
Abstract
Background:
Glucagon-like peptide 1 (GLP-1) is involved in the regulation of energy and glucose homeostasis. As GLP-1 has similar effects on the energy homeostasis as the hypophysiotropic thyrotropin-releasing hormone (TRH) neurons that regulate the hypothalamic–pituitary–thyroid (HPT) axis, we raised the possibility that the TRH neurons are involved in the mediation of the effects of GLP-1. Therefore, the relationship and interaction of the GLP-1 system and the TRH neurons of the hypothalamic paraventricular nucleus (PVN) were studied.
Methods:
To examine the anatomical and functional relationship of TRH neurons and the GLP-1 system in the PVN, immunocytochemistry, in situ hybridization, in vitro patch-clamp electrophysiology, metabolic phenotyping, and explant experiments were performed.
Results:
Our data demonstrate that the TRH neurons of the PVN are innervated by GLP-1 producing neurons and express the GLP-1 receptor (GLP-1R). However, not only do the GLP-1-innervated TRH neurons express GLP-1R but the receptor is also present in the axons of the hypophysiotropic TRH neurons in the blood–brain barrier free median eminence (ME) suggesting that peripherally derived GLP-1 may also influence the TRH neurons. In vitro, GLP-1 increased the firing rate of TRH neurons and depolarized them. In addition, GLP-1 directly stimulated the GABAergic input of a population of TRH neurons. Furthermore, GLP-1 inhibited the release of TRH from the hypophysiotropic axons in the ME. In vivo, peripheral GLP-1R agonist administration markedly inhibited the food intake and the energy expenditure, but had no effect on the TRH expression in the PVN and resulted in lower circulating free T4 levels.
Conclusions:
Our results indicate that GLP-1R activation has a direct stimulatory effect on TRH neurons in the PVN, but the activation of GLP-1R may also inhibit TRH neurons by facilitating their inhibitory inputs or by inhibiting the axon terminals of these cells in the ME. The innervation of TRH neurons by GLP-1 neurons suggests that TRH neurons might be influenced by both circulating GLP-1 and by GLP-1 neurons of the nucleus tractus solitarii. The lack of GLP-1R agonist-induced regulation of TRH neurons in vivo suggests that the HPT axis does not mediate the GLP-1R agonist-induced weight loss.
Introduction
Glucagon-like peptide-1 (GLP-1) is primarily synthesized by the neuroendocrine L cells of the intestinal mucosa and by a neuronal group in the nucleus tractus solitarii (NTS). 1 GLP-1 decreases circulating glucose levels, has a potent inhibitory effect on food intake, and induces body weight loss. 2,3 Long-acting GLP-1 receptor (GLP-1R) agonists are used for the treatment of diabetes and obesity. 4 These GLP-1R agonists cause a marked decrease of food intake within the first 24 hours, which effect gradually wanes during chronic treatment. However, their effect on body weight is maintained. GLP-1R agonists also prevent the diet-induced compensatory decrease of energy expenditure, 5 indicating a role of GLP-1 signaling in the regulation of energy expenditure. The mechanism of this effect of GLP-1 signaling is not yet understood.
The hypothalamic–pituitary–thyroid (HPT) axis plays a critical role in the regulation of energy expenditure 6 by affecting the basal metabolic rate, 7 and therefore may mediate the effect of GLP-1 agonists on the energy expenditure. The primary central regulators of the HPT axis are the hypophysiotropic thyrotropin-releasing hormone (TRH)-synthesizing neurons of the hypothalamic paraventricular nucleus (PVN). 6 The TRH neurons integrate the neuronal and humoral inputs and transmit this information toward the peripheral parts of the HPT axis. 6
Our earlier studies demonstrated the presence of GLP-1R in neuronal elements of the PVN and also in the hypophysiotropic axons in the median eminence (ME). 8 Therefore, we hypothesized that the hypophysiotropic TRH neurons could be targeted by GLP-1 signaling. As GLP-1R signaling increases cyclic adenosine monophosphate (cAMP) production, 9 and cAMP stimulates the synthesis of TRH, 6 we raised the possibility that activation of the HPT axis could mediate the effects of GLP-1 on energy expenditure. To test this hypothesis, we explored the anatomical relationship between the GLP-1 system and TRH neurons and studied the effect of GLP-1 signaling on components of the HPT axis.
Materials and Methods
Animals
Adult male TRH-IRES-tdTomato mice, 10 FVB/AntJ mice weighing 25–35 g, were used for the anatomical and electrophysiological investigations. Adult male Wistar rats weighing 250 g from AnimaLab Hungary Kft. (Budapest, Hungary) were used for ME explant experiments. Adult male CD1 mice weighing ∼35 g were used for the in vivo experiments. Animals were housed under standard environmental conditions (12-hour light/dark cycle, lights on at 06.00 hours, temperature 22°C ± 1°C, food and water ad libitum). The tissue preparation for immunocytochemistry is described in the Supplementary Materials and Methods.
Ethical statement
All experimental protocols were reviewed and approved by the Animal Welfare Committees at the Institute of Experimental Medicine and the Animal Health and Food Control Station, Budapest (PE/EA/100-2/2021).
