
Abstract
Background:
The DIO2 Thr92Ala polymorphism (rs225014), which occurs in about 15–30% of Caucasian people, determines a less efficient type 2 deiodinase (D2) enzyme. The aim of this study was to determine the impact of DIO2 Thr92Ala polymorphism on the serum thyrotropin (TSH) levels in thyroidectomized patients with hypothyroidism and to evaluate whether TSH levels and aging could be related, at pituitary level, to D2 activity.
Methods:
This prospective study was performed on 145 thyroid cancer patients, treated with total thyroidectomy, and undergoing radioiodine treatment after 3 weeks of levothyroxine (LT4) withdrawal. A mouse model has been used to determine D2 protein and mRNA levels in pituitary during aging.
Results:
Genetic analysis identified DIO2 Thr92Ala polymorphism in 56% of participants: 64/145 (44%) patients were homozygous wild type (WT) (Thr/Thr), 64 (44%) heterozygous (Thr/Ala), and 17 (12%) homozygous mutant (Ala/Ala). A significant negative relationship was observed between aging and the rise in serum TSH levels during LT4 withdrawal. However, this negative correlation found in WT was reduced in heterozygous and lost in mutant homozygous patients (Thr/Thr r = −0.45, p = 0.0002, 95% confidence interval [CI] −0.63 to −0.23; Ala/Thr r = −0.39, p = 0.0012, CI −0.60 to −0.67; and Ala/Ala r = −0.30, p = 0.2347; CI −0.70 to 0.20). Accordingly, when we compared the TSH measured in each patient to its theoretical value predicted from age, the TSH did not reach its putative target in 47% of WT patients, in 70% of Ala/Thr, and 76% of Ala/Ala carrying patients (p = 0.0036). This difference was lost in individuals older than 60 years, suggesting a decline of D2 associated with aging. The hypothesis that the pituitary D2 decreases with age was confirmed by the evidence that D2 mRNA and protein levels were lower in pituitary from old versus young mice.
Conclusion:
An age-related decline in TSH production in response to hypothyroidism was correlated with decreased D2 levels in pituitary. The presence of DIO2 homozygous Ala/Ala polymorphism was associated with a reduced level of TSH secretion in response to hypothyroidism, indicating a decreased pituitary sensitivity to serum thyroxine variation (Institutional Research Ethics board approval number no. 433/21).
Introduction
Thyroid hormone (TH) replacement with levothyroxine (LT4) is the standard treatment for hypothyroid patients. 1 Although the thyroid gland secretes both thyroxine (T4) and triiodothyronine (T3), in euthyroid individuals, deiodinase D2 accounts for about 70% of circulating T3. 2 In athyreotic patients, the T3 production depends exclusively from the peripheral conversion of T4 to T3 mediated by type 1 and type 2 deiodinases (DIO, D1 and D2). At physiological concentrations, the Km of D2 for T4 (1–4 nM) is about a 1000-fold lower compared with D1 (1–10 μM), which makes T4 a much better substrate for D2 than for D1. 3 Several polymorphisms have been identified in the DIO genes, which could potentially affect TH plasma level. 4 –6
The DIO2 Thr92Ala polymorphism occurs in about 15–30% of Caucasian people and results in a decreased enzymatic activity. 5,7 It is associated with metabolic diseases, mental disorders an increased risk of developing Alzheimer disease, although it is associated with serum TH levels in the reference range. 5,7 –12 This evidence, together with the observation that about 15% of patients treated with LT4 have persistent complaints of hypothyroidism, despite reaching normal thyrotropin (TSH) levels, raises the questions of whether TSH represents a reliable biochemical marker of the thyroid status for all patients. 13
It is well known that D2 is also essential for the pituitary feedback regulation of TSH by T4. The acute suppression of TSH secretion by T4 does not require the formation of circulating T3, but depends on locally D2-generated T3 production. In a mouse model of Dio2 global knockout (D2KO), the loss of D2 activity resulted in a phenotype of pituitary resistance to T4, and in hypothyroid D2KO mice, serum TSH levels were not suppressed by T4 administration, but only by T3. 14 Furthermore, mice with a reduced D2 enzymatic activity were less sensitive to serum T4 level variation. Interestingly, in primary mouse thyrotropes expressing the D2 Thr92Ala enzyme, the suppression of TSH secretion by T4 was reduced versus pituitary cells carrying a wild-type (WT) enzyme. 5
Therefore, even if D2 Thr92Ala enzymatic activity is estimated to be reduced by only 20%, this reduction is sufficient to induce significant changes in TH homeostasis. 5,15 In humans, little is known about the in vivo effects of DIO2 Thr92Ala polymorphism on T4 pituitary sensitivity and TSH secretion. In this setting, defining whether DIO2 Thr92Ala polymorphism affects serum TSH levels and its response to serum T4 variations represents an important issue. The aim of this study was to evaluate in hypothyroid patients the impact of DIO2 Thr92Ala polymorphism on pituitary TSH secretion, and to determine whether TSH levels and aging could be related to D2 activity.
