
Abstract
Background:
Thyrotropin receptor (TSHR) plays a central role in maintaining thyroid function and TSHR impairment causes hypothyroidism, which is often associated with metabolic disarrangement. The most common type of hypothyroidism is autoimmune disease-related and the mechanism, particularly with respect to the role of microRNAs (miRNAs), has not been delineated.
Methods:
Serum from 30 patients with subclinical hypothyroidism (SCH) and 30 healthy individuals were collected and exosomal miR-146a (exo-miR-146a) was examined, followed by extensive mechanistic investigation using various molecular and cellular experimental approaches and genetic-knockout mouse models.
Results:
Our clinical investigation showed that exo-miR-146a was systemically elevated in the serum of patients with SCH (p = 0.04) compared with healthy individuals, prompting us to investigate the biological effects of miR-146a in cells. We found that miR-146a could target and down-regulate neuron-glial antigen 2 (Ng2), with consequent down-regulation of TSHR. We next generated a thyroid-specific Ng2 knockout (Thy-Ng2−/− ) mouse model and found a significant down-regulation of TSHR in Thy-Ng2−/− mice, accompanied by the development of hypothyroidism and metabolic disorders. We further found that a decrease in NG2 resulted in decreased receptor tyrosine kinase-linked downstream signaling and down-regulation of c-Myc, consequently resulting in up-regulation of miR-142 and miR-146a in thyroid cells. Up-regulated miR-142 targeted the 3′-untranslated region (UTR) of TSHR messenger RNA (mRNA) and post-transcriptionally down-regulated TSHR, explaining the development of hypothyroidism above. Local up-regulation of miR-146a in thyroid cells augments the earlier cited processes initiated by systemically elevated miR-146a, thereby forming a feedback loop to propel the development and progression of hypothyroidism.
Conclusions:
This study has uncovered a self-augmenting molecular loop initiated by elevated exo-miR-146a to suppress TSHR through targeting and down-regulating NG2, thereby initiating and propelling the development and progression of hypothyroidism.
Introduction
Hypothyroidism is a common endocrine disorder in humans with a prevalence of 4.3% in the form of subclinical hypothyroidism (SCH) and 0.3% in the form of clinically overt hypothyroidism; SCH is defined as elevated thyrotropin (TSH) with normal thyroid hormones and often progresses into overt hypothyroidism. 1 Hypothyroidism is often associated with several metabolic features such as obesity, hyperlipidemia, and fatty liver, leading to increased risk of cardiovascular diseases (CVDs). 2 This can be explained by deficient thyroid hormone actions, including decreased basal metabolic rate and heat production, thereby affecting body weight and lipid metabolism. 3
Thyrotropin receptor (TSHR) is the master regulator of thyroid hormone production, whose deficiency is well known to cause hypothyroidism. 4,5 TSH/TSHR exerts this function by activating the cAMP/protein kinase A (PKA) cascade, which regulates several downstream molecules that play important roles in normal thyroid function, such as thyroid transcription factors TTF1 and PAX8. 4,5 Although the fact that TSHR deficiency can cause hypothyroidism has long been recognized, 6 –8 whether this is a molecular mechanism in the development of hypothyroidism has not been defined and mechanistically delineated.
Exosomes are a major form of cell-cell communication by transporting biologically active molecules, including microRNAs (miRNAs), which regulate protein expression profiles and cellular functions in target cells. 9 On entry into cells, miRNAs usually suppresses protein translation of target messenger RNAs (mRNAs). 10 MiRNAs vary quantitatively between patients and healthy individuals or in different physiological conditions, suggesting their potential role in human diseases.
An example is that serum miR-146a level was elevated in patients with SCH and positively associated with TSH level. 11 Other studies showed that serum miR-146a was systemically elevated in various autoimmune diseases other than autoimmune thyroiditis. 12,13 The biological and clinical significance of serum miR-146a or exosomal miR-146a (exo-miR-146a) in inflammatory, metabolic, and hypothyroid settings remains unknown.
Interestingly, neuron-glial antigen 2 (NG2) or chondroitin sulfate proteoglycan 4 (CSPG4), which acts as a co-activator of receptor tyrosine kinases (RTKs), 14 –17 was also linked to metabolic disorders. For example, obesity and fat disarrangement developed in mice with genetic ablation of Ng2. 18 NG2 is widely expressed in the central nervous system (CNS), 19 but a role of NG2 from the CNS system to cause metabolic disorders has been ruled out as specific knockout of Ng2 in CNS failed to produce metabolic disorder phenotypes. 18 Thus, how NG2 defect causes metabolic disorders remains elusive.
The earlier cited data raise an interesting question of whether and how hypothyroidism associated with metabolic disorders can be linked to exo-miR-146a and NG2 mechanistically. Here, we addressed this question by examining their relationship and discovered a self-augmenting molecular mechanistic loop involving these molecules to down-regulate TSHR and hence propel the development and progression of hypothyroidism and associated metabolic disorders.
Materials and Methods
Animals
Ng2flox/flox C57Bl/6 and TPO-Cre mouse strains were kindly provided by Prof. William Stallcup (Sanford Burnham Prebys Medical Discovery Institute) and Prof. Kimura Shioko (National Institutes of Health), respectively. Thyroid-specific Ng2 knockout (Thy-Ng2−/− ) mice were established by crossing Ng2 flox/flox mice with TPO-Cre mice, yielding the thyroid follicular epithelial cell (thyrocyte)-specific NG2-deficient mice. Wild-type mice (Thy-Ng2+/+ ) generated in the same batch were used as the controls. This study was approved by the Animal Ethics Committee of Xi'an Jiaotong University (No. XJTU2018-167).
Clinical samples
This study was approved by the Ethics Committee of the First Affiliated Hospital of Xi'an Jiaotong University (No. XJTU2018-167). Informed consent was acquired from all patients before they were included in the study. To evaluate the expression of NG2 and TSHR, 22 non-cancerous thyroid tissues were consecutively obtained from the First Affiliated Hospital of Xi'an Jiaotong University who underwent surgery between March 2019 and April 2019. The patients were newly diagnosed thyroid cancer patients not yet given any treatment before surgery.
To measure serum exosomal miRNAs, serum samples of 30 consecutive patients with SCH (with TSH ≥10 mU/L and normal levels of free thyroxine [fT4]) were collected from the First Affiliated Hospital of Xi'an Jiaotong University between February 2019 and September 2019. The patients were newly diagnosed SCH patients not yet given any treatment.
