) X. tropicalis tadpoles treated with T3. (Note that without TR, the immediate action of T3 is to affect cellular events via nongenomic pathways. Such effects in turn affect downstream changes in mRNA levels by altering mRNA stability and/or transcription. For simplicity, we consider such changes in the absence of TR to be nongenomic effects of T3.) We discovered that TRDKO reduced T3-regulated genes in both organs with few commonly regulated genes between wild-type and TRDKO organs. We showed that many Gene Ontology (GO) terms and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways likely involved in metamorphosis were enriched among T3-regulated genes in wild-type but not TRDKO organs, while many GO terms and KEGG pathways related to metabolism and immune response were enriched in TRDKO but not wild-type organs.
Materials and Methods
All experiments involving X. tropicalis animals were carried out as approved by the Eunice Kennedy Shriver National Institute of Child Health and Human Development (NICHD) Animal Use and Care Committee. The details are presented in the Supplementary Data.
Results
TRDKO reduces the number of genes regulated by T3 in both the intestine and tail
To reveal the role of TR in gene expression by T3 during Xenopus development, we carried out RNA-seq analysis on the intestine and tail from wild-type and TRDKO tadpoles at stage 54 with or without 18-hour T3 treatment to identify differentially expressed genes (DEGs), that is, T3-regulated genes (Supplementary Table S1). To validate the data, we selected 13 DEGs with T3 regulation of fivefold or more and analyzed their expression independently by reverse transcription polymerase chain reaction (RT-qPCR) and found that all were consistent with RNA-seq data (Supplementary Fig. S1). Additionally, the regulation of three well-known direct T3 target genes, hal2, 31 mmp11, and thrb, 20 was confirmed by both RNA-seq and RT-qPCR analyses (Supplementary Fig. S2).
There were 2060 DEGs with T3 regulation of twofold or more in wild-type intestine but only 869 DEGs in TRDKO intestine, with 356 DEGs in common (Fig. 1A and Supplementary Table S3). In wild-type intestine, there were 1002 upregulated and 1058 downregulated DEGs, compared with only 338 upregulated and 531 downregulated DEGs in TRDKO intestine (Fig. 1B). Interestingly, T3 regulated distinct genes in wild-type and TRDKO intestine (Fig. 1C). Even among 356 DEGs genes common between wild-type and TRDKO intestine, more than half were regulated by T3 in opposite manner in wild-type and TRDKO intestine (Fig. 1C).

There are more DEGs in the intestine of premetamorphic wild-type tadpoles than TRDKO tadpoles after T3 treatment. (
The reason for this is unclear. It is possible that T3 causes opposite effects on their expression via TR-dependent pathway and TR-independent pathway, with the TR-dependent pathway having a bigger effect. Thus, in the wild-type tadpoles, the TR-dependent effect dominates, while in the TRDKO tadpoles, the T3 effect from TR-independent pathway becomes detectable. In any case, our findings indicate that T3 regulates mostly different genes in the presence and absence of TR in premetamorphic tadpole intestine.
Similar analysis identified 1848 DEGs in wild-type tail while only 774 DEGs in TRDKO tail, with only 177 common DEGs (Fig. 2A and Supplementary Table S3). There were 840 upregulated and 1008 downregulated DEGs in wild-type tail compared with only 256 upregulated and 518 downregulated DEGs in TRDKO tail (Fig. 2B). Like in the intestine, T3 regulated distinct genes in wild-type and TRDKO tail (Fig. 2C). Even among the 177 common DEGs, more than half were regulated by T3 in opposite manner in wild-type and TRDKO tail (Fig. 2C).