Materials and Methods
Protocol design
We prospectively analyzed 167 differentiated thyroid cancer patients, who have been treated with total thyroidectomy and who were referred for 131I radioiodine treatment (RAI) to the Nuclear Medicine Department of University Hospital “Federico II” of Naples between January 2021 and July 2022. Medical records were reviewed to obtain the following information: (1) demographic data (gender and age at RAI treatment), (2) thyroid cancer type, and (3) dates of LT4 withdrawal and of blood sampling for determination of TSH and free thyroxine (fT4)-free triiodothyronine (fT3) levels three weeks after the LT4 withdrawal (the patients were not on any transitional T3 treatment preceding the LT4 withdrawal).
The exclusion criteria were as follows: (1) treatment with drugs interfering with TH metabolism (e.g., amiodarone, propranolol, corticosteroid) and (2) comorbidity interfering with TH metabolism (chronic renal failure, liver failure, and heart failure class NYHA III/IV). The study was approved by the Ethical Committees of our institution (Institutional Research Ethics board approval number no. 433/21), and all participants provided written informed consent. The research was completed in accordance with the Declaration of Helsinki as revised in 2013.
TSH and TH assessments
Serum levels of fT3 and T4 and TSH were measured using an automated chemiluminescence assay system (Immulite 2000; DPC, Los Angeles, CA).
DIO2 gene analysis
Genomic DNA was extracted from blood with the QIAamp DNA Micro Kit (Qiagen, Milan, Italy) according to the kit's instructions. DNA concentration was assessed with a fluorometer. Primers have been designed to cover the second DIO2 exon using Primer 3 (version 0.4.0). Annealing temperature was 60°C and 51°C (35 cycles). Polymerase chain reaction (PCR) products were digested with the CviQI restriction enzyme (New England Biolabs, Milan, Italy) at 37°C overnight. Fragments were run in a 2% agarose gel stained with ethidium bromide. Genotype determination on the gel is based on the following pattern: WT individuals show three fragments at 105, 57, and 30 bp; heterozygous had four fragments at 105, 87, 57, and 30 bp; and mutant homozygous show two fragments at 105 and 87 bp.
Mouse studies
C57BL/6J mice were obtained from Jackson Laboratory (Bar Harbor, ME). Mice were fed with standard rodent chow and water ad libitum and maintained under a 12-hour light/12-hour dark cycle. Animals were handled according to national and European Community guidelines, and protocols were approved by the Animal Research Committee of the University of Naples “Federico II” (MIUR, Approval Code: 354/2019-PR).