A total of 30 healthy patients without thyroid-related disorders or infection within the past 3 months were recruited for the control group and were matched by age and sex. The exclusion criteria include the following: (1) Pituitary disease and other endocrine diseases; (2) The presence of cancer; (3) Acute cardiovascular and cerebrovascular disease; (4) Severe infection, liver or kidney dysfunction; (5) Using medications that affect thyroid function (such as an amine iodine ketones and other iodine-containing drugs) within the previous 3 months before enrollment.
Specific inclusion and exclusion criteria are shown in Supplementary Figure S1 in the Supplementary Materials. Exosomes were isolated by ultracentrifugation, as previously described. 20 The exosome morphology was examined by a Hitachi transmission electron microscope (H-7650; Hitachi). The size distribution of exosomes was analyzed using a Malvern Zetaszier Nano ZS (Malvern Instruments Ltd., Malvern, United Kingdom), and Western blotting analysis was performed for further validation.
Additional methods are presented in Supplementary Materials.
Results
Exo-miR-146a potentially causes hypothyroidism by targeting NG2
A previous study showed that serum miR-146a level was significantly elevated in patients with SCH. 11 Here, we examined miR-146a level in serum exosomes of SCH patients and control subjects by ultracentrifugation, and we identified them using transmission electron microscopy, particle size analysis, and Western blotting (Fig. 1A–C). Exo-miR-146a was then quantitatively analyzed by real-time quantitative polymerase chain reaction, which showed that exo-miR-146a was significantly elevated in SCH patients (p = 0.04; Fig. 1D).
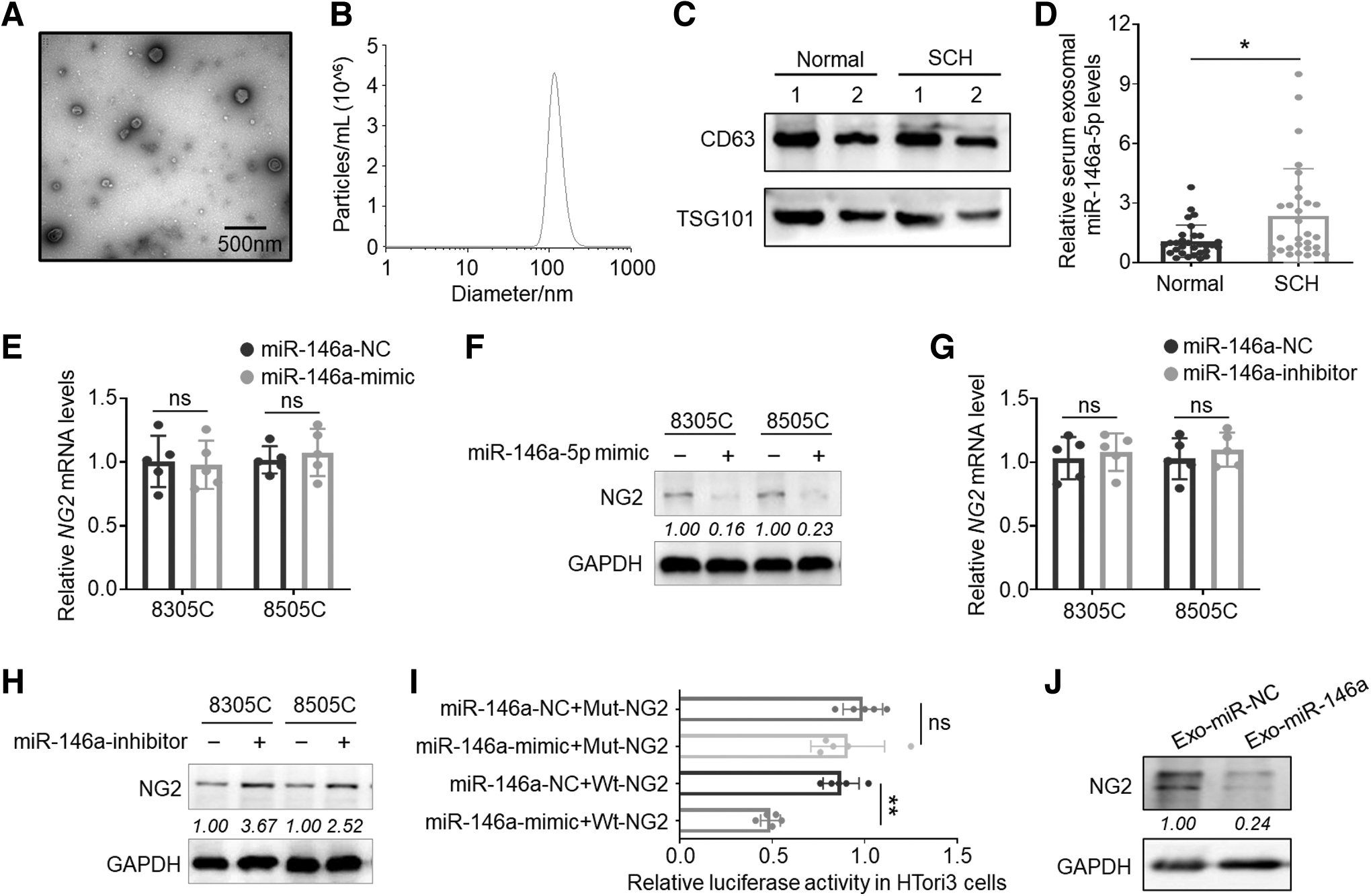
Potential role of exo-miR-146a in regulating thyroid function by targeting NG2. (
Using bioinformatic prediction tools (
We next constructed luciferase reporter plasmids containing coding sequence (CDS) of NG2 mRNA (Supplementary Fig. S2A, B in the Supplementary Materials) and demonstrated that miR-146a mimic significantly decreased luciferase activity of the wild-type (p < 0.001) but not the plasmid with binding site mutation in HTori3 cells (Fig. 1I). We also isolated exosomes from cell culture supernatants of cells treated with miR-146a mimic or the control and added them to untreated cells. The results showed that the former strongly inhibited the protein expression of NG2 compared with the control (Fig. 1J). These data, thus, functionally demonstrate that NG2 is, indeed, a target of miR-146a and the latter negatively regulates the former by affecting its mRNA translation.
As an established co-activator of RTKs, NG2 plays a key role in sustaining their activity 18 and ablation of NG2 suppressed the activity of RTKs and their downstream pathways. 14 –17 Besides, the Gene ontology/Kyoto Encyclopedia of Genes and Genomes pathway enrichment analysis using the GSE126698 dataset showed that NG2 was involved in the “MAPK signaling” and “thyroid hormone synthesis” pathways (Table 1). Based on these observations and the fact that tyrosine kinases inhibitors (TKIs) in the treatment of cancer patients commonly cause hypothyroidism, 21,22 we hypothesize that exo-miR-146a may potentially cause hypothyroidism and associated metabolic disorders by targeting NG2.
