
Abstract
Introduction:
Patients lacking functional monocarboxylate transporter 8 (MCT8), a highly specific thyroid hormone (TH) transporter, present severe psychomotor disabilities. MCT8 deficiency leads to peripheral hyperthyroidism and brain hypothyroidism, the latter due to impaired transport of TH across brain barriers. Available treatments for patients are limited and aim to overcome the limited TH transport across brain barriers. The use of TH analogues such as 3,3′,5-triiodothyroacetic acid (TRIAC) that do not require MCT8 to cross the cellular membranes is considered a potential therapy for MCT8 deficiency. Previous studies have shown that systemic administration of TRIAC at therapeutic doses does not increase TRIAC content in the brain, while intracerebroventricular (ICV) administration of therapeutic doses of TRIAC increases TRIAC content in the brain but does not mediate thyromimetic effects. In view of this, we hypothesize that ICV administration of high doses of TRIAC can mediate thyromimetic effects in the brain without worsening the brain hypothyroidism or peripheral hyperthyroidism of patients.
Methods:
We administered 400 ng/g of body weight per day of ICV TRIAC in a mouse model of MCT8 deficiency: Mct8−/y and deiodinase 2 (Dio2)−/− double knockout mice. The effects of this treatment on TH and TRIAC levels/content in blood and tissues were determined by radioimmunoassay and effects on TH-regulated genes were assessed by real time-quantitative polymerase chain reaction in peripheral and central tissues.
Results:
ICV administration of high doses of TRIAC ameliorated the peripheral hyperthyroidism. In the brain, this treatment did not further aggravate brain hypothyroidism and increased TRIAC content in several brain regions; however, only moderate thyromimetic activity was observed in restricted brain areas.
Conclusion:
Administration of high doses of TRIAC by ICV delivery at juvenile stages in a mouse model of MCT8 deficiency is effective in normalizing peripheral hyperthyroidism but exerts minimal thyromimetic activity in the brain.
Introduction
Mutations in the X
MCT8-deficient patients present altered circulating levels of TH with elevated T3, causing hyperthyroidism in peripheral tissues, low T4 and 3,5′,3′-T3 (reverse T3), and normal or slightly elevated thyrotropin (TSH). 1,2 Paradoxically, patients present a state of brain hypothyroidism 7 due to impaired TH transport across the brain barriers (BBs; including the blood-brain and the blood-cerebrospinal fluid barriers) in the absence of MCT8. 8 –11
Therapeutic options for MCT8-deficient patients are limited. An optimal therapeutic strategy should improve the neurological defects derived from TH deficiency in the brain, while alleviating the peripheral thyrotoxicosis resulting from peripheral hyperthyroidism. In recent years, the TH analogue 3,3′,5-triiodothyroacetic acid (TRIAC) has become a popular experimental alternative treatment for MCT8 deficiency, as an orphan medicinal product. TRIAC is a TH analogue that can access brain cells in the absence of MCT812 and can bind to TH receptors (TRs) 13,14 to mediate thyromimetic actions. 15
In addition, clinical experience with TRIAC has demonstrated its ability to inhibit TSH secretion with a concomitant decrease in the production of endogenous TH. 16,17 In view of this, a clinical trial was initiated to assess the safety of TRIAC treatment in MCT8-deficient patients and the effectiveness of this analogue in improving their peripheral thyrotoxicosis. 18 TRIAC treatment reduced serum T3 levels to the normal range, improving weight, heart rate, and blood pressure. However, a trend toward gross motor function improvements was only observed when children initiated the treatment before four years of age.
Preclinical studies in animal models of the syndrome have indicated that systemic administration of TRIAC has the ability to prevent abnormal neuronal development and myelination impairments when administered in high doses (200–400 ng/g of body weight [BW] per day) at early postnatal stages, 12,15 suggesting that treatment with high doses of TRIAC might be beneficial in MCT8 deficiency. However, additional preclinical studies have demonstrated that oral treatment with therapeutic doses of TRIAC (30 ng/g BW per day) restores T3 plasma levels but severely decreases T4 levels, aggravating brain hypothyroidism in MCT8 deficiency by further decreasing brain T3 content. 19
This suggests that systemic administration of high doses of TRIAC might be detrimental for MCT8-deficient subjects. Preclinical studies in animal models of MCT8 deficiency have also explored the administration of therapeutic doses of TRIAC (30 ng/g BW per day) into the lateral ventricle of the brain by intracerebroventricular (ICV) infusion, bypassing the limitation of TH entry through the BBs in the absence of MCT8. 20 This approach increased 2.2-fold TRIAC levels in plasma, but it did not alter TH plasma levels or the expression of T3-target genes in peripheral tissues. In the brain, this treatment did not alter TH content, but it increased TRIAC content 1.5-fold; however, it did not induce TH-mediated actions on the explored target genes.