Pituitary gland was collected and total RNA was extracted using TRIzol (Invitrogen), according to the manufacturer's instructions. Two hundred fifty nanograms of total RNA was reverse transcribed into cDNA using LunaScript RT SuperMix Kit (New England BioLabs). The cDNAs were amplified by PCR in an i-Cycler (Bio-Rad) using iTaq Universal SYBR Green Supermix (Bio-Rad). Sequences of primers used are as follows: mDio2s_new CTCTTCCTGGCGCTCTATGA, mDio2r_new CACACTGGAATTGGGAGCAT, mTSHb R AAAAGCACGGAGAGGAGGAC, mTSHb for GTGGGTGGAGAAGAGTGAGC, mCyAS CGCCACTGTCGCTTTTCG, and mCyAR AACTTTGTCTGCAAACAGCTC). Quantitative real-time PCR was performed as described previously using the delta cycle threshold method, with cyclophilin A used as the housekeeping gene for normalization. 16
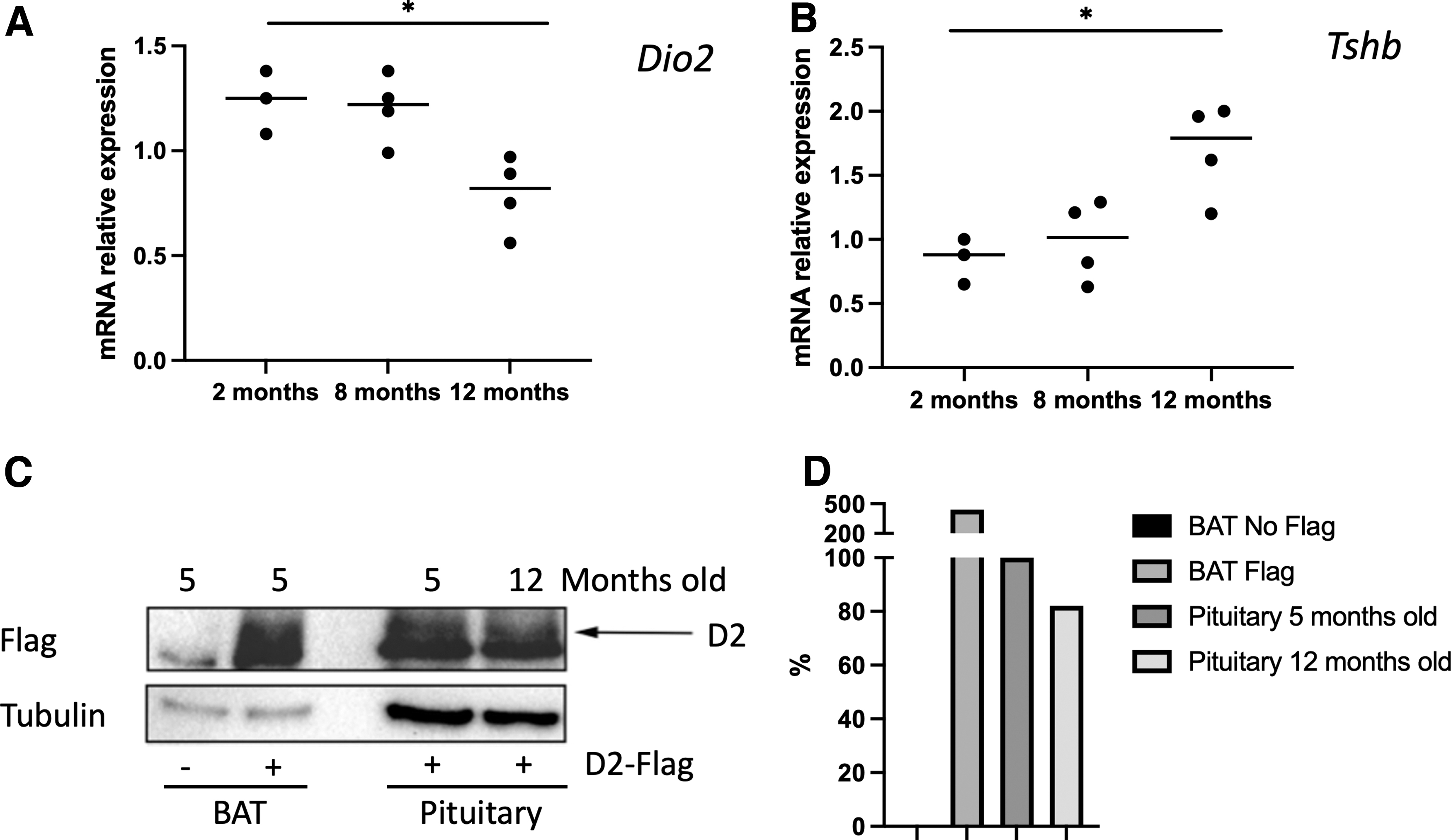
For Western blot analysis, we collected pituitaries from D2 3xFlag mouse 5 and from a D2 WT mouse at the age of 5 and 12 months (n = 3). Three pituitaries from each different mouse model and age were pooled together and mechanically dissociated in 100 μL 1 × Laemmli Sample Buffer. Cold stimulated brown adipose tissue from D2 3xFlag mouse 5 and from a D2 WT mouse have been utilized as positive and negative control, respectively. Fifteen microliters (about 30 μg) of each sample were run on a 10% sodium dodecyl sulphate-polyacrilamide gel electroforesis gel and transferred on polyvinylidene fluoride membrane (Amersham Hybond-P). The membrane was then blocked with 5% non-fat dry milk in phosphate-buffered saline, probed with anti-FLAG (1:1000; Sigma), washed and incubated with horseradish peroxidase-conjugated donkey anti-mouse IgG secondary antibody (1:3000), and detected by chemiluminescence (Biorad). After extensive washing, the membrane was incubated with anti-Tubulin-specific antibodies (1:500; Abcam) as loading control.
Statistical analysis
Statistical tests performed were frequency analysis, the Chi Square test for the evaluation of the significance between the nominal variables, and Mann–Whitney U test and Student's t test for unpaired data to compare values and average of continuous variables. Univariate analysis of variance was used for comparison between groups. Spearman correlation analysis was used to identify associations between the variables of interest. A p-value <0.05 was assumed as statistical significance. The statistical analysis was performed using the GraphPad software 8.
