
Abstract
Background:
Iodine is required for the synthesis of thyroid hormone (TH), but its natural availability is limited. Dehalogenase1 (Dehal1) recycles iodine from mono- and diiodotyrosines (MIT, DIT) to sustain TH synthesis when iodine supplies are scarce, but its role in the dynamics of storage and conservation of iodine is unknown.
Methods:
Dehal1-knockout (Dehal1KO) mice were generated by gene trapping. The timing of expression and distribution was investigated by X-Gal staining and immunofluorescence using recombinant Dehal1-beta-galactosidase protein produced in fetuses and adult mice. Adult Dehal1KO and wild-type (Wt) animals were fed normal and iodine-deficient diets for 1 month, and plasma, urine, and tissues were isolated for analyses. TH status was monitored, including thyroxine, triiodothyronine, MIT, DIT, and urinary iodine concentration (UIC) using a novel liquid chromatography with tandem mass spectrometry method and the Sandell–Kolthoff (S-K) technique throughout the experimental period.
Results:
Dehal1 is highly expressed in the thyroid and is also present in the kidneys, liver, and, unexpectedly, the choroid plexus. In vivo transcription of Dehal1 was induced by iodine deficiency only in the thyroid tissue. Under normal iodine intake, Dehal1KO mice were euthyroid, but they showed negative iodine balance due to a continuous loss of iodotyrosines in the urine. Counterintuitively, the UIC of Dehal1KO mice is twofold higher than that of Wt mice, indicating that S-K measures both inorganic and organic iodine. Under iodine restriction, Dehal1KO mice rapidly develop profound hypothyroidism, while Wt mice remain euthyroid, suggesting reduced retention of iodine in the thyroids of Dehal1KO mice. Urinary and plasma iodotyrosines were continually elevated throughout the life cycles of Dehal1KO mice, including the neonatal period, when pups were still euthyroid.
Conclusions:
Plasma and urine iodotyrosine elevation occurs in Dehal1-deficient mice throughout life. Therefore, measurement of iodotyrosines predicts an eventual iodine shortage and development of hypothyroidism in the preclinical phase. The prompt establishment of hypothyroidism upon the start of iodine restriction suggests that Dehal1KO mice have low iodine reserves in their thyroid glands, pointing to defective capacity for iodine storage.
Introduction
Iodine deficiency (ID) has severe adverse effects on human health, due to inadequate thyroid hormone (TH) production. 1 Nearly 2 billion individuals in the world are at risk for iodine deficiency, with pregnant women and children being the most vulnerable. 1,2 Despite its global prevalence and its dramatic consequences during pregnancy and childhood, early detection of incipient ID in individuals is not currently possible. For decades, international efforts were implemented to identify human populations with poor iodine intakes using the median urinary iodine concentration (UIC). However, UIC is not useful to diagnose ID individually since it depends on the recent iodine intake, which is typically variable, and not on iodine reserves. In fact, the need for more efficient markers of ID reflecting individual iodine status was recently emphasized. 3,4 Such biomarkers must relate to genetic factors modulating the efficiency of iodine handling pathways.
Dehalogenase1 (DEHAL1) also known as iodotyrosine deiodinase (IYD) is a key enzyme for iodine metabolism in the thyroid gland. It removes iodine from mono- and diiodotyrosines (MIT, DIT), making the former available for recycling and reuse in the thyroid gland to sustain thyroxine (T4) and triiodothyronine (T3) synthesis. 5,6 Loss-of-function DEHAL1 mutations were shown to cause severe goitrous hypothyroidism and mental retardation. 7 However, the time of onset of such hypothyroidism is unpredictable, 7,8 suggesting the influence of environmental conditions in the expression of the disorder. Unfortunately, neonatal screening programs do not detect hypothyroidism of this origin, but the reason is elusive. Therefore, there is a need for further investigation of the role of iodine recycling and the consequences of abnormal iodine conservation in humans throughout life. 9
To this aim, we generated a Dehal1-knockout (Dehal1KO) mouse using gene trap technology, allowing us to investigate the dynamic relationship between the external iodine supplies and the development of hypothyroidism under controlled iodine intake.