Morphological studies
Double- and triple-label immunocytochemistry was performed on sections of TRH-IRES-tdTomato mice to detect the GLP-1-immunoreactive (IR) innervation and GLP-1R content of TRH neurons and for the simultaneous detection of GLP-1 innervation and GLP-1R content of TRH neurons. Double-label immunoelectron microscopy was performed to examine the GLP-1-IR innervation of TRH neurons in the PVN and the relationship of GLP-1R-IR structures and the TRH neurons in the PVN and ME. The coexpression of Glp1r and Trh mRNAs in the PVN was studied using double-labeling in situ hybridization. The methods of these morphological studies are described in the Supplementary Materials and Methods.
In vitro whole-cell patch-clamp electrophysiology
Whole-cell patch-clamp electrophysiology was performed on slices of TRH-IRES-tdTomato mice. The TRH neurons were identified based on the red fluorescence of tdTomato expressed exclusively in the TRH expressing cells. The effects of the long-acting GLP-1R agonist, Exendin-4 (Ex4), were studied on the firing and membrane potential of TRH neurons and on the excitatory and inhibitory inputs of these cells.
The details of the experimental procedure are described in the Supplementary Materials and Methods.
Determination of the effect of GLP-1 signaling on the regulation of TRH release from the ME
To understand the effects of GLP-1 signaling on the TRH release of the ME, explant experiment was performed as described in detail in the Supplementary Materials and Methods.
Examination of the effect of liraglutide treatment on the HPT axis
To investigate the short-term effects of liraglutide, after habituation to the metabolic cages, two groups of mice (N = 8/group) were treated peripherally via intraperitoneal injection of vehicle or liraglutide diluted in vehicle in 0.4 μg/BWg dose and sacrificed 24 hours after treatment. The body composition was determined by an EchoMRI whole-body magnetic resonance analyzer (Zinsser Analytic, Germany). After the liraglutide injection, the metabolic parameters were monitored with the TSE Phenomaster system. The experiments are described in detail in the Supplementary Materials and Methods.
Results
GLP-1-containing axons innervate TRH neurons in the PVN
To determine the relationship of the GLP-1-containing axons and TRH neurons, double-label immunocytochemistry was performed for GLP-1 and tdTomato, since the TRH-IRES-tdTomato mice express the red fluorescent protein tdTomato marker specifically in the TRH expressing cells. 10 The dense network of GLP-1-IR fibers was observed around the tdTomato-IR TRH neurons in all anteroposterior levels of the PVN. GLP-1-IR axons were frequently in close association with the TRH neurons, suggesting GLP-1-containing innervation of TRH neurons in the entire PVN (Fig. 1A, B). Quantitative analysis revealed that almost half (46.80% ± 6.60%) of the TRH neurons were in contact with GLP-1-IR boutons. The average number of GLP-1-IR boutons on the contacted TRH neurons was 1.51 ± 0.15.
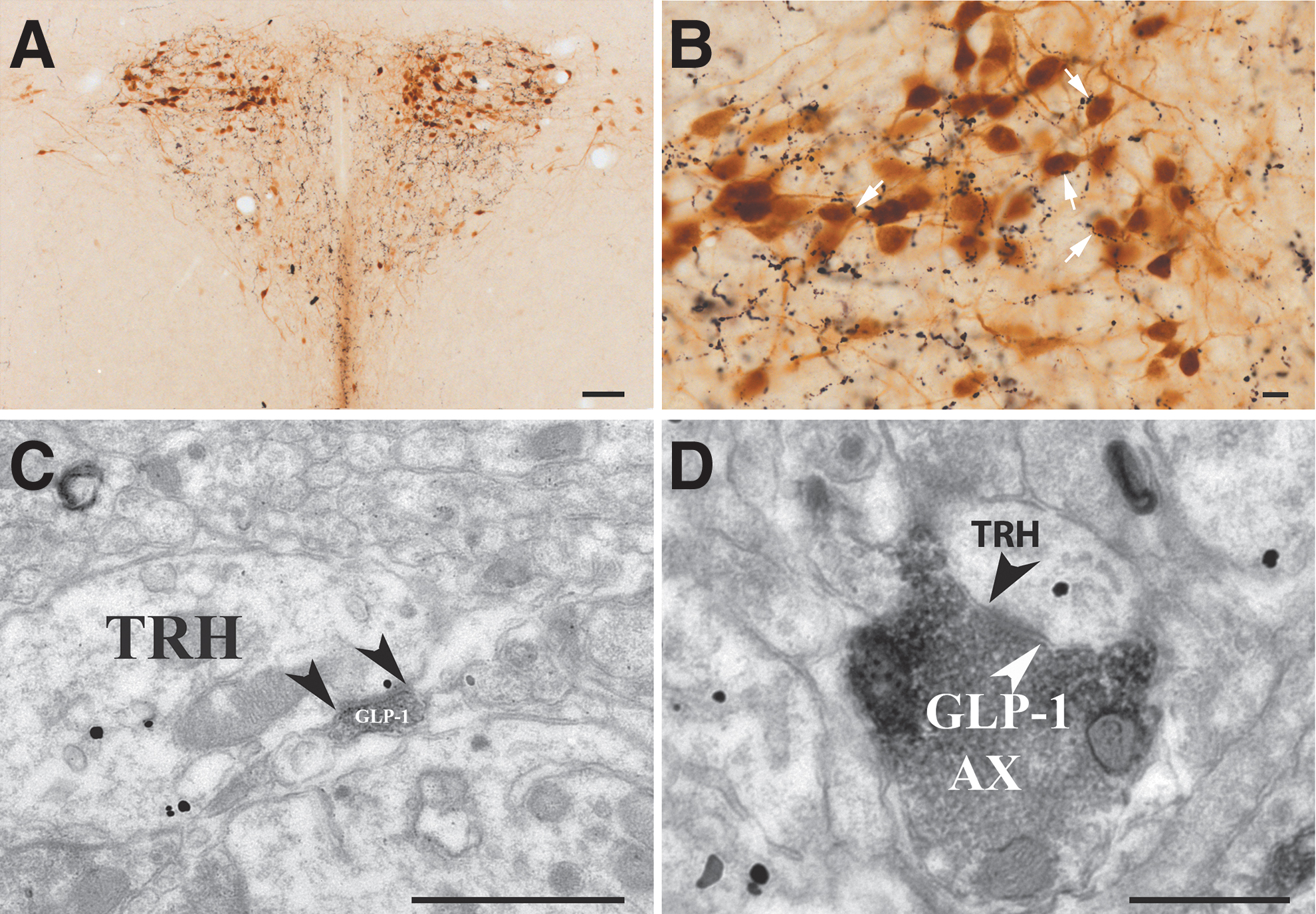
The TRH neurons of the PVN receive GLP-1 containing innervation. (
At the ultrastructural level (Fig. 1C, D), GLP-1-IR axons were observed to form asymmetric type synapses, suggesting that GLP-1-containing inputs convey an excitatory effect to the TRH neurons.