To determine the predicted TSH target for age, we extrapolated the equation of line for Thr/Thr group: Y = a * X + b, where Y is the expected TSH, and X is the age (eTSH = a * Age + b). 17,18 We applied this formula to calculate the predicted TSH for each single patient on the basis of age.
Results
A total of 145 out of 167 screened patients satisfied the inclusion criteria (Supplementary Fig. S1) and form the basis of the study: 30.3% were males, 99% were white/Caucasian, and the median age was 46 years at the time of the 131I treatment (range: 14–82 years). For 81% of participants, this was the first treatment course of RAI, and for the remaining 19%, it was the second treatment course (Table 1). TSH levels were measured three weeks after LT4 withdrawal before RAI treatment. At that time, all patients were severely hypothyroid (all fT4 and fT3 levels were below the reference ranges).
Patient Characteristics
TSH, thyrotropin.
The analysis of the exon 2 of the DIO2 gene showed that 64/145 (44%) patients were homozygous WT (Thr/Thr), 64/145 (44%) heterozygous (Thr/Ala), and 17/145 (12%) were mutant homozygous (Ala/Ala; Table 2). The three groups did not statistically differ for sex, age, TSH levels, stage of the disease, time since thyroidectomy, and number of RAI treatments (Table 1).
Genotype Distribution
We, first, confirmed that TSH level after LT4 withdrawal is correlated with the age of the individuals: TSH level increases to a lesser extent in older than in younger patients (r = −0.39, p < 0.001, 95% confidence interval [CI] −0.52 to −0.24; Fig. 1a). Next, we evaluated whether the TSH rise was correlated with deiodinase gene polymorphism. At univariate analysis, age was still negatively related to serum TSH levels in Thr/Thr (r = −0.45, p = 0.0002, CI −0.63 to −0.23) and Ala/Thr (r = −0.39, p = 0.0012, CI −0.60 to −0.67) groups, but not in the Ala/Ala group (r = −0.30, p = 0.2347, CI −0.70 to 0.20; Fig. 2b). The lowest slope is associated with the mutant Ala/Ala phenotype, while the highest with the WT phenotype (Fig. 1b).