Methods
Generation of Dehal1-knockout mice
Mouse embryonic stem cells (ESCs) containing the Dehal1-knockout allele engineered through gene trap were obtained from the knockout mouse project repository. 10 Essentially, a targeting vector containing the bacterial β-galactosidase (lacZ) as reporter gene and neomycin (Neo) as selection gene were inserted between exons 1 and 2 of the wild-type (Wt) Dehal1 allele (Fig. 1A). Therefore, the Dehal1 promoter and exon 1 are conserved in the recombinant allele, while exons 2 to 6 are excluded from the messenger RNA (mRNA), which now contains lacZ, allowing the characterization of the Dehal1 expression pattern (Fig. 1A). The targeting vector was electroporated into C57BL/6J ES cells, and recombinant clones were selected on antibiotic and expanded. ESC clones verified for Dehal1 Flox/Wt were microinjected into blastocysts derived from albino C57BL/6J-Tyrc-Brd mice. Chimeric male mice with germline transmission were backcrossed to C57BL/6J mice, and the heterozygous mutants were crossbred to obtain homozygous mutant mice.
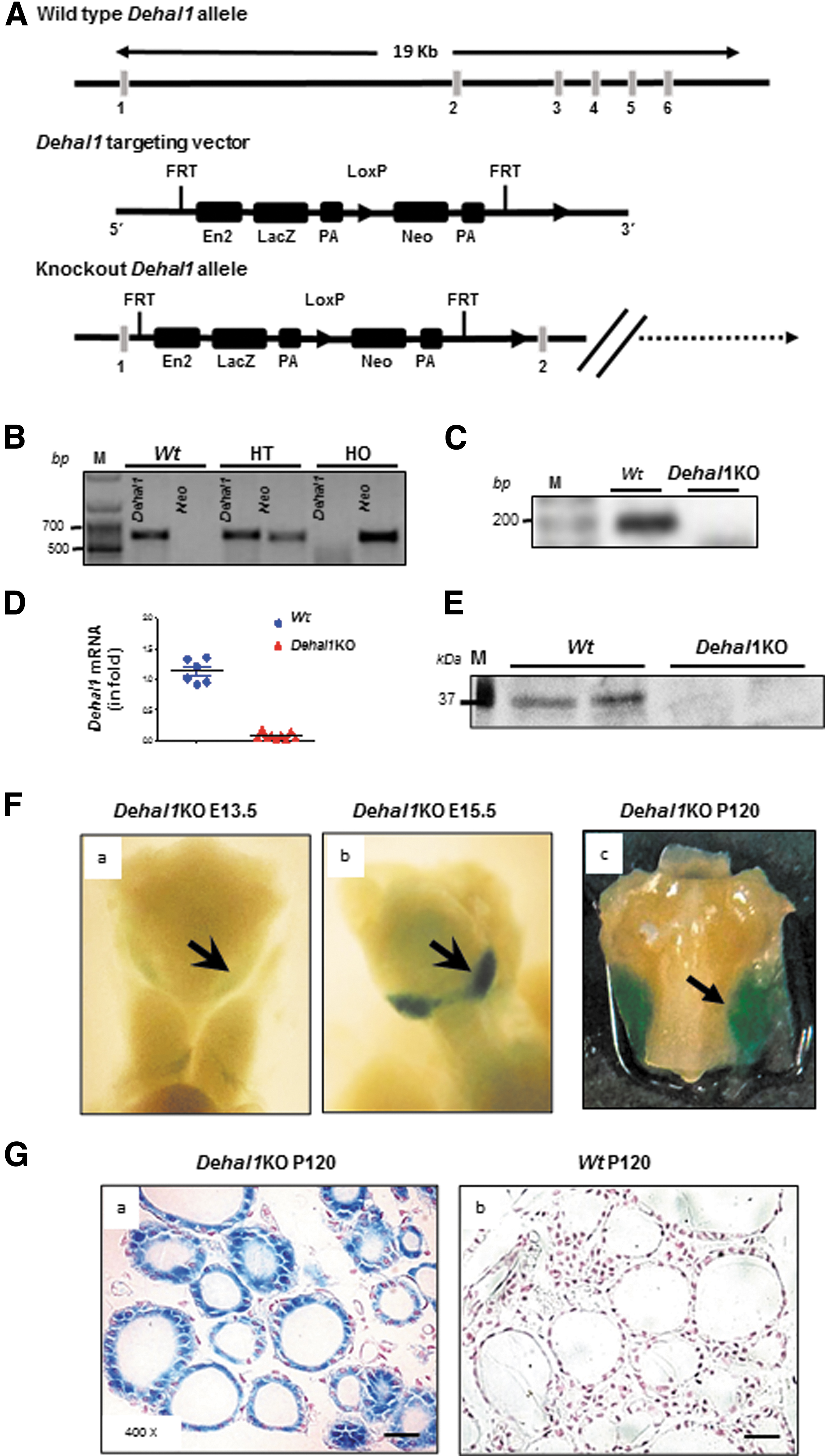
Dehal1KO mouse generation and characterization. (
Mouse genotyping
Mice were genotyped by touchdown-PCR using DNA extracted from tail. Primer sequences and conditions are available in Supplementary Table S1.