GLP-1R immunoreactivity is associated with the perikarya of TRH neurons and with axons innervating TRH neurons in the PVN
To reveal whether GLP-1R is present in the TRH neurons that are innervated by GLP-1-containing axons, triple-label immunocytochemistry was performed for GLP-1, GLP-1R, and tdTomato. GLP-1R-immunoreactivity was observed both intracellularly and on the surface of a population of TRH neurons in the PVN (Fig. 2A–C). Some of the TRH neurons contained GLP-1R-immunoreactivity and were also contacted by GLP-1-IR axons, but GLP-1R-IR TRH neurons were also observed without GLP-1-containing axons in their vicinity (Fig. 2A–C).
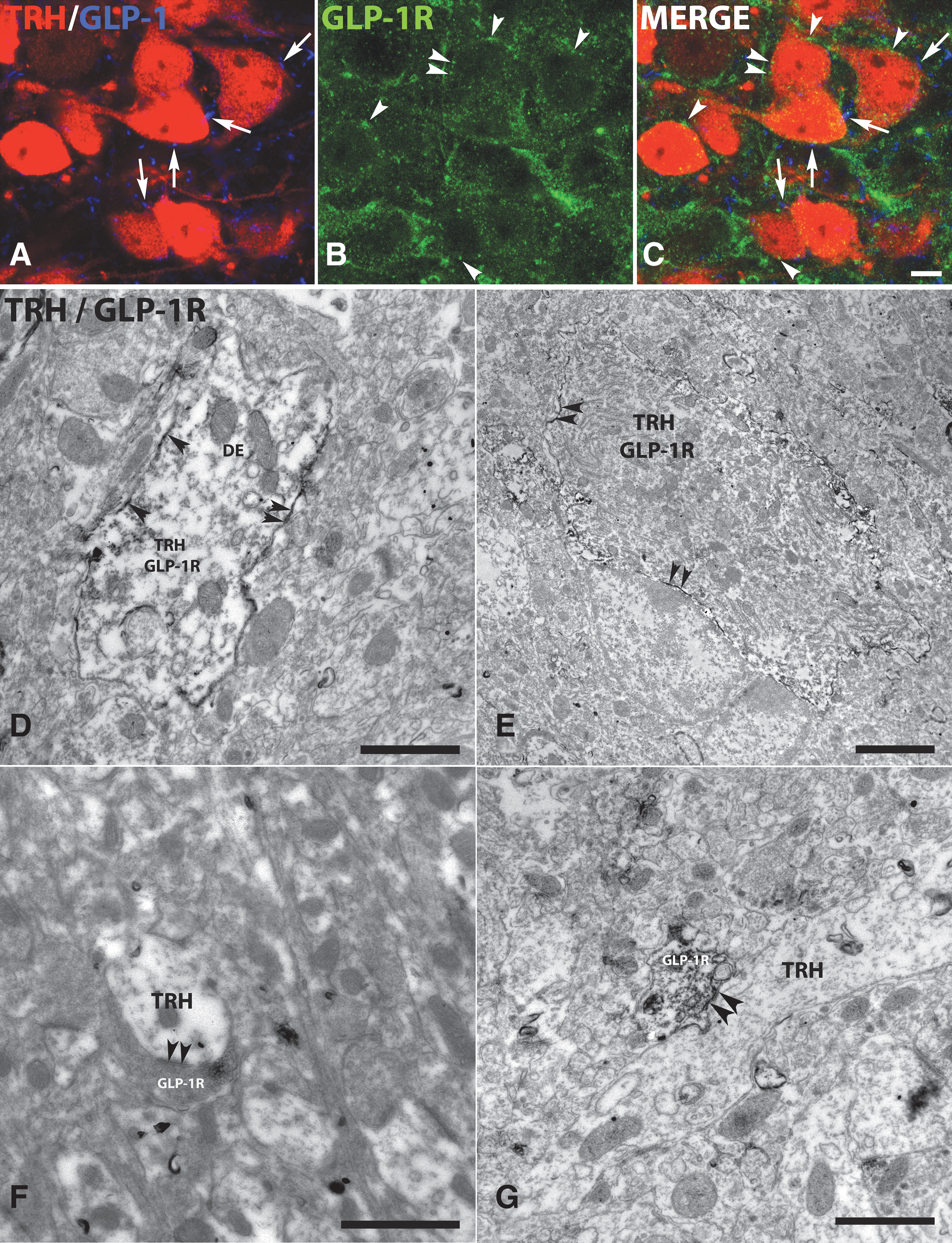
GLP-1R immunoreactivity is present on the surface of TRH neurons and on axons innervating the TRH neurons in the PVN. Triple-labeled immunofluorescent preparation (
Ultrastructural examinations of double-labeled preparations revealed that GLP-1R-immunoreactivity was associated with the outer cell membrane of dendrites (Fig. 2D) and perikarya of TRH neurons (Fig. 2E). GLP-1R-immunoreactivity was also observed in association with axons forming symmetric type synapses on the TRH neurons (Fig. 2F, G). In these axons, GLP-1R-immunoreactivity was associated with the outer cell membrane and with synaptic vesicles.