Kyoto Encyclopedia of Genes and Genomes Pathway Enrichment Analysis of NG2
NG2, neuron-glial antigen 2.
Thyroid-specific Ng2 knockout causes hypothyroidism in mice
To test the role of NG2 in regulating thyroid function, we generated a thyroid-specific Ng2 knockout mouse model (Thy-Ng2−/− ) by crossing Ng2 flox/flox mice with thyroid peroxidase (Tpo)-Cre mice (Fig. 2A). The desired genotypes were identified by the polymerase chain reaction (PCR) assay (Supplementary Fig. S3A in the Supplementary Materials), and the efficiency of Ng2 knockout was confirmed by immunohistostaining (Fig. 2B).

Thyroid-specific Ng2 KO causing hypothyroidism in mice. (
We also validated thyroid-specific Ng2 knockout by showing no significant difference in hepatic Ng2 expression between Thy-Ng2−/− and Thy-Ng2+/+ (wild-type) mice by immunohistostaining (Supplementary Fig. S3B in the Supplementary Materials).
We next examined the impact of thyroid-specific Ng2 knockout on thyroid functions. At the age of 5 weeks, mice showed no difference in serum total triiodothyronine (TT3), total tetraiodothyronine (TT4), free triiodothyronine (fT3), and fT4 (Supplementary Fig. S4A in the Supplementary Materials). At this age, there were slight histopathological abnormalities in thyroid gland of Thy-Ng2−/− mice compared with Thy-Ng2+/+ mice, such as increased colloid droplets, endoplasmic reticulum expansion, vessel dilatation, and increase in mitochondria (Supplementary Fig. S4B, C in the Supplementary Materials).
At the age of 36 weeks, TT3, TT4, fT3, and fT4 were significantly decreased while TSH was significantly increased in Thy-Ng2−/− mice compared with Thy-Ng2+/+ mice (Fig. 2C), showing the development of hypothyroidism. Moreover, TSHβ mRNA levels were significantly elevated in the pituitary gland of Thy-Ng2−/− mice (p = 0.02; Supplementary Fig. S4D in the Supplementary Materials), consistent with primary hypothyroidism-associated compensatory production of TSH.
We next assessed the levels of thyroid peroxidase antibody (TPOAb), thyroglobulin antibody (TGAb), and thyrotrophin receptor antibody (TRAb) by enzyme-linked immunosorbent assay (ELISA) assay and did not find significant difference in these antibodies between Thy-Ng2−/− mice and Thy-Ng2+/+ mice (Supplementary Fig. S4E in the Supplementary Materials), thereby ruling out immune-related factors. However, we observed the shrinkage of nuclear membrane, enlargement of perinuclear space, and dilation of endoplasmic reticulum in Thy-Ng2−/− mice (Fig. 2D), indicating the aging of thyroid follicular cells.
Patients with hypothyroidism are more likely to have uric acid and alkaline phosphatase (ALP) disorders. 23,24 We found an upward, but not significant, trend in uric acid level (p = 0.16) and a significant decrease in ALP level in Thy-Ng2−/− mice (p < 0.05) (Fig. 2E, F). Also, we found that hypothyroid Thy-Ng2−/− mice had reduced heart rate (p = 0.02) and pulse pressure (p = 0.04) (Fig. 2G, H).
Thyroid-specific Ng2 knockout causes metabolic disorders in mice
We speculated that hypothyroidism from Ng2 knockout might cause metabolic disorders as seen in humans. Indeed, we observed that body weight gain in Thy-Ng2−/− mice was faster than that in Thy-Ng2+/+ mice (Fig. 3A, B). We also found that body shape of Thy-Ng2−/− mice at the age of 36 weeks showed a significant expansion of the abdomen with an increase in abdominal circumference compared with Thy-Ng2+/+ mice (Fig. 3C–E), but not in body length (Fig. 3F, G).
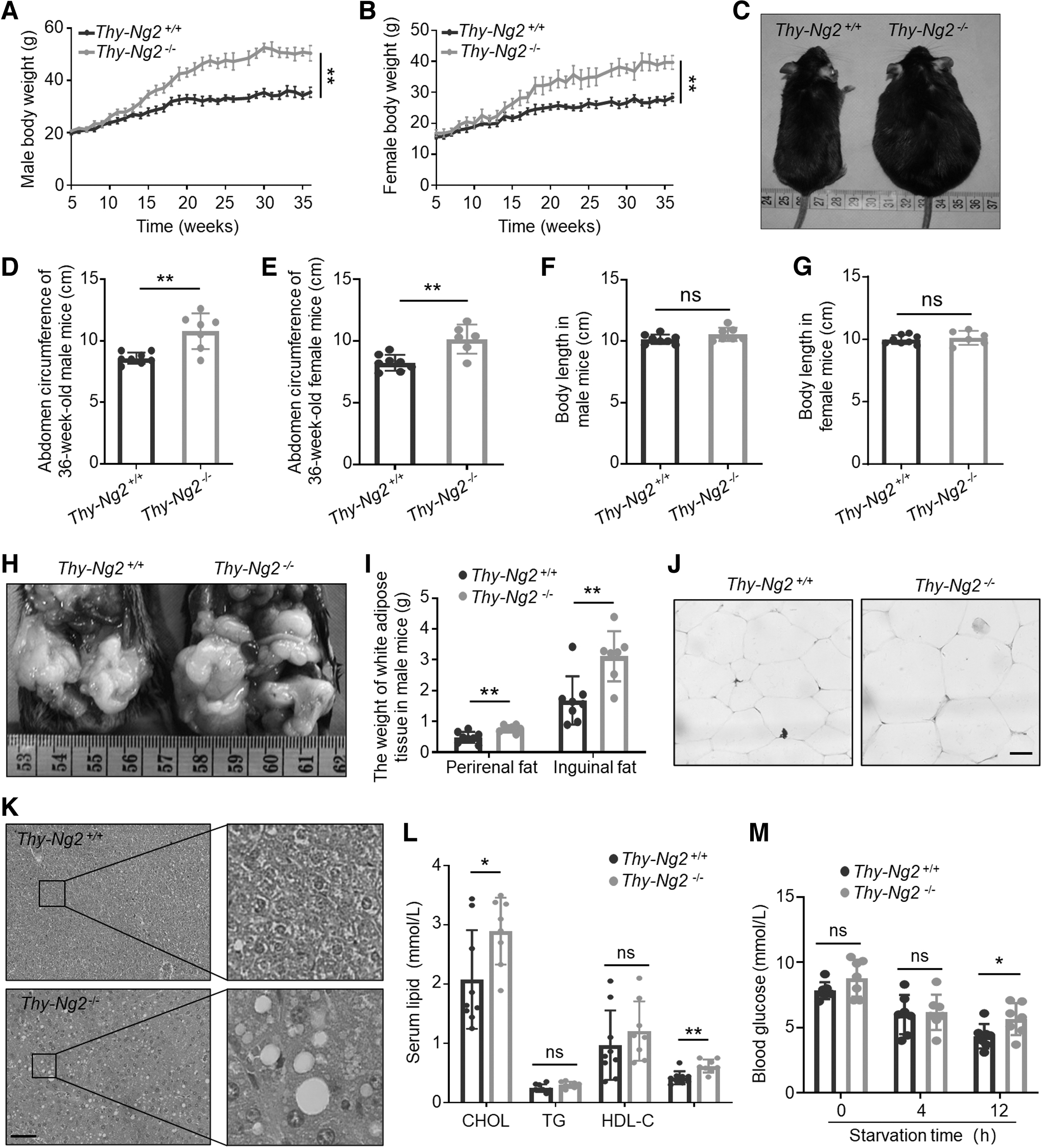
Thyroid-specific Ng2 KO causing metabolic disorders in mice. Growth curves of body weight in male (
The perirenal fat and inguinal fat were significantly increased in Thy-Ng2−/− mice compared with Thy-Ng2+/+ mice (Fig. 3H, I). We also found expansile adipocytes in Thy-Ng2−/− mice (Fig. 3J); however, mRNA levels of Ucp1 and Pgc-1α in brown adipose tissues did not significantly differ between Thy-Ng2+/+ mice and Thy-Ng2−/− mice (Supplementary Fig. S5 in the Supplementary Materials).