Quantitative reverse-transcription polymerase chain reaction
Mouse tissue RNA was isolated according to the manufacturer's protocol (Macherey-Nagel, Düren, Germany). One microgram of RNA was reverse transcribed with Transcriptor cDNA Synthesis Kit (Roche Diagnostics, Basel, Switzerland), and quantitative PCR was performed with SYBR Green dye (Quantabio, Beverly, MA) on a ABI Prism 7900HT Fast Real-Time PCR System (Applied Biosystems, Thermo Fisher Scientific, Waltham, MA) using the Delta Ct method. Dehal1 was amplified with specific primers (Sigma–Aldrich, St. Louis, MO) (Supplementary Table S1). Results were normalized with Gapdh and expressed relative to controls.
X-Gal staining
Whole-mount or tissue sections from Dehal1KO and Wt adult and fetal mice were X-Gal stained following published protocols. 11
Immunofluorescence
Β-Galactosidase immunofluorescence was performed after antigen retrieval using sodium citrate buffer at pH = 6 and blocked for 1 hour with 10% goat serum in phosphate buffered saline. Sections were incubated overnight at 4°C with primary antibody β-galactosidase (1:300 dilution; Thermo Fisher Scientific). Specific secondary Alexa-488 antibody (1:200 dilution; Life Technologies, Carlsbad, CA) was used for 1 hour at room temperature, and sections were mounted with an aqueous agent (ProLong Gold, Thermo Fisher Scientific). Nuclei were counterstained with DAPI (Sigma–Aldrich). Images were obtained using an SP5 confocal microscope (Leica Microsystems, Wetzlar, Germany).
Experimental iodine deficiency
Female 5- to 6-month-old Wt and Dehal1KO mice were fed a low-iodine diet (TD180914; Envigo, Indianapolis, IN) containing 0.05 μgI/g, and separated into three groups receiving 1400 μgI/L, 200 μgI/L, or no-iodine in the drinking water, resulting in total estimated iodine intakes of 5.8 (normal iodine diet [NID]), 1.0 (low iodine diet [LID]) and 0.2 μgI (very low iodine diet [VLID]) per animal per day, respectively. 12 Urine was collected using hydrophobic sand (Sodispan Research, Madrid, Spain). Mice were anesthetized with 1.5–2% isoflurane (Esteve Laboratories, Barcelona, Spain) inhalation in O2 (0.9L O2/min), and blood was collected by cardiac puncture. Tissues (thyroid, kidney, and liver) were collected on days 0, 12, and 28. Urine and blood from 10-day-old pups, 1-month juveniles, and 3-month adults, fed regular diet (1.4 μgI/day; SAFE diet), were also collected.
Determination of plasma thyrotropin
Plasma thyrotropin (TSH) levels were measured by radioimmunoassay as described in detail previously. 13
Determination of MIT, DIT, T3, and T4 in plasma and/or urine
Previously published methods 14 –17 were modified to allow the simultaneous quantification of MIT, DIT, T3, and T4.
Sample preparation
One hundred microliters of urine/plasma was added with 5.4 pmol 13 C9-MIT, 4.0 pmol 13 C9-DIT, 2.8 pmol 13 C6-T3, and 2.4 pmol 13 C6-T4, vortex mixed, equilibrated for 30 min at room temperature; 300 μL of cold acetone was added, and the obtained mixtures were kept at 4°C for 30 min to achieve an optimal sample deproteinization. Then, samples were centrifuged 10 min at 22,780 g, and the resulting supernatants were warmed up to 40°C and dried under a nitrogen stream. The residues were reconstituted with 200 μL of 3.0 N hydrochloric acid in n-butanol and incubated for 45 min at 60°C for the formation of the corresponding butyl esters of tyrosines, THs, and their internal standards.
After evaporation to dryness, the samples were reconstituted with 500 μL of 0.1 M potassium acetate buffer (pH = 4) and loaded onto Agilent (Santa Clara, CA) Bond Elut Certify 130 mg SPE cartridges to perform the extraction according to a procedure previously described. 15 The eluate was dried once again under nitrogen, and the dried residue was reconstituted with 100 μL of acetonitrile/0.1 M hydrochloric acid (50/50 by volume), and 1 μL of it was injected into the liquid chromatography tandem mass spectrometry (LC-MS-MS) system. Calibration curves ranged from 0.1 to 100.0 ng/mL and were daily prepared by serial dilution in methanol and derivatized by the same procedure used for the samples.