In view of all the aforementioned, we hypothesized that high doses of TRIAC (400 ng/g BW per day), administered by ICV infusion, might mediate thyromimetic actions in the brain by regulating TH-target genes without affecting circulating TH levels or worsening peripheral hyperthyroidism and brain hypothyroidism. Mice lacking MCT8 do not replicate the gross neurological abnormalities present in patients, 21 due to a compensatory mechanism involving the higher presence of the T4 transporter organic anion-transporting polypeptide 1c1 at the mouse BB in comparison with humans 22 –25 and increased activity of the enzyme deiodinase type 2 (DIO2), 26,27 which converts T4 into the transcriptionally active TH form T3.
For this reason, we have administered 400 ng/g BW of TRIAC per day by ICV delivery in mice deficient in MCT8 and DIO2 (Mct8/Dio2KO), a mouse model that replicates the endocrine symptoms and neurological impairments of patients. 28,29 Moreover, since most of the patients diagnosed to date are now in their late childhood years, adolescence, or even adulthood, we have performed the studies at juvenile stages to assess if ICV administration of high doses of TRIAC is able to restore neurological impairments.
Methods
Animal models and experimental design
All experimental procedures on research animals were conducted in strict accordance with the European Union Council guidelines (directive 2010/63/UE) and Spanish regulations (R.D. 53/2013), and were approved by the ethics committee “Comité de Ética y Experimentación Humana y Animal” (CEEHA) at CSIC and by the Comunidad Autónoma de Madrid Review Board (Proex 162/17) on the use of animals for scientific purposes.
Mice were maintained under a 12:12 light–dark cycle, at a constant temperature of 22°C ± 2°C and ad libitum access to food and water. Experiments were carried out in wild-type (Wt) and MCT8-DIO2-deficient mice (Mct8/Dio2KO) that were bred and genotyped as described. 28 Since MCT8 deficiency only affects male patients, studies were performed exclusively in male mice. See Supplementary Methods for more details on experimental design.
The surgical implantation of osmotic minipumps into the right lateral ventricle was performed as described. 20 In brief, 3-month-old animals were anesthetized with ketamine (75 μg/g BW) and medetomidine hydrochloride (1 μg/g BW) and placed on the stereotaxic frame. A hole was drilled at coordinates bregma −0.5 mm, 1.0 mm lateral in the skull. An Alzet Brain Infusion Kit 3 (0008851; Alzet) catheter connected to a 2002 Alzet osmotic minipump (0000296; Alzet) was implanted into the lateral ventricle, at a depth of 2 mm, in Wt and Mct8/Dio2KO male mice.
Wt animals were treated with artificial cerebrospinal fluid (aCSF or vehicle-treated) (n = 9) to assess if TRIAC treatment can revert the alterations in Mct8/Dio2KO mice to Wt values. Mct8/Dio2KO animals were treated with aCSF (n = 8) or with high doses of TRIAC [400 ng/g BW per day (n = 7)]. Mice were anesthetized 12 days after surgery as described earlier, blood was extracted by retro-orbital collection for TH and TRIAC determinations, and they were transcardially perfused with saline to remove blood from tissues. Plasma, liver, heart, cerebral cortex, cerebellum, striatum, hippocampus, and a ventral brain region containing the midbrain, thalamus, and hypothalamus were harvested and kept at −80°C until further use.
Radioimmunoassays of T4, T3, and TRIAC
T4, T3, and TRIAC were extracted from individual 80 μL aliquots of plasma. T3 and T4 from the cerebral cortex were extracted and purified as previously described 30,31 by a method involving the use of DOWEX AG 1-X2 resin (140-1251; Bio-Rad), which retains TRIAC during the extraction, avoiding any interference of TRIAC in the T3 radioimmunoassay (RIA). TRIAC from the cerebral cortex, cerebellum, and ventral brain region was extracted as described, without the use of DOWEX columns. The purified extracts were used for T4, T3, and TRIAC determinations by sensitive RIAs. 20,30 In plasma samples, T3 data were corrected for the cross-reactivity of TRIAC on the T3 antiserum (17%).