It is well known that patients with obesity are more likely to develop fatty liver, hyperlipidemia, and diabetes. 25,26 Here, we observed abundant vacuoles in hepatic tissue sections of Thy-Ng2−/− mice, but not in Thy-Ng2+/+ mice (Fig. 3K). Moreover, there was an increase in blood cholesterol (CHOL, p = 0.03) and low density lipoprotein (LDL, p = 0.002) in Thy-Ng2−/− mice (Fig. 3L). Thy-Ng2−/− mice also had a slight increase in fasting glucose level (Fig. 3M).
These can all be seen in human hypothyroidism 1 and are risk factors for human cardiovascular and metabolic disorders. 27 At this stage, we did not find significant difference in hepatic or renal function between Thy-Ng2−/− mice and Thy-Ng2+/+ mice (Supplementary Fig. S6A–C in the Supplementary Materials). These results demonstrate that thyroid-specific Ng2 knockout mice are predisposed to develop hypothyroidism, obesity, and metabolic abnormalities.
NG2 knockout results in the suppression of the TSH/TSHR pathway
We next examined the effect of NG2 knockout on the TSH/TSHR pathway given its critical role in maintaining normal thyroid function. 5 We found that TSHR protein level was significantly decreased but its mRNA level was virtually unchanged in Thy-Ng2−/− mice compared with Thy-Ng2+/+ mice (Fig. 4A, B). This was also supported by correlation analysis between mRNA (R = 0.03, p = 0.5) and protein (R = 0.34, p = 0.03) expression of TSHR and NG2 (Supplementary Fig. S7A, B in the Supplementary Materials).
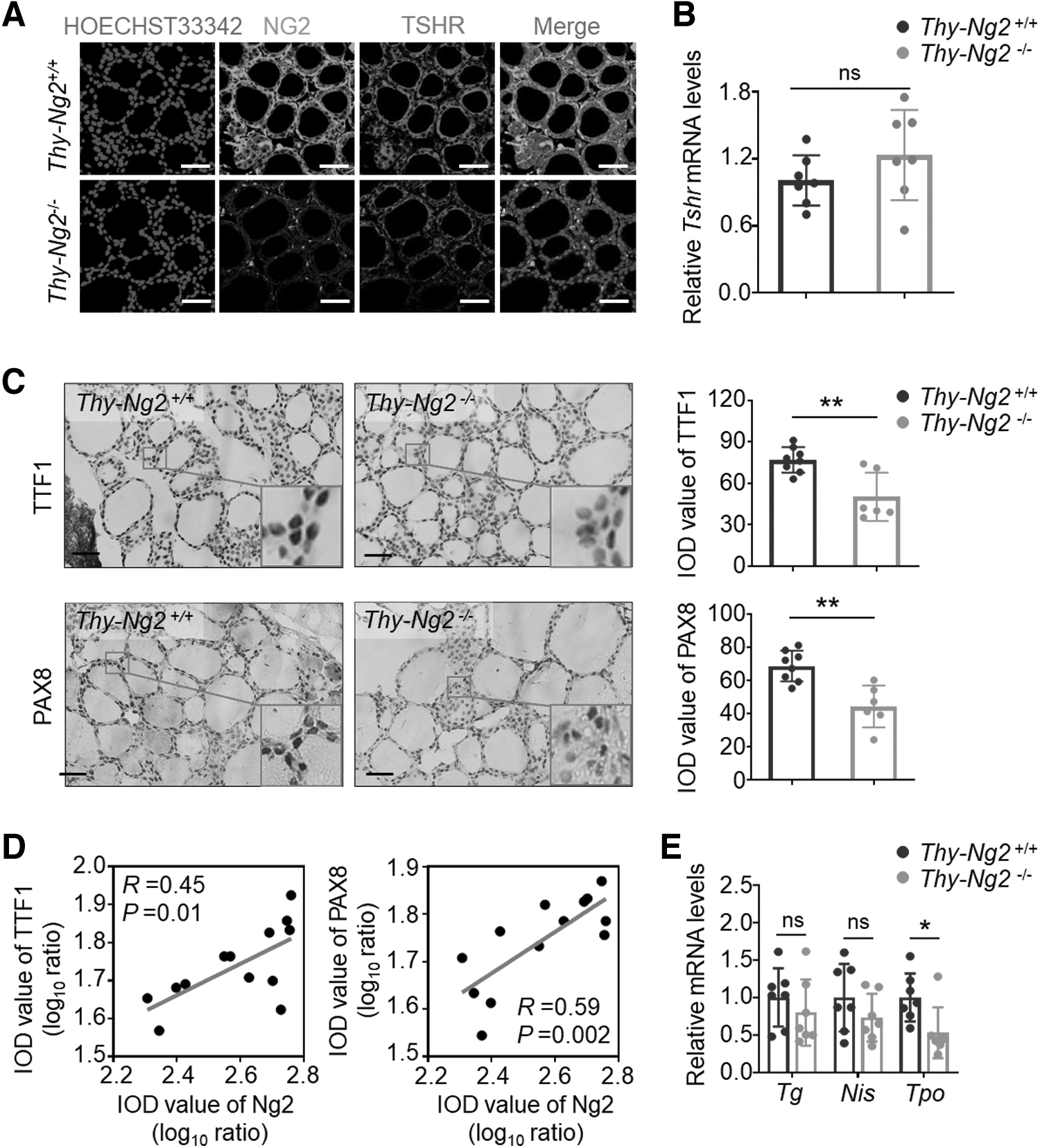
Inhibitory effect of Ng2 KO on the activity of TSH/TSHR signaling pathway. (
These results suggest that Ng2 positively regulates TSHR expression at a post-transcriptional level. Treatment with proteasome inhibitor MG132 and autophagy inhibitor chloroquine (CQ) to block proteasome-mediated degradation and autophagy-mediated lysosomal degradation failed to rescue NG2 ablation-mediated TSHR reduction (Supplementary Fig. S8 in the Supplementary Materials), suggesting that protein degradation is not a mechanism for NG2 ablation-induced TSHR reduction.