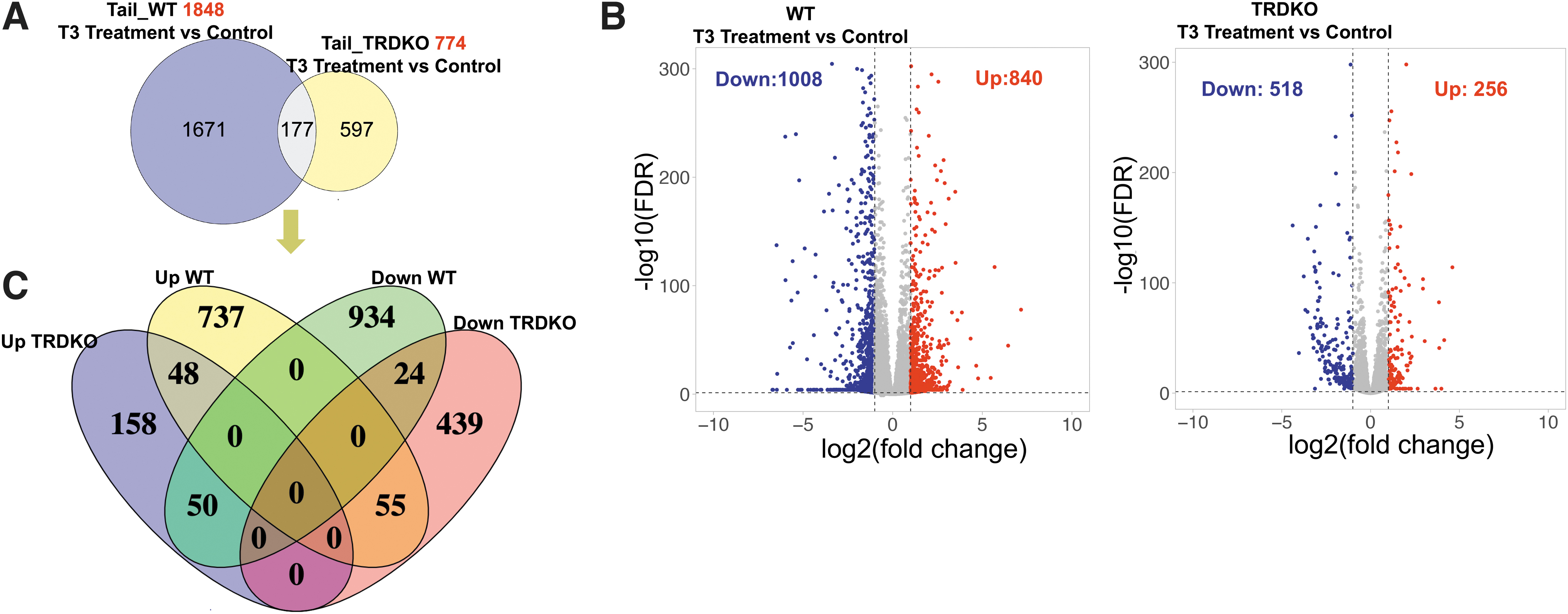
There are more DEGs in the tail of premetamorphic wild-type tadpoles than TRDKO tadpoles after T3 treatment. (
TRDKO affects GO terms and KEGG pathways regulated by T3 in the intestine
Earlier studies have shown that TRDKO delays/inhibits intestinal remodeling and TRDKO animals die at stage 61, before any significant tail length reduction. 9,20 Thus, TRDKO likely affects T3 regulation of processes involved in metamorphosis. To investigate this, we carried out GO and KEGG pathway analyses on the 2060 and 869 DEGs in wild-type and TRDKO intestine, respectively (Supplementary Tables S4 and S5). The most significantly enriched GO term was “development process” in wild-type intestine (Fig. 3A and Supplementary Table S4) and “response to organic substance” in TRDKO intestine (Fig. 3C and Supplementary Table S5).
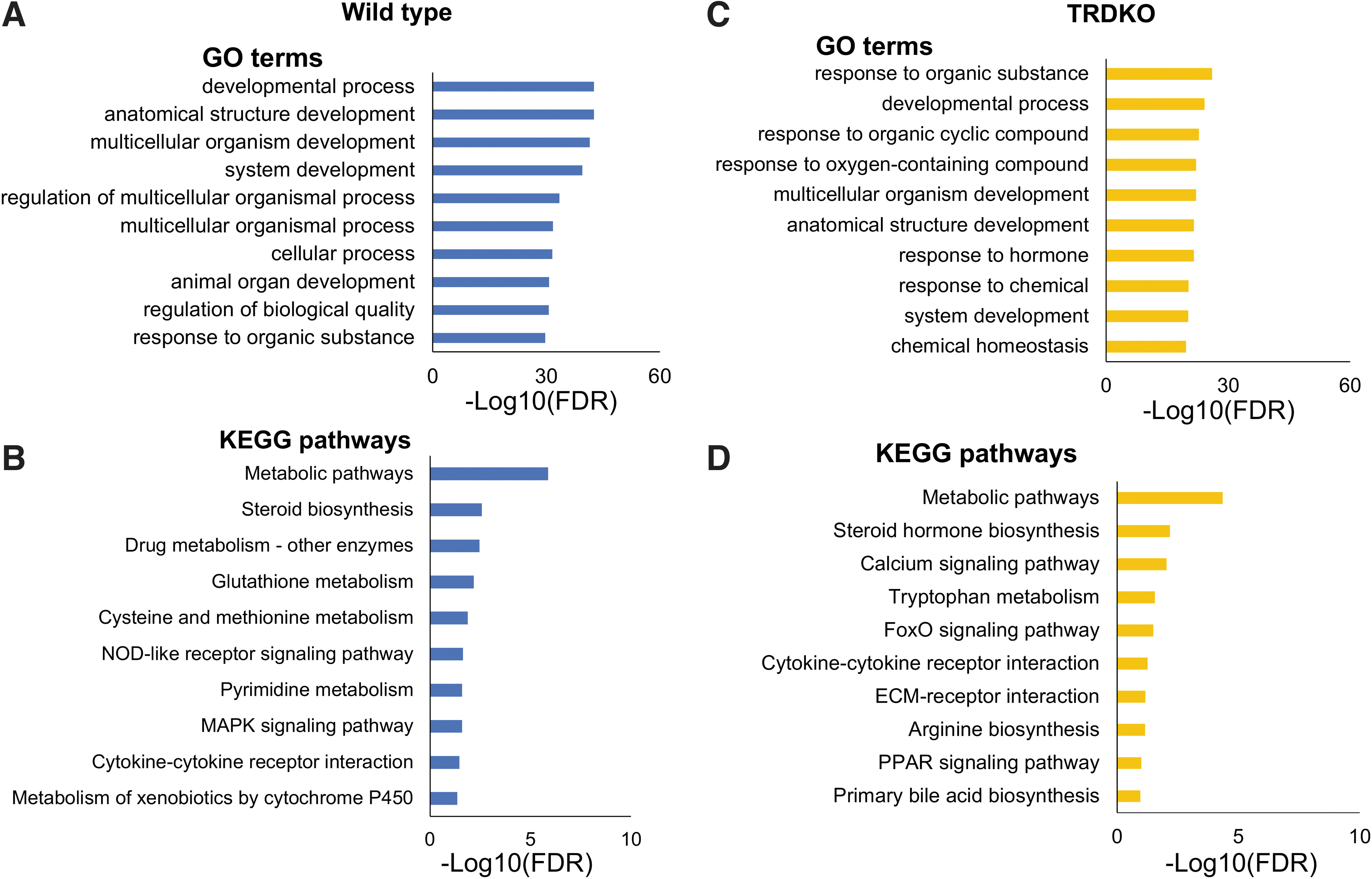
GO and KEGG analyses reveal that distinct GO terms/pathways are regulated by T3 in the intestine of wild-type (
In addition, there were many more development-related GO terms among the top 10 most significantly enriched GO terms in wild-type intestine than in TRDKO intestine (Fig. 3A, C). The top 10 most significantly enriched KEGG pathways were also different between wild-type and TRDKO intestine. Of note, glutathione metabolism pathways, involved in cell cycle and apoptosis, and pyrimidine metabolism pathways, important in stem cell function and DNA replication, were enriched in wild-type intestine (Fig. 3B), while the pathway for extracellular matrix (ECM)-receptor interaction, likely involved in epithelial folding, and PPAR pathway, important for lipid metabolism, were enriched in TRDKO intestine (Fig. 3D).
GO terms related to development, cell cycle, apoptosis, and stem cell are expected to be critical for intestinal remodeling during metamorphosis. Consistently, we found many of them enriched among DEGs in wild-type intestine, but fewer in TRDKO intestine (Fig. 4A and Supplementary Tables S4 and S5). Interestingly, most of the GO terms enriched in TRDKO intestine were also enriched in wild-type intestine. Taking cell cycle-related GO terms as an example, 43 of the 48 GO terms enriched in TRDKO intestine were also enriched in wild-type intestine (Fig. 4B). We identified T3-regulated genes in these 43 GO terms and found 191 and 76 DEGs in wild-type and TRDKO intestine, respectively.