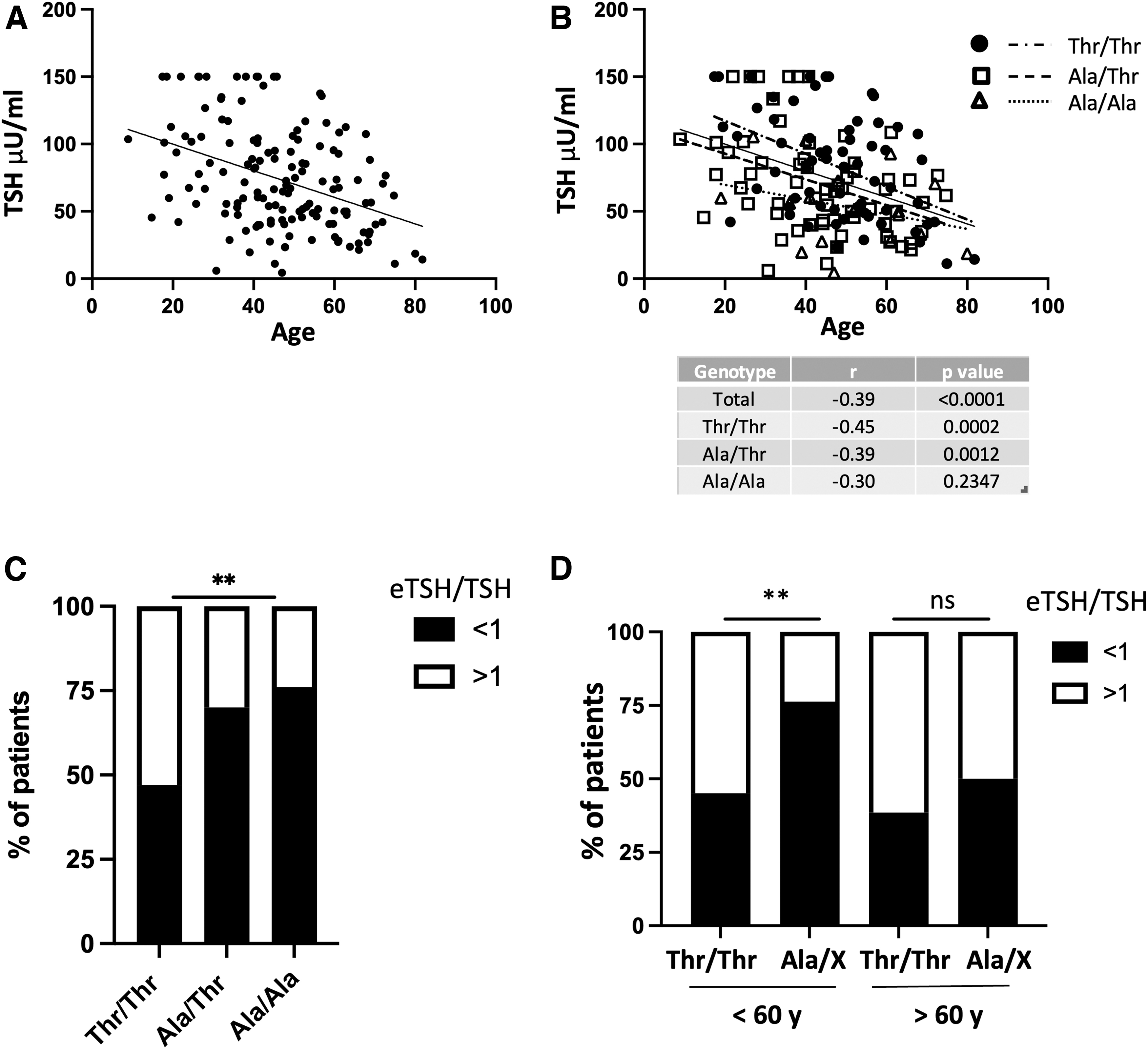
Correlation between age and TSH levels after three weeks of LT4 withdrawal (
(
To assess the impact of the presence of Dio2 Thr92Ala polymorphism on TSH levels after LT4 withdrawal, we determined the percentage of individuals carrying the polymorphism that did not reach the predicted TSH target for age, and this occurred in 47% of patients of the WT Thr/Thr group and in 70% of the Ala/Thr and 76% of Ala/Ala patients (p = 0.0036) (Fig. 1c). Our data indicate that there is a correlation between the magnitude of rise in TSH level and D2 activity.