Given that the TSH/TSHR pathway regulates thyroid cellular functions by the cAMP/PKA system, 28 we examined the effect of Ng2 knockout on cAMP production and found that cAMP levels were significantly decreased in Thy-Ng2−/− mice compared with Thy-Ng2+/+ mice (Supplementary Fig. S9 in the Supplementary Materials).
As the downstream targets of the TSHR/cAMP pathway, mRNA and protein levels of thyroid-specific transcription factors Ttf1 and Pax8 were all down-regulated in Thy-Ng2−/− mice, particularly the former (Fig. 4C and Supplementary Fig. S10A in the Supplementary Materials). Besides, there was a positive association of Ng2 expression with the expression of Ttf1 (protein level: R = 0.45, p = 0.01; mRNA level: R = 0.69, p = 0.001) or Pax8 (protein level: R = 0.59, p = 0.002; mRNA level: R = 0.41, p = 0.005) (Fig. 4D and Supplementary Fig. S10B, C in the Supplementary Materials).
These data suggest that Ng2 positively regulates the expression of Ttf1 and Pax8 through the TSH/TSHR/cAMP pathway. Consistent with the well-known fact that thyroglobulin (Tg), sodium iodide symporter (Nis), and Tpo are the downstream targets of Ttf1 and Pax8, 29 their mRNA levels were all decreased in Thy-Ng2−/− mice compared with Thy-Ng2+/+ mice, particularly Tpo (p < 0.05; Fig. 4E).
Correlation analyses further supported this conclusion (Supplementary Fig. S10D–F in the Supplementary Materials). In addition, we also investigated the levels of NG2, TTF1, and PAX8 in a panel of para-cancerous thyroid tissues, and we found that the levels of TTF1 and PAX8 were relatively higher in tissues with high NG2 expression than those with low NG2 expression (Supplementary Fig. S11 in the Supplementary Materials), further supporting the conclusion. These data, taken together, indicate that thyroid-specific NG2 knockout causes hypothyroidism through down-regulating TSHR and related downstream molecules.
NG2 knockout reduces TSHR expression by blocking PI3K/AKT and MAPK/ERK pathways and down-regulating c-Myc
As a co-activator of RTKs, NG2 can regulate several major signaling pathways, including the PI3K/AKT and MAPK/ERK pathways. 30,31 As supported, our results showed that the phosphorylation of Akt and Erk and the expression of c-Myc, a major target of these pathways, 32 were strongly inhibited in thyroid tissues of Thy-Ng2−/− mice compared with Thy-Ng2+/+ mice (Fig. 5A, B). NG2 knockout in thyroid cancer cells produced results fully consistent with the earlier cited results in mice (Fig. 5C).
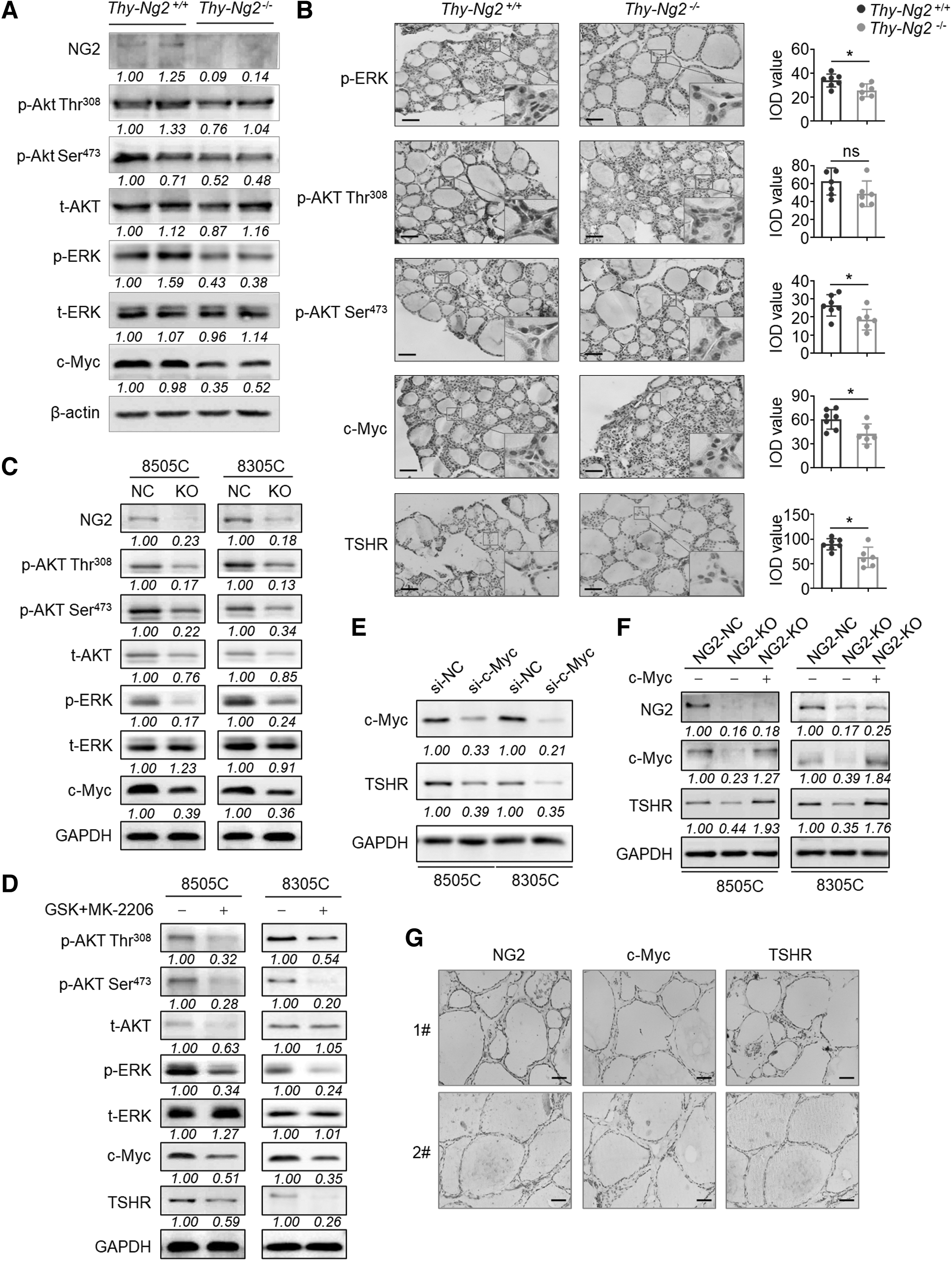
Inhibition of TSHR expression by NG2 KO via the suppression of PI3K/Akt and MAPK/Erk pathways and down-regulation of c-Myc. (
A combined treatment of MEK inhibitor GSK1120212 and AKT inhibitor MK-2206 not only suppressed the phosphorylation of AKT and ERK, but also down-regulated the expression of c-Myc and TSHR (Fig. 5D). Besides, c-Myc knockdown decreased TSHR expression (Fig. 5E), while ectopic expression of c-Myc reversed the inhibitory effect of NG2 knockout on TSHR expression (Fig. 5F).