Instrument layout
AB Sciex (Concord, Ontario, Canada) QTRAP 6500+ triple quadrupole mass spectrometer with ESI source, coupled to an Agilent 1290 UHPLC system, fitted with a 110 Å, 2 × 50 mm, 3 μm particle size, Gemini C18 column (Phenomenex, Torrance, CA).
Operative conditions
The chromatographic separation was carried out as previously reported by Guzzolino et al. 17 Mass spectrometry operated in positive ions selected reaction monitoring mode, by using optimized source and compound parameters to achieve the best possible performances. Three transitions were used to monitor each compound and internal standard: the one with the highest signal-to-noise ratio was used as a quantifier (Q), while the other two as qualifiers (q). They were the following: MIT-But: 363.9 → 261.8 daltons (Da) (Q), 363.9 → 135 Da (q), 363.9 → 291.0 Da (q); 13C9-MIT-But: 373.1 → 270.1 Da (Q), 373.1 → 143.2 Da (q), 373.1 → 300.2 Da (q); DIT-But: 489.9 → 387.9 Da (Q), 489.9 → 260.9 Da (q), 489.9 → 290.0 Da (q); 13C9-DIT-But: 498.8 → 395.8 Da (Q), 498.8 → 268.8 Da (q), 498.8 → 298.9 Da (q); T3-But: 707.9 → 605.9 Da (Q), 707.9 → 479.1 Da (q), 707.9 → 651.9 Da (q); 13C6-T3-But: 713.9 → 611.9 Da (Q), 713.9 → 485.1 Da (q), 713.9 → 657.9 Da (q); T4-But: 833.9 → 731.99 Da (Q), 833.9 → 605.0 Da (q), 833.9 → 777.9 Da (q); 13C6-T4-But: 839.9 → 737.9 Da (Q), 839.9 → 611.0 Da (q), 839.9 → 783.8 Da (q).
Validation of the method is presented in Supplementary Figure S1.
Urinary iodine concentration
Mice urine samples were processed by the modified Sandell–Kolthoff (S-K) method 18 and expressed relative to creatinine concentrations.
Study approval
Animal studies followed the guidelines from the European Animal Research Ethical Committee and were approved by the Animal Research Ethical Committee of the Autonomous University of Madrid, Spain (Proex-13/17).
Statistics
GraphPad Prism software v5 (San Diego, CA) was used for statistical analysis. Results are expressed as mean ± standard error of the mean. Differences were assessed by the nonparametric Mann–Whitney test for two independent samples and were considered statistically significant when the p-value was <0.05 (*p < 0.05, **p < 0.01, ***p < 0.001).
Results
Characterization of Dehal1KO mice
The generation of the Dehal1KO mouse by gene trap technology results in a recombinant protein composed of a truncated Dehal1 protein encoded by exon 1, followed by the β-galactosidase enzyme (trDehal1-βG) under the control of the endogenous Dehal1 promoter (Fig. 1A). Chimeric male mice with germline transmission were bred to successfully generate heterozygous and homozygous Dehal1KO mice (Fig. 1B). The absence of Wt Dehal1 RNA and protein was confirmed in thyroids of homozygous Dehal1KO mice (Figs. 1C–E). Whole-mount X-Gal labeling at low-power magnification showed the presence of the recombinant Dehal1-β-Gal protein in developing thyroids of KO mice from embryonic days 13.5 (E13.5) and E15, as well as in postnatal day 120 (P120) adults (Fig. 1F.a–F.c). In P120 KO mice, the recombinant protein was highly expressed in the thyroid, staining follicular structures, but not parafollicular cells (Fig. 1G.a, G.b).
Tissue expression of Dehal1
The activity of the endogenous promoter driving trDehal1-β-Gal expression was investigated in the developing thyroid, kidney, and liver, sites of reported IYD activity. 19 At E13.5, X-Gal labeling was detected in the pre-follicular stage of the thyroid, in tubular structures of the kidneys, and was not detected in the liver (Fig. 2A). Unexpectedly, intense labeling was identified in epithelial cells of the developing choroid plexus (Fig. 2A).
