
Abstract
Background:
Graves' disease (GD), one of the most common forms of autoimmune thyroid disorders, is characterized by hyperthyroidism caused by antibodies (Abs) against the extracellular A-subunit of the thyrotropin receptor (TSHR). Various approaches have been used to create mouse models of GD, including transfected fibroblasts and immunization with plasmids or adenoviruses expressing human TSHR A-subunit (hTSHR A-subunit). These models, however, require repeated immunization and produce inconsistent results. In this study, we established a novel Cre-loxP system-based mouse model that is able to generate the hTSHR A-subunit, mimicking human GD, and characterized the histological changes in Graves' orbitopathy (GO) progression after a single injection.
Materials and Methods:
A Cre-loxP system-based mouse model was constructed by inserting the CAG-loxP-STOP-loxP-hTSHR A-subunit cassette into the Rosa26 locus of the mouse genome. Conditional expression of the hTSHR A-subunit was successfully achieved by intramuscular injection of the transactivator of transcription-Cre recombinase (GD mice). Blood tests for anti-TSHR Abs and the total thyroxine (T4) level were performed. Magnetic resonance imaging (MRI) was used to monitor morphological changes in the eyes. A histological examination of the thyroid gland and retrobulbar tissues was performed to observe pathological changes.
Results:
Twenty-four (8 control and 16 GD) mice were investigated. All GD mice exhibited higher levels of TSHR Abs compared with the control group. Moreover, more than 80% of the mouse models showed elevated T4 levels accompanied by thyroid goiter. MRI analysis revealed an increased volume of retrobulbar tissue, while immunohistochemical staining of orbital tissues exhibited macrophage infiltration and muscle fibrosis in the GD mice, contrasting with the control group.
Conclusions:
Our novel mouse model for GD, which showed the histological features of GO, was successfully established using the Cre-loxP system. This animal model offers improved insights and contributes to advancing methodological developments for GD and GO.
Introduction
Graves' disease (GD) is one of the most common forms of autoimmune thyroid disorders. It is characterized by hyperthyroidism caused by activation of the thyroid gland by autoantibodies against the thyrotropin receptor (TSHR). 1 Graves' orbitopathy (GO), also known as thyroid eye disease, is an inflammatory autoimmune disorder of the orbit that occurs in up to 50% of patients with GD. 2,3 In 3–5% of patients with GO, severe symptoms and noticeable changes in appearance may lead to impairment in quality of life. Around one-third of the patients who have GO suffer from unsatisfactory treatment outcomes. 4
An animal model exhibiting GD in combination with GO is crucial for understanding their pathological features and developing therapeutic approaches. The TSHR, which plays a significant role in development of both GD and GO, serves as a key factor in the establishment of animal models for these diseases. 5 In GD, the loss of immune tolerance to the TSHR results in production of autoantibodies that target the TSHR on thyroid epithelial cells, leading to excessive secretion of thyroid hormones.
Likewise, the primary cause of GO is the immune response triggered by antibodies (Abs) binding to TSHR expressed in the retrobulbar adipose tissue. 6 Various approaches have been explored to replicate human GD in mice, 7 that is, by transfection of fibroblasts 8 or plasmids 9 and immunization with adenovirus expressing the TSHR. 10 Recognizing that the TSHR A-subunit (significantly more than the TSHR holoreceptor) induces TSHR Abs biased toward thyroid stimulation, 11 the subsequent approaches predominantly favor utilization of the TSHR A-subunit. 12 –14
These methods have been proven to induce serum TSHR Abs and hyperthyroidism. Nonetheless, multiple electroporation pulses are required to establish a disease model using a TSHR plasmid, which can lead to significant mortality. 12 Recently, research has focused on immunization with the adenovirus-hTSHR A-subunit to generate TSHR Abs and induce hyperthyroidism. 15 The animal model generated through this approach also reproduced several GO orbital features, including adipogenesis, fibrosis, and infiltration by F4/80+ macrophages.
However, this method requires time for adenovirus production and involves the use of unnecessary antigens, 12 along with nine repeated injections. 14 It is challenging to use existing animal models of GD for research since they are time-consuming and labor-intensive and yield inconsistent results. 16 A genetically engineered animal model for GD and GO may be a convenient method for shortening the process of developing the model and improving research efficiency. 17
In this study, we established the GD animal model by using a single injection of transactivator of transcription (TAT)-Cre recombinase to induce hTSHR A-subunit expression in Cre-loxP genetically engineered mice. This study presents a report on genetically modified mice with GD and GO, demonstrating a model that mimicked GD with an 81.25% frequency and the progression to GO through changes in orbital tissue, making this an ideal tool for further advancements in research for GD and GO.