We also found a positive correlation among the expression of NG2, c-Myc, and TSHR in a panel of para-cancerous thyroid tissues (Fig. 5G and Supplementary Fig. S12 in the Supplementary Materials), further supporting the earlier cited conclusions. As the downstream thyroid-specific transcription factors, TTF1 and PAX8 were relatively higher in NG2 high samples.
NG2 knockout post-transcriptionally down-regulates TSHR expression by the c-Myc/miR-142 signaling axis
We next determined how c-Myc mediated NG2 ablation-induced TSHR reduction. Given the role of c-Myc in repressing a wide range of miRNAs,
33
we tested a panel of miRNAs that were known to be negatively regulated by c-Myc
33
and were predicted to target the 3′-untranslated region (UTR) of TSHR mRNA using online tools (
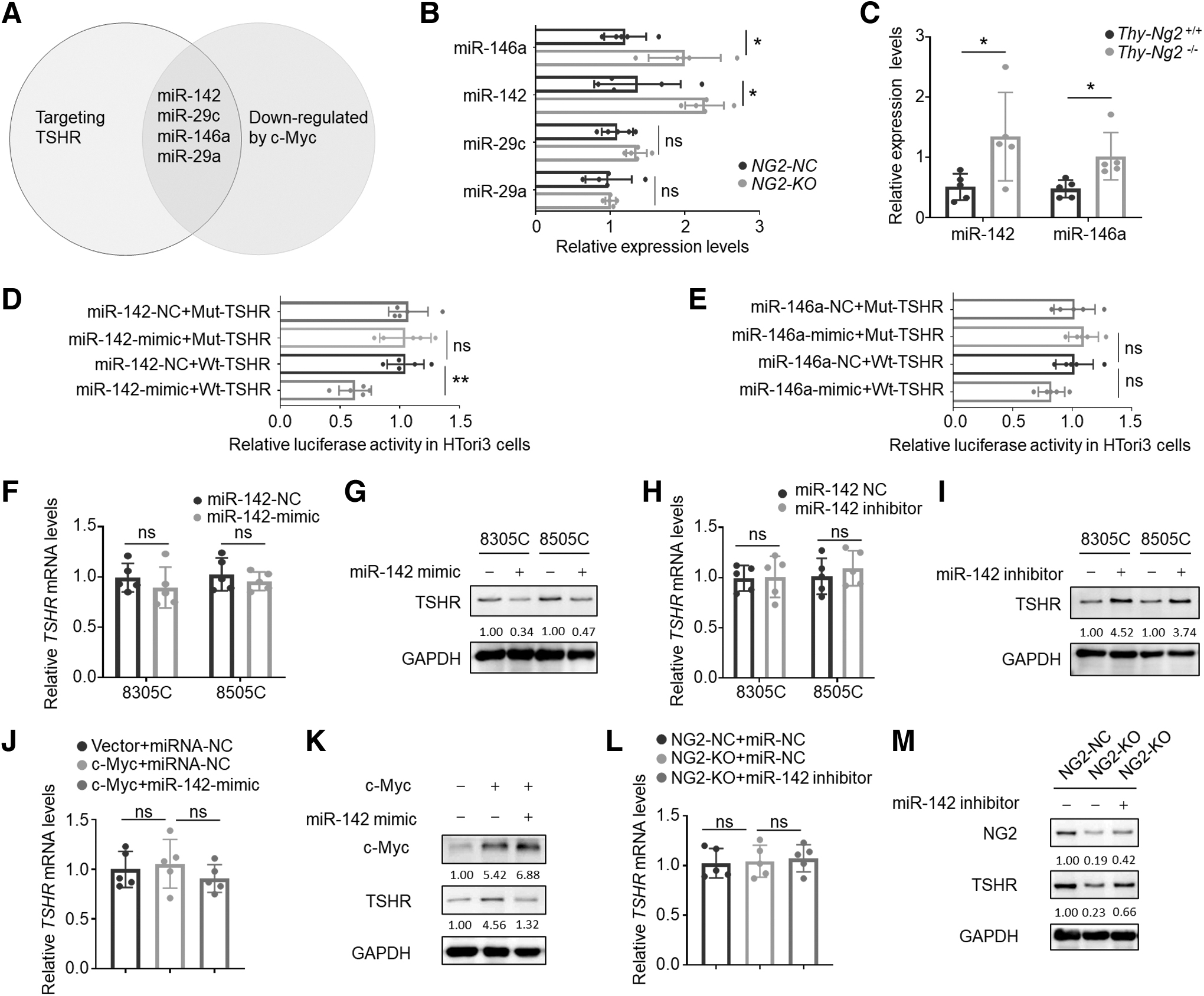
Post-transcriptional down-regulation of TSHR expression by NG2 KO via c-Myc/miR-142 signaling axis. (
The results showed that the expression of miR-142 and miR-146a was up-regulated on NG2 knockout compared with the control (Fig. 6B). This finding was confirmed in Ng2 knockout mice (Fig. 6C). In addition, to determine the effect of the levels of thyroid hormones on the expression of miR-142 and miR-146a, we treated HTori3 and 8505C cells with 10 μM Methylthiouracil, a thiourea anti-thyroid agent, and found that Methylthiouracil up-regulated their expression compared with the control (Supplementary Fig. S13 in the Supplementary Materials); however, the related molecular mechanism needs to be further investigated.