Gene expression
RNA extraction from liver, heart, cerebral cortex, striatum and hippocampus, cDNA synthesis, and real time-quantitative polymerase chain reaction (qPCR) were performed as described. 20 The 18S gene was used as internal standard and, for analysis, the cycle threshold was used. The expression of the T3-regulated genes in Table 1 was evaluated using Applied Biosystems TaqMan probes.
Taqman Probes Used for Real-Time Quantitative Polymerase Chain Reaction
Statistics
Data are expressed in scatter dot plots as mean ± SD. Data were tested for normal distribution using the Kolmogorov–Smirnov test. Parametric data sets were tested using one-way analysis of variance and the Tukey's multiple comparisons test and nonparametric data set by Kruskal–Wallis test. Detection of outliers from a Gaussian distribution was performed by ROUT tests and were excluded from further analyses. 32 A complete list of outlier exclusion can be found in Supplementary Methods. The number of samples used for each test (that may vary due to loss of tissue at collection) is indicated in each figure. Significant differences are represented as *p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001. All analyses were conducted using GraphPad Prism 9.
Results
ICV TRIAC treatment further decreases plasma T4 and T3 levels
ICV administration of 400 ng/g BW of TRIAC for 12 days caused more than a 2-fold increase in plasma TRIAC levels compared with Wt and vehicle-treated Mct8/Dio2KO controls (p < 0.0001 and p = 0.0036, respectively) (Fig. 1A). Furthermore, the plasma T4 levels were decreased 1.6-fold in vehicle-treated Mct8/Dio2KO animals (p < 0.0001), and were further decreased 5-fold after ICV TRIAC treatment (p = 0.0012; Fig. 1B) compared with Wt. The plasma T3 levels were 3.5-fold higher in vehicle-treated Mct8/Dio2KO compared with Wt animals (p < 0.0001; Fig. 1C), as previously described. 28 After ICV administration of TRIAC, T3 levels decreased in Mct8/Dio2KO to nondetectable values (Fig. 1C).

Effects of treatment on analogue/hormone plasma levels. (
ICV TRIAC treatment restores several T3-regulated genes in peripheral organs
We explored the expression of several T3-regulated genes in the liver and heart to assess the effects of the treatment on peripheral organs. In the liver, Dio1 expression was significantly increased in vehicle-treated Mct8/Dio2KO in comparison with Wt mice (p < 0.0001), due to their high T3 plasma levels. TRIAC treatment significantly decreased Dio1 expression (p < 0.0001) (Fig. 2A), most probably due to the decrease in plasma T3 levels after treatment, although it was not fully normalized. Gsta2 expression was not different in any of the experimental conditions (p = 0.0992; Fig. 2B) and Ucp2 expression, which was increased in the liver of Mct8/Dio2KO in comparison with Wt animals (p = 0.0250), was normalized by ICV TRIAC treatment (p = 0.1038; Fig. 2C).

Effects of treatment on T3-regulated genes in peripheral organs. (
In the heart, expression of the T3-regulated gene Hcn2 33 was increased in Mct8/Dio2KO in comparison with Wt mice (p < 0.0001) and expression of this gene was normalized after treatment with TRIAC (Wt vs. Mct8/Dio2KO+TRIAC, p = 0.9554; Fig. 2D). While the expression of the T3-regulated genes Atp2a2, 34 Dio1, and Myh6 35 was not altered in vehicle or TRIAC-treated Mct8/Dio2KO (Fig. 2E, F, G), Myh7 35 highly increased in Mct8/Dio2KO mice after TRIAC treatment (p < 0.0001; Fig. 2H).
ICV TRIAC treatment increases TRIAC content in the brain and decreases T4 content
To assess whether ICV TRIAC treatment resulted in an increase of TRIAC content in the brain, we directly measured TRIAC content in several brain regions by RIA. TRIAC content was not significantly different in Wt and vehicle-treated Mct8/Dio2KO in all the brain regions explored, which included the cerebral cortex (p = 0.2729), the cerebellum (p = 0.9991), and a ventral brain region containing the midbrain, thalamus, and hypothalamus (p = 0.9995; Fig. 3A–C). ICV infusion of TRIAC increased TRIAC content in all of the regions assessed: 3.5-fold in the cerebral cortex (p < 0.0001), 3.6-fold in the cerebellum (p = 0.0048), and 16.5-fold in the ventral region (p < 0.0001) in TRIAC-treated Mct8/Dio2KO mice in comparison with vehicle-treated Mct8/Dio2KO animals (Fig. 3A–C). Moreover, we assessed the effects of ICV TRIAC treatment on the brain T4 and T3 content.