We then hypothesize that reduced TSH response observed with aging could depend on an impaired D2 activity. Interestingly, when our population was stratified into two groups according to the age, using a cutoff of 60 years, we found that in patients younger than 60 years, the percentage of patients who did not reach the predicted TSH target was higher in Ala-carrying patients versus WT patients, but such a difference was lost in the older population, and this might be related to a decline of D2 activity associated with aging (p = 0.01 and p = 1.000, respectively) (Fig. 1d).
To confirm this hypothesis, we measured Dio2 expression levels in mouse pituitary glands of mice of different ages (5 and 12 months, respectively). We found that Dio2 mRNA and protein expression levels in mouse pituitary decrease with aging, whereas TSH beta subunit mRNA expression increases, reinforcing our hypothesis (Fig. 2).
Discussion
In this study, we showed that in hypothyroid patients carrying DIO2 Thr92Ala polymorphism, the increase in serum TSH levels in response to hypothyroidism was lower in magnitude versus WT patients. This suggests that, in humans, DIO2 Thr92Ala polymorphism is associated with a decreased pituitary sensitivity to T4. This is in accordance with the phenotype observed in both global and thyrotroph-specific D2KO mice models, which have a blunted response to T4 treatment in terms of reduction in TSH levels. 14,19,20 In human, this is in line with a previous study showing that thyroidectomized patients carrying D2 Thr92Ala polymorphism need higher amount of LT4 to fully suppress TSH, 21 although this was not confirmed in a later study. 22
Herein, we observed that, in agreement with previous studies, the raise in TSH upon LT4 withdrawal is more significant in younger than in older individuals. 23,24 An interesting finding of our study was that the correlation TSH/age was lost in Ala/Ala group. In a graph plotting TSH level versus age, the slope of the curve negatively correlates with the number of the copy of polymorphic DIO2 allele. This suggests that the correlation TSH/age might depend on D2 activity, which in turn decreases with age.
This supposed age-dependent decrease in D2 activity could explain why the increment of TSH is reduced in aging individuals. The condition of aging individuals is partially reproduced in young individuals with the genetic D2 deficiency due to DIO2 polymorphism. This led us to speculate that there is a minimal D2 activity, which confers adequate rise of TSH in response to reduced T4 levels. This rise is reduced in WT individuals with age, and in younger patients, it is affected by the genetic DIO2 polymorphism.
Importantly, the decreased response of TSH to serum T4 reduction in individuals with the D2 Thr92Ala polymorphism could simply be a slower response in timing than in WT individuals, with a longer time needed by the pituitary to reach a definitive stable TSH target. In this scenario, a subclinical hypothyroidism or hyperthyroidism in these subgroups of individuals could represent a more challenging condition than overt clinical hypothyroidism or hyperthyroidism. However, this hypothesis could potentially be investigated by evaluating TSH rise timing in populations of patients with spontaneous hypothyroidism (i.e., in untreated primary, spontaneous autoimmune hypothyroidism cohort references). 17 Notably, Torlontano et al found that homozygous differentiated thyroid cancer carriers of the DIO2 Thr92Ala allele need higher T4 dosages, only in the near-suppressed TSH group, but not in the suppressed group. 21
Of note, deiodinase enzymes are involved in Hypothalamus-Pituitary-Thyroid (HPT) axis maturation. 25 In pituitary-D2KO mice, the absence of D2 activity was associated with a variation of TSH in a narrower range compared to WT mice, probably related to an altered HPT axis maturation due to the higher perinatal levels of T4. 19 We could suppose that a similar condition might be associated also with DIO2 Thr92Ala polymorphism in humans. Celi et al showed that in humans, DIO2 Thr92Ala polymorphism is associated with an altered HPT axis characterized by a decreased rate of acute TSH-stimulated T3 release from the thyroid. 8 Of note, D2 protein stability is regulated at multiple levels, including a tissue-dependent ubiquitination process. 26 Hypothalamus has an increased sensitivity to T4 due to a lesser D2 ubiquitination, compared to other tissues. This differential regulation is important to allow the thyrotropin-releasing hormone/TSH feedback. 26
Furthermore, some T3 generated by D2 in mediobasal hypothalamic tanycytes, also reaches the pituitary through the portal capillaries contributing to the pituitary T3 content. 27 Thus, the presence of a D2 enzyme with a decreased activity, as the polymorphic D2, might impact this process and thus be responsible, at least in part, for the decreased TSH secretion observed in hypothyroid patients carrying the D2 Thr92Ala polymorphism.