Materials and Methods
Construction of the Rosa26_hTSHR A-subunit vector
The complementary deoxyribonucleic acid (cDNA) encompassing the hTSHR A-subunit (amino acid residues 1–289) (NCBI Reference Sequence: NM_000369.5) was obtained by amplifying human cDNA in a normal thyroid tissue sample. The DNA and Rosa26 plasmid were then digested with the restriction enzymes, AscI and XmaI (New England Biolabs, Ipswich, MA, USA).
The Rosa26 plasmid refers to the Rosa26 arm that contains a neomycin resistance gene with two loxP sites in the same direction on both sides downstream of the cytomegalovirus (CMV) early enhancer/chicken β-actin (CAG) promoter (Macrogen, Seoul, South Korea). The digested DNA was ligated to fuse the insert to the recipient plasmid. The constructs were then subcloned and confirmed by DNA sequencing.
Establishment of homologous recombinant embryonic stem cell clones
The construct was then transfected to J1 embryonic stem (ES) cells derived from 129S4/SvJae mice (Macrogen) by electroporation to form recombinant ES cell clones. Positive clones were selected using G418 (#10131035; Thermo Fisher Scientific, MA, USA) at a concentration of 200 μg/mL in an ES cell culture medium.
After transfecting the established recombinant ES cells with a pBS513 EF1alpha-cre plasmid, purchased from Addgene (Cambridge, MA, USA), expression of the TSHR A-subunit was confirmed by reverse transcription–polymerase chain reaction (RT-PCR). Primer sets for the hTSHR A-subunit and Gapdh are shown in Table 1.
Primers Used for the Polymerase Chain Reaction
Primer sets Rosa26_ex_F + CMV_enhancer_R targeting the external region of the Rosa26_hTSHR A-subunit and primer sets, Chimeric_intron_F + Neo_R and OS_Neo_F + bGH_polyA_R, targeting different parts of the internal region of the Rosa26_hTSHR A-subunit.
PCR, polymerase chain reaction; F, forward; R, reverse.
Obtaining hTSHR A-subunit heterozygous mice and backcrossing
Rosa26_hTSHR A-subunit ES cell clones were microinjected into 129S4/SvJae mouse embryos ([i.e., the donor] Macrogen); these embryos were transplanted into recipient C57BL/6 mice (Macrogen) to obtain chimeric mice. The chimeric mice were then crossed with wild-type C57BL/6 mice to obtain the F1 generation. Rosa26_hTSHR A-subunit-positive F1 generation screening was followed by PCR using the primers listed in Table 1.
The F1 individuals were backcrossed to wild-type BALB/c mice (OrientBio, Inc., Seongnam, South Korea). After five successive generations, the mice were used for the experiment.
Mouse genotyping
All mice were genotyped by PCR using genomic DNA isolated from toe cuts. For each toe sample, 100 μL of direct PCR lysis reagent (#102; Viagen Biotech, Irvine, USA) containing 0.1 mg/mL proteinase K (#p2308; Sigma-Aldrich, St. Louis, MO, USA) was added. The lysis procedure was conducted as per the guidelines of the manufacturer. Subsequently, PCR was carried out using the primers listed in Table 1.
Induction of hTSHR A-subunit expression in genetically engineered mice
The mice were housed under standard light (12-hour light–12-hour dark) and temperature (22–24°C) conditions with free access to a standard rodent chow diet and water. The Institutional Animal Care and Use Committees of Yonsei University Health System in Seoul, South Korea, approved all animal experiments (Approval No. 2020-0111).
At 8 weeks of age, 500 U (50 μL) of TAT-Cre recombinase (Sigma-Aldrich) was injected intramuscularly into the leg muscle of each mouse (GD group, n = 16 [male = 6 and female = 10]). At the same time, 50 μL of saline was administered to the control mice using the same method (control group, n = 8 [male = 4 and female = 4]). The body weight was measured, and blood was collected every 4 weeks for TSHR Ab and total thyroxine (T4) measurements.
At 13 weeks after injection, orbital magnetic resonance imaging (MRI) was conducted using a 9.4T BioSpec 94/20 MRI scanner (Bruker BioSpin GmbH, Ettlingen, Germany). The ImageJ program (National Institutes of Health [NIH], Bethesda, MD, USA) allowed us to label retrobulbar tissues with a “free hand line” and calculate the retrobulbar tissue volume.
At 14 weeks after the injection, mice were sacrificed, and blood was collected for thyroid-stimulating antibody (TSAb) and free T4 (fT4) measurements. The thyroid gland and retrobulbar tissues were microsurgically excised for histopathological studies.