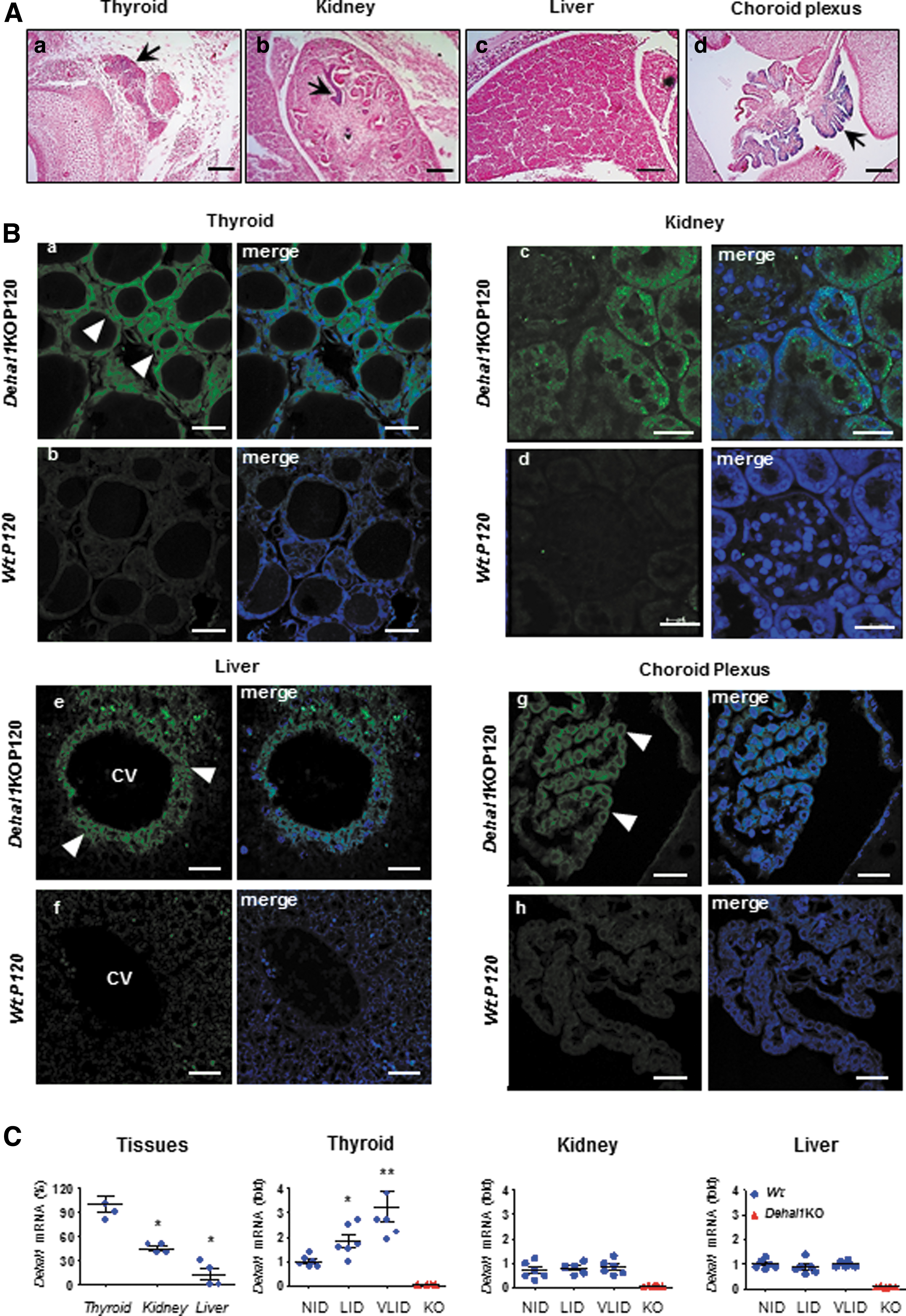
Extrathyroidal expression of Dehal1 in embryos and adult mice. (
In adult Dehal1KO mice, thyrocytes were β-Gal immunoreactive (ir)-positive (Fig. 2B.a). In the kidneys, β-Gal ir-positive cells were observed in proximal convoluted tubules (Fig. 2B.c) showing a cytoplasmic granular pattern. In the liver, this sensitive technique identified ir-positive hepatocytes lining the central vein (Fig. 2B.e). In the choroid plexus β-Gal ir-positive epithelial cells and neighbor, ependymocytes were also observed lining the ventricular surface (Fig. 2B.g).
Dehal1 RNA was most abundant in the thyroid and expressed in the kidneys and liver at two- and threefold lower levels, respectively (Fig. 2C), suggesting that Dehal1 could have additional roles in mammalians. Therefore, Dehal1 expression was investigated in response to iodine deficiency in the three tissues from Wt mice. LID and VLID triggered 2- and 3.5-fold increase of Dehal1 mRNA in the thyroid, but not in the kidneys or liver (Fig. 2C), indicating that the Dehal1 exerts iodine homeostasis through the thyroid gland.