Trh-mRNA-containing neurons in the PVN express Glp1r mRNA
Double-label fluorescent in situ hybridization for Trh and Glp1r mRNAs revealed that Glp1r mRNA expression was variable among PVN neurons, from very low expression, represented by 1–3 distinct fluorescent “dots” per cell, to very high expression where the fluorescent signal of densely packed dots virtually filled the cytoplasm (Fig. 3). In the midlevel of the PVN (Allen Reference Atlas levels 60–62) where hypophysiotropic TRH neurons are highly concentrated, 11 46.9% ± 1.2% of the Trh neurons were positive for Glp1r mRNA (n = 3 brains, 244 ± 20 Trh neurons counted per brain). Virtually all Glp1r-positive Trh neurons in the mid PVN had low- to medium-level Glp1r hybridization signal (Fig. 3A1–3).
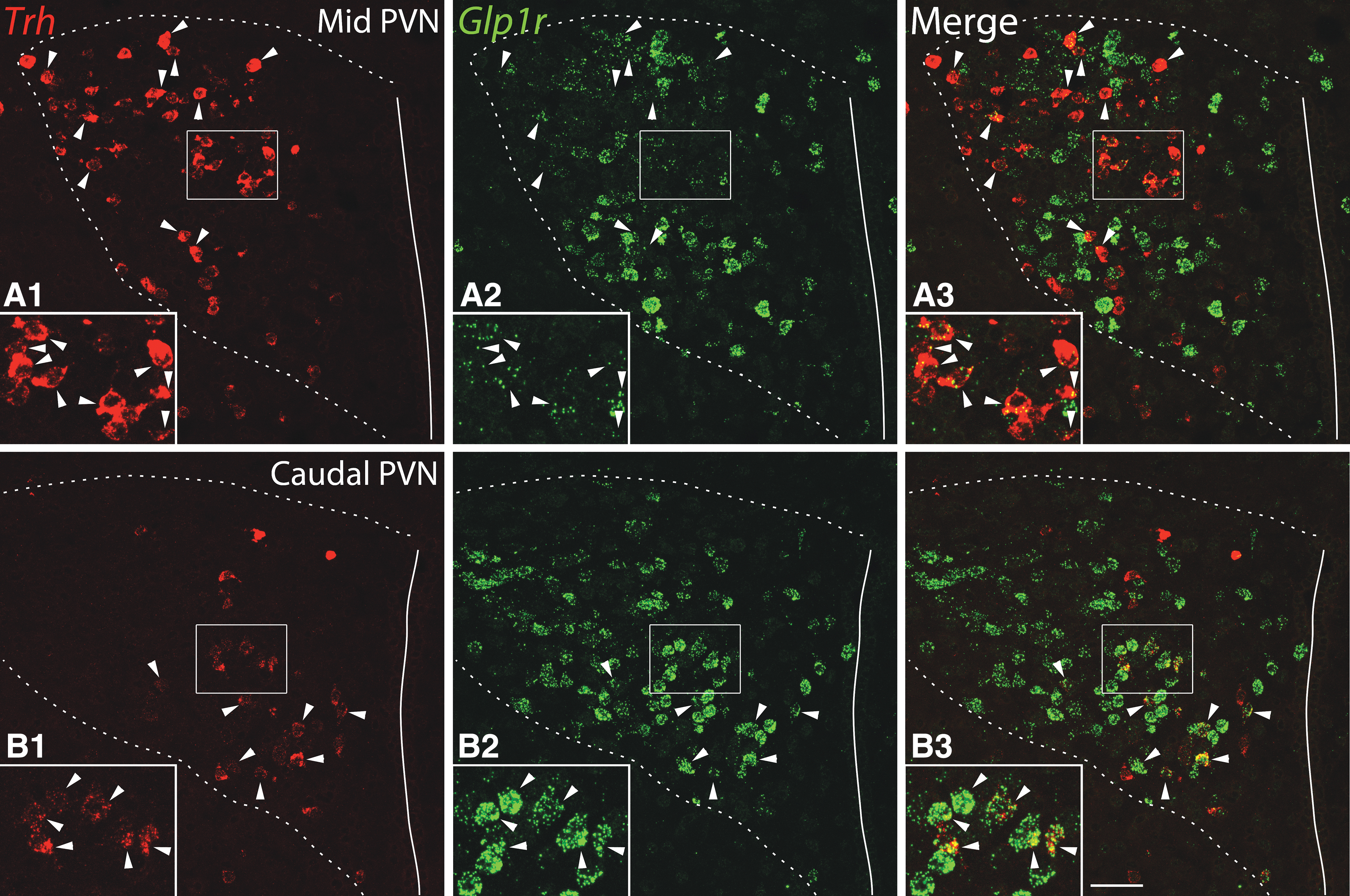
Expression of Glp1r mRNA in the TRH neurons of the PVN. (
In the caudal PVN (Allen Reference Atlas levels 63–66, as defined by Biag et al. 12 ), we observed a subpopulation of Trh neurons with very high Glp1r hybridization signal in the ventral zone of the medial parvicellular part (Fig. 3B1–3). This subnucleus is known to contain preautonomic neurons projecting to the medulla and spinal cord. 12
GLP-1 signaling stimulates TRH neurons and their GABAergic inputs in the PVN
To understand the effects of GLP-1 signaling on TRH neurons, the effects of GLP-1R agonist exendin-4 (Ex4) were studied using patch-clamp electrophysiology. Ex4 (1 μM) significantly increased the firing rate of all examined TRH neurons (242.41% ± 