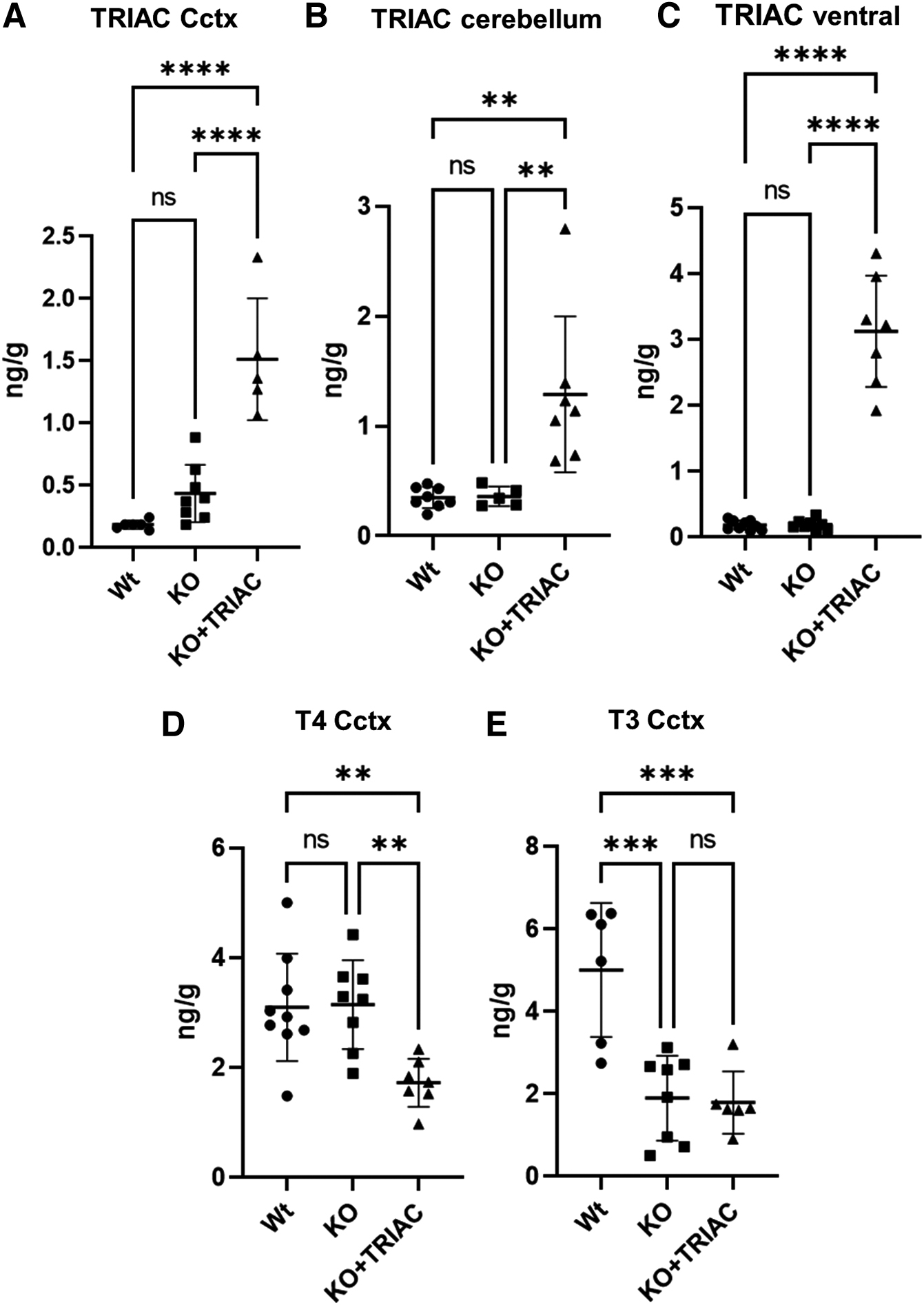
Effects of treatment on analogue/hormone brain content. (
T4 content was not altered in the cerebral cortex of Mct8/Dio2KO mice compared with Wt counterparts (Fig. 3D), as described. 28 ICV TRIAC treatment in Mct8/Dio2KO animals decreased T4 content 1.8-fold in the cerebral cortex (p = 0.0066) in comparison with vehicle-treated Mct8/Dio2KO littermates (Fig. 3D), most probably due to the decrease in plasma T4 levels after treatment with TRIAC. This finding contrasts with previous results where ICV administration of therapeutic doses of TRIAC did not impact T4 brain content. 20 Despite the decreases in T4 brain content, T3 did not further decrease in Mct8/Dio2KO mice after ICV TRIAC administration in comparison with vehicle-treated Mct8/Dio2KO animals (p = 0.9846), in which cerebral cortex T3 content was 2.6-fold lower than in Wt animals (p = 0.0004; Fig. 3E).