To functionally test whether these miRNAs can target TSHR mRNA, we constructed luciferase reporter plasmids containing their potential binding sites or mutated binding sites (Supplementary Fig. S14A, B in the Supplementary Materials). The results showed that miR-142 but not miR-146a mimic significantly decreased the luciferase activity in HTori3 cells transfected with wild-type plasmid, with virtually no effect on mutated one (Fig. 6D, E).
We also demonstrated that miR-142 mimic virtually had no effect on TSHR mRNA level (Fig. 6F), but strongly decreased its protein level in 8305C and 8505C cells (Fig. 6G), and vice versa (Fig. 6H, I). These results indicate that miR-142 can bind to 3′-UTR of TSHR mRNA and suppress its translation. Ectopic expression of c-Myc up-regulated TSHR protein levels but not mRNA levels, and this effect was reversed by miR-142 mimic (Fig. 6J, K).
Moreover, miR-142 inhibitor could rescue NG2 ablation-mediated down-regulation of TSHR at the protein level but not mRNA level (Fig. 6L, M). Also, we measured serum exo-miR-142 levels in SCH patients and control subjects and did not find the difference between them (Supplementary Fig. S15 in the Supplementary Materials), suggesting that, unlike miR-146a, miR-142 is not systemically changed in the blood, but changes locally in thyroid cells in the development of hypothyroidism.
Notably, miR-146a has been shown to target and down-regulate TRβ in the context of thyroid cancer. 34 Thus, we measured mRNA expression of Thrb in thyroid tissues of thyroid-specific NG2-deficient mice and control mice, and we found that the former exhibited a decreased expression of Thrb compared with the latter (p = 0.048) (Supplementary Fig. S16 in the Supplementary Materials). These data suggest that NG2 also may modulate TRβ signaling in the thyroid gland; however, more evidence needs to be provided in the future.
Discussion
In this study, we uncover a new mechanism that initiates and propels the development and progression of hypothyroidism (Fig. 7). Specifically, systemically increased exo-miR-146a in the serum of patients with autoimmune diseases targets and down-regulates NG2 in thyroid cells. Suppression of NG2 decreases the activities of PI3K/AKT and MAPK/ERK pathways and the expression of their downstream target c-Myc.
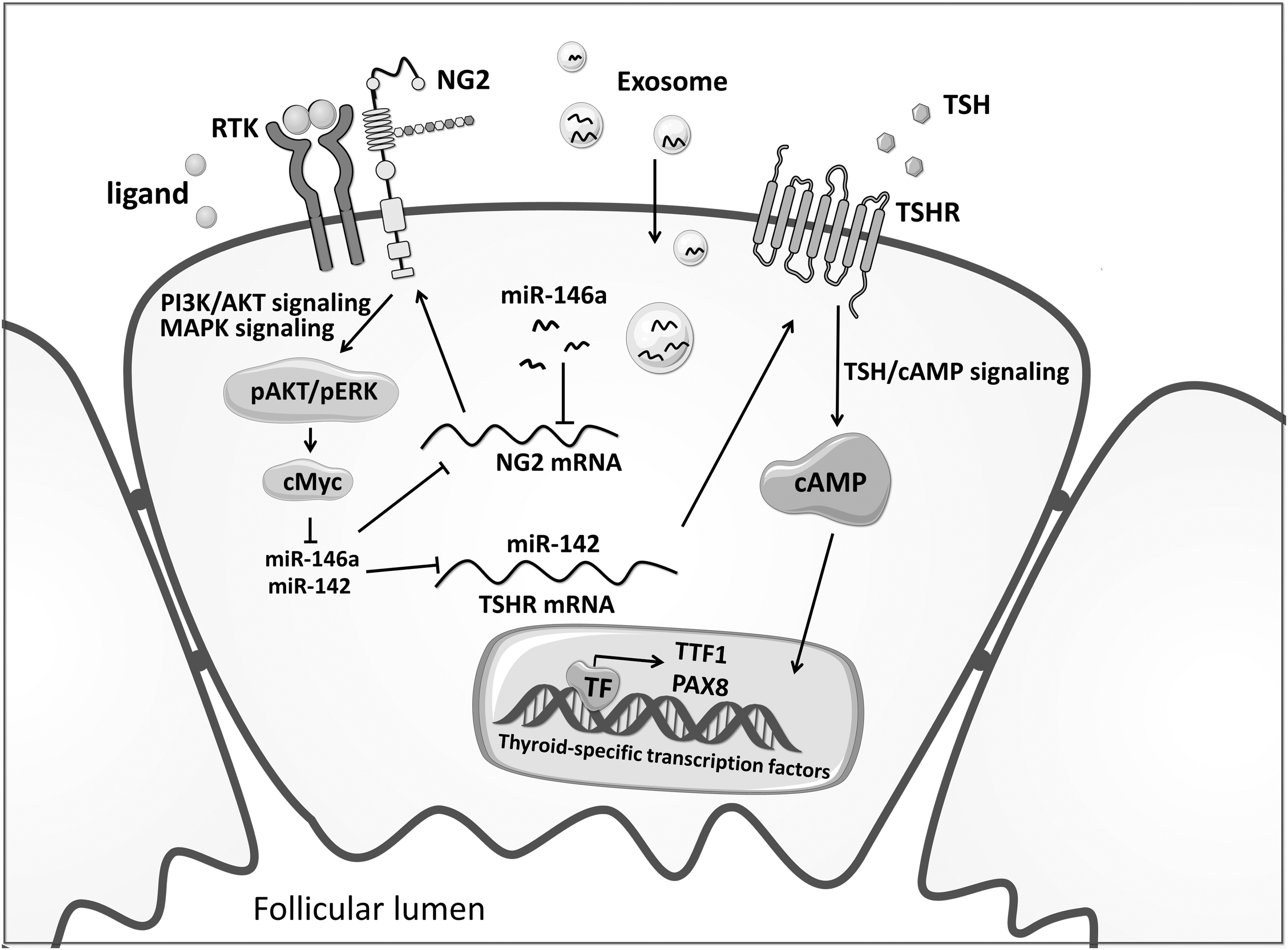
The schematic model of this study. The schematic model of the mechanism involving exo-miR-146a and NG2 to initiate and propel the development and progression of hypothyroidism.
c-Myc down-regulation leads to the elevation of miR-142 and miR-146a in thyroid cells. Increased miR-142 post-transcriptionally suppresses TSHR expression by targeting the 3′-UTR of its mRNA, thereby down-regulating the TSHR/cAMP/PKA signaling and subsequently impairing the function of thyroid follicular cells. Local elevation of miR-146a in thyroid cells caused by c-Myc down-regulation further enhances this process, forming a self-augmenting cycle loop to propel the progression of hypothyroidism.