21.93%; p < 0.001, N = 30) (Fig. 4) and depolarized their membrane potential (+3.05 ± 1.2 mV; p < 0.001, N = 30). The Ex4-induced depolarization of TRH neurons was also observed in the presence of tetrodotoxin (TTX; inhibitor of voltage-gated sodium channels 13 ) in the artificial cerebrospinal fluid (aCSF) (+2.25 ± 1.3 mV; p = 0.004, N = 6), indicating a direct excitatory effect of GLP-1 signaling on the TRH neurons of the PVN. Ex4 treatment had no effect on the amplitude of the action potentials.
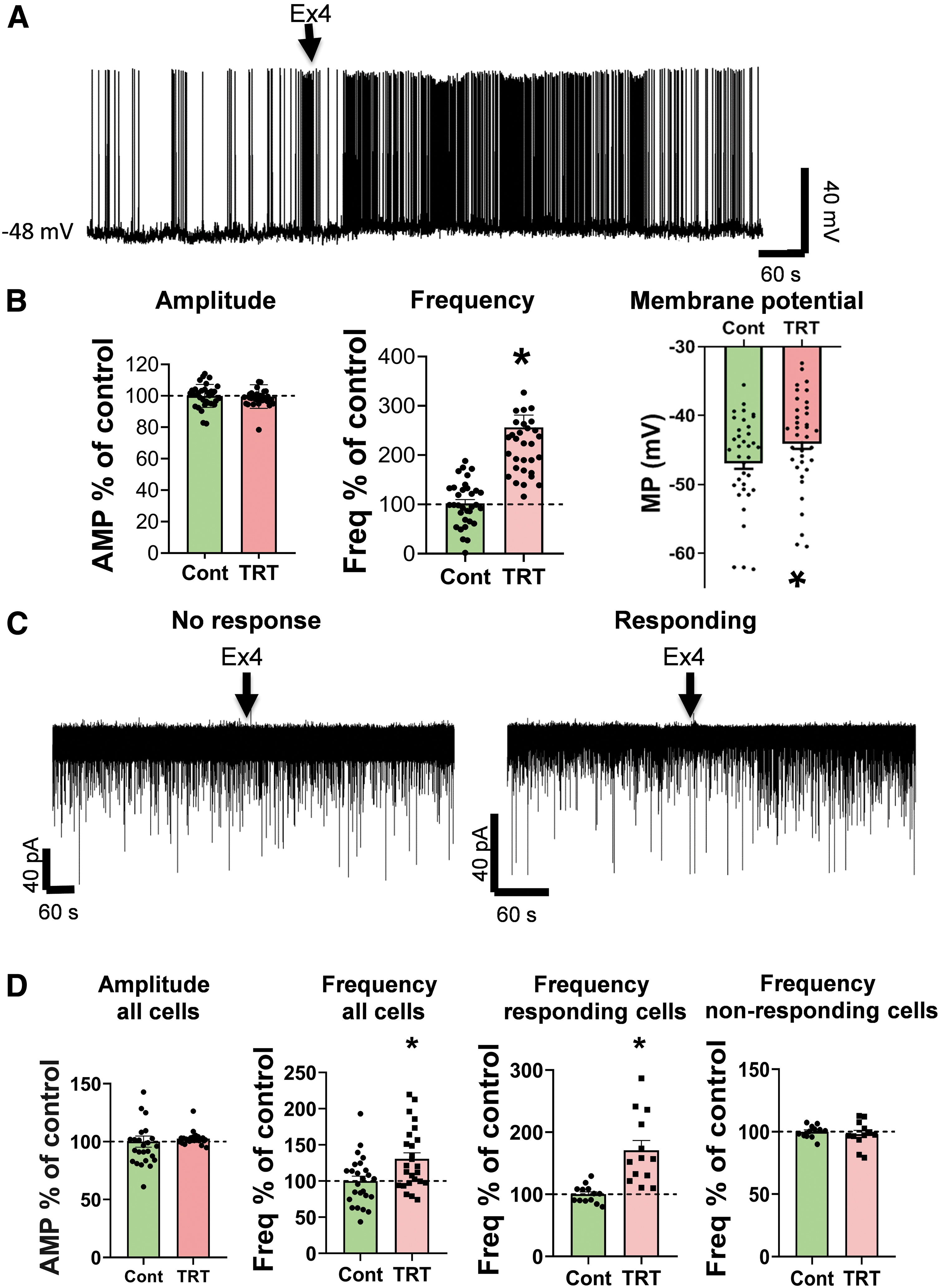
Effects of GLP-1 analog Ex4 (1 μM) on the spontaneous firing and mIPSC of the TRH neurons in the PVN of mice. (
The presence of GLP-1R in inhibitory axon terminals innervating TRH neurons suggested that in addition to its direct effect on TRH neurons, GLP1-R signaling may also influence these cells by acting on the GLP-1R-containing inputs. Therefore, we studied the effects of GLP-1R signaling on the miniature postsynaptic currents (mPSCs) of TRH neurons in the PVN. Ex4 (1 μM) increased the frequency of miniature inhibitory postsynaptic currents (mIPSCs) of TRH neurons (130.71% ± 8.44%; p = 0.007; N = 25) (Fig. 4); while it had no effect on other mIPSC parameters (amplitude, half-width, rise tau, rise time). This effect of Ex4-treatment was not uniformly observed on all TRH neurons in the PVN. In approximately half of the TRH neurons (52%, 13 out of 25 studied neurons), the Ex4 treatment increased the frequency of mIPSCs (170.92% ± 15.61% of control values; p = 0.001).