ICV TRIAC treatment regulates the expression of few T3-regulated genes in specific brain regions
As THs mediate most of their actions by binding to TRs and modulating the expression of T3-regulated genes, we explored whether increases in brain TRIAC content after ICV delivery are enough to modulate the expression of T3-regulated genes. To this aim, we studied the expression of Cbr2, Cntn2, Dio3, Flywch2, Hr, Nefm, and Sema7a genes, known to be downregulated in the cerebral cortex of Mct8/Dio2KO mice. 26,36 As expected, the expression of all these genes was downregulated in Mct8/Dio2KO mice in comparison with the Wt group (Cbr2, p = 0.0007; Cntn2, p = 0.0002; Dio3, p = 0.0449; Flywch2, p = 0.0023; Hr, p < 0.0001; Nefm, p < 0.0001 and Sema7a, p = 0.0053). ICV treatment with TRIAC was not able to modulate the expression of any of them (Fig. 4A–G).
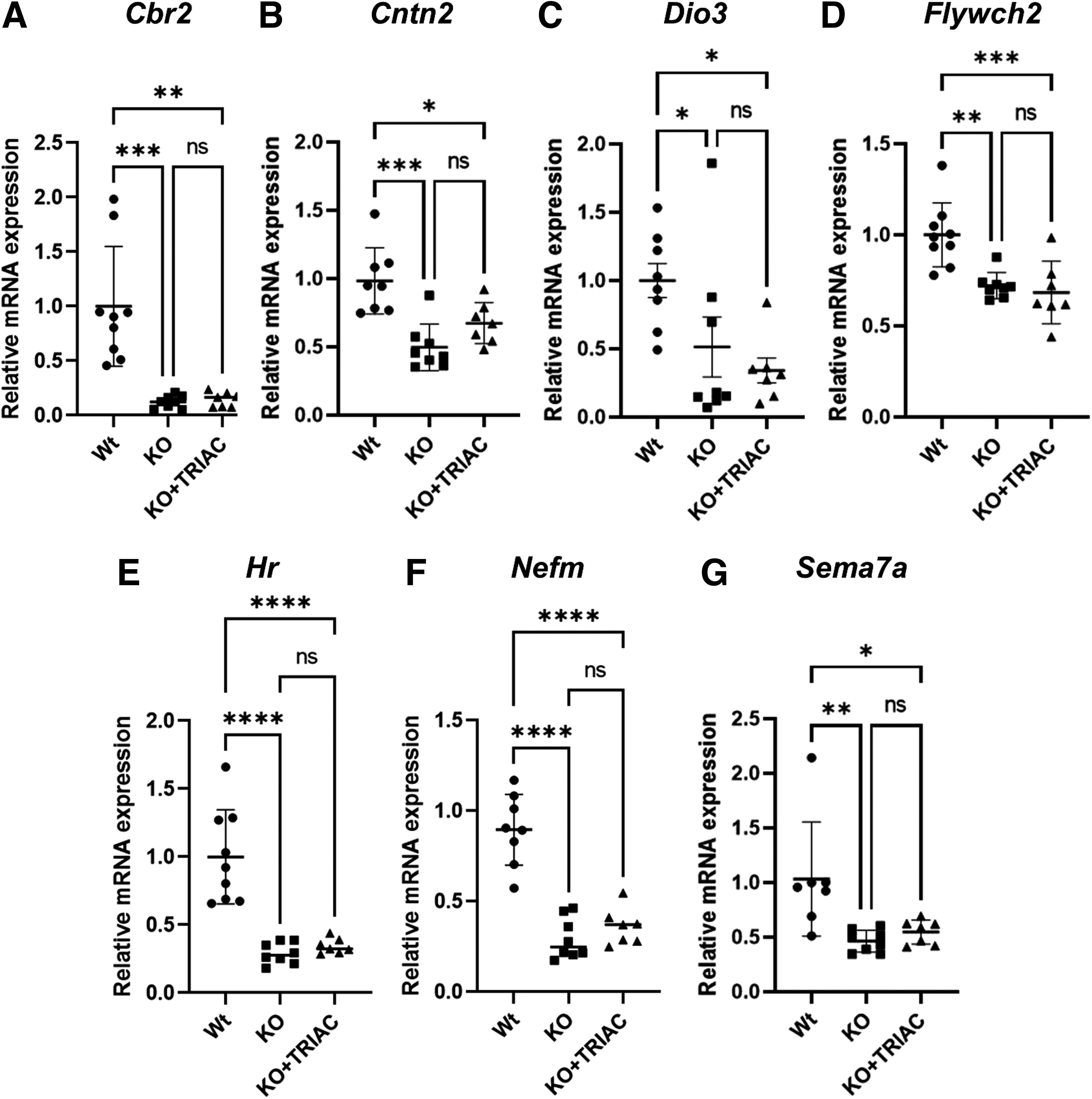
Effects of treatment on T3-regulated genes in the cerebral cortex. (
The ability of ICV delivery of TRIAC to modulate the expression of T3-regulated genes was further assessed in other brain regions: the striatum and the hippocampus. In the striatum, Dio3 was not different in any of the experimental conditions (p = 0.0604; Fig. 5A). The expression of the T3-regulated gene Hr was downregulated in both vehicle- (p = 0.0072) and ICV TRIAC-treated Mct8/Dio2KO animals (p = 0.0046; Fig. 5B). We also explored the expression of Nrgn, a T3-target gene that has been shown to be highly sensitive to T3 in the striatum. 37
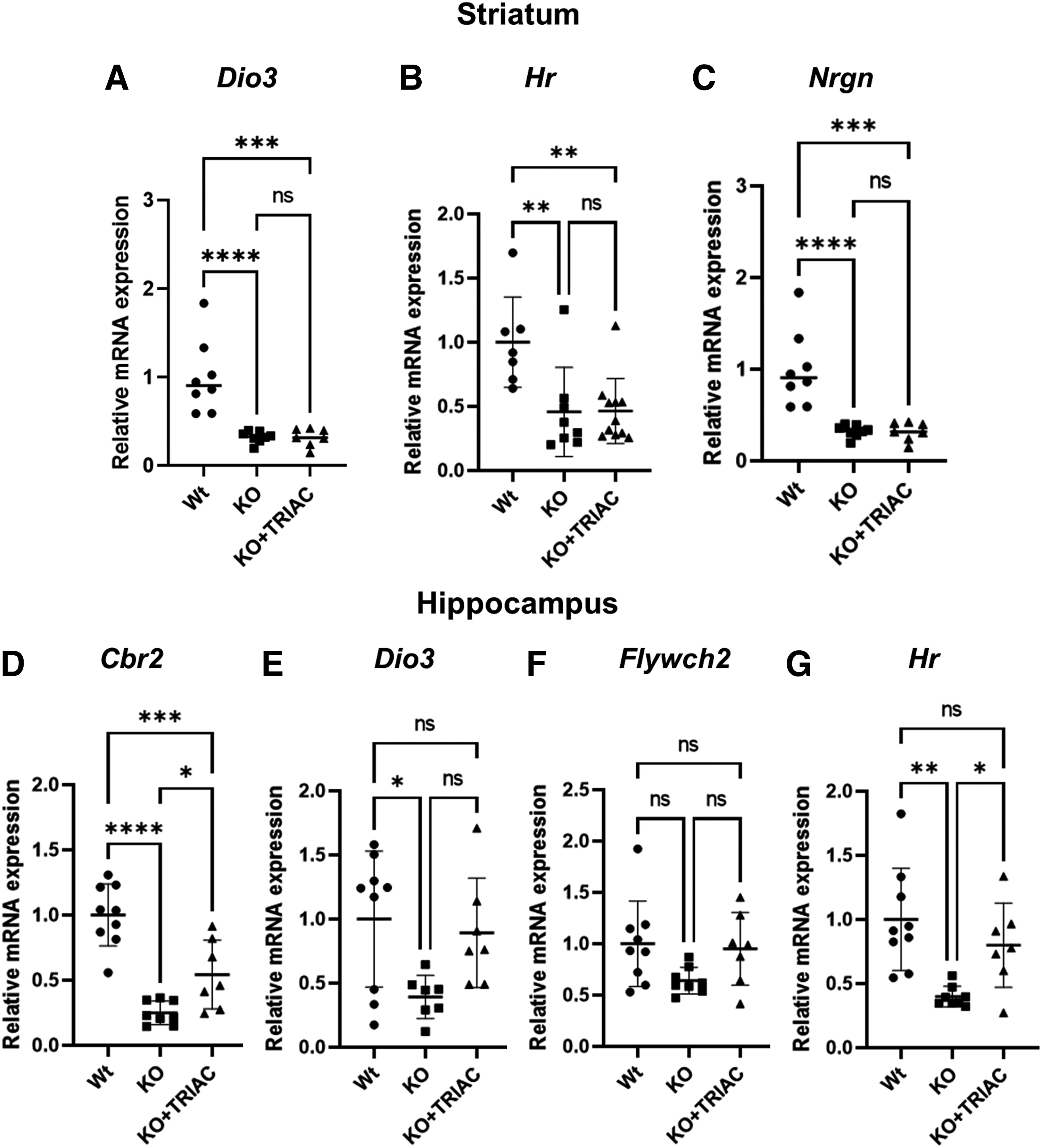