Dehal1KO mice have negative iodine balance and iodotyrosines urinary loss
A novel LC-MS-MS method with high sensitivity and specificity was developed for the concomitant determination of iodotyrosines and iodothyronines (Fig. 3A). Adult Dehal1KO mice showed normal plasma concentrations of TSH, T4, and T3 when fed NID (Fig. 3B). However, they showed 10-fold increased plasma MIT and DIT compared with Wt mice (p < 0.0001) (Fig. 3B). In urine, MIT and DIT reached a 30-fold increase in Dehal1KO mice versus Wt mice (p < 0.0001) (Fig. 3B), indicating that both plasma and urine iodotyrosines are highly discriminative to unveil Dehal1 defects in rodents. Finally, Dehal1KO mice excrete nearly double the amount of iodine in urine, compared with Wt mice (p < 0.001) (Fig. 3B).
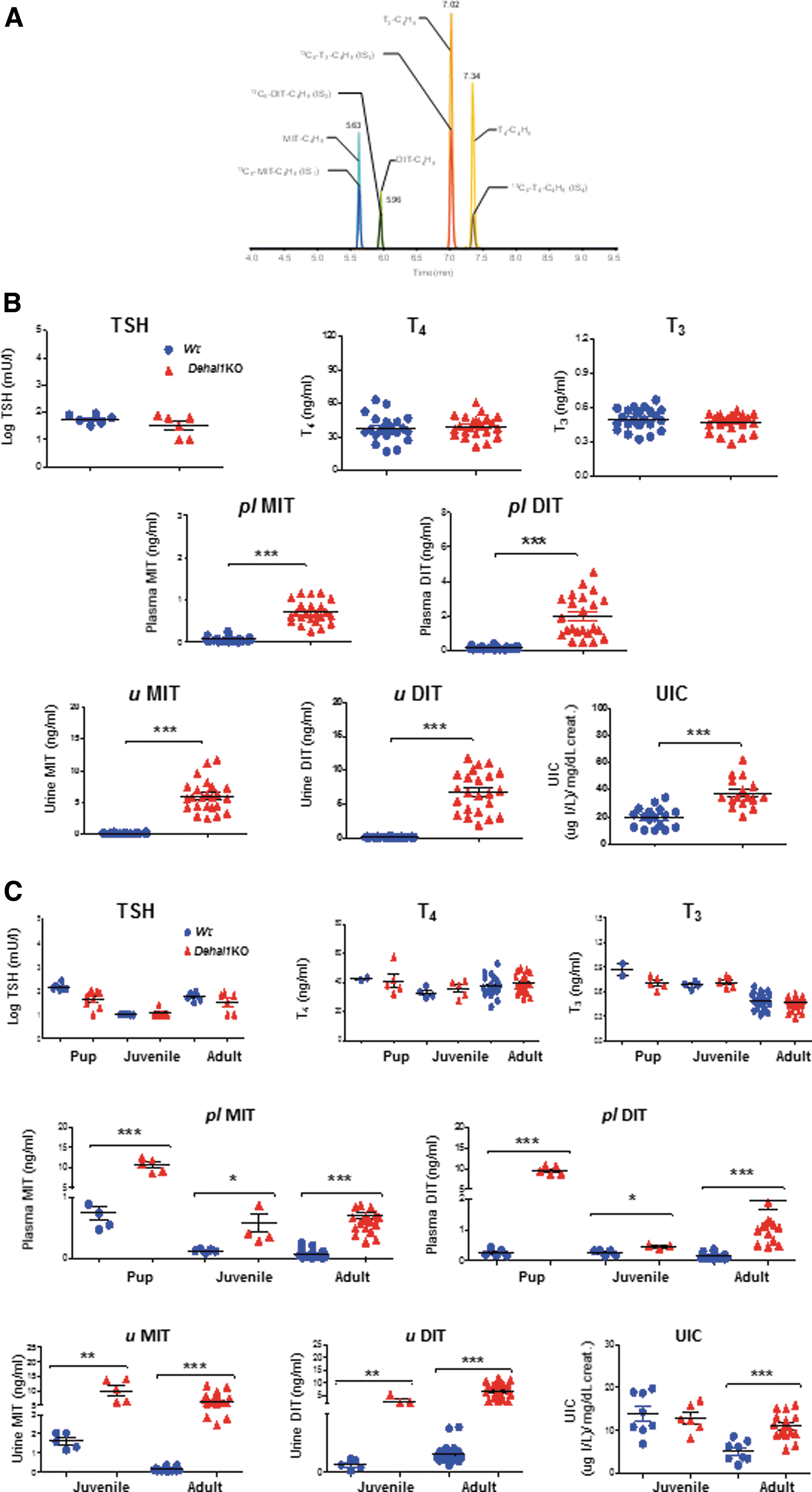
Hormonal and biochemical characterization of Wt and Dehal1KO mice. (
Since mice of both genotypes were fed the same amount of iodine, this indicates that (a) Dehal1KO mice have negative iodine balance through excess iodine excretion caused, at least in part, by the chronic urinary loss of MIT and DIT, and (b) the organic iodide contained in iodotyrosines is measured by S-K, while this technique is usually assumed to report only the inorganic (dietary) iodine content of the urine. This finding may have relevant methodological implications for ID studies (Supplementary Fig. S2). In summary, under sufficient iodine supply, Dehal1KO mice can sustain euthyroidism, although they have a negative iodine balance. This makes Dehal1KO mice extremely vulnerable to iodine shortages. Furthermore, this finding indicates that elevated plasma or urinary iodotyrosines are reliable indexes of IYD deficiency, even under conditions of iodine sufficiency.