Measurement of thyroid Abs and thyroid hormones
Serum TSHR Ab levels were measured using a fast enzyme-linked immunoassay (ELISA) commercial kit (EUROIMMUN Medical Laboratory Diagnostics Ltd., Luebeck, Germany), and total T4 concentrations were measured using an ELISA kit (DRG International, Inc., Springfield, NJ, USA). The level of TSAb was measured using the Thyretain TSI Reporter BioAssay system (Diagnostic Hybrids, Inc., Athens, OH, USA), and the level of fT4 was measured using the FT4 II electrochemiluminescence immunoassay system (Roche, Mannheim, Germany).
Histological examination of thyroid and retrobulbar tissues
Thyroid and orbital tissues were fixed in 10% buffered formalin, dehydrated, and embedded in paraffin and sectioned for hematoxylin and eosin staining, Masson's trichrome staining, immunohistochemical staining for CD3 (1:100, ab11089; Abcam, Cambridge, United Kingdom) to identify T cells and for F4/80 (1:100, NB600-404; Novus, Littleton, USA) to identify macrophage cells, and alcian blue staining to identify hyaluronic acid. The adipose tissue was quantified using ImageJ and normalized to the optic nerve area.
Statistical analyses
All data are presented as mean and standard error of the mean (SEM). Statistical differences between the two groups were analyzed by an unpaired t-test using Prism, v6.01 (GraphPad, San Diego, CA, USA). Significance was set at the following p-values: *p < 0.05, **p < 0.01, and ***p < 0.001.
Results
Generating hTSHR A-subunit genetically engineered mice
The schematic diagram of the Rosa26_hTSHR A-subunit vector is shown in Figure 1A. After hTSHR A-subunit-containing mouse ES cells were established, whether the cells conditionally expressed the hTSHR A-subunit was confirmed using RT-PCR when the stop sequence between loxP sites was cut with Cre (Fig. 1B). The F1 generation of mice was screened by PCR using specific primers for the Rosa26_hTSHR A-subunit shown in Table 1 (Fig. 1C).
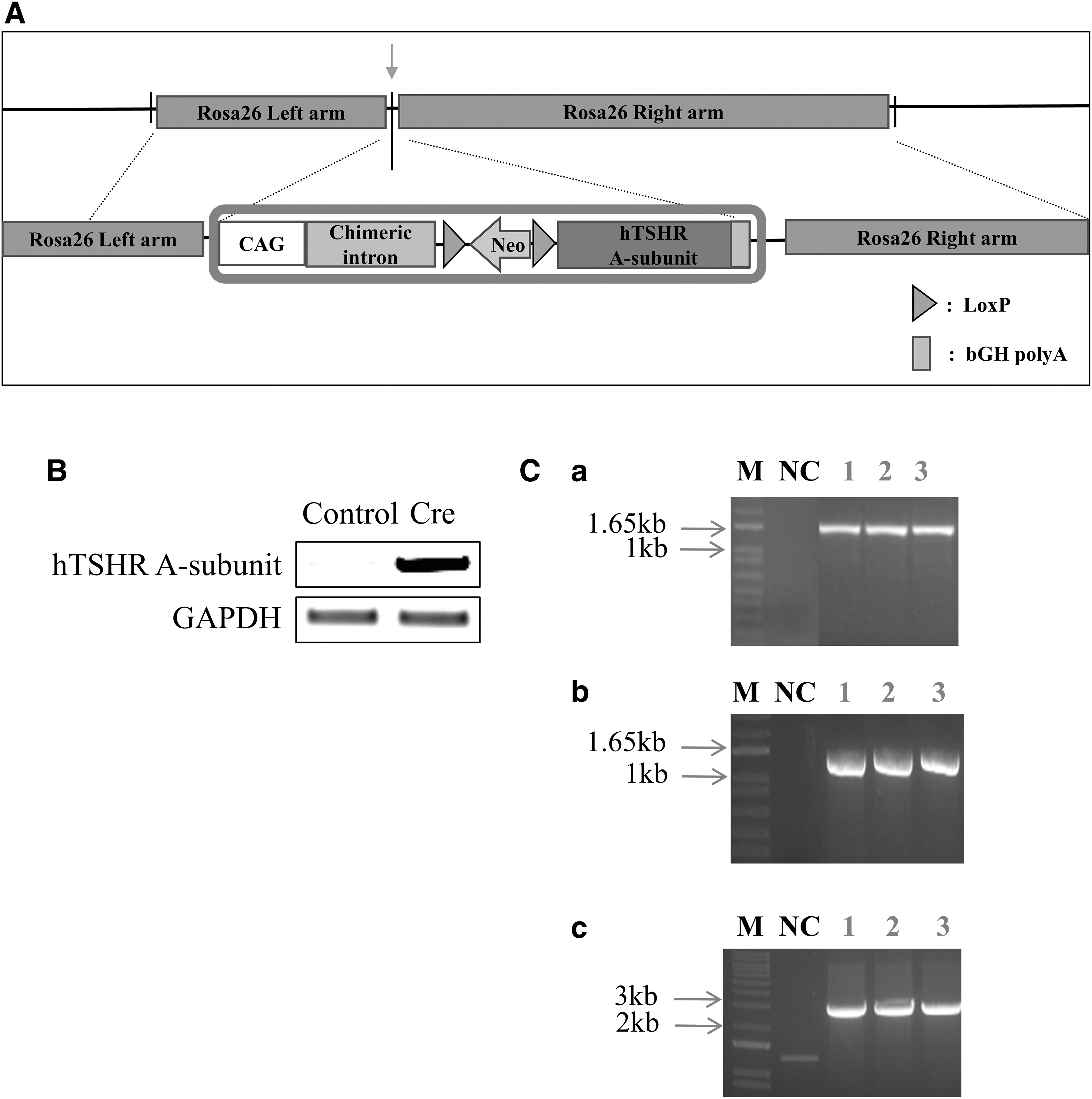
Production of hTSHR A-subunit genetically engineered mice.