Effects of treatment on T3-regulated genes in the striatum and hippocampus. (
Nrgn expression was highly downregulated in vehicle-treated Mct8/Dio2KO mice in comparison with Wt (p < 0.0001) and its expression remained downregulated after ICV-TRIAC delivery (p = 0.0001; Fig. 5C). In the hippocampus, expression of Cbr2 was downregulated in vehicle-treated Mct8/Dio2KO mice in comparison with Wt (p < 0.0001; Fig. 5D). A twofold increase in Cbr2 was observed in ICV TRIAC-treated animals in comparison with vehicle-treated Mct8/Dio2KO mice (p = 0.0338), although the expression of this gene did not reach Wt values (Fig. 5D).
Dio3 expression was decreased in vehicle-treated Mct8/Dio2KO mice (p = 0.0243; Fig. 5E). Even though Dio3 expression in ICV TRIAC-treated Mct8/Dio2KO animals was not different from vehicle-treated Mct8/Dio2KO (p = 0.0900), it was not different from Wt values either (p = 0.8681), which might suggest mild Dio3 upregulation by ICV TRIAC treatment in the hippocampus (Fig. 5E). Expression of Flywch2 was not statistically significantly different between any of the experimental conditions (p = 0.0780; Fig. 5F). Hr was downregulated in vehicle-treated Mct8/Dio2KO mice (p = 0.0016) and its expression increased twofold as a result of ICV TRIAC treatment (p = 0.0494), reaching Wt values (Fig. 5G).
Altogether the data suggest that despite the significant increases in TRIAC content in the brain after ICV delivery, these are only able to modulate the expression of a limited number of T3-regulated genes in specific brain regions.