Iodotyrosines are increased from early life in Dehal1KO mice
As MIT and DIT are increased in adult Dehal1KO mice, we investigated iodotyrosine concentrations in early postnatal life. Plasma from 10-day-old pups and plasma and urine from juvenile and young adult Dehal1KO and Wt mice were analyzed. On NID, pups, juveniles, and young adults of both genotypes showed comparable levels of plasma TSH, T4, and T3 (Fig. 3C). However, only Dehal1KO pups, juveniles, and young adults exhibited a marked increase in plasma MIT and DIT (Fig. 3C). This iodotyrosine elevation was particularly large at the neonatal stage (>10-fold increased MIT/DIT in Dehal1KO vs. Wt pups), while urine from juvenile and young adult KO mice showed a fivefold increase compared with Wt mice (Fig. 3C). Of note, UIC was elevated in Dehal1KO mice versus Wt mice only when they reached the adult stage (Fig. 3C). This could be attributed to functional immaturity of juvenile (and likely neonatal) kidneys, conferring a unique advantage to iodotyrosines to identify the postnatal disease timely.
The findings show that even normal iodine intake significantly increased iodotyrosines, detectable in plasma and urine from 10-day-old Dehal1KO pups, which is equivalent to birth in humans, potentially allowing an early diagnosis of the disorder.
Iodine shortage rapidly triggers hypothyroidism in Dehal1KO mice
To analyze the role of Dehal1 as a function of controlled iodine supplies, adult Wt and Dehal1KO mice were fed NID, LID, and VLID for 28 days. Plasma TSH, T4, and T3, plasma and urinary iodotyrosines, and UIC were determined at 0, 12, and 28 days (Fig. 4A). On NID and LID, Wt and Dehal1KO mice showed comparable TSH, T4, and T3 levels (Fig. 4B). However, Dehal1KO mice fed VLID showed marked elevation of TSH and decreased T4 and T3, developing profound hypothyroidism by day 28, while Wt mice were euthyroid. Hypothyroidism in Dehal1KOs developed in a very short time, being already present on day 12. A transient increase of T3 was detectable at day 12 in Dehal1KO mice, likely related to the reported preferential synthesis of T3 in ID, mediated by D1 deiodination of T4. 20 After the T3 peak, a complete failure of TH synthesis occurred in Dehal1KO, while Wt mice stayed euthyroid (Fig. 4B).

Model of incremental iodine deficiency and biochemical characterization of plasma and urine in Wt and Dehal1KO mice. (
Regarding iodotyrosines, Dehal1KO mice on NID showed a large elevation of plasma MIT and DIT compared with Wt mice, while, on LID, iodotyrosine levels declined progressively but always remained higher than in Wt mice (p > 0.005). By contrast, VLID (along with severe hypothyroidism) caused a steep decrease of iodotyrosines in Dehal1KO mice so that significant differences between genotypes became no longer detectable at the end of the experiment (Fig. 4B).
In urine, iodotyrosines mirrored the changes in plasma (Fig. 4C); however, despite a decreasing trend when on iodine restriction, MIT and DIT concentrations remained markedly higher in Dehal1KO mice throughout the experiment (p < 0.001 at day 28) (Fig. 4C). Notwithstanding, DIT, which has two iodine atoms, showed a steeper decrease than MIT in Dehal1KO mice on VLID, probably reflecting the depletion of iodine reserves. Finally, UIC was significantly higher in Dehal1KO mice versus Wt mice only on NID and LID diets, with no difference detectable between genotypes on VLID (Fig. 4C).
In summary, the findings indicate a marked sensitivity of Dehal1-deficient mice to ID, developing severe hypothyroidism in a very short time.
Discussion
Considering the central role of Dehal1 in iodine metabolism, we investigated the first mammalian model of dehalogenase deficiency under graded iodine supplies.