In contrast, in the remaining TRH neurons (48%, 12 out of 25 studied neurons), the Ex4 treatment had no effect on the mIPSC frequency (98.06% ± 2.99%; p = 0.541). This effect of Ex4 on the mIPSC frequency of TRH neurons was observed in the presence of G protein inhibitor, GDP-β-S, in the intracellular solution indicating that GLP-1 signaling has a direct facilitatory effect on the inhibitory inputs of approximately half of the TRH neurons in the PVN.
In contrast, Ex4 (1 μM) treatment had no effect on the miniature excitatory postsynaptic currents (mEPSC) frequency of the TRH neurons (94.57 ± 10.19; p = 0.30; N = 12).
GLP-1R is present in hypophysiotropic TRH axons in the external zone of the ME
Earlier, we observed a large number of GLP-1R-IR hypophysiotropic axons in the external zone of the ME in rats, 8 where the axons of the hypophysiotropic TRH neurons terminate. 6 Therefore, we studied whether GLP-1R is present in the axons of hypophysiotropic TRH neurons in the external zone of ME in mice using double-label immunofluorescence. GLP-1R-immunoreactivity was observed in a large proportion of the axon terminals of TRH neurons in this location (Fig. 5A). At the ultrastructural level, strong GLP-1R-immunoreactivity was associated to the surface of the TRH axons in the ME (Fig. 5B, C).
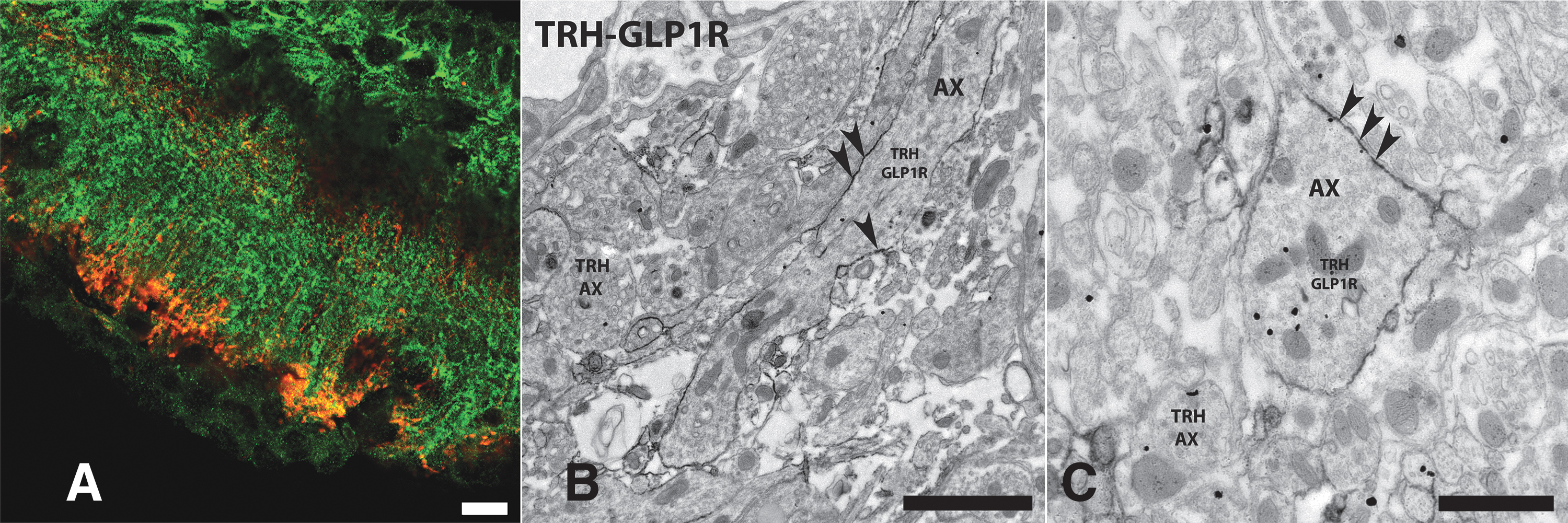
Presence of GLP-1R-immunoreactivity on the surface of TRH axons in the external zone of the ME. (
GLP-1R signaling inhibits the TRH release from the ME
To determine, how circulating GLP-1 or GLP-1 agonists influence the TRH release from the hypophysiotropic axon terminals, ME explants were used. The ME explants were prepared from Wistar rats, as in preliminary experiments, the weight of mouse ME explants was under the detection limit. The ME explants were treated with Ex4 (0.5 μM) in the presence of TRH-degrading enzyme inhibitor. During the first 10 minutes of incubation, Ex4 treatment did not influence the TRH release (control vs. Ex4 ng/mL/ng tissue: 23.22 ± 4.72 vs. 22.39 ± 7.56; p = 0.93). However, the Ex4 treatment significantly decreased the TRH release during the second 10 minutes of treatment (control vs. Ex4 ng/mL/ng tissue: 38.31 ± 9.68 vs. 14.15 ± 3.18; p = 0.028).