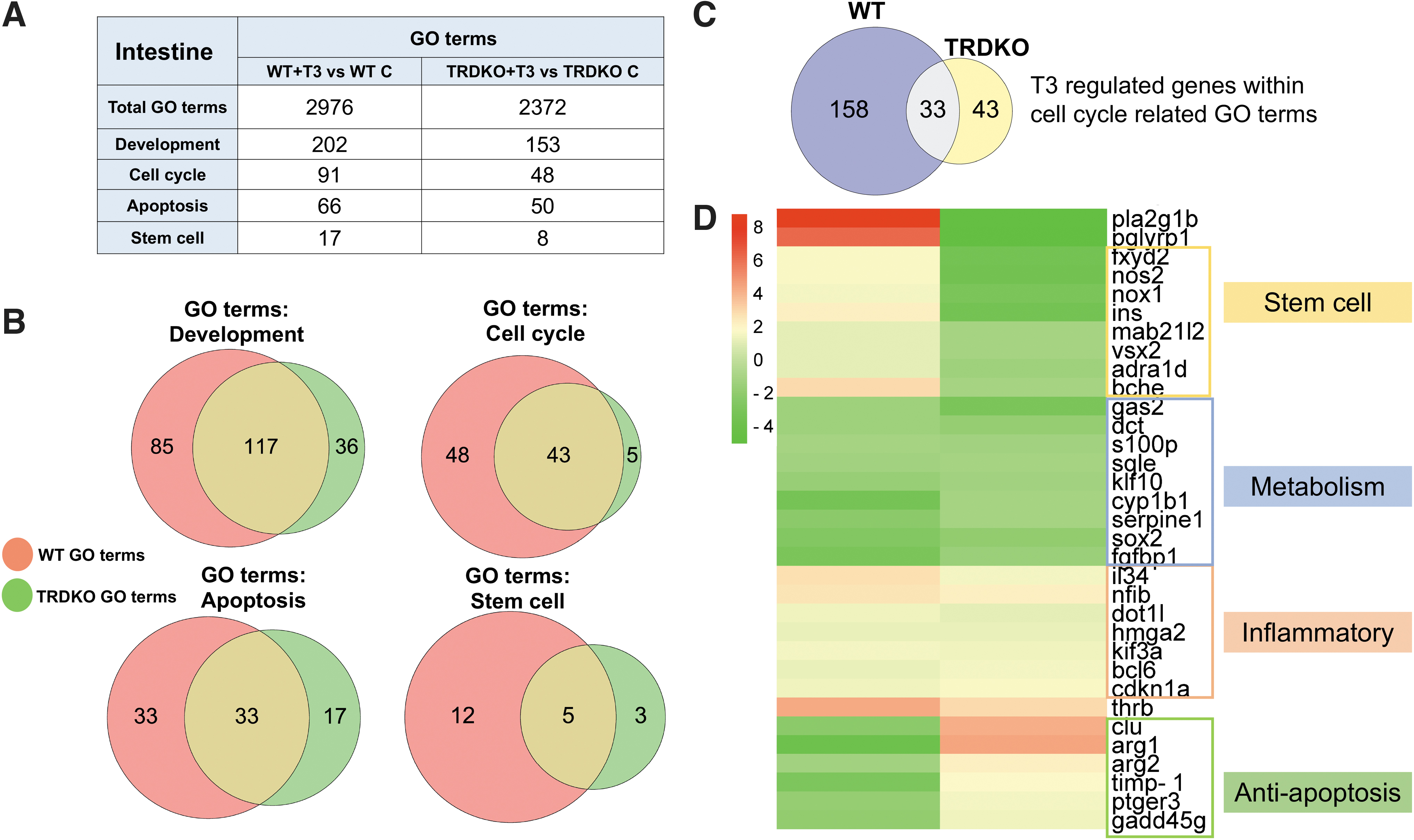
TRDKO reduced the number of enriched GO terms related to major intestinal remodeling processes: apoptosis, cell proliferation/cell cycle, development, and stem cell. (
Among them, only 33 DEGs were common between wild-type and TRDKO intestine (Fig. 4C). Furthermore, clustered heat map comparison of gene regulation patterns showed that most of these 33 genes were regulated in opposite manner by T3 in wild-type and TRDKO tadpoles, and they could be grouped into two functional groups: stem cells and anti-apoptosis (Fig. 4D). Their opposite regulation patterns are consistent with T3-induced stem cell formation and larval epithelial cell death during intestine remodeling in wild-type but not TRDKO tadpoles. These findings suggest that T3 affects mostly distinct biological processes in the presence and absence of TRs, but there are common effects on metabolism and inflammatory response commonly in wild-type and TRDKO intestine.
Effects of TRDKO on GO terms and KEGG pathways regulated by T3 in the tail
Similar analyses on the 1848 and 774 T3-regulated DEGs in wild-type and TRDKO tail showed that many development-related GO terms were enriched in wild-type tail (Fig. 5A and Supplementary Table S6), while lipid transport and metabolism GO terms were highly enriched in TRDKO tail (Fig. 5C and Supplementary Table S7). Among KEGG pathways, cell cycle, cellular senescence, and p53 signaling pathways were enriched in wild-type tail, while lipid metabolism pathways were enriched in TRDKO tail (Fig. 5B, D and Supplementary Tables S6 and S7). Notably, a higher fraction of metabolism-related pathways (17/27) were enriched in TRDKO tail than in wild-type tail (13/40), suggesting that many more of DEGs were involved in metabolism in TRDKO tail compared with wild-type tail.