Thr92Ala DIO2 polymorphism was found associated with several pathological conditions as hypertension, insulin resistance, type 2 diabetes, and Alzheimer's disease, although these associations have not been observed in all populations studied. 7,5,11,28 –30 Importantly, the generation Ala-D2 transgenic mouse model showed that, despite the normal serum TSH levels, the Thr92-to-Ala substitution caused endoplasmic stress, the accumulation of Thr92Ala-D2 in the trans-Golgi, and the generation of less T3. 31 This was associated with a cognitive phenotype characterized by a decrease in physical activity, an increase in sleep, and a need for a longer time to memorize objects. 31 This study indicates that, even if Thr92Ala D2 enzymatic activity is estimated to be reduced by only 20–30%, this reduction is sufficient to induce significant changes in TH homeostasis under specific circumstances.
This study has some limitations and strengths. Regarding the individuals enrolled in the study, a limitation is that the evaluation was performed before RAI ablation. We cannot exclude the presence of residual thyroid tissue that produces some THs and could affect TSH levels. In addition, although the sample 145 was appropriate to ensure adequate statistical power, the number of Ala/Ala patients was relatively small. Indeed, when patients were stratified based on age, we had to consider only two groups (the Thr/Thr and Ala/X groups).
Regarding mouse studies, we showed that both D2 protein and mRNA expression were decreased, which together represent a strong indicator, but does not correspond to the D2 enzymatic activity. On the other hand, in our opinion, it has the following strengths: it is a prospective study involving a hypothyroid human model condition, which will be useful in the future to better characterize the correlation between the TSH rise and TH level decline in individuals carrying the DIO2 Thr92Ala polymorphism. Moreover, this study confirmed evidence derived from studies in mice.
Conclusions
During the past decade, serum TSH level has been questioned as a good marker for hypothyroidism, especially in athyreotic patients. About 15% of patients treated with LT4 have persistent clinical complaints, despite reaching biochemical euthyroidism. 32 –34 TH homeostasis is complex and depends on the integration of peripheral and central information, and TSH represents a sensor in this homeostatic system. In some circumstances, this control could be stressed and the adaptation could be suboptimal. In hypothyroid patients, the presence of DIO2 polymorphism could represent one of these difficult conditions. In summary, we found that D2 polymorphism is associated with a decreased pituitary sensitivity to T4. More studies are needed to confirm our findings and determine possible clinical implications.
Footnotes
Authors' Contributions
C.L.: conceptualization (lead) and writing—original draft review and editing (equal). M.A.S.: methodology (equal). R.A.: formal analysis (equal); F.V.: formal analysis (equal); T.P.: methodology (equal); V.G.: formal analysis (equal); C.B.: writing–review and editing (equal); G.T.: conceptualization (supporting); S.M.: writing—original draft (supporting); V.D.: formal analysis (supporting); E.M.: writing—review and editing (equal); M.K.: conceptualization (supporting); M.S.: conceptualization (equal) and writing—review and editing (equal); D.S.: conceptualization (equal) and writing—review and editing (equal).
Author Disclosure Statement
No competing financial interests exist.
Funding Information
PRIN 2017, Italian Ministry of University and Research, Rome, grant number Italian Ministry of University and Research, grant number 2017YTWKWH_004, Prof. Domenico Salvatore.
Supplementary Material
Supplementary Figure S1