Effects of peripheral liraglutide treatment on the HPT axis
Intraperitonial injection of the long-acting GLP-1R agonist 14 liraglutide to male CD1 mice had a potent effect on the parameters of energy homeostasis in the first 24 hours after the treatment and markedly inhibited the Npy gene expression in the arcuate nucleus (ARC) (Supplementary Results), but had no effect on the TRH mRNA level in the PVN (Fig. 6). The integrated density values of TRH mRNA in the PVN of the control and liraglutide-treated groups were similar 24 hours after the treatment (control vs. liraglutide: 504.34 ± 34.25 vs. 521.15 ± 56.18; p = 0.80). Liraglutide treatment, however, significantly decreased the free thyroxine (fT4) levels (control vs. liraglutide [ng/dL]: 2.05 ± 0.06 vs. 1.77 ± 0.04; p = 0.0027) but did not influence the free triiodothyronine (fT3) levels (control vs. liraglutide [ng/dL]: 5.38 ± 0.53 vs. 5.12 ± 0.32; p = 0.64).

Influence of liraglutide treatment on the HPT axis. Peripheral liraglutide treatment had no effect on the Trh mRNA level of the PVN (
Thyrotropin (TSH)β mRNA level in the pituitary had a tendency for decrease in liraglutide-treated mice that did not reach the level of significance (control vs. liraglutide [relative quotient]: 1 ± 0.08 vs. 0.85 ± 0.07; p = 0.18) (Fig. 6).
Discussion
Our data demonstrated that approximately half of the TRH neurons in the PVN are innervated by GLP-1-synthesizing neurons of the brain stem. In accordance with the glutamatergic phenotype of the GLP-1 neurons, the GLP-1-containing axons established asymmetric type synapses on the surface of TRH neurons indicating the excitatory nature of these synapses. 15
Approximately 47% of Trh neurons in the neuroendocrine midportion of the PVN were observed to express Glp1r mRNA. This is almost certainly an underestimate, however, as in many cases the Glp1r hybridization signal resulted in only 1–3 distinct dots in a small part of the cytoplasm, but the thin (16 μm) tissue sections rarely contained the entire cell bodies of Trh neurons. Trh neurons in the preautonomic part of the caudal PVN, corresponding to a group of previously described nonhypophysiotropic TRH neurons, 11 also expressed Glp1r mRNA. In most cases, these nonhypophysiotropic TRH neurons were more intensely labeled with the GLP-1R hybridization signal than the hypophysiotropic TRH neurons. This ventral parvocellular subdivision of the PVN is known to project to autonomic centers of the medulla and the spinal cord. 12
GLP-1R-immunoreactivity was observed in association with the surface of TRH neurons. Intriguingly, GLP-1R was observed not only in TRH neurons that were contacted by GLP-1-containing axons, but also in TRH neurons without GLP-1-containing inputs. This would suggest that either GLP-1 of neuronal origin can diffuse in the PVN by volume transmission, or peripheral GLP-1 can access TRH neurons in the PVN.
GLP-1R was also observed in axons forming symmetric type synapses on the surface of TRH neurons, raising the possibility that GLP-1 signaling also affects TRH neurons indirectly by regulating the activity of the inhibitory inputs of these cells.
The presence of GLP-1R on the TRH neurons of the PVN is also supported by the Ex4-induced depolarization and increased firing of these neurons, and by the inability of TTX to prevent the Ex4-induced depolarization.
In accordance with the presence of GLP-1R in axons establishing symmetric type synapses with TRH neurons, Ex4 increased the frequency of the mIPSCs of approximately half of the TRH neurons. Ex4 did not influence the mEPSCs. Intracellularly administered G-protein inhibitor, GDP-β-S, did not prevent this effect of Ex4 demonstrating that Ex4 facilitates the inhibitory input of TRH neurons by binding to presynaptic GLP-1R.
Thus, GLP-1 influences the TRH neurons both directly and indirectly. Therefore, the net effect of GLP-1 signaling on the TRH neurons of the PVN may depend on the activity of the GLP-1R-containing inhibitory inputs of these cells. When these inhibitory inputs are inactive, GLP-1 should stimulate the TRH neurons. However, when the GLP-1R-containing inhibitory inputs are active, the GLP-1-induced facilitation of inhibition could diminish the GLP-1-induced direct stimulation of TRH neurons. The PVN receives GABAergic inputs from several brain areas where GLP-1R is expressed, such as the hypothalamic dorsomedial nucleus, bed nucleus of stria terminalis, and preoptic area, 8,16 but identifying the sources of GLP-1R-containing GABAergic inputs of TRH neurons requires further studies.