These results indicate that we have successfully constructed genetically modified mice that can conditionally express the hTSHR A-subunit.
Evaluation of thyroid function
We induced hTSHR A-subunit expression in genetically engineered mice by injecting them with TAT-Cre recombinase and assessed the progression of GD at predetermined time points (Fig. 2A). After injection, no significant differences were noted in body weight between the GD and control group mice: at week 4, 24.629 ± 0.725 g versus 23.675 ± 1.245 g (p = 0.54); at week 8, 27.194 ± 0.973 g versus 25.337 ± 1.331 g (p = 0.24); and at week 12, 28.869 ± 0.964 g versus 26.9 ± 1.331 g (p = 0.21) (Fig. 2B).
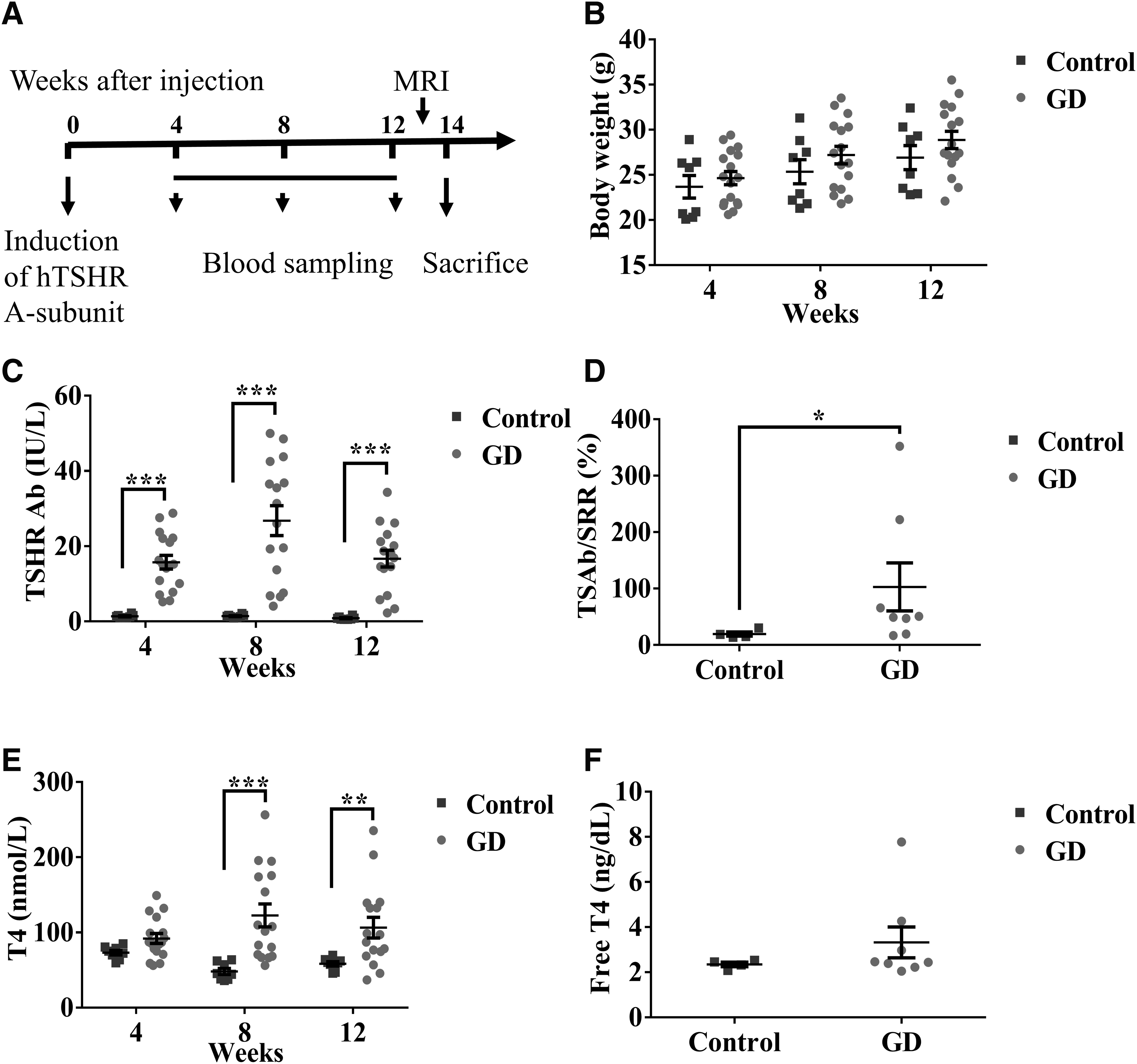
Induction of hTSHR A-subunit expression in hTSHR A-subunit genetically engineered mice.
The presence of hyperthyroidism was determined by measuring serum thyroid Abs and thyroid hormone levels. TSHR Ab levels were higher in the GD group mice than in the control group mice at all three time points: at week 4, 15.748 ± 1.815 IU/L versus 1.374 ± 0.146 IU/L (p < 0.001); at week 8, 26.796 ± 3.985 IU/L versus 1.401 ± 0.131 IU/L (p < 0.0001); and at week 12, 16.695 ± 2.234 IU/L versus 0.879 ± 0.130 IU/L (p < 0.001) (Fig. 2C).