GO and KEGG analyses reveal that distinct GO terms/KEGG pathways are regulated by T3 in the tail of wild-type (
Organ-specific nongenomic actions of T3
To investigate if the nongenomic effects were organ-dependent, we compared the T3-regulated DEGs in TRDKO tail and intestine. We found that 598 DEGs were unique to the tail and 693 DEGs were unique to the intestine, with only 176 common DEGs, much less than the common DEGs between the wild-type intestine and tail (Fig. 6A). Heat map of the 176 common DEGs showed that many were regulated by T3 in opposite manner in the 2 organs (i.e., upregulated [red] by T3 in one organ but downregulated [green] in the other) (Fig. 6B). In addition, of these 176 DEGs, 55 DEGs were upregulated and 97 DEGs downregulated in both organs, while 24 genes had opposite regulation patterns (Fig. 6C–F, C′–F′). Thus, TR-independent nongenomic effects of T3 on gene expression are mostly organ-specific.
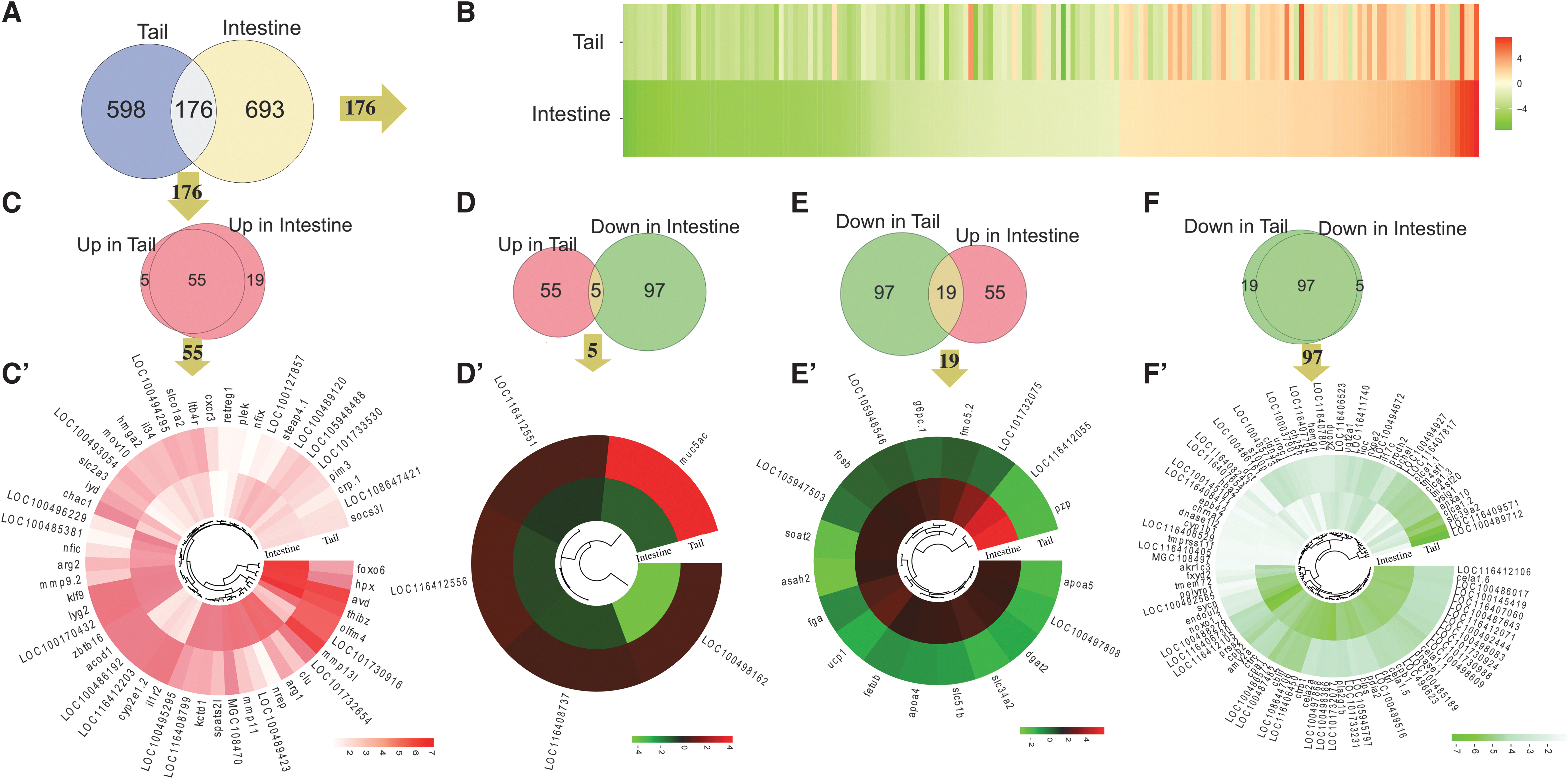
The nongenomic effects of T3 affect distinct genes in the tail and intestine. (
To identify common nongenomic effects of T3, we performed GO/KEGG analysis on the 152 DEGs similarly regulated by T3 in TRDKO intestine and tail and found that the enriched GO terms and pathways were mostly related to metabolism and cytokine–cytokine receptor interaction (Supplementary Fig. S4).