The axons of hypophysiotropic TRH neurons terminate in the blood–brain barrier free external zone of the ME, where circulating hormones can easily access these terminals. We observed that the majority of the TRH axons contained GLP-1R in this brain region, suggesting that circulating GLP-1 may directly regulate the activity of hypophysiotropic TRH axons. Indeed, ME explant experiment showed that activation of GLP-1Rs had a delayed inhibitory effect on the TRH release of these axons. Similar delayed inhibitory effect of GLP-1R activation was described earlier on the nucleus accumbens-projecting neurons of the paraventricular nucleus of the thalamus. 17 These data suggest that different second messengers may mediate the effects of GLP-1R activation in the perikaryal and axonal compartments of TRH neurons.
Based on the potent stimulatory effect of GLP-1R activation on the activity of the TRH neurons in the PVN, we expected that administration of the long-acting GLP-1R agonist liraglutide increases circulating thyroid hormone levels. Despite the marked inhibitory effect of liraglutide on the food intake and energy expenditure during the 24 hours of the experiment, the treatment did not influence the TRH gene expression in the PVN. This is surprising as both the direct stimulatory effect of GLP-1R signaling and decreased NPY synthesis in the ARC should increase the activity of TRH neurons. Thus, it is not clear why the TRH neurons did not respond to liraglutide treatment. One possibility is that peripherally administered liraglutide cannot access the hypophysiotropic TRH neurons.
This hypothesis is supported by the very limited distribution of the neurons labeled with peripherally administered fluorescent liraglutide. 18 In addition, it is also conceivable that the direct stimulatory effect of liraglutide on TRH neurons is counterbalanced by its effect on the inhibitory inputs of the TRH neurons.
Heppner et al. published a very similar observation about the interaction of GLP-1R signaling and the kisspeptin neurons of the ARC. 19 They reported that peripheral liraglutide treatment did not stimulate kisspeptin neurons despite of the direct stimulatory effect of GLP-1R signaling on these cells observed with patch-clamp electrophysiology. 19
To assess the effect of liraglutide treatment on the regulation of TSH levels, the TSHβ mRNA level of the pituitary gland was determined as an alternative of the serum TSH level determination as suggested by the American Thyroid Association Guide, 20 because of the problem of most mouse TSH assays. We have observed a decreasing tendency of TSHβ mRNA expression in the pituitary.
Accordingly, the fT4 level is decreased in the blood 24 hours after the liraglutide treatment. As the axon terminals of hypophysiotropic TRH neurons are located outside the blood–brain barrier, 6 the liraglutide-induced decrease of fT4 levels might be due to the effect of liraglutide on the activity of the hypophysiotropic TRH terminals. However, we cannot exclude the possibility that liraglutide could affect peripheral T4 metabolism.
The proopiomelanocortin (POMC) neurons of the ARC are considered to be important mediators of the effects of liraglutide on energy homeostasis. 18 The unchanged POMC and cocaine- and amphetamine-regulated transcript (CART) expressions in the ARC of mice 24 hours after liraglutide treatment suggest that the liraglutide-induced inhibition of food intake is independent from these anorexigenic neurons in this early phase. In contrast, the markedly decreased synthesis of the orexigenic NPY in the ARC may contribute to the anorexigenic effect of liraglutide in the first 24 hours of treatment. Interestingly, the treatment had no effect on the AGRP expression in these cells. The inhibition of NPY expression is in accordance with the inhibitory effect of GLP-1R agonists on the NPY/AGRP neurons. 18 This effect was thought to be exclusively indirect via the facilitation of the GABAergic inputs of NPY neurons. 18
However, we recently showed that a population of NPY neurons express GLP-1R, 21 indicating that GLP-1R agonists may have direct effects on these neurons.
Similar to the liraglutide treatment, central glucose administration decreases NPY gene expression in the ARC without influencing AGRP, POMC, or CART expression 22 raising the possibility that GLP-1R agonists and glucose may act via a common signaling mechanism in these feeding-related neurons.
In summary, our data demonstrate that GLP-1R activation has direct stimulatory effect on TRH neurons in the PVN, but the activation of GLP-1R may also inhibit TRH neurons by facilitating the inhibitory inputs of these cells or by inhibiting their axon terminals. The innervation of TRH neurons by GLP-1 neurons suggests that TRH neurons might be influenced by both circulating GLP-1 and GLP-1 released from the GLP-1 neurons of the NTS. The lack of regulation of TRH neurons by liraglutide treatment indicates that the HPT axis does not mediate liraglutide-induced weight loss.
Footnotes
Acknowledgments
The authors thank Veronika Penksza for her technical help and Charles Pyke (Novo Nordisk A/S, Måløv, Denmark) for providing the GLP-1 and GLP-1R antibodies and the liraglutide.
Authors' Contributions
Y.R.: investigation, methodology, and writing original draft; Z.P.: investigation, methodology, formal analysis, and writing original draft; A.S.-S.: investigation, methodology, and writing original draft; D.K.: investigation; B.T.: investigation; M.M.: methodology; G.W.: investigation; C.F.: conceptualization, methodology, writing—review and editing, supervision, and funding acquisition.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The study was supported by the National Brain Research Program (NAP 3.0) of the Hungarian Academy of Sciences and by Project no. RRF-2.3.1-21-2022-00011, titled National Laboratory of Translational Neuroscience has been implemented with the support provided by the Recovery and Resilience Facility of the European Union within the framework of Programme Széchenyi Plan Plus.
Supplementary Material
Supplementary Data