TSAb levels at week 14 were significantly elevated in the GD group mice (Fig. 2D). Most mice in the GD group showed higher thyroid hormone levels than those in the control group. The total T4 levels in the GD and control groups were as follows: at week 4, 91.962 ± 6.545 nmol/L versus 73.139 ± 3.035 nmol/L (p = 0.291); at week 8, 122.572 ± 15.182 nmol/L versus 48.092 ± 3.989 nmol/L (p < 0.0001); and at week 12, 106.461 ± 13.716 nmol/L versus 58.278 ± 2.847 nmol/L (p = 0.009) (Fig. 2E). The fT4 levels at week 14 showed the same trend as the total T4 level (Fig. 2F).
These above results demonstrate successful establishment of an animal model resembling hyperthyroidism in GD, characterized by notable pathological features, including elevated T4 levels and significant upregulation in expression of TSHR Abs.
Histological study of the thyroid gland
Diffuse enlargement and hypertrophy of the thyroid gland were noted in GD group mice, in contrast to the gland in the control group mice (Fig. 3A). We found a significant difference in the area of thyroid tissue between the groups: GD group 34.125 ± 2 mm2 versus control group 13.125 ± 1.856 mm2 (p < 0.001) (Fig. 3B). Morphological examination of the thyroid gland showed tall and columnar morphology in the GD mice, suggesting hyperthyroidism (Fig. 3C).