We next analyzed KEGG pathways enriched among 256 upregulated and 518 downregulated DEGs in TRDKO tail or 338 upregulated and 531 downregulated DEGs in TRDKO intestine (Supplementary Fig. S5). We combined the pathways enriched among the upregulated and downregulated DEGs for each organ and compared them. We found that 13 pathways were enriched in both organs, including “PPAR signaling pathway” and “Cytokine–cytokine receptor interaction” (Supplementary Fig. S5). Intriguingly, more than half of pathways were enriched from DEGs regulated by T3 in opposite patterns in the tail and intestine, such as “PPAR signaling pathway” (Supplementary Fig. S5).
Further analysis showed that the pathways enriched among DEGs regulated by T3 in either the same or opposite manner in the two organs were mainly related to different metabolic processes with exception of the pathway for “Cytokine–cytokine receptor interaction” and “FoxO signaling pathway,” both of which were enriched among DEGs upregulated in both organs. The regulation of genes such as il34/il1r2 in “Cytokine–cytokine receptor interaction pathway” and cdkn2b and cdkn2d/cdkn1a in “FoxO signaling pathway” suggests that both pathways are likely involved immune regulation in both organs (Supplementary Fig. S6). 32 –35
Given that integrin αvβ3 is one of the key targets of T4 for its nongenomic actions and “ECM-receptor interaction pathway” was enriched in TRDKO intestine (Supplementary Table S5), we compared the DEGs in this pathway between TRDKO tail and intestine. The results showed that all DEGs in the pathway were downregulated, such as laminin, collagen, and α8, in TRDKO intestine (Fig. 7A). However, in the tail, laminin was upregulated, while other DEGs were distinct from the DEGs in the intestine (Fig. 7B). Additionally, majority of the DEGs in “PPAR pathway,” enriched in both TRDKO intestine and tail (Figs. 3D and 5D), were distinct in the two organs (Supplementary Fig. S7A, B). Thus, nongenomic effects of T3 affect different genes in the intestine and tail.
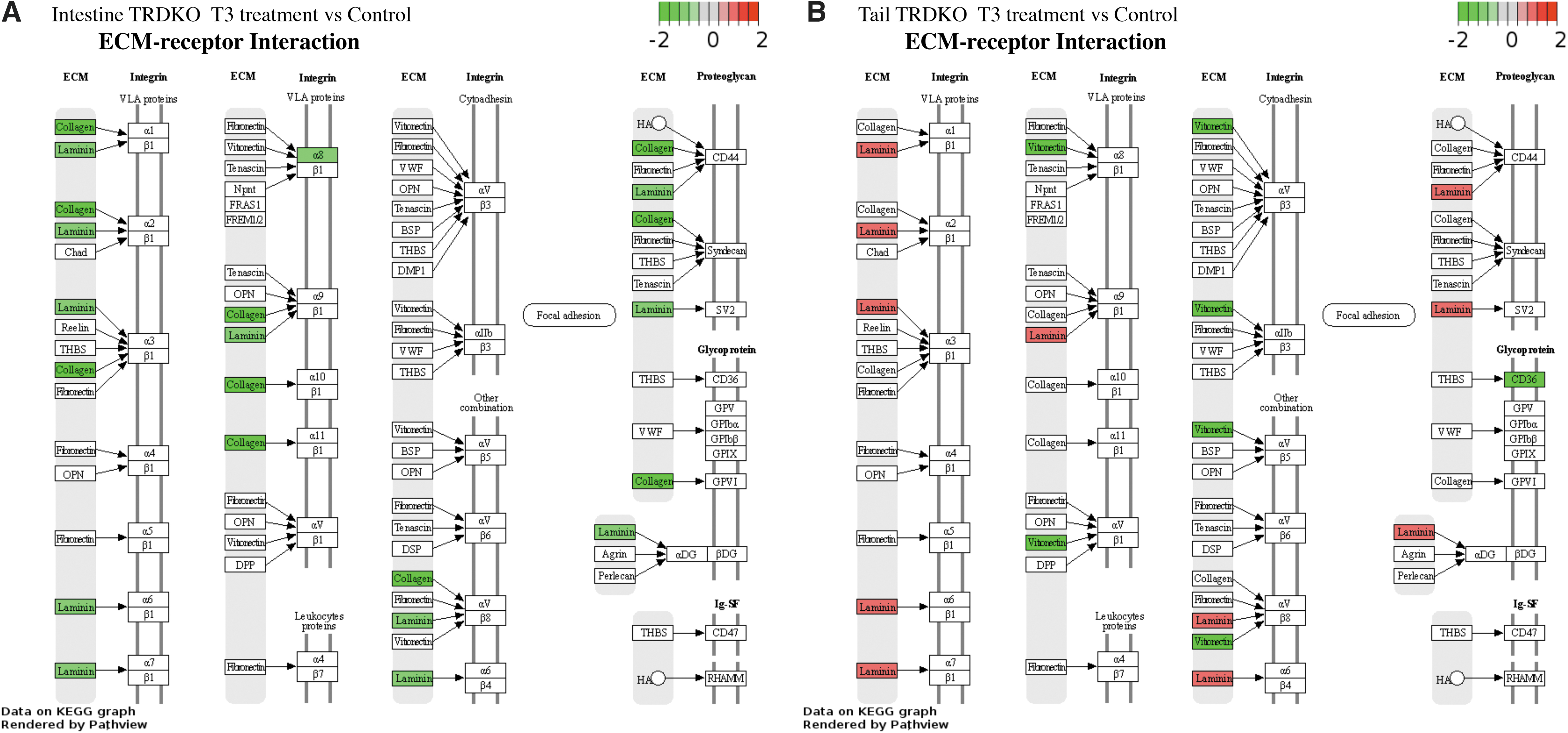
KEGG pathway for ECM-receptor interaction is enriched among T3-regulated genes in both the tail (
TRDKO derepresses T3-regulated programs