In this study, we uncover a new mechanism that initiates and propels the development and progression of hypothyroidism (Fig. 7). Specifically, systemically increased exo-miR-146a in the serum of patients with autoimmune diseases targets and down-regulates NG2 in thyroid cells. Suppression of NG2 decreases the activities of PI3K/AKT and MAPK/ERK pathways and the expression of their downstream target c-Myc.
c-Myc down-regulation leads to the elevation of miR-142 and miR-146a in thyroid cells. Increased miR-142 post-transcriptionally suppresses TSHR expression by targeting the 3′-UTR of its mRNA, thereby down-regulating the TSHR/cAMP/PKA signaling and subsequently impairing the function of thyroid follicular cells. Local elevation of miR-146a in thyroid cells caused by c-Myc down-regulation further enhances this process, forming a self-augmenting cycle loop to propel the progression of hypothyroidism.
Besides, in the transgenic mouse model with Ng2 conditional knockout, the levels of ALP, heart rate, and pulse pressure are also basically consistent with the phenotype of hypothyroidism patients. 35 –37
An apparently plausible explanation for hypothyroidism is the physically damage of thyroid cells. Our present study provides a molecular mechanism that elevated miR-146a, both systemically in the blood stream and locally in thyroid cells, triggers and promotes TSHR down-regulation and hence the development of hypothyroidism. This mechanism may likely operate even before thyroid cells are physically impaired.
The self-augmenting molecular loop through exo-miR-146a may mechanistically contribute to the well-known progressive nature of hypothyroidism, such as the progression from SCH to overt hypothyroidism. In addition, circulating miR-146a level has shown an age-related decline in men, and this difference only existed in the group older than 75 years, compared with other age groups. 38
Aberrant miR-146a was also observed in autoimmune diseases, such as rheumatoid arthritis, autoimmune uveoretinitis, and Graves' disease. 12,13,39 A previous study provided the evidence that the level of miR-146a was higher in papillary thyroid cancers (PTCs), but not in goiter tissues, compared with heathy control. 40 These observations, taken together, suggest the important role of miR-146a in the pathogenesis and progression of human diseases, including thyroid disorders.
Hypothyroidism, even in the form of SCH, is commonly associated with metabolic disorders such as obesity, hyperlipidemia, and fatty liver, which predispose patients to CVDs. 41 This was reproduced in hypothyroidism induced by thyroid-specific Ng2 knockout in mice in the present study.
As this was thyroid-specific knockout of Ng resulting in hypothyroidism, the metabolic disorder phenotype that developed in this mouse model most likely reflected the impacts of hypothyroidism as seen in hypothyroid patients. A previous study showed that global knockout of Ng2 caused obesity and other metabolic disorders in mice. 18
Thyroid function was not evaluated in this study, but, based on the findings in our present study on thyroid-specific knockout of Ng2, it is likely that metabolic disorders found in the global Ng2-knockout animals were caused by hypothyroidism. This speculation is also supported by decreased body temperature and O2 consumption in global Ng2-knockout animals, features consistent with hypothyroidism.
In contrast, CNS-specific knockout of Ng2 did not cause metabolic disorders and probably also not hypothyroidism (thyroid condition was not examined in the study). 18 However, whether the hypothyroidism initiated and propelled by the miR-146a/NG2 feedback loop is somehow particularly associated with metabolic disorder remains to be answered.
A key mechanistic step for NG2 in the development of hypothyroidism was the NG2 loss-induced down-regulation of RTKs. Interestingly, tyrosine kinase inhibitors targeting RTKs in the treatment of cancer patients are commonly seen to cause hypothyroidism. 21 This is consistent with the finding in the present study that compromised RTKs-linked signaling as a result of miR-146a down-regulation of NG2 is a critical step in the miR-146a/NG2 loop-initiated hypothyroidism.
Exosomes serve as nanocarriers for the delivery of biologically important molecules such as miRNAs that affect disease process and may therefore be used as diagnostic or prognostic markers for disease. 42,43 An example is exosomal miR-106b, which serves as a promising marker for lung cancer. 44
By analogy, serum exo-miR-146a found in the present study may be used to help identify patients at high risk for the development of hypothyroidism. Its level may possibly be used to predict the progression risk for hypothyroidism. This is supported by the previous finding that serum miR-146a level was positively associated with TSH level and hence possibly the severity of hypothyroidism. 11
In summary, the present study identifies a new mechanism of a self-augmenting molecular loop involving exo-miR-146a and NG2 and its downstream molecules to propel the development and progression of hypothyroidism. This study may help better understand the mechanistic molecular pathogenesis in the development and progression of hypothyroidism.
However, this study has some limitations. Most important, patients who just have hypothyroidism or SCH usually do not need surgery, making it difficult to obtain clinical samples. Thus, we did not validate the role of miR-146a, NG2 and its downstream molecules in thyroid tissues of patients with hypothyroidism or SCH.
Footnotes
Acknowledgments
The authors would like to thank Profs. William Stallcup (Sanford Burnham Prebys Medical Discovery Institute) and Kimura Shioko (National Institutes of Health) for kindly providing transgenic mouse lines.
Authors' Contributions
All authors read and approved the final version of the manuscript. P.H., M.X., and H.G. conceived the projects, designed the experiments, and wrote the manuscript. F.S. and P.C. conducted the animal experiments. M.J. provided financial support and helped to design experiments. C.F. and Y.W. performed the statistical analysis. F.S. and Q.Y. performed the cytological and biological experiments. S.Z. contributed to patient samples.
Author Disclosure Statement
H.G. is an Associate Editor at Thyroid but was blinded to the peer review process. All other authors have no competing financial interests.
Funding Information
This work was supported by the National Natural Science Foundation of China (Nos. 82170803 and 82072949).
Supplementary Material
Supplementary Materials