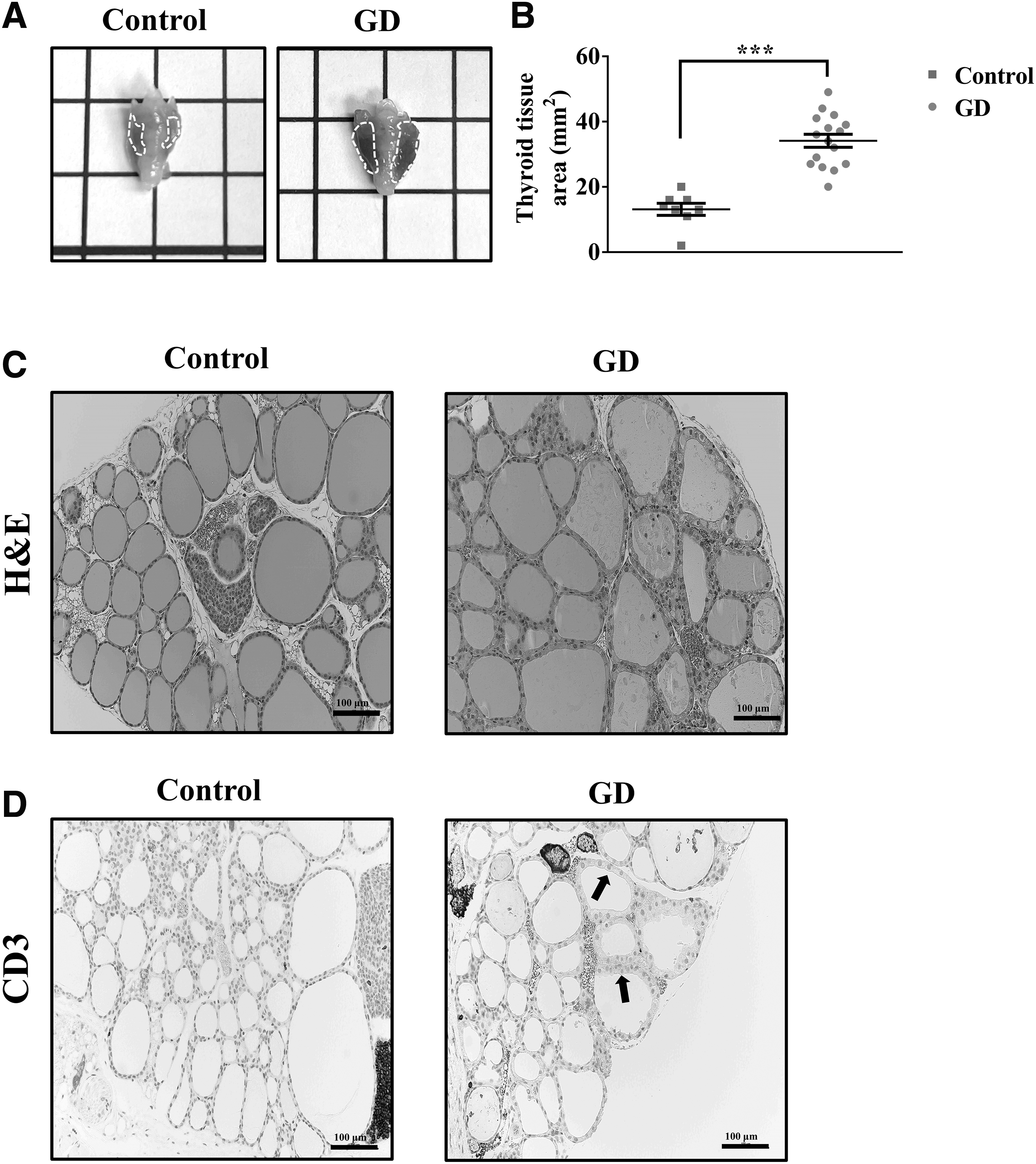
Histological study of the thyroid gland from GD animal models.
In addition, staining for CD3 in the GD group mice revealed extensive T cell infiltration (Fig. 3D). These histological changes in the thyroid gland show that the animal model demonstrates features of hyperthyroid glands.
Histological study of retrobulbar tissues
Before the animals were sacrificed, the orbit was subjected to MRI. There was no significant protrusion of the eyeball in the coronal images (Fig. 4A). However, in the GD group mice, a significant increase in the volume of retrobulbar tissues was observed. We found significant differences in volume of the two sides of the retrobulbar adipose tissue between the GD group mice and control group mice: left eye: 0.618 ± 0.042 mm3 versus 0.485 ± 0.029 mm3 (p < 0.05); and right eye: 0.642 ± 0.039 mm3 versus 0390.426 ± 0.02 mm3 (p < 0.001) (Fig. 4B).

MRI study of the GD animal models.
We also found statistical differences in volume of the two sides of the retrobulbar muscle between the GD and control groups: left eye: 4.622 ± 0.262 mm3 versus 3.509 ± 0.329 mm3 (p < 0.05); and right eye: 5.213 ± 0.217 mm3 versus 4.114 ± 0.207 mm3 (p < 0.05) (Fig. 4B).
The histological examination showed that the volume of the retrobulbar adipose tissue was increased in GD group mice (Fig. 5A). Additionally, as shown in Figure 5B, F4/80+ macrophages in the retrobulbar tissue were seen more frequently in the GD group mice, indicating recruitment of macrophages to the site of inflammation, than in the control group mice. Masson's trichrome staining of the retrobulbar tissue of the GD group mice revealed fibrosis in the muscle tissue (Fig. 5C). In summary, the GD mouse model also exhibits histological features of GO.

Histological analysis of the retrobulbar tissue of animals with GD.