Discussion
In this study, we have evaluated the possibility of administrating high doses of TRIAC by ICV delivery in a mouse model of MCT8 deficiency to mediate thyromimetic actions in the brain without worsening the peripheral hyperthyroidism or brain hypothyroidism characteristic of MCT8 deficiency.
We have found that ICV administration of high doses of TRIAC increases TRIAC serum levels to the same, or an even higher, extent than when TRIAC is administered peripherally at therapeutic doses. 19 In plasma, T4 and T3 levels decreased in Mct8/Dio2KO after ICV treatment with high doses of TRIAC, which contrasts with previous observations where ICV administration of therapeutic doses of TRIAC did not affect T4 and T3 plasma levels, 20 and resembles the observations obtained after systemic treatment with therapeutic doses of TRIAC that also decreased T4 and T3 plasma levels. 19
A decrease in T3 plasma levels restored or improved the expression of several T3-regulated genes affected by peripheral hyperthyroidism in the liver and heart, with the exception of Myh7 which greatly increased after ICV TRIAC treatment. Interestingly, Myh7 is a T3-negatively regulated gene, 35 indicating that increases in its expression are probably reflecting the reduction in plasma T3 levels rather than peripheral thyromimetic effects of TRIAC. In the brain, treatment with TRIAC decreased T4 content, probably due to the decreases in plasma T4 levels; however, this event did not affect brain T3 content.