Our study showed that Dehal1KO mice have a negative iodide balance compared with normal mice under conditions of iodine sufficiency due to the large excretion of iodotyrosines in plasma and urine. This abnormality is reflected in the UIC of Dehal1KO mice since, contrary to standard thinking, the S-K technique measured both inorganic and organic (iodotyrosines) iodine. In a translational view, this could reduce the efficacy of S-K to detect “low iodine intakes” from urines eventually containing iodotyrosines.
Furthermore, we identified that Dehal1KO mice on iodine restriction develop profound hypothyroidism in a surprisingly short time compared with normal mice, which remain euthyroid in our experimental design. This indicates that the thyroids of Dehal1KO mice do not contain enough iodine reserves to compensate (even very short) periods of iodine shortage, suggesting a relevant role of Dehal1 in iodine storage during iodine sufficiency. Thus, Dehal1 activity revealed necessary not only in scenarios of iodine restriction to “salvage iodine” for TH synthesis but also as an enzyme governing the efficient building-up of iodine reserves in the thyroid gland. Further research on the mechanisms how Dehal1 influences iodine storage in the thyroid is warranted.
Our findings may have important clinical implications. In humans, the determination of MIT and DIT may be useful for the diagnosis of DEHAL1 defects when hypothyroidism is present but, most importantly, before hypothyroidism is established. This could be particularly relevant for neonates since, as shown here for mice, affected babies may not show hypothyroidism at birth, but iodotyrosines could be invariably elevated. This would allow detection of the risk for hypothyroidism when the brain is actively developing, thus helping the prevention of mental retardation. 7,8 Our experimental model needs to be extended to precisely define the dynamics of iodine conservation in chronic ID or in circumstances of increased iodine demands, such as pregnancy, where our model can be instrumental to investigate the consequences for the progeny. 21
Finally, the expression of Dehal1 in extrathyroidal tissues is intriguing, especially in the choroid plexus. Further to its capacity for deiodination, Dehal1 can debrominate and dechlorinate tyrosines. 22,23 Recently, essential functions for bromide and chloride were unveiled in the brain, 24,25 encouraging the investigation of whether Dehal1 could participate in cell homeostasis of other halogens in specific tissues.
In conclusion, Dehal1 is a key factor for iodine metabolism in the thyroid gland. In periods of iodine deficiency, it allows the reuse of iodine from iodotyrosines for hormone synthesis, but in iodine sufficiency, it contributes to optimal iodine storage in the thyroid gland. The model of dehalogenase deficiency presented here supports the proposed paradigm that genetic factors mediate the relation between iodine nutrition and iodine status (reserves). From a translational perspective, elevated iodotyrosines are biomarkers for the risk and vulnerability to iodine-deficient hypothyroidism in asymptomatic individuals with defective iodine recycling that now await validation in the clinical arena.
Footnotes
Acknowledgments
The Center for Instrumentation Sharing of the University of Pisa is kindly acknowledged for providing the Sciex QTrap 6500+ mass spectrometer, which was used for the mass spectrometric assays. We are indebted to Prof. Pere Berbel for substantial comments on the article, and to Dr. Carlota Largo, Mrs. Elisa Pulido, and Mrs. Mercedes Tanarro for their technical contribution.
Authors' Contributions
C.G.-G., M.B., and P.A. designed and conducted experiments, analyzed the data, and contributed to the generation of figures and the draft of the article. Y.A., F.S.-L., X.-H.L., J.G.-G., A.B., D.M., A.M., R.M., and A.R.M. contributed to the methodology. A.B.-S., A.R.M., A.S., and M.P.d.M. validated the data and supervised the methodology. A.R.M., M.P.d.M., A.S., J.B., and S.R. provided intellectual input into the design of the study and reviewed and edited the article. J.C.M. and R.Z. conceived the project, wrote the article, and acquired funding and resources for the study. C.G.-G., M.B., and P.A.: The co-first authors contributed to the vast majority of the experimental design and execution at separate and collaborating institutions through time. To assign the order, the relevance and time invested to complete their respective contributions were considered.
Author Disclosure Statement
The authors have declared that no conflict of interest exists.
Funding Information
The work was supported with public funding (AES PI16/00830) to José Carlos Moreno from the Carlos III Health Institute (ISCIII) of the Spanish Ministry of Health and Research, European FEDER Funds and, in part, from the DK15070 grant to Samuel Refetoff from the National Institutes of Health, USA. The rest of the authors have no funding contributions to declare.
Supplementary Material
Supplementary Table S1
Supplementary Figure S1
Supplementary Figure S2