Discussion
In this study, we created a genetically engineered animal model that could exhibit GD pathogenesis and accompanying GO histological manifestations with only one injection of TAT-Cre recombinase. Numerous attempts have been made to establish GD animal models using the TSHR A-subunit, which acts as an autoantigen. 18,19
In previous studies, immunizing mice with a mouse TSHR A-subunit-encoding adenovirus hardly elicited TSHR A-subunit Abs due to established tolerance against the mouse TSHR. 20 Recent studies have demonstrated that genetic immunization of human TSHR A-subunit using adenovirus vectors could generate a GO-like phenotype. However, this widely used animal model requires repeated immunization to generate phenotypes resembling GO. 14
The prolonged time and repetitive procedures required for these models have prompted researchers to make efforts to construct models in more efficient ways and to improve the feasibility of the research and reduce the burden on animals. Rather than administering the exogenous hTSHR A-subunit gene to mice, we investigated the possibility of genetically modified mice conditionally expressing the hTSHR A-subunit.
To achieve expression of the hTSHR A-subunit in mice at specific time points, specific gene-editing methods need to be considered. One widely employed approach is the tamoxifen-inducible CreER system. 21 In the absence of tamoxifen, Cre is commonly considered to be excluded from the nucleus. 22 However, there is a “leaky” phenomenon where Cre nuclear translocation can be induced even without tamoxifen. 23,24
In our preliminary study using the tamoxifen-inducible CreER system, leaky expression may have led to immune tolerance and failure to produce TSHR Abs. In light of the idea that high-dose adenovirus expressing the hTSHR A-subunit partially overcame resistance to the hTSHR A-subunit in mice 25 and, conversely, low-dose immunization with adenovirus expressing the hTSHR A-subunit induced high prevalence of hyperthyroidism, 13 we attempted varying doses of tamoxifen, from low to high.
Unfortunately, we observed no increased TSHR Ab production (data not shown). To prevent immune tolerance caused by a leaky expression, we used the Cre-loxP system to establish mouse models capable of expressing the hTSHR A-subunit. 26 TAT-Cre recombinase, a cell-permeable Cre that can activate the Cre-loxP system, was utilized to achieve conditional expression of the hTSHR A-subunit gene. 27
However, hTSHR A-subunit expression alone does not guarantee establishment of a GD animal model given the potential for antibody neutralization. 28 Therefore, we tested various concentrations of TAT-Cre recombinase to find the optimal dosage that could effectively induce hyperthyroidism (data not shown).
As the lineage of mice is crucial in construction of animal models, both C57BL/6 and BALB/c mice have been used to induce GD and GO models, but the success rates have differed. 29 Additionally, studies have shown that BALB/c mice carry dominant genes associated with susceptibility to induction of hyperthyroidism, distinguishing them from C57BL/6 mice. 30
However, considering that C57BL/6 mice are preferred for site-directed modifications and transgenesis experiments due to their superior reproductive performance, we conducted preliminary trials on C57BL/6 mice for our model. Consequently, TSHR Ab levels were elevated in the GD group compared with those in the control group, but the total T4 levels did not consistently increase (data not shown). Considering these factors, the mice were backcrossed to wild-type BALB/c mice, and the resulting generation was used for developing the GD model.
The main characteristics of GD models are elevated serum T4, TSHR Ab, and TSAb levels and typical histological alterations such as thyroid follicular cell hypertrophy. 31 –33 Our results demonstrated that serum T4, TSHR Ab, and TSAb levels increased in the GD animal group. However, owing to the substantial volume of serum required to measure TSAb levels, we excluded data that could not be measured because of insufficient sample volumes.
In addition to the aforementioned characteristics of GD, the mouse models exhibit several typical pathological features of GO, including swelling of the extraocular muscles, an increase in retrobulbar adipose tissue, 34 and infiltration of F4/80+ macrophages in the retrobulbar tissue. 35 In summary, the model used in this study successfully reproduced the majority of GD and GO characteristics within three months.
It is important to acknowledge that all animal models have limitations. We can gain comprehensive insights by examining the same disease across multiple models than by investigating it with a single model. 36 –38 One limitation of this study is that alcian blue staining, which indicates accumulation of hyaluronic acid in the retrobulbar tissue, showed no differences in the staining pattern between the GD and control group mice (data not shown).
In a recent study on animal models of GO induced by adenovirus, orbital pathological changes were observed after 34 weeks. 39 Therefore, more obvious orbital symptoms can be expected if the experimental period is extended. In addition, another limitation of this study is that it did not achieve equal distribution of the sexes in the experimental group. However, we believe this did not impact our results because one study showed that the occurrence rates and intensities of autoimmune hyperthyroidism and orbitopathy were similar in both males and females. 40
Taken together, in this study, we successfully established a mouse model of GD and GO by recombining the hTSHR A-subunit at the Rosa26 locus of the mouse genome and by injecting TAT-Cre recombinase to enable hTSHR A-subunit expression, thereby resembling the clinical features of GD and exhibiting the histological manifestations of GO (Fig. 6). This animal model offers improved insights and contributes to advancing methodological developments for GD and GO.