Earlier studies showed that TRDKO led to precocious premetamorphic development, enabling tadpoles to enter metamorphosis prematurely, likely due to derepression of target genes. 20,36 To investigate if TRDKO derepresses developmental programs in the tail and intestine, we compared gene expression between wild-type and TRDKO tail and intestine at stage 54 without T3 treatment. We obtained that 2682 (1646 upregulated, 1036 downregulated) DEGs in the tail (Supplementary Table S8A) and 2405 (1295 upregulated, 1110 downregulated) DEGs in the intestine (Supplementary Table S8D).
GO/KEGG analyses revealed that “cell cycle” and “p53 signaling pathway” were enriched among 2682 tail DEGs (Supplementary Table S8B, C and Supplementary Fig. S8A, B), while “development process” and “ECM-receptor interaction” were enriched among intestinal 2405 DEGs (Supplementary Table S8E, F and Supplementary Fig. S8C, D). These results suggest that TRDKO also causes derepression of T3-inducible genes and T3-regulated programs that are likely involved in the metamorphosis of the tail and intestine, although to an extent not sufficient to cause detectable precocious metamorphic changes in these organs.
Discussion
Thyroid hormone signals predominantly via TR to affect biological processes, such as metabolism, cell proliferation and death, and organ development. 1,3,37 Extensive studies in mice have shown that TRα knockout, TRβ knockout, and TRDKO are associated with different phenotypes, suggesting that different TR subtypes may have distant roles for development and organ physiology. 38,39 Similarly, studies in frog have shown that TRα and TRβ knockout have different phenotypes. 3 Interestingly, TRs appear to be not required for the development of many, if not all, adult organs in both frogs and mammals, although tadpoles lacking TRs die at the climax of metamorphosis. 3,9,20,38,40
This offers an interesting and important opportunity to study potential roles of T3 nongenomic actions during development. Our findings here indicate that TR-mediated genomic and TR-independent nongenomic effects of T3 affect the expression of mostly distinct genes in premetamorphic tadpoles. Furthermore, the nongenomic effects of T3 are largely organ-specific with metabolism as a major common target in both the intestine and tail.