ICV administration of high doses of TRIAC significantly increased TRIAC content in all brain areas explored, indicating that this administration of TRIAC is able to distribute throughout the brain reaching remote areas from the injection site, such as the cerebellum. Despite this, increases in TRIAC content were not able to consistently modulate the expression of T3-regulated genes in all the explored regions, and only some effects were observed in the hippocampus. While this represents an improvement over the delivery of therapeutic doses of TRIAC, either systemically or ICV, neither of which modulated the expression of any T3-regulated gene, 19,20 greater effects on T3-regulated genes would be desired to improve neurological impairments in MCT8-deficient patients.
Since TRIAC has been reported to bind with the same affinity as T3 to the TR alpha and with higher affinity to the TR beta, 14 why are the increases in TRIAC content not able to consistently modulate the expression of T3-target genes? Goslings et al 38 demonstrated many years ago that TRIAC has a shorter duration of nuclear occupancy because of rapid fractional metabolism, which might explain the restricted hormonal effect of this compound.
As for why the T3-regulated genes in the hippocampus seem to be more sensitive to TRIAC treatment than other regions, this could be related to the tissue distribution of TRIAC with greater effects in areas in proximity to the injection site at the lateral ventricle, such as the hippocampus; although other possibilities such as differential accessibility of TRIAC to neural cells, or different presence of TRs isoforms in each tissue should not be discarded. Since TRIAC binds T3 receptors directly, 14 and previous studies have shown that increases in TRIAC content in the cerebral cortex of Mct8KO mice (that present increased DIO2 activity in the brain) are not able to regulate the expression of T3-target genes, 20 it is unlikely that lack of DIO2 activity in the present model might be impairing the ability of TRIAC to mediate gene expression.
Altogether the data indicate that, although ICV delivery of TRIAC at high doses seems to have a greater possibility of improving the neurological conditions in MCT8-deficient patients than ICV or systemic delivery of therapeutic doses of TRIAC, the thyromimetic effects elicited by TRIAC at juvenile stages in the brain appear to be limited. While there is evidence supporting that high doses of TRIAC can improve brain development when administered at early postnatal periods, 12,39 suggesting that TRIAC treatment might be beneficial for MCT8-deficient patients at early developmental stages, the findings from this study suggest that administration of high doses of TRIAC elicit minimal thyromimetic effects in the brain when administered at juvenile stages.
MCT8 deficiency is a highly incapacitating disease for patients and their families. Although TRIAC treatment improves the symptoms derived from peripheral hyperthyroidism in MCT8 deficiency, additional strategies such as alternative TH analogues, 36 drug delivery routes, 40 pharmacological chaperones, 41,42 and gene therapy approaches 9,43,44 should be thoroughly explored with the aim to improve neurological defects in patients.
Footnotes
Acknowledgments
TRIAC antibody for RIA determinations was kindly provided by Dr. A. Burger. The authors would like to thank Cristina Crespo for animal care and Dr. Maria Jesus Obregon for her advice on TH determination. We are grateful to Ewan McDonald for language editing and review.
Authors' Contributions
A.G.-F., S.B.-L., and C.G.-M. provided the conceptual framework for the study and designed the experiments. C.G.-M. performed surgical animal treatments and performed brain dissections. C.G.-M. and J.H.-Á. performed real time-quantitative polymerase chain reaction analyses and TH measurements. A.M.-P. performed TH measurements and provided intellectual input. C.G.-M., J.H.-Á., A.M.-P., S.B.-L., and A.G.-F. analyzed and interpreted the data. S.B.-L. wrote the original draft of the article and all authors revised and edited the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported by grants from the Ministerio de Ciencia e Innovacion (MCIN)/AEI/10.13039/501100011033 (Grant No. SAF2017-86342-R to A.G.-F.); MCIN/AEI/10.13039/501100011033/FEDER “Una manera de hacer Europa” (Grant No. PID2020-113139RB-I00 to A.G.-F.); Consejo Superior de Investigaciones Científicas (Grant No. 2020AEP044 to A.G.-F.); the Sherman Foundation (Grant No. OTR02211 to S.B.-L. and A.G.-F.); Asociación Corriendo con el Corazón por Hugo (Grant No. OTR06190 to A.G.-F.); and MCIN/EU (Grant No. IJC2020-043543-I to S.B-L).
Supplementary Material
Supplementary Methods