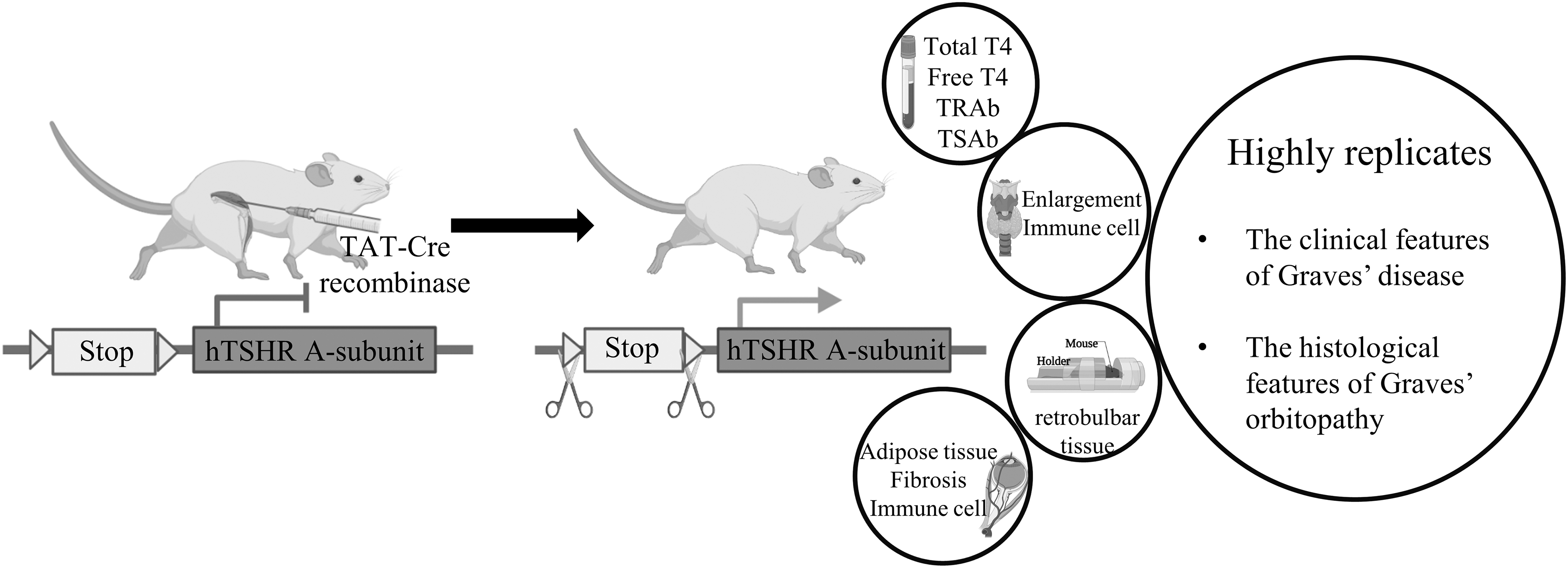
Key insights and conclusive findings from the animal model. A mouse model of GD was developed by injecting TAT-Cre recombinase to enable hTSHR A-subunit expression, resembling the clinical features of GD and exhibiting the histological manifestations of GO. GO, Graves' orbitopathy; TAT, transactivator of transcription.
Footnotes
Acknowledgments
The authors would like to thank Editage for English language editing. They would also like to acknowledge the Annual Meeting of the Endocrine Society—ENDO 2023, where aspects of this research were presented as a poster.
Authors' Contributions
Y.B. was involved in data curation (lead); writing—original draft preparation (lead); visualization (lead); investigation (lead); and formal analysis (lead). D.K. was involved in conceptualization (lead); methodology (lead); funding acquisition (lead); supervision (lead); project administration (lead); and writing—reviewing and editing (lead). Y.H.C., C.R.K., and J.S.Y. were involved in investigation (supporting). E.J.L. was involved in conceptualization (equal); methodology (equal); supervision (supporting); and project administration (supporting).
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by a faculty research grant from Yonsei University College of Medicine [Grant No. 6-2021-0128] (D.K.) and the Korean Endocrine Society of Hyangseol Young Investigator Award 2021 (D.K.). Y.B., Y.H.C., C.R.K., J.S.Y., and E.J.L. declare that they have no funding information.