Unliganded TRs are critical to maintain normal development and prevent premature metamorphosis. 3,9,12 Consistently, our RNA-seq analysis identified many more T3-regulated gene in wild-type tadpoles than those in TRDKO tadpoles. Furthermore, majorities of the genes regulated by T3 in both wild-type and TRDKO organs exhibited opposite T3 regulation patterns. For example, dio3 (deiodinase type 3), responsible for T3 degradation and required for proper development 41 and upregulated by T3 treatment in mouse cerebrocortical cells, 42 is similarly regulated in both wild-type intestine and tail after T3 treatment of premetamorphic tadpoles. However, the expression was repressed in the intestine by T3 treatment of TRDKO (Supplementary Table S3). Thus, TRs are critical to mediate T3 regulation of genes important for metamorphosis, consistent with early transgenic expression of dominant-negative TRs that can block metamorphosis. 43 –45
TR is necessary for intestinal larval epithelial cell death and de novo formation of adult stem cells. 9,12,13,15 Consistently, GO terms and KEGG pathways related to development, apoptosis, and stem cell were highly enriched among DEGs in wild-type intestine while TRDKO dramatically reduced their enrichment. Notable examples include signaling pathways such as Hedgehog, Notch, Wnt, and TGF-beta, which are important for regulation of stem cell and organ development and include DEGs such as bmp4 and wnt5a, important for cell proliferation and differentiation during intestinal remodeling. 46 –52
In this regard, it is worth noting that such pathways are also regulated by T3 to affect stem cells, cell proliferation, and differentiation in the mouse intestine, 53 –56 highlighting a conservation of T3 function in vertebrate intestine. In addition, KEGG pathways “glutathione metabolism” and “pyrimidine metabolism,” reported to be important for cell cycle and apoptosis, stem cell and DNA replication, 57 –59 were enriched in wild-type intestine, again consistent with their likely involvement in intestinal metamorphosis. However, “ECM-receptor interaction” and “lipid metabolism” pathways were enriched in TRDKO intestine, which agrees with our previous findings that TRDKO intestine has premature intestinal epithelial folding and muscle development but lacks larval epithelial cell apoptosis and stem cell development/proliferation. 9
T3 and T4 can induce rapid nongenomic actions through plasma membrane and mitochondrial binding sites. 26,60,61 For instance, T4 (and to a lesser extend T3) binds to integrin αvβ3 to activate MAPK signaling cascade, while T3 is the principal activator of the PI3K/Akt pathway, and such effects in turn affect cell proliferation, angiogenesis, and cell survival. 61 –64 Our analysis for the first time revealed nongenomic effects of T3 on global gene expression in developing animals. We found that GO terms and pathways related to metabolism were highly enriched among DEGs in the absence of TR in both the intestine and tail. T3 is a well-known regulator of metabolism in mammals, including amino acid catabolism and lipid metabolism. 5,26,61,65 –67 While it is generally believed that these effects are mediated by TR-dependent gene regulation, our findings here suggest that TR-independent nongenomic effects of T3 may also play a, if not dominant, role in regulating tissue metabolism.
For example, GO terms and pathways related to metabolism (e.g., α-linolenic acid [18:3n-3]) and immune response (e.g., Cytokine–cytokine receptor interaction) were highly enriched in both TRDKO tail and intestine, suggesting that nongenomic actions of T3 may affect immune response and metabolism of longer chain polyunsaturates, which are required for normal physiology such as brain function. 68,69 Similarly, melanogenesis pathway was enriched among downregulated DEGs in TRDKO intestine and tail, consistent with reports that T3 may inhibit the production of melanin. 70,71 However, despite such common effects in the intestine and tail, most of genes regulated by T3 in the absence of TRs and the corresponding enriched GO terms/pathways are different between the tail and intestine, suggesting that nongenomic actions of T3 are largely dependent on tissues/organs and/or cellular microenvironment. 2,63,67
Conclusions
In conclusion, we have discovered that mostly distinct genes are regulated by T3 via TR-independent nongenomic and TR-dependent processes. While it is possible that RNA-seq of wild-type tissues may miss some genes regulated by T3 via TRs if the genes are regulated by T3 in opposite manner though TR-dependent and TR-independent processes, the enrichment of GO terms and pathways among the DEGs in wild-type tadpoles is consistent with T3-dependent metamorphic changes. However, the most noticeable nongenomic effects of T3 in the absence of TR in both organs are the regulation of GO terms and pathways involved in metabolism. Given the well-known effects of T3 on tissue metabolisms in mammals, it is templating to suggest that perhaps some of the metabolic effects of T3 in mammals are mediated by TR-independent nongenomic processes.
Footnotes
Acknowledgments
We thank Nga Luu for laboratory management and animal care staff for frog husbandry.
Authors' Contributions
S.W., Y.S., Y.T., and Y.-B.S. designed the research plan. S.W., Y.S., and Y.T. carried out the research and data analyses. All authors participated in the article preparation and approved the final version of the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Intramural Research Program of the Eunice Kennedy Shriver National Institute of Child Health and Human Development (NICHD), National Institutes of Health (NIH). Yuta Tanizaki and Yuki Shibata were supported in part by the Japan Society for the Promotion of Science Research Fellowship for Japanese Biomedical and Behavioral Researchers at the NIH. Yuta Tanizaki was also supported in part by an FY22 NICHD Early Career Award.
Supplementary Material
Supplementary Data
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Supplementary Figure S7
Supplementary Figure S8
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
Supplementary Table S5
Supplementary Table S6
Supplementary Table S7
Supplementary Table S8